![]()
![]()
![]()
Scientists and extension agents feel most comfortable when they can give a farmer or hatchery manager an unambiguous piece of advice to solve or prevent a problem. Farmers and hatchery managers feel more comfortable when they receive unambiguous advice, because this means that there is only one way of doing something, and if they follow the advice they will solve or prevent the problem and become successful. To this end, a number of publications have recommended that aquaculturists or hatchery managers should maintain a specific Ne in order to prevent inbreeding - or genetic drift-related problems. These recommendations have ranged from 50 to 1,000; the majority suggest either 500 or 1,000.
Unfortunately, there is no single Ne and there is no simple set of genetic guidelines that can be prescribed for every farmer and every hatchery manager. Unlike disease prevention and treatment, the recommendations and protocols that are needed to prevent genetic problems vary from farmer to farmer, from farm to farm, and from programme to programme.
The reason why recommendations must be site-specific is because each farmer and each hatchery manager have different goals. Those for farmers with small farms will differ from those for farmers with medium to large farms. Those for fingerling producers will be different from those for farmers who simply grow fish for market. Finally, those for hatchery managers who manage populations that are being cultured to restock lakes and rivers will be radically different from all others, because the goals and plans for these programmes are to maintain and restore damaged natural fish stocks, not produce fish for market.
Proper management of a population, whatever the goal may be, begins with acquisition of the stock. The acquisition of a hatchery population is difficult and expensive, and this is where many genetic problems occur. The acquisition is of the utmost importance, and it may be the single most important aspect of brood stock management because this act determines the maximum amount of genetic variance and also determines how much inbreeding will be produced via future matings. What is acquired determines the population's genetic potential. In most cases, brood stock management after the population has been acquired is done just to keep the population from getting worse, through preservation of what was acquired by minimizing inbreeding and genetic drift. The first part of this chapter describes genetic aspects of acquiring a population and provides recommended Ne's that should be used during the acquisition process.
This second portion of this chapter synthesizes the information that was developed in Chapters 4, 5, and 7 to produce a series of recommendations on how large Ne must be to prevent inbreeding and genetic drift from causing genetic problems in hatchery populations when no selective breeding programme will be conducted. Recommendations will be made for populations at small, medium-sized, and large farms that produce fish for food and also for hatcheries that produce fish that will be stocked in fisheries programmes.
Except where noted, most recommendations in this section assume that farmers and hatchery managers cannot mark their fish and follow individual pedigrees. The only way to prevent inbreeding and genetic drift from causing genetic problems when these conditions exist is to manage a population's Ne.
Recommended Ne's were developed using the following assumptions: First, subsistence-level farmers should ignore the problem. Second, the size of the farm and the farmer's degree of sophistication determine how much management is needed; i.e., how much genetic risk the farmer should assume. Farmers who produce fingerlings for the local market should assume a low degree of genetic risk, which means that they should try and control inbreeding and genetic drift. Those who produce fish for themselves and some for the local market and who raise the fish using extensive management can assume more genetic risk; i.e., they can allow inbreeding to accumulate to fairly high levels and can ignore genetic drift. Third, the type of aquaculture programme determines how much genetic risk should be assumed. If the fish are being cultured to restock a lake or river, a very low level of genetic risk should be assumed, which means that a greater degree of genetic management is required than would be necessary if the fish were being raised for food. The only management options that make sense for programmes that raise fish for fisheries programmes are to accept as little genetic risk as possible and to minimize both inbreeding and genetic drift. A major goal in this type of fish culture is to conserve genotypic and genetic variance, and this can be accomplished only by minimizing inbreeding and genetic drift.
The final section discusses inbreeding and its management during a selective breeding programme. When a farmer conducts a selective breeding programme, his primary objective is to alter, not conserve, gene and genotypic frequencies in order to improve the population. Inbreeding is inevitable during a selective breeding programme, because each act of selection creates a bottleneck, and when select brood fish mate, a good percentage of the matings are consanguineous. Because of this, inbreeding is not the concern that it is in the absence of selection, when the only genetic goal is to conserve the status quo.
While inbreeding is not a major concern during selection, you do not want inbreeding to accumulate to levels where selection is being conducted simply to counteract inbreeding depression. Ways to minimize inbreeding during a selective breeding programme, as well as breeding techniques that can be used to produce genetically improved fish with 0% inbreeding for grow-out are described.
Genetic drift will also occur during a selective breeding programme. In fact, genetic drift can actually occur at a faster rate under these circumstances, because comparatively fewer fish are spawned than would be the case if random mating were used. Selection and preventing genetic drift are diametrically opposed breeding programmes. The goal of selection is to change gene frequencies and thus improve productivity, not conserve the status quo. Additionally, the major effects of genetic drift will be on genes that are not under direct or indirect selection. Consequently, prevention of genetic drift is not a major concern during a selective breeding programme.
The acquisition of a population is often the most important step in the management of the population. Mistakes that are made during acquisition can create permanent genetic damage, which can render meaningless the efforts to preserve the genetic quality of the population. You cannot manage and conserve what you do not have.
Before acquiring a population, a farmer or hatchery manager must ask the following questions: What is the purpose of the fish that will be acquired, and what do I want to do with them? For example, if a farmer wants to produce fast-growing fish for the local food market, he might want to purchase fish from another farmer or from a fingerling supplier who has a healthy, fast-growing strain. Conversely, he could acquire wild fish and create his own fast-growing strain. Both approaches will work; one will provide faster returns, while the other will enable the farmer to direct selection from the beginning and to create his own improved strain.
Hatchery managers who are raising fish that will be used to rebuild depleted populations in lakes and rivers should acquire fish from the lakes and rivers that will be stocked. It makes little sense to try and save an ailing population by stocking an exotic strain. This approach will often doom that population, especially if it is unique and small. If the stocked population hybridizes with the natural population, the stocking programme will cause the genetically unique population to disappear. If the stocked population out-competes the natural population for food or for spawning sites, the natural population will go extinct. It is counterproductive to raise a single strain and to use that strain for all stocking programmes. The stocks that are raised and stocked must be customized for the bodies of water where they are to be stocked.
The most important aspect of acquiring a population and the one that will have the greatest effect throughout its culture history is the genetic size of the acquired population. The Ne that is obtained will determine how much genetic variance exists and it will thus determine future inbreeding. If the Ne of the acquired population is small, the foundation generation will be a bottleneck, and this creates what is called the “founder effect.” The effect that this has on inbreeding can be dramatic, but it often plateaus after a few generations. However, the long-term effects on genetic variance can be disastrous. A bottleneck that occurs in the foundation generation can permanently rob the population of many alleles that will be needed if the fish are to be stocked in the wild or if future selective breeding programmes are to be conducted.
This problem even occurs unintentionally, due to the fecundity of fish and the inability to identify individuals. I once saw someone ship 2,000 full-sibs (a single family produced by one mother and one father) to fill a request for a foundation population. The recipient thought he was receiving a population with a potential Ne of 2,000. The Ne of the fish that were received might have been 2,000, but the Ne that produced the foundation population was only 2. That foundation population had little genetic variance, and all first-generation offspring would have had F = 25% (more if the two parents were related). Although this example is extreme, many hatchery stocks have been started with small foundation populations that were not that much larger. Even when better efforts are taken to obtain as large an Ne as possible, the fish that are sampled are often produced by only a handful of brood fish.
