![]()
![]()
![]()
by
A. Bakun (Chairman)*
| ABSTRACT |
| The main primary variables or conditions which directly affect individual fish and, through these, the abundance of fish populations are: food, predation, temperature, salinity and oxygen concentration. These primary variables are altered by several processes, some of which are described in this report, together with different sources of information on environmental variability and biological characteristics. There is a need for more rigorous approaches to the problem of environmental effects on fishery problems, and some suggestions are made in this respect. The information on environmental variations is of great value for management purposes, particularly when environment has proven to be a major source of fluctuation of fish availability. |
Fishermen are very aware that short-term availability of fish is often dependent on environmental conditions such as temperature of the water or state of the tide. In the longer term there are equally important relationships with environmental changes, but the linkages tend to be less clear. The requirement of fishermen and fishery managers is that these relationships be identified so as to enable prediction of catches and of their effect on the abundance and viability of fish resources. For the fishery scientist the problem is a more fundamental one of differentiating between availability and absolute changes in abundance; and of understanding the processes by which various environmentally-dependent rates, such as recruitment, growth, natural mortality and fecundity interact with exploitation to cause negative consequences such as stock dislocation or collapse. The results of the discussions of these and other topics related to the study and monitoring of the environment are summarised in this report. Those who contributed to the discussions and preparation of this report are S. Coombs and F. Robles, who acted as rapporteurs, as well as P. Bernal, J. Cañon, P. Fréon, M.L. Garcia, O. Guillén, R. Jones, R. Jordán, P.M. Kelly, P. Lagos, R. Parrish, G.D. Sharp, P. Shelton, S. Zuta and others.
Lack of understanding of the processes which regulate recruitment, in particular, has been identified as perhaps the most serious problem hindering the advance of fishery science. Studies of historical data bases seem to have been remarkedly unsuccessful in finding clear relationships. One argument has been that one would not expect simple or constant relationships. Some relationships that have been found are undoubtedly mere chance agreements with no basis in reality; with a multitude of conceivable environmental effects and relatively short fishery time series, some degree of empirical “fit” to one or another environmental series can generally be found. Clearly, there is a present requirement for a more rigorous approach to the problem, within a firm conceptual framework which will enable examination to proceed in a more scientific fashion. Rational approaches, which are based on understanding of the processes involved, and empirical approaches which depend on analysis of available data series, should be merged so as to incorporate all the available pertinent information. Design of observational programmes, monitoring systems, and experiments should be integrated so that all elements are mutually supportive. Any possible “shortcuts”, such as interregional comparative studies or other types of approach should be evaluated and used where beneficial.
Above all, a focus must be kept upon overall integration of the various levels of knowledge developed so as to bear directly on real-world needs. For example, we are presently experiencing perhaps the greatest large-scale ocean anomaly in recent decades, the El Niño of 1983. Despite the wealth of theory and data on effects of such anomalies on, for example, productivity, it is an unfortunate fact that at this time there exists no generally-agreed-upon basis for predicting its effect on recruitment and viability of one of the largest fishery stock complexes in the world, the Peruvian-Chilean sardine, Sardinops sagax. On the other hand, accumulated experience and available scientific knowledge are sufficient to indicate increased likelihood of serious abnormalities in such factors as reproductive success, availability to exploitation, etc., during periods of intense environmental anomalies.
The larval stage is often considered to be the most likely segment of the life cycle for environmental effects to strongly impact eventual recruitment. The most promising hypothetical mechanisms are discussed in several recent IOC or FAO reports (e.g. Sharp, 1981; Bakun et al., 1982; SCOR Working Group 67, 1982); thus we have chosen not to repeat any lengthy enumeration here. Also for most of the topics discussed here reference should be made to the Proceedings of this Expert Consultation. Many of the papers presented and discussed at this meeting provide an adequate background to this report.
Considering the interactions within a defined ecosystem, there will be a different emphasis on the physical and biological forcing functions depending on the nature of the system and the implicit time and space scales of the processes involved. For example, consider a simple contrast between a temperate-shelf sea (e.g. North Sea) and a boundary-current (e.g. Peru Current) system (Table 1). By implication there will be greater changes in hydrography, productivity and geography of the biota in the boundary system than in the temperate-shelf sea system.
