![]()
![]()
![]()
Rates of growth and milk production by ruminants grazing tropical pastures or consuming crop residues alone are generally low and are only about 10% of the animal's genetic potential. The reasons for this low productivity are complex, but in order of priority they appear to be:-
the imbalanced nature of the nutrients that arise from digestion of such forages when these are fed without supplements
the high incidence of disease and parasitism, which is exacerbated by poor nutrition
the hot, often humid environment which lowers feed intake of ruminants, particularly by cattle.
Feed concentrates such as grain are often recommended as supplements under the above conditions as the animals are usually thin and believed to be energy deficient. However, the poor condition of cattle in the tropics is more likely to be the result of inefficient digestion in the rumen and inefficient utilization of the nutrients absorbed. In hot environments the heat generated by such inefficiencies is hypothesized to lower overall feed intake, which is then expressed as a frank energy deficiency (Leng, 1990).
Research has shown that large improvements in ruminant productivity and efficiency of feed utilization from low quality forages can be achieved by small amounts of supplements that provide essential nutrients in the basal feed (Preston & Leng, 1986; Leng, 1990). These increases are not restricted to laboratory studies and have been observed on a large scale in terms of milk production in cattle and buffalo on small farms in India (NDDB, 1989). These principles of supplementation are also being used to develop beef production in cropping areas of China, where over six million cattle are being fattened by small farmers using straw treated with ammonia to improve both its digestibility and non-protein nitrogen (NPN) content; the straw is then supplemented with bypass protein (Guo Tingshuang & Yang Zhenhai, 1994; Dolberg & Finlayson, 1995; Sansoucy, 1995). These projects emphasize the large potential impact of the feeding strategies that are discussed in the coming sections
Increased production of meat and milk are the obvious benefits from strategic supplementation of large ruminants fed poor quality forages. The improved body condition of animals, which is a major response to balancing nutrition in dairy, beef or draft animals, also affects work performance and reproductive efficiency; the age at puberty of heifers is lowered, the intercalving interval in mature cows is decreased and the work capacity of animals is improved.
It is possible, through strategic supplementation, to increase efficiency of production by up to five fold over similar animals fed only poor quality forages. This has been done without changing the basal feed resources and has been achieved by identifying and providing critical nutrients that are otherwise deficient in the diet. In this way nutrient availability is balanced with animal requirements and forage intake is stimulated at the same time (see Leng, 1990; Preston & Leng, 1986).
In general, the supplements to such diets for ruminants include a source of fermentable N, minerals for the rumen organisms and a source of protein that is protected from degradation in the rumen but moves rapidly to the lower tract to improve the essential amino acid supply to the animal.
In many tropical countries the appropriate supplements are readily available, e.g.,
urea and molasses as N and trace mineral sources respectively
local minerals
oilseed meals and protein byproducts of other industries.
There is, however, a need to identify the most appropriate sources of non-protein nitrogen (NPN), minerals and protein available locally and provide them in a form to complement the nutrients absorbed from fermentative digestion of a poor quality forage diet.
Within the developing countries the basic resources for ruminant production are pasture, crop residues and other cellulosic biomass; these are generally low in total protein and fermentable N and are quite often deficient in a range of minerals. Soils in the tropical and subtropical climate zones are almost always associated with low content of such minerals as P, S, K, Ca, Mg, Cu, Mo, Co, Se and B (Kerridge et al. 1986) and high aluminium saturation or acidity which are restrictive to root growth (Chen et al. 1992). In tropical soils, phosphorus is often a major mineral deficiency.
Often a source of bypass protein is not available locally, particularly in the grasslands or savannahs. In the rangelands, leguminous forages, tree foliages, their seeds and pods are by far the greatest potential source of protein meals, soluble N and source of minerals. When fed with pasture or straw, they provide many of the nutrients deficient in the basal forage and create a rumen environment more efficient in microbial growth, which is also often accompanied by a higher rate of digestion of the basal forages.