The history of a stock should be examined before it is acquired. Obviously, the production history of the fish needs to be determined: growth rate, survival, fecundity, disease history, ease of spawning, etc. If possible, this information should be determined for every year that the fish have been at the hatchery. A progressive decrease in yield, survival, or fecundity might be clinical signs of inbreeding depression.
One other bit of information is equally important: the Ne for every generation. Even if Ne's are unknown, data sheets that list the number of fish that spawned will allow you to determine approximate Ne's for every generation.
Good stocks should not be discarded simply because the Ne's are smaller than desired. Background information about the strain's Ne's is simply another tool that will enable you to better evaluate the strain and to determine if the fruits of a farmer's labour are sweet or bitter.
One reason you want to determine the strain's history is so that you do not re-acquire the strain you already have. Many farmers acquire their foundation stock from a single good source and then rename the strain. A background check on the fish might reveal that all farmers in a region have the same strain, but it has a different alias on each farm.
If the foundation stock is going to come from a wild population, everything should be done in order to acquire a large Ne. It is often assumed that the most efficient way to do this is to randomly sample the population. This can work, but a population often assumes a non-random distribution, so a random sample will not accurately reflect the make-up of the population. If the population assumes a non-random distribution, use the natural history of the population and the way it is distributed in the lake or river to collect as large an Ne as possible. One way to accomplish this is to sample from many locations and to sample over time.
If the life history of the fish is well known and if the fish lay eggs masses, one way to collect a foundation stock with a large Ne is to obtain samples from many egg masses. If each egg mass is produced by a single male and female (or a known number of multiple males and females), the Ne of the fish that produced the foundation population can be determined. Egg masses should be collected over the entire spawning season, or the collection will be a form of selection that eliminates some genetic variance and reduces Ne from what it could have been. If egg masses are obtained, the families can be raised in individual tanks until they can be marked.
How large should the Ne of the foundation population be? Recommended Ne's that are presented in the next section can be used. However, more fish have to be collected than are needed to create the desired Ne, because of mortalities caused by transportation, disease, etc.
When no selective breeding programme will be conducted, the only genetic goal of brood stock management is to prevent changes in the gene pool; this is accomplished by minimizing inbreeding and genetic drift. Such management will prevent genetic decreases in yield, etc. However, not all farmers need to be concerned about these problems or need to manage their populations to prevent them from occurring. Separate sets of guidelines are presented for different farm sizes and the goals that the farmers or hatchery managers might have for the populations that are being cultured.
Most farmers who own small fish farms (arbitrarily defined as farms <2 ha) should not be concerned about inbreeding or genetic drift. Many of these farmers are subsistence-level farmers, and yields and profits can be increased far more if these farmers learn how to use proper fertilization techniques, to manage water quality, to use supplemental feeding, etc. Improvements in these areas of management will be a more effective use of the farmer's time and financial resources. If the farmer increases yield by using better management, his family will have more food, and he may be able to sell surplus fish.
Not all farmers who own small fish farms are subsistence-level farmers. Some might be quite sophisticated and use state-of-the-art management. Those farmers might be able to incorporate inbreeding control and/or prevention into yearly management plans. Those who could benefit from this additional type of breeding management should follow the recommendations that are outlined in the next section on medium-sized farms.
Farmers who own medium-sized fish farms (arbitrarily defined as those that have 2 ha of ponds) range from subsistence-level to sophisticated, so a wide range of management options is available. The recommendations depend on a farmer's type of management and his long-term goals. As before, subsistence-level farmers should not worry about genetic aspects of management. They need to improve the environmental aspects first (feeding, fertilization, etc.).
Most farmers who own medium-sized fish farms should be concerned with preventing only inbreeding problems, but some will also want to prevent genetic drift-related problems. Table 8 provides guideline minimum constant Ne's for fish farmers who own medium-sized fish farms. The Ne's presented in Tables 8, 9, and 10 were compiled from values given in Tables 4 and 5 (pages 79 and 81).
Four options are presented in Table 8. The first two are for farmers who are willing to accept a high level of “genetic risk.” In this case, risk means that the farmers are willing to allow inbreeding to accumulate to 10% and want to try and save alleles whose frequencies are 0.05. It can be argued that these farmers should ignore genetic drift, and most should not worry about it; however, an examination of Table 4 shows that the Ne's that are needed to prevent inbreeding from exceeding 10% are quite small for the first 10 generations (≤50), so the addition of a few extra brood fish to prevent mild levels of genetic drift is easily accomplished. In this case, the recommendation is to have either a 95% or 99% guarantee of keeping alleles whose frequencies are 0.05. The 0.05 frequency level was chosen because it is an order of magnitude more difficult to save alleles rarer than f = 0.05 (Table 5).
It is also likely that many alleles that are rarer than f = 0.05 are lost due to domestication selection. If rare alleles are valuable (they increase survival, make fish less excitable, improve reproductive success, or improve other phenotypes that are important at fish farms), domestication selection will increase the frequencies dramatically. Additionally, unless the farmer is using state-of-the-art management, improvements in the environmental aspects of management will have a far greater impact on yields and profits than improvements in the genetic aspects of management.
The recommended minimum Ne's for this type of farmer and this level of genetic risk range from 30 to 100 (Table 8), depending on the number of generations that the farmer wants to incorporate into his plan. Since most farmers will at most plan ahead for only a few generations, the recommended Ne's that are needed by most farmers are quite small; in many cases Ne will be ≤50. If a farmer only wants to prevent inbreeding from reaching certain levels, the Ne's listed in Table 4 can be used.
Table 8. Minimum constant effective breeding numbers (Ne's) needed to prevent inbreeding- and genetic drift-related problems on medium-sized fish farms. Minimum Ne's are presented for four options: Two are for farmers who are willing to accept a higher level of risk-who want to keep inbreeding from exceeding 10% and want either a 95% or 99% guarantee of keeping alleles whose frequencies are 0.05. The other two options are for farmers who want less risk-who want to keep inbreeding from exceeding 5% and want either a 99% guarantee of keeping alleles whose f = 0.05 or a 95% guarantee of keeping alleles whose f = 0.01.
| No. generations | Keep F≤10%; 95 % guarantee for alleles f=0.05 | Keep F≤10%; 99% guarantee for alleles f=0.05 | Keep F≤5%; 99% guarantee for alleles f=0.05 | Keep F≤5%; 95% guarantee for alleles f=0.01 |
| 1 | 30 | 45 | 45 | 150 |
| 2 | 36 | 52 | 52 | 183 |
| 3 | 40 | 56 | 56 | 203 |
| 4 | 43 | 59 | 59 | 218 |
| 5 | 45 | 61 | 61 | 229 |
| 6 | 47 | 63 | 63 | 238 |
| 7 | 48 | 64 | 70 | 245 |
| 8 | 50 | 66 | 80 | 252 |
| 9 | 51 | 67 | 90 | 258 |
| 10 | 52 | 68 | 100 | 263 |
| 15 | 75 | 75 | 150 | 283 |
| 20 | 100 | 100 | 200 | 297 |
More sophisticated farmers will want to accept less genetic risk, which is defined as preventing inbreeding from exceeding 5% and producing either a 99% guarantee of keeping alleles whose frequencies are 0.05 or a 95% guarantee of keeping alleles whose frequencies are 0.01. Farmers who are using intensive management and who want to have relatively few genetic problems should try and save alleles whose frequencies are 0.01, because this will virtually guarantee that alleles that are more common than 0.01 will be saved. The recommended minimum Ne's that are needed for this level of genetic risk range from 45 to 297, depending on the number of generations that are incorporated into the management plan. An Ne of 61 will prevent inbreeding from exceeding 5% and will produce a 99% guarantee of keeping alleles whose frequencies are 0.05 for five generations, while an Ne of 229 will prevent inbreeding from exceeding 5% and will produce a 95% guarantee of keeping alleles whose frequencies are 0.01 for five generations.
