![]()
![]()
![]()
Introducción
En biología pesquera la forma más práctica de expresar el declinar (o sea la disminución en número) de un grupo de edad de organismos marinos a través del tiempo, es por medio de tasas instantáneas. Dichas tasas, de las cuales normalmente tres han sido definidas, se presentan en las dos expresiones siguientes:
Nt = No · e-Zt | (18) |
donde:
No es el número (inicial) de peces cuando t = 0
Nt es el número de peces que quedan al final del tiempo “t”
Z es la tasa instantánea de mortalidad total
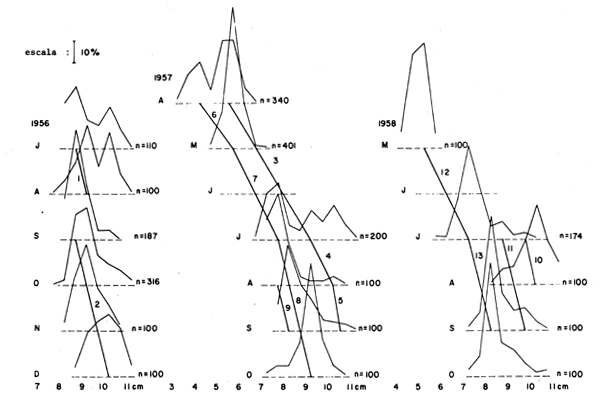
Figura 10 Datos de frecuencia por clases de longitud del “slipmouth” Leiognathus bindus capturado aguas afuera de Calicut, India, en los años 1956, 1957 y 1958, con segmentos de la curva de crecimiento sugeridos para su utilización con el “Ploteo de Gulland y Holt” (ver Tabla 6 y texto). Basado en datos de Balan, 1967
Una ventaja de las tasas instantáneas consiste en que las mismas pueden ser sumadas o restadas. En consecuencia tenemos:
Z = M + F | (19) |
donde:
M es la tasa instantánea de mortalidad natural
F es la tasa instantánea de mortalidad por pesca (ver Figura 11)
Obviamente, cuando:
F = O entonces Z = M; | (20) |
lo cual significa que las mortalidades natural y total tienen el mismo valor cuando no hay pesquería (es decir, en un recurso no explotado).
En lo concerniente al estudio de mortalidad de organismos acuáticos, el biólogo pesquero debe encarar dos tareas principales:
(a) estimar el valor de Z;
(b) cuando ello corresponda, discriminar el valor estimado de Z en sus elementos componentes, M y F.
Estimación de la mortalidad total
Para estimar la mortalidad total(Z) a partir de la talla media en las capturas existen tres tipos de procedimientos:
(a) Cuando se ha obtenido un gran número de datos de frecuencia por clases de longitud
de un recurso dado, con un cierto arte de pesca, Z puede ser estimado a partir
de la longitud media (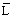 ) en la captura de una población específica mediante:
) en la captura de una población específica mediante:
 donde L∞ y K son parámetros de la ecuación de von Bertalanffy; es la longitud
media en la captura, estimada a partir de tallas mayores de L'; y L' es la longitud
de los animales más pequeños que están plenamente representados en los
muestreos de las capturas (Beverton y Holt, 1956).
donde L∞ y K son parámetros de la ecuación de von Bertalanffy; es la longitud
media en la captura, estimada a partir de tallas mayores de L'; y L' es la longitud
de los animales más pequeños que están plenamente representados en los
muestreos de las capturas (Beverton y Holt, 1956).
(b) Otra ecuación que puede ser utilizada para estimar Z a partir de la talla media
en las capturas es:
 donde:
donde:
L∞, , L' y K ya han sido definidos anteriormente, en tanto que “n” es el número
de peces utilizados para el cálculo de (basados en Ssentongo y Larkin, 1973).
Anotar que cuando n es grande, el término n/n+1 tiende a uno y por lo tanto puede
ser ignorado1.