Although the quantitative production of crop and livestock products in third world countries has increased in the last 25 years, this has been achieved largely by way of increased animal numbers (Jackson, 1981).
In some developing countries, high levels of grain are being fed to ruminants, emulating the feedlot strategies of North America. Increasing pressure on cropping lands and massive demand for grain for monogastric animal production, particularly in China, suggest that this is a transient feature, which is likely to cease as population increases and surplus (subsidized) world grain is reduced.
In the future, ruminants in tropical countries and cattle in particular, are likely to depend almost entirely on crop residues, particularly straws and stovers, pasture biomass and agro-industrial byproducts such as sugar cane tops and molasses.
The fibrous feeds contain large amounts of complex carbohydrates which are not digested by intestinal enzymes and therefore require fermentative digestion by microbes. The common characteristics of such feeds are low digestibility, low protein content and a low mineral content.
It is the contention of the writer that there has been a major misconception throughout the scientific literature: the low productivity of ruminants fed poor quality forages in tropical developing countries is mainly a result of the low energy density of the feed. There is now a large volume of evidence that the primary constraint to productivity is rather an inefficient utilization of the feed because of deficiencies of critical nutrients in the diet. The deficient nutrients may be those critical to the growth of rumen microbes which ferment the feed; they may be those required to balance the protein to energy ratio absorbed. Supplementation of animals to supply these nutrients often results in levels of production that would be highly acceptable under temperate country practices. The evidence for these statements has been considered in detail recently by Leng (1990) who suggested that a marked interaction of heat generation in consumption, digestion, metabolism of ruminants and environmental heat load has major implications for ruminant production in the tropics.
The laws of thermodynamics suggest that inefficient utilization of feed by ruminants for tissue deposition must result in increased heat generation. Such heat generation could have large implications for ruminant production in the humid tropics. The often low intakes of poor quality forages by cattle in hot climates may be imposed by accumulation of heat in the body from both an inefficient rumen microbial ecosystem (Table 2.1) and an inefficient utilization of acetate in metabolism of the animal. This would be exacerbated by a high environmental heat load (Leng, 1990). Correction of a nutrient imbalance by supplying nutrients deficient for microbial growth and feeding a bypass protein often increases a depressed intake of poor quality forages (i.e., 50–60 g/kg0.75/d) to normal, i.e., between 80–100 g/kg0.75/d (see Figure 2.1 on the following page).
| YATP | ||||
|---|---|---|---|---|
| 8 | 14 | 19 | 25 | |
| Microbial protein synthesized (g/d)* | 500 | 800 | 1010 | 1212 |
| VFA produced (MJ/d) | 41 | 34 | 30 | 26 |
| Methane produced (MJ/d) | 9.4 | 8.5 | 8.0 | 7.6 |
| Heat (MJ/d) | 6.4 | 5.1 | 4.3 | 3.1 |
| P/E ratio (g protein/MJ) | 12 | 25 | 34 | 47 |
* Microbial protein may be only 75–85% digestible.
Cattle in the tropics, as compared to those in cool countries, may require less nutrients for maintenance as they do not have to combat cold stress. If the nutrients spared oxidation are used for tissue synthesis, animals in warm climates can be more efficient users of forages than animals in cold climates. Ruminants use acetate or fat as energy sources when cold and such animals have a lower requirement for amino acids relative to energy (P/E ratio) than animals under hot conditions. Thus the requirements for amino acids are higher relative to energy substrate in cattle in the tropics than in animals on the same feed in cool environments: the total amount of nutrients required is lower in animals in the tropics relative to temperate areas.
Similar scenarios are seen in the poultry industry where the protein requirements as a percentage of the diet are increased from 16 to as high as 21% and the feed intake is decreased as temperature/humidity increases (Mastika & Cumming, 1994).