Many farmers will be able to incorporate these recommended Ne's into their management plans without incurring additional expense, because a 2-ha farm requires dozens to hundreds of brood fish in order to produce enough fingerlings for stocking. For example, a tilapia farmer will need 50 to >200 brood fish in order to stock his grow-out ponds.
Farmers who own large fish farms (arbitrarily defined as those that have >2 ha of ponds) generally use intensive levels of management because such farms produce fish for local, regional, and national markets. Some farmers will use low levels of technology, but by and large, such farmers will be more likely to use state-of-the-art management.
Farmers who use extensive management can accept a higher level of risk (less genetic management) than those who use more intensive management. Table 9 provides recommended Ne's for large farms. The higher-level risk option means preventing inbreeding from exceeding 10% and providing 95% and 99% guarantees of keeping alleles whose frequencies are 0.05. Recommended Ne's range from 30 to 250, depending on the number of generations that will be incorporated into the work plan.
Table 9. Minimum constant effective breeding numbers (Ne's) needed to prevent inbreeding- and genetic drift-related problems on large fish farms. Minimum Ne's are presented for four options: Two are for farmers who are willing to accept a higher level of risk-who want to keep inbreeding from exceeding 10% and want either a 95% guarantee of keeping alleles whose f=0.05 or a 99% guarantee of keeping alleles whose f=0.05. The other two options are for farmers who want less risk-who want to keep inbreeding from exceeding 5% and want either a 95% guarantee of keeping alleles whose f=0.01 or a 99% guarantee of keeping alleles whose f= 0.01.
| No. generations | Keep F≤10%; 95% guarantee for alleles f=0.05 | Keep F≤10%; 99% guarantee for alleles f=0.05 | Keep F≤5%; 95% guarantee for alleles f=0.01 | Keep F≤5%; 99% guarantee for alleles f=0.01 |
| 1 | 30 | 45 | 150 | 230 |
| 2 | 36 | 52 | 183 | 264 |
| 3 | 40 | 56 | 203 | 284 |
| 4 | 43 | 59 | 218 | 298 |
| 5 | 45 | 61 | 229 | 309 |
| 6 | 47 | 63 | 238 | 319 |
| 7 | 48 | 64 | 245 | 326 |
| 8 | 50 | 66 | 252 | 333 |
| 9 | 51 | 67 | 258 | 339 |
| 10 | 52 | 68 | 263 | 344 |
| 15 | 75 | 75 | 283 | 364 |
| 20 | 100 | 100 | 297 | 378 |
| 25 | 125 | 125 | 308 | 390 |
| 30 | 150 | 150 | 318 | 399 |
| 35 | 175 | 175 | 350 | 406 |
| 40 | 200 | 200 | 400 | 413 |
| 45 | 225 | 225 | 450 | 450 |
| 50 | 250 | 250 | 500 | 500 |
Farmers who use more intensive levels of management to produce food or who are fingerling producers should accept less risk (more genetic management). In this case, the farmer should try and prevent inbreeding from exceeding 5% and should try and produce 95% or 99% guarantees of keeping alleles whose frequencies are 0.01. Recommended constant Ne's for this option range from 150 to 500, depending on the number of generations that are incorporated into the work plan.
As was the case with medium-sized farms, most farmers who own or manage large farms should be able to incorporate these recommendations into hatchery management plans. For example, the Ne's needed for the higher risk option for five generations are 45 and 61, while those for the lesser risk option are 229 and 309. A constant Ne of 344 will keep inbreeding from exceeding 5% and will produce a guarantee of keeping alleles whose frequencies are 0.01 for 10 generations. It should be relatively easy to incorporate this type of genetic management into yearly work plans on large farms, because the Ne's listed in Table 9 should be no larger than those that most farmers use to achieve fingerling production quotas.
Hatchery managers who produce fish that will be stocked in lakes and rivers in order to restore damaged fisheries have an entirely different management goal. The goal in this type of aquaculture is to restore an ailing fisheries by stocking a self-reproducing population. Consequently, the genetic goal is to maintain as much genetic diversity as possible. The hatchery manager does not want to alter phenotypic or genotypic frequencies by selection, domestication, inbreeding, or genetic drift.
Table 10. Minimum constant effective breeding numbers (Ne's) needed to prevent inbreeding- and genetic drift-related problems in hatchery stocks that are used for fisheries management projects to restock lakes and rivers. Minimum Ne's are presented for four options: moderate risk, which is to keep inbreeding from exceeding 5% and to produce a 99% guarantee of keeping alleles whose f = 0.01; low risk, which is to keep inbreeding from exceeding 5% and to produce a 99% guarantee of keeping alleles whose f = 0.005; little risk, which is to keep inbreeding from exceeding 1% and to produce a 99% guarantee of keeping alleles whose f= 0.005; and no risk, which is to keep inbreeding from exceeding 1% and to produce a 99% guarantee of keeping alleles whose f=0.001.
| No. generations | Keep F≤5%; 99% guarantee for alleles f=0.01 | Keep F≤5%; 99% guarantee for alleles f=0.005 | Keep F≤1%; 99% guarantee for alleles f=0.005 | Keep F≤1%; 99% guarantee for alleles f=0.001 |
| 10 | 344 | 689 | 689 | 3450 |
| 15 | 364 | 730 | 730 | 3653 |
| 20 | 378 | 758 | 1000 | 3797 |
| 25 | 390 | 780 | 1250 | 3908 |
| 30 | 399 | 799 | 1500 | 3999 |
| 35 | 406 | 814 | 1750 | 4076 |
| 40 | 413 | 827 | 2000 | 4143 |
| 45 | 450 | 839 | 2250 | 4202 |
| 50 | 500 | 850 | 2500 | 4255 |
| 55 | 550 | 859 | 2750 | 4302 |
| 60 | 600 | 868 | 3000 | 4346 |
| 65 | 650 | 876 | 3250 | 4386 |
| 70 | 700 | 883 | 3500 | 4423 |
| 75 | 750 | 890 | 3750 | 4457 |
| 80 | 800 | 896 | 4000 | 4489 |
| 85 | 850 | 903 | 4250 | 4520 |
| 90 | 900 | 908 | 4500 | 4548 |
| 95 | 950 | 950 | 4750 | 4750 |
| 100 | 1000 | 1000 | 5000 | 5000 |
The reason why the genetic goal is to maintain as much genetic diversity as possible and not alter it during the fish culture phase of the operation is that no one knows what alleles or allelic combinations are important for survival in the wild. Also, no one can predict which alleles will become important in the future. Environments change, and it is difficult to predict what they will be like in the future and what alleles will be important.