Table 1. Some contrasting characteristics of temperate-shelf seas and boundary-current systems
| Temperate-shelf sea | Boundary Current |
| Relatively-closed system | Relatively-open system |
| Slow biological turnover | Rapid biological turnover |
| Locally driven | Affected by distant physical events |
| Physically and biologically buffered against change | Weakly buffered against change |
Furthermore an examination of the lifestyle of the fish species will suggest the relative importance of links with other components of its system. An oversimplified example is presented in Figure 1.
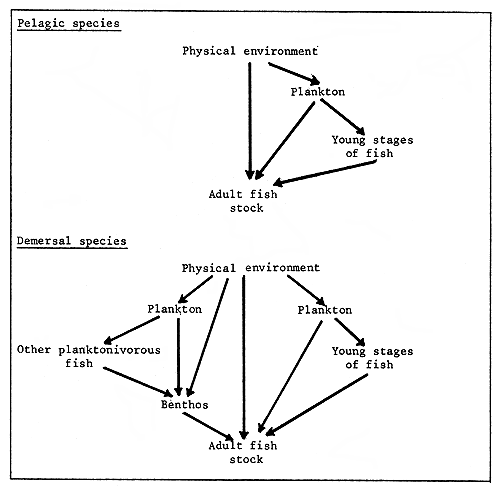
Figure 1. Simplified links of pelagic and demersal species with other components of their systems
Eastern-boundary currents are dominated by pelagic fishes and much of the exploited biomass is composed of species at relatively low trophic levels. Temperate-sea shelf systems tend to be dominated by demersal species; however, they also have considerable quantities of pelagic species. The demersal species typically are at the higher trophic levels and they tend to have complex food webs which interact with, and include, the food webs of the pelagic species. The more direct links between the physical environment and the adult fish stock for a pelagic species of eastern boundary currents suggest that there is less buffering between the response of the fish to environmental change and therefore the effects may be more readily observable.
Clearly a wide range of processes whereby environmental change affects fish populations can occur in most systems. The important consideration is that an appreciation of the relative influence of the system-specific characteristics will enable some assessment to be made of the relative importance of the various mechanisms. Obviously, collecting data sufficient to account for all environmentally-caused variance will not be possible. Focus must be directed toward a single process, or at most a very limited number of processes, having a large enough impact to be discernible through the noise level generated by the multitude of other processes which are occurring.
The range of scales to be considered is wide. For example, the geographical scales of populations can range from hundreds of metres in coral reefs to thousands of kilometres in boundary currents; individual environment scales can range from centimetres (e.g. for larval feeding) to thousands of kilometres (e.g. ambit of an adult fish); relevant time scales can range from one day (for larval starvation) to a couple of years (generation time of populations).
Certainly, it is essential to have a sound conceptual basis upon which to organise any study. The alternative is to expend resources collecting various pieces of information which may never add up to the answers required for necessary actions. Progress is expected to occur as a result of an iterative feed-back process, more or less as follows:
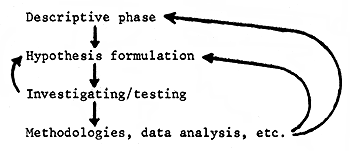
From the formalisation of a hypothesis concerning availability of fish, a logical appraisal should be undertaken following a scheme generally similar to that outlined in Figure 2, where particular research pathways are identified.