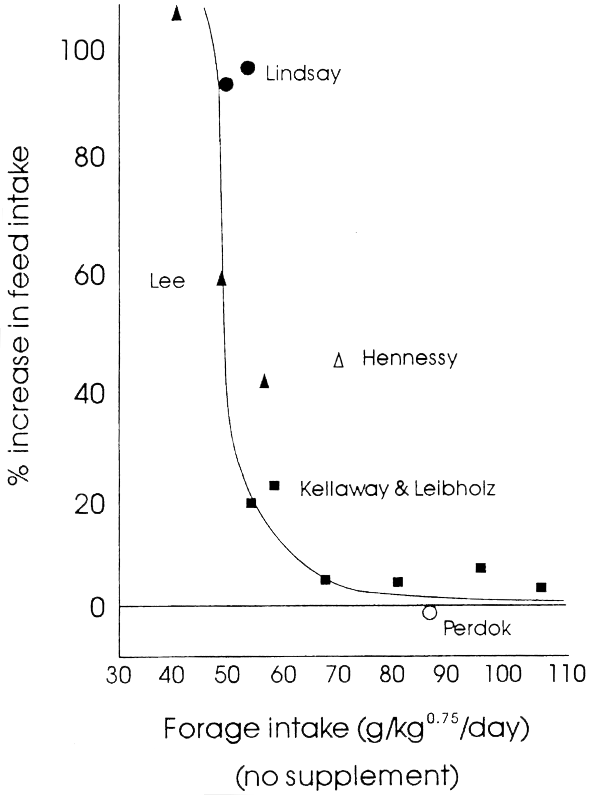
The rationale on which the following discussions centred has been reviewed by Leng et al. (1977), Preston & Leng, (1986) and Leng (1989, 1990). The basic concepts are as follows. Ruminants fed low quality forages require supplementation with critically deficient nutrients to optimize productivity. The required supplements in order of priority must:—
correct nutrient deficiencies for the rumen microbes, particularly ammonia and sulphur and phosphorus
increase the ratio of protein (absorbed amino acid) to energy (VFA) available from digestion so that it more closely corresponds to the animal's requirements.
Ruminant animals obtain their nutrient requirements mainly from the products of rumen fermentation (i.e., microbial cells and VFA) and, in some situations, dietary bypass nutrients. They receive the majority of their essential amino acids from microbial protein on forage based diets, particularly when they are low in true protein. It is, therefore, important to consider how microbial growth efficiency, and therefore amino acid availability from this source, can be maximized so as to minimize the need for expensive bypass protein supplements.
The composition of rumen microbial cells is relatively constant (Hespell & Bryant, 1979; Czerkawski, 1985). A typical analysis is:—
true protein 32–42%
small nitrogenous molecules 10%,
nucleic acids 8%,
lipid 11–15%
polysaccharide 17% and
ash 13%.
Higher lipid contents have been noted in bacterial dry matter isolated from the particle-associated phase of rumen contents (Merry & McAllan, 1983) and from rumens where the microbial modifier rumensin has been fed to animals.
Growth of microbial cells in the rumen requires:—
a source of fermentable carbohydrate to supply:—
monomers (building blocks) for synthesis of cellular material
ATP to provide the energy for polymer synthesis. ATP is generated during carbohydrate fermentation by substrate level phosphorylation and also by other processes associated with electron transfer and transmembrane vectorial metabolism
N compounds for synthesis of proteins and nucleic acids, including:—
ammonia
perhaps some extremely small amounts of some amino acids
an array of minerals and vitamins.
Ammonia can be the sole source of N for the synthesis of protein and other nitrogenous compounds in the majority of rumen bacteria (Bryant, 1973). Forage fed to ruminants may contain high levels of soluble protein, however and 30–80% of the microbial N of mixed bacteria is apparently derived from dietary peptides and amino acids under these circumstances (Nolan et al. 1986; Nolan, 1975). For starch-or sugar-based diets, peptides or amino acids in small quantities appear to be essential N sources for efficient growth of rumen microbes (Broderick et al. 1988; Maeng et al. 1986). The organisms fermenting fibrous carbohydrate in the rumen, however, appear to require little, if any, peptides or amino acids and can grow efficiently on ammonia alone (Maeng et al. 1989; Kanjanapruthipong & Leng, 1996). The cellulolytic species which utilize ammonia for amino acid synthesis may, however, require some branched chain VFA.