This means the genetic goal is to prevent or minimize inbreeding and genetic drift. Table 10 contains recommended Ne's for four management options. The management options presented in Table 10 go from the 10th to the 100th generation. The reason why no options are given for generations 1–10 is because this type of management requires long-term planning. If short-term options are unavailable, hatchery managers and fishery department officials are not tempted to take shortcuts and go for short-term benefits at the expense of the long-term success of the programme. The number of generations that should be incorporated into the programme will depend on the generation interval. Fish with generation intervals ≤1 year will require more than those with generation intervals of 3–4 years, simply because the smaller the generation interval, the more quickly inbreeding can accumulate during a specific time period. For example, if you want to manage a population for 12 years, a species with a 4-year generations interval requires only three generations of management; a species with a 1-year generation interval requires 12 generations of management. A minimum of 10 years should be incorporated into the work plan, but if possible, 25 generations should be considered a minimum. Conservation biology (stocking to restore a fisheries is a type of conservation biology) requires long-term planning if it is to succeed.
In this type of aquaculture, the definitions of risk are different than they are for fish farming. Here, moderate and low risk means preventing inbreeding from exceeding 5% and trying to produce 99% guarantees of keeping alleles whose frequencies are either 0.01 (moderate risk) or 0.005 (low risk), respectively. Effective breeding numbers for these options range from 344 to 1,000, depending on the number of generations.
A third option is to accept little risk, which means preventing inbreeding from exceeding 1% and producing a 99% guarantee of keeping alleles whose frequencies are 0.005. Effective breeding numbers for this option range from 689 to 5,000, depending on the number of generations.
The best option, in terms of genetic conservation, is to accept no genetic risk-to prevent inbreeding from exceeding 1% and to produce a 99% guarantee of keeping alleles whose frequencies are 0.001. This option requires Ne's that range from 3,450 to 5,000, depending on the number of generations.
Obviously, there will be a compromise between what is best, in terms of genetic conservation (the no risk option), and what is practical, in terms of the budget and available manpower. Effective breeding numbers needed for the first two options are not unreasonably large. For example, Ne's needed to achieve the genetic goals for 25 and 50 generations are 390 and 500, respectively, for the moderate risk option and 780 and 850, respectively, for the low risk option. Although these options are not as effective as the others, both should produce populations that have minimal genetic damage, although the effects of 5% inbreeding on survival in the wild are not known. One study found that inbreeding in Atlantic salmon reduced the return rate of stocked fish.
The third option (little risk) is an improvement on the second (moderate risk), in that inbreeding will never exceed 1%. Effective breeding numbers needed to achieve the genetic goals for 25 and 50 generations are 1,250 and 2,500, respectively. While the Ne's required for this level of genetic risk might be larger than the number of brood fish that can be maintained or spawned at some hatcheries, such Ne's are realistic for many hatcheries that routinely spawn several thousand fish.
The no risk option will prevent inbreeding from exceeding 1% and will produce a virtual guarantee of keeping polymorphic genes in the polymorphic state. Effective breeding numbers needed to produce the genetic goals for this management option are 3,908 for 25 generations and 4,255 for 50 generations. Most hatcheries will not be able to use this option, simply because they will not be able to hold or spawn this many fish. However, if this option can be used, it will produce a population of fish that has a better chance of surviving to become a self-reproducing population. If this occurs, the hatchery aspect of the project will have succeeded and can be ended.
An option that can be used to prevent hatchery-induced inbreeding or genetic drift problems is to wild-source new brood fish every two to three generations. This option is not always available, especially if the target lake or river has been damaged by pollution or by over-fishing; also, the option is expensive and requires additional skilled labour. But if this option is incorporated into the work plan, it might be a way to prevent domestication selection from altering the gene pool, and it is a way of preventing inbreeding and can minimize genetic drift. Even if the population cannot be replaced every few years, the same effect might be achieved if 10–25% of the brood fish are replaced on a yearly basis.
Some aspects of management might have to be altered in order to achieve the desired Ne. The sex ratio should be 1:1 or as close to that as possible. If possible, more matings should be made than are needed, and only an equal portion of each egg mass should be used. If gametes are stripped, eggs and sperm should not be pooled before fertilization. After fertilization, families should be raised in individual tanks so that mortalities can be determined and the effects on Ne can be quantified. If each brood fish is allowed to spawn only once and if it mates only with one other fish, Ne will be easy to determine.
In addition, the hatchery manager must be able to practice what is called “no selection.” No selection is the absence of any type of selection that will alter the population's gene pool. Matings should be random, and no fish should be chosen or excluded for any reason. This means fish must be spawned over the entire spawning season, not when it is convenient for the hatchery manager. Fish that mature at the beginning and at the end of the season must also be spawned. The number of matings that are made during a week should be determined
by the percentage of fish that ripen during that period. There is a tendency to spawn only those fish that are ready during the middle of the spawning season, because it is an efficient use of labour. However, this is a form of selection, and it will lower Ne and will change gene frequencies. Studies have shown that spawning time can be radically altered by selective breeding.
If possible, fish should be tagged so that pedigrees can be maintained. This will enable hatchery managers to determine Ne with greater confidence. New tagging techniques have been developed that are easy, reliable, and inexpensive.
Finally, there must be some way to assess the effect of the management programme on inbreeding and genetic drift. Effective breeding number can be determined, and this will enable hatchery managers to predict what the effects on inbreeding and genetic drift will be; however, this is not empirical proof that the management programme has produced the desired results. Studies have found that genetic drift occurs despite efforts to prevent it.
The effects of the management programme on the population's gene pool can be determined by using electrophoresis. Electrophoresis is a biochemical technique that enables geneticists to determine protein phenotypes and their genotypes. This technology has become the most important genetic tool for population biologists, because it enables them to study a population's genes over time and space.
Electrophoresis is a technique that examines proteins that are taken from eye, liver, heart, blood, or muscle. The tissue is ground and turned into a liquid. Small amounts of the liquid are then inserted into sheets of starch or other material called “gels”. An electric current is run through the gels, and this causes the proteins to migrate through the gel. The distance the proteins move is determined by their size and chemical composition. After a predetermined time, the electric current is stopped, and the gel is stained to reveal specific protein phenotypes and their genotypes. The way protein phenotypes and genotypes are determined is illustrated in Figure 34.
These evaluations should be done with what are called “neutral” genes (genes where different protein phenotypes are of equal value; i.e., both work equally well, and neither phenotype provides a fish with an increase in fitness). If non-neutral genes are used, the effects of selection could be confounded with inbreeding or genetic drift.
If this is done every generation, gene frequencies can be determined, and the frequencies over time can be compared. This will provide a good estimate of genetic drift. Additionally, this technique can be used to determine inbreeding. The percentage of homozygotes can be determined each generation. Since inbreeding increases homozygosity, any increase that is measured will approximate the amount of inbreeding.
If the population is examined electrophoretically every generation, information on gene frequencies and linkage disequilibrium can be used to assess the population's Ne, which will provide a good estimate of inbreeding. Effective breeding numbers that are determined using this information provide valuable information about the population, because Ne is often smaller than that which is determined by counting the fish that spawn. If Ne is determined indirectly from electrophoretic data, differential reproduction and survival are taken into account, and this provides a more accurate estimate of Ne.