1 Ver nota al pie de la página 26.
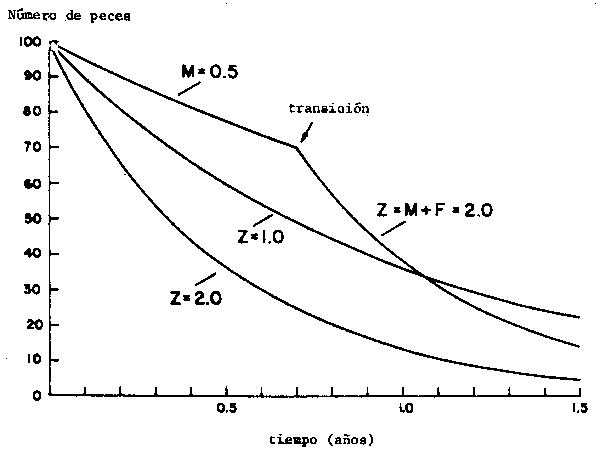
Figura 11 Mostrando la declinación en número de una clase anual de peces, a lo largo del tiempo, comenzando con 100 ejemplares de edad cero, con tres niveles de mortalidad
Figura 12 Nomograma para el estimado del factor de seleción de peces a partir de mediciones de sus proporciones corporales. Información basada en Shindo et al., 1979 y Meemeskul (1979). Original (Nota: todos los factores de slección estimados por medio del presente nomograma están referidos a longitud total. En consecuencia: SF · dimensión de malla = Lc (longitud total))
| Año | Esfuerzo | n | | Z1 | Z2 |
|---|---|---|---|---|---|
| 1966 | 2,08 | 4 733 | 13,25 | 2,41 | 2,95 |
| 1967 | 2,08 | 11 902 | 13,01 | 2,69 | 3,24 |
| 1968 | 3,50 | 12 503 | 12,99 | 2,72 | 3,27 |
| 1969 | 3,60 | 9 060 | 13,07 | 2,62 | 3,16 |
| 1970 | 3,80 | 8 132 | 12,37 | 3,73 | 4,29 |
| 1972 | 7,19 | 3 635 | 12,30 | 3,88 | 4,44 |
| 1973 | 9,94 | 10510 | 12,01 | 4,61 | 5,14 |
| 1974 | 6,06 | 7 960 | 12,60 | 3,30 | 3,85 |
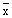 | 4,87 | 12,70 | 3,25 | 3,79 | |
Constantes: L∞ = 20,0; K = 1,16; Lc = 10,0 | |||||
(c) Cuando el crecimiento es isométrico, la expresión basada en peso correspondiente a la Ecuación 22 es la siguiente:

donde W∞ y K son parámetros de la función de crecimiento de von Bertalanffy para crecimiento en peso, la cual tiene la forma:
| Wt = W∞ (1-e-K.(t-to)³) | (24) |
donde  y W' son los pesos correspondientes a y L' que pueden obtenerse (así
como W∞) a partir de la conversión de largo en peso, por medio de la apropiada
relación largo/peso.
y W' son los pesos correspondientes a y L' que pueden obtenerse (así
como W∞) a partir de la conversión de largo en peso, por medio de la apropiada
relación largo/peso.
La Ecuación 24 es particularmente útil, por cuanto resulta bastante fácil pesar la
captura y dividirla entre el número de ejemplares, a fin de obtener el valor de .
Un parámetro estrechamente relacionado con L' (ver la definición anterior) es la longitud promedio a la primera captura (Lc), o la captura a la cual el 50% de los animales muestreados son retenidos por el arte de pesca.
Lc normalmente se estima por experimentos de selectividad (ver Gulland, 1971) los que, sin embargo, requieren de mucho tiempo y esfuerzo. Para facilitar la estimación de Lc se incluye una figura que permite obtener estimados de Lc, a partir de características de los peces que son fáciles de obtener, tales como la relación largo/ancho, o el factor de circunferencia (girth factor) de los peces (Figura 12).
La Figura 12 no permite estimar directamente Lc. Cuando la relación largo/ancho o el factor de circunferencia ha sido estimado, la Figura 12 puede ser empleada para estimar un factor de selección (SF) tal que:
Lc = SF x tamaño de malla
(de la malla del copo cuando se trata de una red de arrastre) | (25) |
El parámetro Lc se usa en una serie de modelos de evaluación de recursos particularmente en el de Beverton y Holt (1966). En casos en que la selección tiene forma de “filo de cuchillo” (ver Beverton y Holt, 1966 o Gulland, 1971), Lc = L'; sin embargo, por lo general L'> Lc.
Estimación de Z por medio de una curva de captura
Otro método de determinar Z consiste en el muestreo de una población de peces integrada por varias clases de edad, seguido del trazado de un gráfico con el valor del logaritmo natural (loge) del número (N) de peces en la muestra, contra su respectiva edad (t) en el eje de las “x”. Es decir:
logeN = a + bt | (26) |
donde el valor de “b”, con signo cambiado, proporciona una estimación de Z.