It seems reasonable to suggest that, since a large number of soil microbes can establish in the rumen, an efficient microbial ecosystem will develop independent of an amino acid supply if the microbes are fed diets without amino acids but with ample non-protein nitrogen.
The yield to the intestines of microbial cells per mole of dietary OM digested in the rumen and therefore the P/E ratio, is always considerably below that theoretically achievable. A major reason is that a variable part of the ATP generated in the production of volatile fatty acid (VFA) is not available for synthesis and is used to maintain cellular homoeostasis (i.e., maintenance of intracellular ion concentrations, enzyme and other protein molecule turnover), mobility and feeding activity in the case of protozoa. This proportion is termed ‘maintenance ATP’ (Leng, 1981; Czerkawski, 1985). Microbes are able, at times, to partially or totally dissociate catabolism from anabolism. Energy-spilling reactions within the cell (uncoupling) may also be responsible for dissipating ATP energy as heat without cell division and growth. This will occur when there is a deficiency of minerals, ammonia or other nutrients essential for microbial growth and also when microbial cells lyse in the rumen.
In the mixed microbial system in the rumen, bacteria are engulfed and digested by protozoa and lysed by bacteriophages. Materials excreted by living cells and the debris from lysed bacterial, protozoal and fungal cells may be fermented by other microbes, inevitably yielding less ATP than that used in their initial synthesis. These extracellular processes and those associated with maintenance and uncoupling dissipate the potential energy in ATP to heat without achieving synthesis. This is termed the heat of fermentation. In theory this can vary from 4–10% of the gross energy of the carbohydrate fermented depending on the microbial growth efficiency (Table 2.1 on page 12) and therefore the availability of microbial growth requirements.
It needs stressing here that the higher levels of heat production in fermentation may be capable of creating a heat stress in animals existing in climates with high humidity and temperature.
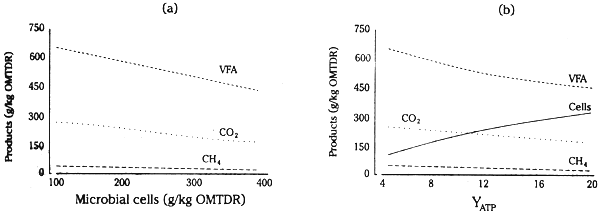
In practice, on some poor quality forages, deficiencies of essential nutrients can reduce microbial growth to very low levels. Under these circumstances, the efficiency of ATP use for microbial cell synthesis (YATP) must be below 2–4 and considerable heat and methane are generated. Supplementation to correct these deficiencies often increases YATP to 14 or higher. This alters considerably the balance of nutrients available. As microbial cell yields per unit of digested substrate (organic matter truly digested in the rumen, OMTDR) is increased (Figure 2.2), or YATP is increased (Figure 2.3 on page 19), a greater fraction of the digested OM is used to provide building block material for the synthesis of cell polymers and proportionally less is channelled into VFA, CH4 and heat production (see Table 2.1 on page 12). The P/E ratio in the products available to the host animal is increased as the efficiency of microbial synthesis or YATP increases (Table 2.2 on the next page) by providing deficient essential nutrients to increase microbial growth and provide increased amounts of bypass protein from diets of low quality forages (Figure 2.2).