The only drawback to electrophoresis is the cost. A well-equipped lab costs up to US$100,000, and it can cost several dollars to examine each fish. In addition, these tests require highly trained personnel. Many universities and research businesses have electrophoresis labs and market these services, which means a hatchery does not need to have its own genetics lab in order to examine its population electrophoretically. Even though these tests can be expensive, if the prevention of inbreeding and the problems associated with genetic drift are a crucial aspect of brood stock management, this technique is a valuable tool that will reveal whether the management programme is succeeding. And it is the only way to quantify the effects of management on the population's gene pool.
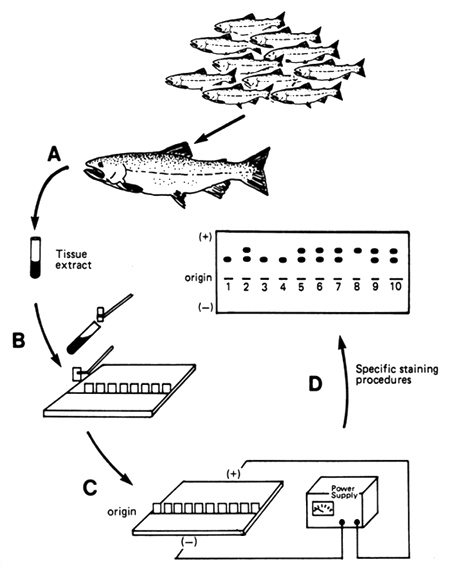
Figure 34. Schematic diagram of how protein phenotypes and genotypes are determined electrophoretically. Protein from various tissues are taken from fish (A), and extracts are placed in gels (B). Electricity is run though the gels (C) to cause the protein to migrate. The gels are stained to reveal the protein phenotypes (D). The stained gel in this figure reveals the phenotypes for 10 fish. The phenotypes are the bands that are revealed by the stain. In this case, there are three possible phenotypes. Heterozygotes have two bands and the two homozygotes each have a single band; consequently, the gene that produces these phenotypes has two alleles, and each allele produces a band. The frequency of each allele can be determined by adding the number of alleles that are revealed by the phenotypes (bands) and dividing that by the total number of alleles. If only one band is present, it is counted twice, because that fish is a homozygote (two copies of the same allele). Since there are 10 fish, there are 20 total alleles. If the top allele is the D allele and the bottom is the D' allele, the frequency of the D allele is 8/20 or 0.4, and the frequency of the D' allele is 12/20 or 0.6.
Source: Utter, F., P. Aebersold, and G. Winans. 1988. Interpreting genetic variation detected by electrophoresis. Pages 21–45 in N. Ryman and F. Utter, eds. Population Genetics & Fishery Management. Washington Sea Grant Program, University of Washington Press, Seattle, Washington, USA. Reprinted by permission from Washington Sea Grant Program.
The recommended Ne's presented in Tables 8, 9, and 10 were created from a combination of published information and educated guesswork. The effects that these Ne's have on inbreeding and genetic drift in hatchery populations have not been tested empirically. It is unlikely that they ever will be evaluated, because such studies would be difficult and expensive; furthermore, such studies would take many generations-in some cases, >25 generations-simply to determine if a population behaved as expected.
Because of this and because Ne's must be customized for each farmer or hatchery manager and for each farm or hatchery, many fish culturists will want to create their own set of recommendations and determine a constant Ne that best suits their needs. This can be accomplished by using the Ne's presented in Tables 4 and 5 (pages 79 and 81). Fish culturists who only want to prevent inbreeding depression can use the Ne's in Table 4, while those who wish to prevent both inbreeding depression and the ravages of genetic drift need to use both tables.
The information in Tables 4 and 5 can be used in two ways: One, a farmer or hatchery manager can use the tables to determine the constant Ne that is needed to keep inbreeding from exceeding a predetermined level and which is needed to produce a certain guarantee of keeping alleles of a given frequency for a predetermined number of generations. Two, a farmer or hatchery manager can use the tables to determine how much inbreeding and what guarantee of keeping alleles a specific constant Ne will produce over various numbers of generations.
For example, if a farmer decides he wants to keep inbreeding from exceeding 6% and he wants to produce a 95% guarantee of keeping alleles whose frequencies are 0.05 for eight generations, he will find that the Ne from Table 4 is 67, while that from Table 5 is 50. Since he wants to achieve both goals, he needs to use the larger Ne, which means he needs a constant Ne of 67 to achieve his goals.
Conversely, if a hatchery manager knows that he can maintain a constant Ne of 1,250 but would have difficulty in maintaining one larger than that, he can can use Tables 4 and 5 to determine how effective this Ne would be in minimizing inbreeding and the effects of genetic drift. Table 4 reveals that a constant Ne of 1,250 will produce: F = 1% at generation 25; F = 2% at generation 50; F = 3% at generation 75; F = 4% at generation 100. Table 5 shows that this Ne will produce a >99% guarantee of keeping alleles whose frequencies are 0.005 for 100 generations. This information reveals that a constant Ne of 1,250 will do an excellent job of preventing inbreeding and genetic drift from adversely affecting the population, provided bottlenecks have not already crippled it genetically.
The ability to customize Ne's is important. If a farmer or hatchery manager is given a set of guidelines and is told that he must maintain an Ne which he feels is impossible, impractical, or prohibitively expensive, he will simply ignore the advice and feel that the genetic aspects of brood stock management are more trouble than they are worth. On the other hand, if a farmer or hatchery manager can customize his Ne and knows what that Ne can accomplish genetically, he is more likely to incorporate genetics into brood stock management.
Selection and prevention of inbreeding and genetic drift are, for practical purposes, incompatible genetic goals. The way to minimize inbreeding is to maintain a large Ne or to prevent consanguineous matings. Selection is a breeding programme that reduces the population's Ne by culling (eliminating) fish that fall below a cut-off value (minimal acceptable phenotypic value) and that allows only select brood fish (those whose phenotype[s] meet or exceed the cut-off value) to mate. When you mate the best with the best, you often mate relatives, which produces inbreeding. This has led some to call selection a form of directed inbreeding. All populations that have been improved by selection are inbred to a degree; some are extremely inbred.
Even though some inbreeding is inevitable, there are some techniques that can be used to moderate inbreeding so that it does not counteract selection. There is no point in conducting a selective breeding programme simply to offset inbreeding depression. The idea is to minimize inbreeding-but not at the expense of the selective breeding programme.
Genetic drift will also occur during a selective breeding programme. Selection and the conservation of genetic variance are opposite types of genetic management. Selection is designed and conducted to change gene frequencies and eliminate many alleles-that is its purpose. The goal is to change the frequencies of the alleles at the loci that control the phenotypes that are under selection. But selection will also change other gene frequencies because of linkage groups (groups of genes are “linked” when they are located on the same chromosome) and because of indirect selection (selection for one phenotype that affects another; e.g., selection for length also selects for weight and food conversion). Genetic drift is not the concern that it is when no selection is being conducted, because the major effects of genetic drift will be on genes that are not affected by the selective breeding programme. Consequently, farmers and hatchery managers should not be overly concerned about genetic drift when they conduct a selective breeding programme.
There are two basic types of selection. The simplest is individual (also called “mass”) selection, where selection is based on individual phenotypic values. Fish that meet or exceed the cut-off value are saved, and the others are culled. Family relationships are totally ignored; an individual is selected or culled based on its own merit.