Sin embargo, es preciso que se cumpla una serie de requisitos para que el valor de “-b” resulte una buena aproximación del de Z. Entre ellos, cabe mencionar:
(a) Sólo deben ser considerados aquellos valores de logeN que correspondan a clases de edad completamente vulnerables frente al arte de pesca de referencia (entre otras cosas, la longitud de los individuos debe ser mayor que Lc, tal como se lo definiera anteriormente). Esto implica utilizar nada más que la “porción descendente” de la curva de captura (ver Figura 13).
(b) El reclutamiento debe haber sido parejo dentro del período considerado; o en todo caso haber tenido cambios al azar únicamente.
Cuando se dispone de apropiadas muestras de frecuencia por clases de longitud o de peso, una curva de captura puede también ser confeccionada a través de la conversión previa de tamaño a edad en base a los respectivos parámetros de crecimiento. En tal caso, sin embargo, debe tomarse la precaución de no incluir ejemplares cuya talla se halla próxima al valor asintótico de longitud, puesto que ello podría implicar que su edad fuera groseramente sobrestimada. Esta última circunstancia, precisamente, determina la imperiosa necesidad de confeccionar un diagrama de puntos dispersos a partir de pares de valores de logeN y de t, a los fines de poder identificar adecuadamente la sección de la curva de captura que puede ser utilizada para el cálculo de Z (Figura 13). Asimismo, debe destacarse que, en tanto Z es igual a la pendiente (con signo cambiado) de la curva de captura, la edad real - que requiere un estimado de to - puede en este caso reemplazarse por una edad relativa; por ejemplo haciendo to = 0 (Tabla 8, Figura 13).
| Longitud media de la claseb | N | logeN | Edad relativac | Advertencias |
|---|---|---|---|---|
| 6,5 | 3 | 1,10 | 0,56 |  No debe utilizarse la parte ascendente de la curva No debe utilizarse la parte ascendente de la curva |
| 7,5 | 143 | 4,96 | 0,67 | |
| 8,5 | 271 | 5,60 | 0,78 | |
| 9,5 | 318 | 5,76 | 0,90 | |
| 10,5 | 416 | 6,03 | 1,03 | |
| 11,5 | 488 | 6,19 | 1,17 | |
| 12,5 | 614 | 6,42 | 1,32 | |
| 13,5 | 613 | 6,42 | 1,49 | Porción utilizada para la estimación de Z |
| 14,5 | 493 | 6,20 | 1,67 | |
| 15,5 | 278 | 5,63 | 1,88 | |
| 16,5 | 93 | 4,53 | 2,12 | |
| 17,5 | 73 | 4,29 | 2,40 | |
| 18,5 | 7 | 1,95 | 2,74 | |
| 19,5 | 2 | 0,69 | 3,15 | |
| 20,5 | 2 | 0,69 | 3,70 |  No debe utilizarse pues las longitudes se encuentran de masiado próximas a L∞ No debe utilizarse pues las longitudes se encuentran de masiado próximas a L∞ |
| 21,5 | 0 | - | 4,53 | |
| 22,5 | 1 | 0,00 | 6,19 | |
| 23,5d | 1 | 0,00 | - |
a Según Ziegler (1979) quien asimismo proporciona: L∞ = 23,1; K = 0,59 y T = 28°C
b Longitud = longitud total, en cm
c Calculada mediante la Ecuación 32, con to = 0
Discriminación de Z en M y F
Dividiendo Z en M y F por medio del trazado de un gráfico de Z contra esfuerzo
Cuando, para un período de varios años, se dispone de valores de Z correspondientes a diferentes magnitudes anuales de esfuerzo (f), el valor de M puede ser calculado a partir de:
Z = M + qf | (27) |
donde “q” es el coeficiente de capturabilidad que relaciona “f” y mortalidad por pesca (F) según:
F = q · f | (28) |
En consecuencia, una serie de valores de Z (media anual) pueden ser graficados contra sus respectivos valores de “f”, y mediante la técnica de regresión lineal cabe trazar una línea recta ajustada a dichos puntos, la cual responderá a una ecuación del tipo:
y = a + bx | (29) |
donde:
Z = y
x = f
b = pendiente, que proporciona un estimado del coeficiente de capturabilidad “q”
a = intercepto, corresponde a un estimado de M
(ver Figura 14, donde se han graficado los valores de Z obtenidos en base a las Ecuaciones 21 y 231).