Changes in the proportions of VFA produced have a relatively minor effect on ATP generation by rumen micro-organisms and therefore on the quantity of cells synthesized per mole OM digested. Microbial mixes in the rumen producing higher proportions of butyrate in the VFA, however, appear to be associated with lower net yields of cells. Higher rates of production and absorption of propionate benefit the animal tissues by potentially generating more ATP per mole VFA oxidized in the body, and by supplying a higher level of glucose availability. Glucose availability could, at times, limit growth (Preston & Leng, 1986) or milk production (Linzell, 1967) in ruminants.
| Rumen environment | Protein bypass (g/d) | Microbial cells produced (gDM/d) | Protein from microbial cells (g/d) | VFA produced (MJ/d) | Total protein available (g/d) | P/E* (g/MJ) |
|---|---|---|---|---|---|---|
| Deficient | 0 | 830 | 500 | 41 | 500 | 12 |
| Supplements | 0 | 1680 | 1010 | 30 | 1010 | 33 |
| Deficient** | 400 | 830 | 500 | 41 | 900 | 22 |
| Supplements | 400 | 1680 | 1010 | 30 | 1410 | 47 |
* Ratio of microbial protein plus dietary protein to VFA energy
The diets given to ruminants in developing countries seldom approach the high nutrient densities of those used in the temperate countries and the benefits of supplementation are more obvious the poorer the quality of the diet. Responses in cattle grazing dry pastures to molasses urea multinutrient blocks in the tropics are greater than those in animals on similar diets in the temperate areas. These observed different effects of supplementation are explained when climate/nutrition interactions are considered (see page 11).
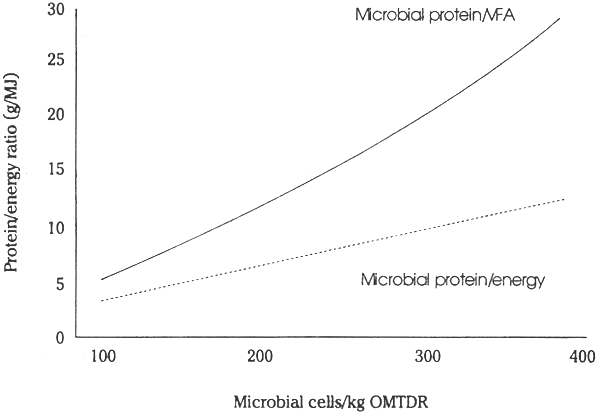
Forages contain mineral components which reflect soil conditions and may therefore be low in essential nutrients such as Na, Mg, P, Ca and S as well as trace minerals such as Co, Cu, Se and Zn. The nett effect of such deficiencies in the rumen is similar to a deficiency of N or ammonia, that is, a lowered microbial cell yield per unit of carbohydrate digested (Figure 2.2 on page 17 and see Leng, 1981) and at times a lowered digestibility. The first effect of any mineral deficiency will be on microbial growth; thus the symptom that first arises is one of malnutrition or protein deficiency.
It is pertinent to stress here that microbial growth efficiency in the rumen may decrease without a concomitant effect on digestibility or feed intake until the washout of the rumen of microbes causes a decrease in microbial pool size (see Kanjanapruthipong & Leng, 1996).
The levels of sulphur and ammonia in rumen fluid which maximize digestibility of fibrous carbohydrates appear to be 1–2 mg S/l and 50–80 mg ammonia N/l respectively (Bray & Till, 1975; Satter & Slyter, 1974), whereas maximum microbial growth efficiency seems to require 4–10 mg S/l and 150–200 mg ammonia N/l respectively (Kandylis, 1984; Moir, 1975; Perdok & Leng, 1990; Boniface et al. 1986; Leng et al. 1993). Growth of fungi is highly dependent on sulphur availability (Akin et al. 1983) and fungi are absent from the rumen on low sulphur diets; this could be a major constraint to digestion of poor quality forages by ruminants in the nonindustrialized tropics. Sulphur deficiency appears to be widespread in the tropics as this mineral is leached from soils readily and quickly. A partial or total loss of fungi from the rumen could reduce digestibility of a mature forage by up to 5 digestibility units, which in turn would lower feed intake by 20–40%.