Inbreeding is inevitable when conducting individual selection. Because family relationships are ignored during the selection process, a majority of the select brood fish can come from only a few families, particularly if one or two matings produce outstanding offspring. When this occurs, the population goes through a severe bottleneck which generates inbreeding. Figure 35 shows how a bottleneck can be produced during individual selection and why inbreeding occurs.
Inbreeding is also produced because family relationships are usually ignored during the mating process when select brood fish are spawned. This is inevitable if fish are not marked. If family relationships are ignored when matings are made, relatives will be mated; among these matings will be brother-sister and half-sib matings, which produce considerable inbreeding.
Even though inbreeding cannot be prevented when conducting individual selection and when the fish are not marked, the rate of inbreeding might be minimized by maintaining Ne at a predetermined size. The Ne's in Table 4 can be used as guidelines; however, inbreeding will be greater than that indicated in Table 4. The Ne's in Table 4 are those that will keep inbreeding from exceeding undesired levels when there is no selection and when random mating occurs. When a selective breeding programme is being conducted, matings are not random; they are what is called “assortative matings” (the best are mated with the best). Additionally, each act of selection creates a mini-bottleneck, which can retroactively lower the Ne of the previous generation.
One way to prevent or reduce inbreeding is to mark fish and avoid consanguineous matings when select brood fish mate. This might decrease the genetic gain, but it will prevent inbreeding. If this mating programme is used, the population must be sufficiently large so that adequate numbers of select brood fish can be spawned without consanguineous matings. If relatives are not allowed to mate, the number of potential matings that can be made among the select brood fish will decrease dramatically.
After a few generations, it might not be possible to prevent consanguineous matings, but if brother-sister matings or half-sib matings can be avoided, large accumulations of inbreeding can be prevented. If only cousins are allowed to mate, relatively little inbreeding will accumulate in the select population. Five generations of first cousin matings will produce less inbreeding than a single generation of brother-sister matings. If only second cousins are allowed to mate, inbreeding will never exceed 2%.
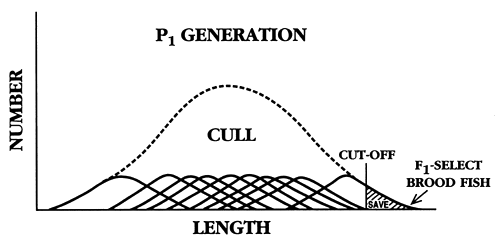
Figure 35. This graph shows why inbreeding occurs during individual selection. The size distribution of a population and the size distributions of the 10 families that make up the population are shown. The placement of the cut-off value produced a population of select brood fish in which every individual came from a single family. The selection process retroactively lowered the Ne of the P1 generation to 2. When the F1-select brood fish are spawned, all fish in the F1-select generation will have F = 25%. This example is a bit extreme, but it illustrates why inbreeding is inevitable when individual selection is conducted.
After: Newkirk, G.F. 1979. A discussion of possible sources of inbreeding in hatchery stock and associated problems. Proceedings of the World Mariculture Society 10:93–100.
A second approach that can be used to moderate inbreeding is to divide the population undergoing selection into cohorts (sub-populations). Dividing a population into cohorts is a breeding technique that can be used to improve the efficiency of selection. For example, a population can be divided into cohorts based on spawning date to neutralize the effect of age (number of growing days and water temperature during those days) on size; this prevents these environmental sources of variance for growth from becoming confounded with heritable differences during selection. This approach can also be used to minimize inbreeding.
If a population is divided into a number of cohorts, selection is conducted independently in each cohort, and select brood fish from each cohort are mixed before they are spawned. By conducting selection in this manner, the number of parents that produce select brood fish will increase, which reduces the inbreeding that will be produced. No further selection is allowed after select brood fish are mixed, because a second round of selection could eliminate one or more cohorts, which would negate some of the effort that has been expended, and it would also increase the rate of inbreeding.
If select brood fish from each cohort are marked or are held in separate ponds or tanks until they can be spawned, a breeding programme called “rotational mating” can be used to prevent inbreeding for a number of generations (Figure 36). The number of inbreeding-free generations of selection that can be produced depends on two factors: the number of cohorts and how cohorts are produced after the first generation of selection.
During the first generation of rotational mating, F1-select brood fish are mated as follows: females from cohort 1 are mated to males from cohort 2, females from cohort 2 are mated to males from cohort 3, etc. Each mating combination is used to produce an F1-select generation cohort. A second generation of selection occurs in each F1-generation cohort, and when F2-select brood fish are mated, the mating pattern is staggered: females from the first cohort are mated to males from the third cohort, etc. Each mating combination is used to produce an F2-generation cohort. A third generation of selection occurs in each F2-generation cohort, and when F3-select brood fish are mated, the mating pattern is staggered: females from the first cohort and mated to males from the fifth cohort, etc. The number of cohorts needed to prevent inbreeding after the first generation of selection doubles for each additional generation:
| Number of cohorts | Select generation when inbreeding will be produced. Select brood fish that produced these fish will have F = 0% |
| 2 | F2 |
| 4 | F3 |
| 8 | F4 |
| 16 | F5 |
| 32 | F6 |
If mating combinations cannot be used to produce the cohorts as described above as and as illustrated in Figure 36 after the first generation of selection, new cohorts must be created from a pooled population of select brood fish every generation. If this approach is used, rotational mating will prevent inbreeding in the F1-select generation, but it will only minimize it thereafter. After the first generation of selection, rotational mating can be used to prevent brother-sister matings, which will slow the rate at which inbreeding is produced.
A major liability of this method of preventing or minimizing inbreeding is the cost. It is more expensive to produce and maintain a number of cohorts over successive generations than to conduct selection in a population as a whole, and the cost of the programme escalates as the number of cohorts increases.
A second liability is the fact that if the cohorts aren't needed to control a major source of VE (e.g., spawning date) and to prevent it from being confounded with VA during selection, dividing a population into cohorts will decrease response to selection. Response to selection is greater in a single large population than in one of equal size that is divided into subpopulations.
The use of cohorts prevents inbreeding for a limited number of generations. Eventually, inbreeding will be produced. Once inbreeding is produced it usually accumulates rather rapidly; eventually the level of inbreeding will approach that which would have occurred without the use of cohorts. However, the use of cohorts can be quite useful in preventing inbreeding for four to six generations and in minimizing inbreeding for several generations thereafter.
If the production of inbreeding-free fish for grow-out is a top priority, crossbreeding can be combined with selection to produce fish with no inbreeding generation after generation. In this breeding programme, selection is conducted simultaneously in two strains that were previously shown to produce outstanding F1 hybrids; selection in the two strains can be for either the same phenotype or for different traits. After the first generation of selection, some of the select brood fish are hybridized to produce F1 hybrids for grow-out or for sale to other farmers. The F1 hybrids will have F = 0%, since the parents are not related. Other select brood fish from both strains undergo a second generation of selection. This process is continued every generation (Figure 37). All F1 hybrids are sold, and none is allowed to reproduce. If all F1 hybrids are sold and if the two strains are kept genetically isolated from each other, F1 hybrids with F = 0% can be produced generation after generation.