Dividiendo Z en M y F por medio de un estimado independiente de M
Cuando se dispone de sólo un valor de Z, o cuando los valores estimados de Z y “f” cubren un rango demasiado pequeño como para que resulte posible obtener valores razonables de M y de “q”, el coeficiente de capturabilidad (q) pudiera ser estimado de acuerdo a:

donde:
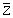 es la media de los valores disponibles de Z (o un único valor de la misma)
es la media de los valores disponibles de Z (o un único valor de la misma)
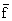 es la media de los valores de “f” (o de un sólo valor de “f”)
es la media de los valores de “f” (o de un sólo valor de “f”)
M es un estimado independiente de la mortalidad natural (ver Tabla 7 y Figura 14).
Método para obtener estimados independientes de M
Ha sido demostrado por varios autores que el valor del parámetro K de la VBGF en peces se encuentra estrechamente vinculado con su longevidad. Esto se fundamenta sobre la observación según la cual, en la naturaleza, los peces más viejos de un recurso crecen hasta alcanzar un 95% de su longitud asintótica (Taylor, 1962; Beverton, 1963). A partir de:
Lt = L∞ (1-e-K(t-to)) | (31) |
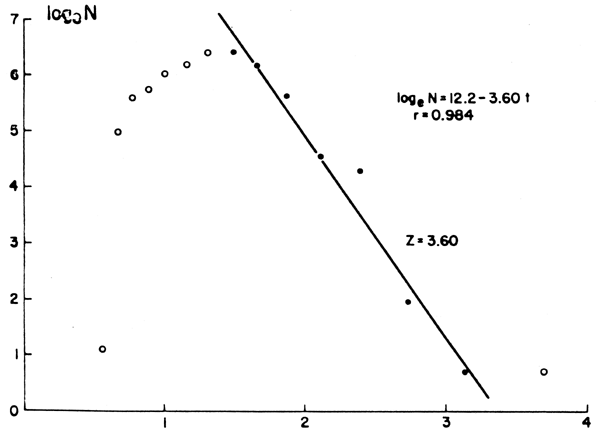
Figura 13 Curva de captura basada en longitudes convertidas a edades. Nótese que la parte “ascendente” de la curva no ha sido considerada, así como tampoco un punto correspondiente a una longitud próxima a L∞ (ver texto)
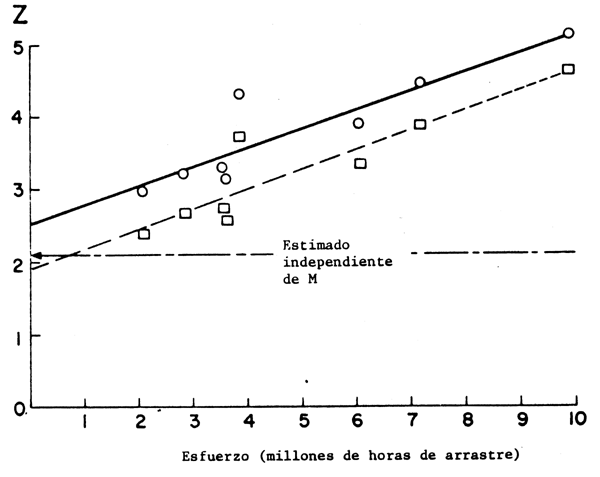
Figura 14 Gráfico de la mortalidad total (Z) contra el esfuerzo de pesca (f) a los fines
de estimar la mortalidad natural y el coeficiente de capturabilidad en Salaroides
leptolepis del Golfo de Tailandia. Basado en la Tabla 7. Las ecuaciones son:
Z1 = 1,96 + 0,263 f (cuadrados)
Z2 = 2,52 + 0,263 f (círculos)
podemos obtener:

Si, para los peces más viejos, asumimos una longitud del 95% de L∞, resultará que:

y en consecuencia:

donde “tmax” es la longevidad de la especie en cuestión.
Resulta evidente que, en los peces, la mortalidad natural tendría que estar correlacionada con la longevidad, y por lo tanto con K. Asimismo, el valor de M debiera estar también determinado por el tamaño, puesto que los individuos mayores han de tener, en general, menor número de predatores que los pequeños.
Puede demostrarse, por otra parte, que la mortalidad natural en peces se encuentra vinculada con la temperatura del medio ambiente (Pauly, 1978a, 1980). Estas variadas interrelaciones pueden ser expresadas, con respecto al crecimiento en longitud, por medio de la regresión múltiple:
log10M = -0,0066-0,279 log10L∞ + 0,6543 log10K + 0,4634 log10T | (35) |
y con respecto al crecimiento en peso, por:
log10M = -0,2107-0,0824 log10W∞ + 0,6757 log10K + 0,4687 log10T | (36) |
donde:
M es la mortalidad natural de un cierto recurso;
L∞ (longitud total en cm) y W∞ (peso vivo en g) son las dimensiones asintóticas de la especie;
K es un coeficiente de crecimiento;
T es la temperatura media anual (en °C) del agua en la cual vive el recurso de referencia,
obtenida por ejemplo a partir de un atlas oceanográfico.