It is probable that similar relationships to those applying to sulphur and ammonia also apply to the relative availability of other critical minerals, particularly phosphorus, which also affect digestibility and microbial growth efficiency.
A marginal deficiency in the diet of a number of essential microbial nutrients may be undetectable and have no effect on forage digestibility but may significantly lower the protein/energy ratio without effecting metabolizable energy availability (see Leng, 1981). These marginal deficiencies, through their effects on the balance of nutrients from the rumen also affect the efficiency with which the absorbed nutrients are used by the animal.
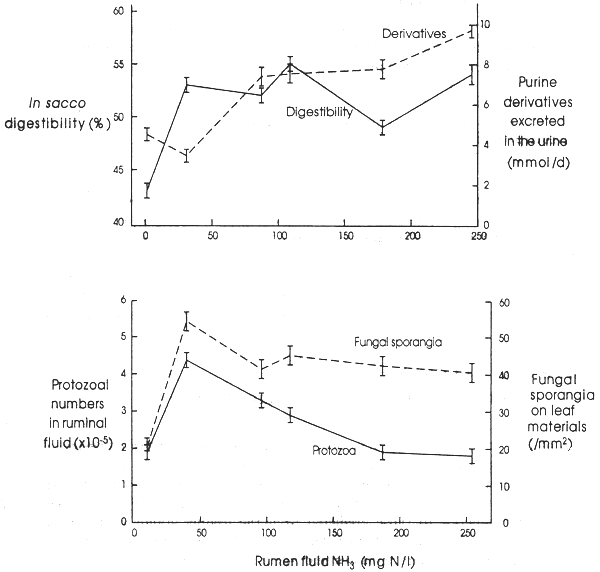
The effects of ammonia availability on digestibility and straw intake of cattle (Figure 2.4 on the following page) suggest that there are two critical ammonia levels. These are 80– 100 mg N/l for optimum fibrolytic activity and 150–250 mg N/l for optimal microbial growth and therefore optimum P/E ratio in the nutrients absorbed.
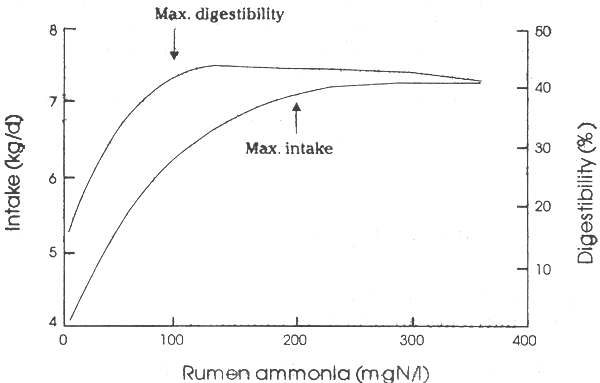
Studies at the University of New England, Australia (Kanjanapruthipong & Leng, 1996) have recorded that on a low protein forage (oaten chaff) the rumen environment of sheep is markedly changed with increasing levels of ammonia. The availability of microbial protein was assessed by using purine excretion as an index of microbial biomass reaching the intestines to be digested and absorbed (Chen & Gomes, 1992). Initially, increases in rumen fluid ammonia of between 10 and 50mg N/l optimized fibre digestibility and apparently stimulated the rumen microbial populations of protozoa and fungi. Thereafter increasing the rumen ammonia level further led to linear decreases in protozoal density in rumen fluid (Figure 2.5 on page 23)
Excretion of purine derivatives in the urine of the sheep decreased initially as ammonia levels increased (which is an indication of the extent of movement of microbes to the lower gut for digestion and absorption of the amino acids (Smith & McAllan, 1971; Chen & Gomes, 1992) and then increased thereafter. Thus, net microbial protein reaching the small intestine with increasing ammonia in the rumen was initially depressed and then increased almost linearly with ammonia concentration. These results indicate a complex series of changes within the rumen ecosystem as ammonia is increased in rumen fluid. It seems that much more ammonia is needed to maximize P/E in the nutrients absorbed than has been previously recommended. This indicates the importance of feeding strategies that maintain ammonia levels in the rumen at high levels at all times, such as occurs when ammoniated straws are fed to cattle or tree foliages are included in low N-forages fed to ruminants. The results also point to a reason why urea supplementation does not always promote increased levels of production from low quality forages. It seems that at the low ammonia levels, protozoa are highly efficient and competitive, reducing bacterial cell flow to the lower gut. Therefore a small amount of extra N in such diets can be detrimental unless it provides an ammonia level that exceeds 40 mgN/l. Even though feed intake and digestibility are increased under such conditions, P/E ratios may be decreased and the benefits of more intake are balanced by a decreased efficiency of feed utilization.