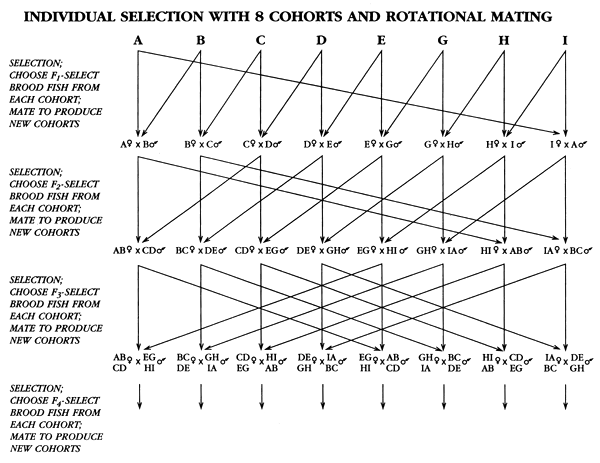
Figure 36. Schematic diagram of rotational crossing, a mating programme that can be used to prevent or minimize inbreeding during individual selection when the population is broken into cohorts. In this figure, there are eight cohorts (A–I). After the first generation of selection, F1-select brood fish from each cohort are mated as follows: Females from cohort A are mated to males from cohort B; females from cohort B are mated to males from cohort C, etc. Each of these mating combinations forms a new cohort. A second generation of selection occurs, and F2-select brood fish are mated as follows: females from the first cohort (A♀ × B♂) are mated to males from the third cohort (C♀ × D♂), etc. After the third generation of selection, F3-select brood fish are mated as follows: females from the first cohort (AB♀ × CD♂) are mated to males from the fifth cohort EG♀ × HI♂). etc. The rotational mating programme outlined in this figure for eight cohorts produced no inbreeding during this selective breeding programme. Inbreeding will occur when the F4-select brood fish, which have no inbreeding, spawn and produce the F4-select generation. Once inbreeding cannot be prevented, it can be minimized if pedigrees are followed, full-sib matings are prevented, and half-sib matings are minimized.
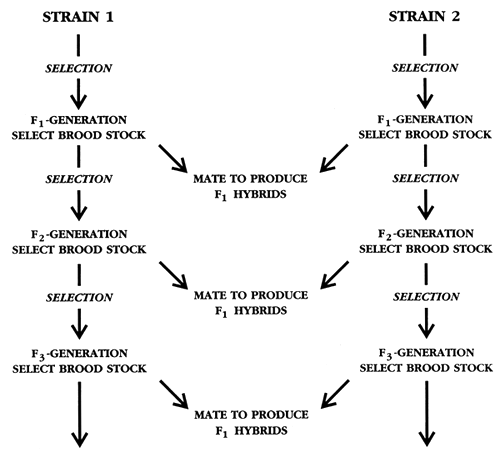
Figure 37. Schematic diagram of how hybridization can be combined with selection to produce fish that have F = 0% for grow-out. The F1 hybrids that are continually produced in this breeding programme will always have F = 0% if the hybrids are not allowed to mate and if the two strains are kept genetically pure. This breeding programme enables a breeder to combine improvements from selection and hybridization. Since F1 hybrids cannot breed true, this type of breeding programme is used to prevent farmers from using the animals or seed for brood stock, and it also ensures that customers must purchase new animals or seed every year.
By combining selection and hybridization, a farmer can produce fish that are genetically superior (as a result of selection) and that exhibit hybrid vigour (as a result of crossbreeding), and as a bonus, he can advertise the fact that the fish have no inbreeding. This combined breeding programme also enables a breeder to protect his investment. By selling hybrid fingerlings, a fingerling farmer sells fish that will not breed true. Selling hybrids to farmers is a traditional way breeding companies protect their investment-farmers have to buy new seed or animals every year.
This type of breeding programme does not guarantee that inbreeding will not cause problems. Inbreeding will occur in both strains. If inbreeding reaches certain levels, it could effect fecundity, which will make the production of hybrid fingerlings more difficult and expensive. One way to prevent inbreeding-induced decreases in fecundity from affecting fingerling production is to select simultaneously in four strains and produce di-hybrids; the breeding programme needed to do this is similar to that illustrated in Figure 22 (page 55). This type of breeding programme is twice as expensive as that needed to produce F1 hybrids, but this is how seed companies circumvent the problem of lowered fecundity in their select but highly inbred lines of corn (maize) and other plants.
It is often difficult to determine inbreeding in a select population when the fish are not marked. When it is difficult to measure inbreeding, the select population can be examined electrophoretically each generation, and the results can be compared to those which existed before selection and to those from a contemporary control population, in order to determine how much inbreeding has accumulated. This will not help prevent inbreeding, but it will enable a farmer or hatchery manager to measure it.
If inbreeding accumulates to levels that cause significant inbreeding depression, selection can be relaxed for a generation by mating the select fish to fish from another strain. This will temporarily reduce inbreeding to 0%. If the farmer imports only superior fish, this might have little effect on the phenotypes that are under selection.
The second basic type of selection is family selection. There are two types of family selection: between-family selection, where whole families are selected or culled based on family means; within-family selection, where the best fish from each of a number of families are saved. Inbreeding is also inevitable with this type of selection. When between-family selection is used, relatively few families are saved. This retroactively lowers Ne of the previous generation to a very small number. If select brood fish are not marked and are mated at random, then because the select brood fish came from relatively few families, relatives will mate, and the inbreeding that will be produced can become quite large.
The easiest way to prevent inbreeding during between-family selection is to mark the select brood fish and to prevent consanguineous matings. This approach can prevent inbreeding for a number of generations and can minimize it thereafter by preventing brother-sister or half-sib matings.
When the fish cannot be marked, the only way to prevent inbreeding or to minimize it during between-family selection is to treat each of the select families as if it were a cohort and use rotational mating to produce successive generations. After the first generation of selection, between-family selection will choose the best families from each “cohort.” The rotational mating program that is used is similar to that illustrated in Figure 36.
When within-family selection is used, if fish can be marked, it is fairly easy to prevent inbreeding for many generations. This type of selective breeding program can maintain Ne at a constant or at an increasing level. Because this type of selection does not put the population though a bottleneck every generation, it will be possible to prevent consanguineous matings for many generations, as long as a sufficient number of families are entered into this selective breeding programme.
Another way to prevent or minimize inbreeding is to use rotational mating. The mating programme that would be used is similar to that illustrated in Figure 36.
As was the case with individual selection, fish with no inbreeding can be produced for grow-out by hybridizing select brood fish from two select strains.
No universal prescription can be written for every farmer and hatchery manager that will cure or prevent problems associated with unwanted inbreeding or the effects of genetic drift. Recommendations must be customized, and depend on: the size of the farm; the goals the farmer or hatchery manager has; his level of expertise; the type of production management that he uses to raise fish; whether the fish can be marked; the presence or absence of a selective breeding programme; and the number of generations that are to be incorporated in the work plan before genetic trouble occurs.
Subsistence-level farmers who own small farms (<2 ha) do not need to worry about the genetic aspects of productivity. They need to become better managers and learn how to increase yields and profits by proper feeding, fertilization, etc.
Farmers who are more technologically advanced should be encouraged to incorporate some genetic management into brood stock management plans. Most farmers are not going to conduct a selective breeding programme; when this is the case, the only genetic goals will be to conserve some genetic variance and to minimize inbreeding. Since most farmers do not mark their fish, the only way to accomplish these goals is to manage the population's Ne at a predetermined level.