Ambas regresiones han sido deducidas en base a una serie de 175 valores de L∞ (o de W∞), K, T y M, como resultado de una búsqueda de literatura pertinente (Pauly, 1980); y aplicada, de esta manera, a rangos de temperatura desde 5°C a 30°C. Las fórmulas proporcionan adecuados estimados en M en peces, que en caso de ser posible por supuesto debieran ser confirmados por medio de análisis de la particular especie de referencia.
Aunque las Ecuaciones 35 y 36 generalmente proporcionan razonables estimados de M para casi cualquier serie de valores de los parámetros de crecimiento y de temperatura, existe un grupo de peces tropicales en los cuales dicha magnitud pudiera encontrarse sesgada. Así, en el caso de peces pelágicos fuertemente encardumados - especialmente en los clupeidos - la mortalidad natural está generalmente sobrestimada por tales ecuaciones. En consecuencia, podría ser conveniente reducir en alguna forma el cálculo de M\?\, por ejemplo multiplicándolo por 0,8.
Evaluando el estado de un recurso por medio de sus tasas de mortalidad únicamente
Una vez que se dispone de los valores de M y de F, puede calcularse la tasa de explotación (E) mediante:

lo cual permite una grosera determinación acerca de si el recurso es objeto de sobrepesca o no, en la asunción de que el valor óptimo de E (Eopt) es aproximadamente igual a 0,5. La utilización de E ≈ 0,5 como valor óptimo para la tasa de explotación se basa en el acerto según el cual el rendimiento sostenible se encuentra optimizado cuando F ≈ M (Gulland, 1971a).
| Ejercicio: | (a) | Determine la disminución en número de una clase anual de peces (No = 100 000), con M = 0,5 y F = 1,0, cuya explotación comienza a la edad de 0,5 años. |
| (b) | Calcule los valores anuales de Z a partir de la Tabla 9 y en base a las Ecuaciones 21 y 23, y utilice estos estimados para obtener valores de M y “q” por medio de la Ecuación 27. | |
| (c) | Compare el valor de M obtenido en (b) con el valor de M resultante de la Ecuación 35, si se sabe que L∞ =29, K = 1,2 y T = 28°C. | |
| (d) | Utilice los datos de la Tabla 10 para trazar una curva de captura en base a la Ecuación 32, con to = 0, a fin de convertir los valores de longitud media en estimados de la edad relativa; y determine Z. Estime M mediante la Ecuación 35 y los adecuados parámetros de crecimiento. Calcule F y E, y determine si el recurso de referencia se encuentra en su etapa óptima de explotación o no. |
| Año | Esfuerzob | c | n |
|---|---|---|---|
| 1966 | 2,08 | 15,7 | 12 370 |
| 1967 | 2,80 | 15,5 | 14 231 |
| 1968 | 3,50 | 16,1 | 10 956 |
| 1969 | 3,60 | 14,9 | 9 738 |
| 1970 | 3,80 | 14,4 | 12 631 |
| 1973 | 9,94 | 12,8 | 9 091 |
| 1974 | 6,06 | 12,8 | 15 229 |
a Información basada en Boonyubul y Hongskul (1978) y SCS (1978)
b En millones de horas de arrastre
c A ser utilizada junto con L∞ = 29, K = 1,2 y Lc = 7,6
| Longitud mediab | N | Longitud mediab | N | |
|---|---|---|---|---|
| 7,5 | 11 | 17,5 | 428 | |
| 8,5 | 69 | 18,5 | 338 | |
| 9,5 | 187 | 19,5 | 184 | |
| 10,5 | 133 | 20,5 | 73 | |
| 11,5 | 114 | 21,5 | 37 | |
| 12,5 | 261 | 22,5 | 21 | |
| 13,5 | 386 | 23,5 | 19 | |
| 14,5 | 445 | 24,5 | 8 | |
| 15,5 | 535 | 25,5 | No debe utilizarse | 7 |
| 16,5 | 407 | 26,5 | 2 |
a Basado en Ziegler (1979)
b Para ser usado junto con L∞ = 29,2, K = 0,607 y T = 28°C;
longitud = longitud total en cm
![]()
![]()
![]()