There are a number of dietary manipulations which will alter the P/E ratio in the nutrients absorbed. These include manipulation of:—
the rumen. As has been discussed above, a suboptimal level of any nutrient critical for microbial growth will result in low protein to energy (P/E) ratio in the nutrients absorbed. The first nutritional strategy for ruminants is, therefore, to ensure a nutrient non-limited microbial digestion in the rumen by complete supplementation. This automatically improves the P/E ratio in the nutrients available for digestion in the animal from poor quality feeds.
the protozoa in the rumen (Bird & Leng, 1978). This is not discussed at great length here because, as yet, there are no practical methods that are economic to accomplish the defaunated state in ruminants. However, a number of plants have been identified that contain antiprotozoal compounds which may have application in the use of these trees foliages in the future (see Leng et al. 1992). Immunisation against rumen protozoa has recently shown promise. In general, defaunation increases microbial growth yield by 20–25% as shown in trials with sheep cannulated at the level of the intestines (Viera et al. 1983) or with wool growth of sheep (Bird et al. 1979).
the diet. Feeding a supplement in which the protein has been made insoluble or otherwise non-degradable by rumen microbes is a major strategy to adjust the P/E ratio upwards, provided there is adequate ammonia production in the rumen to meet microbial requirements. Where the diet contains a soluble readily fermentable protein (e.g., leaf protein), it has been shown that including small amounts of bentonite results in more dietary amino acids reaching the small intestines of ruminants (Fenn & Leng, 1989). This is a new area for research, particularly where tree foliage is fed.
The reason for discussing the theoretical calculations of P/E ratio in the nutrients absorbed is to emphasize the large differences that can occur depending on the adequacy of the diet.
Ruminants on a diet containing a high level of bypass protein already have an appropriate level of essential amino acids absorbed relative to VFA energy (see Table 2.2 on page 18) and therefore it is not always necessary for the rumen microbes to grow efficiently as long as digestibility is optimized. The inefficient rumen will, however, have a higher heat of fermentation and produce more methane per unit of carbohydrate fermented; these are disadvantagous and again the increased heat from rumen fermentation may result in different effects in cool as compared with hot environments.
The ratio of P/E in the nutrients absorbed, through its effects on the efficiency of feed utilization has a large effect on growth, milk yield and, in particular, reproductive performance of ruminants (Perdok et al. 1988). The levels of production achieved when the P/E ratio is increased by supplementation in ruminants fed roughage based diets have been greatly superior to those predicted from the feeding standards based on metabolizable energy of a feed (see Figure 2.6 on the next page). The results for the efficiency of cattle growth where molasses and sugar cane are fed with supplements illustrate that low efficiency of utilization of even high energy diets may occur at times. The latter are the diets low in protein and which promote large population densities of protozoa in the rumen (Leng & Preston, 1976) and therefore the ratio of protein to energy in the nutrients absorbed is lower than in ruminants on high grain based diets where protozoal density is low or even zero (see Preston & Leng, 1986).