Farmers who own either small farms or medium-sized farms (2 ha), who are good managers, and who raise fish under extensive conditions can manage the genetics of the population. These farmers should accept a high level of genetic risk (minimal genetic management). They should try and keep inbreeding from exceeding 10% and try and produce either 95% or 99% guarantees of keeping alleles whose frequencies are 0.05. Most of these farmers should not worry about genetic drift, but the Ne's that are needed to protect the population from the ravages of genetic drift are about the same as those needed to prevent inbreeding problems. These goals will require Ne's of between 30 and 100 per generation, depending on the number of generations in the work plan; Ne's needed for five generations are only 45 and 61, respectively, so this type of management should be easy to incorporate into yearly work plans.
Farmers who own medium-sized farms and who raise their fish under intensive conditions should try to incorporate less genetic risk (more genetic management) into their brood stock management plan. They should try and prevent inbreeding from exceeding 5% and should try and produce either a 99% guarantee of keeping alleles whose frequencies are 0.05 or a 95% guarantee of keeping alleles whose frequencies are 0.01. Effective breeding numbers needed to achieve these goals range from 45 to 297, depending on the number of generations in the work plan; Ne's needed for five generations are 61 and 229, respectively. Even though these Ne's are larger, it should be possible for many farmers to incorporate them into yearly work plans without incurring additional costs. For example, these recommendations can easily be incorporated by tilapia farmers, because they need dozens to hundreds of brood fish in order to produce the fingerlings that are needed to stock the ponds. It might be more difficult for some carp farmers, but the Ne's that are suggested are not excessively large.
Farmers who own large farms (>2 ha) have a broad range of options. Those who use extensive farming methods should accept more genetic risk than those who use intensive farming technology or those who produce fingerlings for the local fish farming industry. Those who use extensive farming methods should try and keep inbreeding from exceeding 10% and should try and produce either 95% and 99% guarantees of keeping alleles whose frequencies are 0.05. This will require Ne's that range from 30 to 250, depending on the number of generations in the work plan; Ne's needed for five generations are only 45 and 61, respectively.
Farmers who use intensive farming or those who are fingerling producers should try to prevent inbreeding from exceeding 5% and should try and produce either 95% or 99% guarantees of keeping alleles whose frequencies are 0.01. Effective breeding numbers needed to achieve these goals range from 230 to 500, depending on the number of generations in the work plan; Ne's needed for five generations are 229 and 309, respectively.
Although these goals require larger Ne's, most farmers should be able to incorporate them into brood stock management plans because of the number of brood fish needed to produce the required fingerlings for stocking or sale.
Hatchery managers who operate public hatcheries designed to produce fish that will be stocked in lakes and rivers culture populations that require considerable genetic management. In fact, managing the population's gene pool so that there are few changes should be the top management goal, and only long-term planning should be incorporated; the minimum should be 25 generations, but if this is not possible, 10 generations should be used. In this context, considerable genetic risk is defined as trying to keep inbreeding from exceeding 5% and trying to produce a 99% guarantee of keeping alleles whose frequencies are 0.01. Effective breeding numbers needed for these goals range from 344 to 1,000, depending on the number of generations in the work plan. Moderate risk is defined as trying to prevent inbreeding from exceeding 5% and trying to produce a 99% guarantee of keeping alleles whose frequencies are 0.005. These goals will require Ne's that range from 689 to 1,000, depending on the number of generations in the work plan. Little risk is defined as preventing inbreeding from exceeding 1% and trying to produce a 99% guarantee of keeping alleles whose frequencies are 0.005. These goals will require Ne's that range from 689 to 5,000, depending on the number of generations in the work plan. In terms of genetic conservation, the best management option is to accept no genetic risk-to try and prevent inbreeding from exceeding 1%, and to try and produce a 99% guarantee of keeping alleles whose frequencies are 0.001. These goals will require Ne's that range from 3,450 to 5,000.
Although the “no risk” option is most desirable in terms of genetic management, it is probably an unrealistic goal at most hatcheries. The best options are compromises between what is best (the no risk option) and what is achievable in terms of the budget, labour, and facilities. Because of this, the low risk and little risk options are those that should be incorporated; Ne's needed to achieve these goals for 25 generations are 780 and 1,250, respectively.
Even though the Ne's presented in Tables 8, 9, and 10 can be used by most farmers and hatchery managers, some will want to customize Ne for their farms or hatcheries and for their budgets and will want to use a combination of genetic goals not listed in these tables. This can be accomplished by using the Ne's presented in Table 4 and 5.
A number of publications that have produced a single recommended Ne have suggested using either 500 or 1,000. The logic behind these values is that many population geneticists feel that when Ne is 500–1,000, the population behaves as if it were infinitely large. The values presented in Tables 9 and 10 show that a constant Ne of 500 will prevent inbreeding- and genetic drift-related problems in farmed populations and that an Ne of 1,000 can do a fairly effective job of conserving genetic variance and minimizing inbreeding in populations that are used for stocking programmes. However, the values presented in Tables 8, 9, and 10 also show that: an Ne of 500 is excessively large for most farmers, while an Ne of 1,000 might be too small for some hatchery managers. Although it might be easier to conduct an extension programme by recommending a single Ne that should be maintained by all farmers, Ne's should be customized, because many farmers can maintain an Ne as small as 45. If an excessively large Ne is recommended, many farmers will simply ignore the advice.
When managing a population's Ne, the most important aspect is to maintain a constant Ne and to prevent a bottleneck. A bottleneck will reduce the Ne of the population and will make it difficult, if not impossible, to achieve genetic goals. The time when most bottlenecks occur is during acquisition of the population. It is difficult to ship large numbers of fish, so there is a tendency to ship fish that have come from a handful of spawns; furthermore, mortalities that occur during and after transportation can drastically lower Ne. If Ne goes though a bottleneck during acquisition of a population, the population may be crippled genetically, and efforts to properly manage it are of little value.
Inbreeding and genetic drift are of great concern when a farmer or hatchery manager wants to conserve the population's genetic and genotypic variance and does not want to alter the population in any way, including via selection. However, when a farmer wants to improve growth or other phenotypes by using a selective breeding programme, the problems associated with inbreeding and genetic drift become less important. Selection produces inbreeding and alters gene frequencies.
Inbreeding will increase in a select population because each act of selection creates a bottleneck and because when the best are mated, the mating of relatives often occurs. Farmers who conduct individual selection can prevent inbreeding from counteracting selection by marking fish and preventing consanguineous matings. If consanguineous matings cannot be prevented, the accumulation of inbreeding can be lessened by preventing brother-sister or half-sib matings. If consanguineous matings are restricted to fish less related than first cousins, inbreeding will rarely be a problem.
If fish cannot be marked, inbreeding will occur. Minimum Ne's can be maintained to try and prevent inbreeding, but inbreeding will occur because each act of selection is a bottleneck. Inbreeding can be prevented by breaking the population into cohorts, conducting selection independently in each cohort, and mating select brood fish from the cohorts by rotational mating.
If family selection is used, inbreeding can be prevented by marking fish and preventing consanguineous matings. If fish are not marked, rotational mating can be used to prevent or minimize inbreeding.
If inbreeding depression become significant in a select line, selection can be relaxed for a generation by mating the select brood fish with outstanding fish from an unrelated strain.
Finally, a farmer can produce fish with no inbreeding for grow-out by hybridizing two unrelated select strains; this not only produces fish with F = 0% for grow-out, but it prevents customers from using the hybrids as brood fish, because F1 hybrids do not breed true.
![]()
![]()
![]()