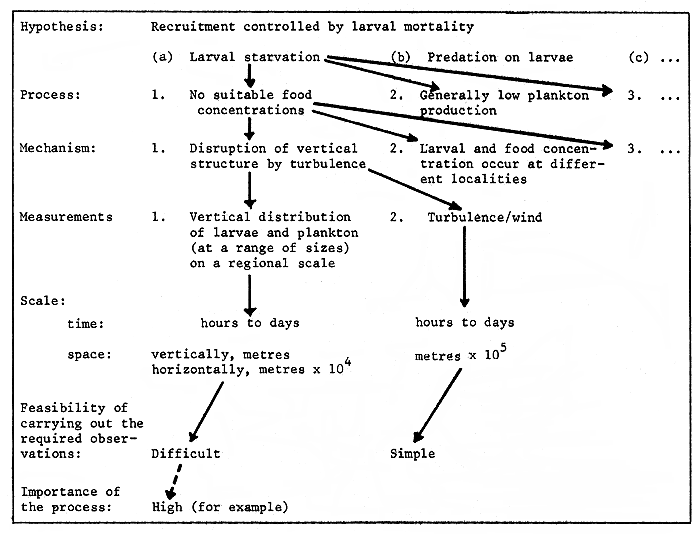
Figure 2. Example of pathways for the appraisal of biological and physical measurements that need to be made when testing the hypothesis that recruitment is controlled by larval mortality.
From this type of analysis we can determine the biological and physical measurements which are to be made. The resulting benefits are efficiency of sampling and improved interpretation of results. Equally, the importance of each process or mechanism can be assessed from a consideration of the system-specific attributes and the feasibility of carrying out the required investigations can be judged; by this means some priority can be assigned to the investigation on a cost/benefit basis.
Complementary to the formalised-hypothesis-testing approach there is still a need for a monitoring function to build up a data base on potentially useful variables. Ease of measurement is of prime importance to ensure continuity in data collection.
In order for a stock to achieve and maintain a large population, a number of physical, chemical and biological conditions must be present on the proper time and space scales. Conditions which directly affect individual fish (i.e. food, predation, temperature, salinity, and oxygen concentration) we are calling “primary variables”. In addition to being affected directly by the primary variables, fish are affected indirectly by processes which alter the primary variables (i.e. insolation, turbulent mixing, transport, divergence and convergence). For development and growth to occur food of the right nutritional quality and particle size must exist at sufficient concentrations. In addition, temperature, salinity and oxygen conditions must not only be within the physiological limits of the stock but a correct mixture of these factors must exist. For example, food may be adequate for rapid growth at one combination of temperature, salinity and oxygen concentration but increased physiological stress caused by changes in temperature or oxygen concentration could limit growth. This could occur even when all of the factors were individually within the physiological limits of the stock.
Rapid growth during the early development stages is particularly important in the eventual recruitment of a stock as increased body size is an important factor in reducing susceptibility to predation or cannibalism. In this regard it is noteworthy that a great majority of neritic fishes, including demersal and littoral forms, have epipelagic larvae. Mortality rates of epipelagic larvae are known to be quite high; however, predation on demersal or littoral larvae might be expected to be even higher due to the increased concentration of vertebrate and invertebrate predators in the benthic and littoral regions.
The high mortality rates that characterise the egg and larval stages make these stages likely susceptible periods for environmental perturbations to substantially alter recruitment. However, to date the number of larvae, as measured by extensive surveys, has proven to be a better index of parent-stock size than of recruitment. This, of course, does not mean that recruitment is, or is not, established during the early life history; but it does suggest that recruitment may be affected by environmental conditions that relate to the juvenile or adult segments of the life cycle.
There is essentially nothing known, for a surprising number of exploited fishes, from the time the larvae become mobile enough to avoid egg and larvae survey nets (usually about a month after hatching) until they arrive at nursery grounds which are usually located in shallow water. This “larval drift period” usually lasts for about a month to nearly a year depending on the species. Environmentally-induced variation in growth, mortality or the larval drift pattern are potential factors in recruitment regulation during this period. Recruitment, in many stocks, is considered to be established by the time the nursery grounds are occupied.
Environmentally-related changes in the natural mortality rate of post-recruits obviously cannot be excluded as having great potential to affect population size, particularly when there are multiyear trends in population size. In addition the adult segment of the stock may directly alter recruitment through changes in egg production. For example, feeding conditions could alter the factors related to egg production, i.e. growth, condition factor, age at maturity, batch fecundity, number of spawnings. Large-scale anomalies are also known to greatly alter the location, seasonality, and volume of spawning; the anchoveta response to the 1972 El Niño is a good example of this type of alteration.