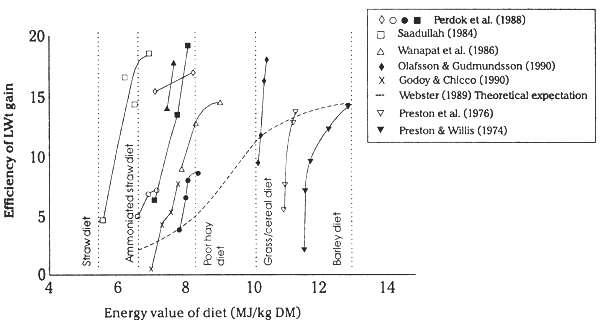
Forage for cattle in the tropics is usually 40–55% digestible. The metabolizable energy in the dry matter (M/D) thus ranges from 8.5 to 4.8 MJ/kg DM. According to feeding standards, a metabolizable energy content in a feed of 8.5 will support a liveweight gain of approximately 3g/MJ of ME intake in cattle. At 4.8MJ/kg DM intake, cattle should be in negative energy balance (see ARC, 1980 and also Webster, 1989). Contrast this with results of supplementary feeding trials based on balancing the nutrition of animals with urea/minerals and bypass protein, where growth rates equivalent to 18g/MJ of ME intake have been achieved in cattle fed straw (see Figure 2.6 on the preceding page). Obviously the presently accepted feeding standards are misleading for roughage based diets. The application of the basic concept of balanced nutrition, as discussed here, can improve animal growth by 2–3 fold and the efficiency of animal growth by as much as 6 fold over previous estimates (a range of 2–10 fold). Also, although growth rates of cattle are below those on grain based diets, cattle in the tropics on forage based diets can be as efficient in converting feed to liveweight gain as those in the temperate countries grazing quality pastures. Overall the ability of cattle to consume supplemented roughage has been underestimated by the feeding standards once the rumen is efficient and the balance of protein to energy is improved (McLennan et al. 1995).
The low productivity of tropical ruminant livestock has been accepted as an inevitable result of a poor feed base and a low feed conversion efficiency. This is no longer tenable. Application of the new feeding strategies detailed here will have a flow-on effect of improved reproduction, increased percentage of a herd in production, and an increased offtake from a herd. The benefit of improved reproduction outstrips the direct effects on liveweight gain or milk yield per se from the application of such strategies.
The challenge is to develop the necessary supplements and to get these to the animals in the various production systems. This is an important deviation from past practice. Vastly superior levels of production can be achieved with this approach as it is much more appropriate than replacing forage with concentrates to achieve increased levels of production.
With the above discussion in mind, the rôle of fodder trees in ruminant diets can be seen as three fold:—
as a N and mineral supplement to enhance fermentative digestion and microbial growth efficiency in the rumen of cattle on poor quality forage
as a source of post-ruminal protein for digestion. In this rôle the influence of secondary plant compounds in binding protein and making it insoluble is of particular importance
as a total feed, supplying almost all the biomass and other nutrients needed to support high levels of animal production.
A further rôle is presently emerging, that is the potential of constituent secondary plant compounds to manipulate the balance of micro-organisms in the rumen. The ability of secondary plant compounds to reduce protozoal populations has important implications for protein availability to ruminants (see Leng et al. 1992).
In the following chapters, the practical implications of tree foliages in providing deficient nutrients in the rumen are discussed. The comparisons made are between the data from feeding studies with foliages, as compared with data from feeding multinutrients aimed at correcting deficiencies for rumen microbes. The potential results of using bypass protein in addition to feeding deficient microbial nutrients are discussed in order to understand the potential benefits of using different foliages in the two roles or being able to process the same foliage to provide either rumen nutrients, bypass protein or both. This discussion, then, identifies the present inefficient use that is made of tree foliages in ruminant nutrition.
In their rôle as a complete food, fodder tree plantations offer alternatives to pastures. Such tree pastures are costly to establish and maintain, but the payoff in the yields of high protein foliages may, in some countries, warrant the management and fertilizer input necessary.
![]()
![]()
![]()