Primary productivity depends on such factors as insolation, nutrients, temperature, circulation and mixing processes, grazing and sinking of phytoplankton. Major productivity centres tend to be found in regions of vertical mixing and horizontal divergence of the upper layers. Turbulent mixing and sinking may adversely affect the rate of primary productivity by removing organisms from the illuminated surface layer. Variation in intensity of coastal upwelling may cause variations in the supply of nutrients supporting organic production in the upper layers.
The transfer of nutrients to the surface layers is not only dependent on upwelling strength (vertical velocity), it also depends on the nutrient content of the waters at the depths at which the upwelling originates. For example, in the Peru current during El Niño occurrences the nutrient-poor surface waters may extend to greater depths and the upwelling process may not be deep enough to reach high nutrient concentrations.
There is a possibility that changes in species composition in the plankton community may be important in determining availability of suitable food particles. During the 1976 El Niño, for example, dinoflagellates were dominant off Peru where normally diatoms are dominant.
The spatial variation of food is known to be extremely patchy, and micro-scale patchiness is thought to be a significant factor controlling recruitment of fishes. The development and dispersal of physical microstructure is related to various processes including solar insolation, tidal currents and wind-induced turbulent mixing. Insolation alters the micro-structure by decreasing the density of the surface waters, resulting in the development of vertical density gradients. On the continental shelf and other shallow bank areas, turbulence generated by tidal flows can destroy vertical stratification, resulting in frontal formations between stratified and mixed zones. Wind-induced turbulent mixing, which is proportional to the cube of the wind speed alters patchiness by dispersing vertically stratified physical and biological microstructure, by bringing nutrients into the upper mixed layers. and by altering the salinity, temperature and oxygen concentration in the surface layer.
The epipelagic phase of neritic fishes varies from only the early life history phase in many demersal and littoral fishes, to the whole life cycle in the case of many small pelagic fishes. The epipelagic zone generally contains appropriate food for fish larvae; it is also the area where transport is the largest. Thus, in addition to the problems of securing sufficient food for early growth and of avoiding predation, larvae must also be located in a geographical position which favours larval transport to nursery areas where a new combination of food, physical, and chemical conditions may be required for juvenile growth.
Transport in the ocean may involve one or several flow mechanisms. The large-scale geostrophic currents are related to pressure gradients which can be estimated from the ocean temperature and salinity structure and which may be associated with sea-level variations at coastal stations. Surface Ekman transport is directly driven by the local wind; upwelling regions are characterised by offshore-directed Ekman transport which may affect recruitment by offshore dispersion of larvae. Oscillating tidal flows can result in net transport over a number of cycles, particularly if organisms can adjust their depth level at different phases of a cycle or even cling to the ocean bottom during unfavourable flow phases. Vertical shear flows of various types are common in the ocean; vertical migratory behaviour may be adapted to utilise vertical variations in horizontal flow. Various surface skin effects, Langmuir circulations, nonlinear surface wave transports, etc. might also be involved in crucial life-cycle transport processes.
Anomalies in atmospheric circulation strongly influence the ocean. There are indications that the upper ocean and the atmosphere are closely coupled over extensive areas from short- to long-term climatic time-scales. The air-sea coupling is accomplished by atmospheric forcing in the ocean. The effects of these interactions appear in the ocean's thermal, velocity and salinity fields, which are affected by the winds and the temperature in the lower atmosphere.
El Niño is a good example of large-scale air-sea interaction and primary variable change by environmental conditions. Anomalous behaviour of the southeast trade winds in the central equatorial Pacific generates a large-scale perturbation in the upper ocean in the form of equatorial Kelvin wave, which propagates eastward, and deepens the thermocline along with the intrusion of equatorial surface waters of higher temperature and low salinity toward the west coast of South America. The effect of the anomalous water and deep thermocline is to produce large fluctuations in all of the primary variables over a very large proportion of the Peru current region.
Some of the most common means of acquiring information and monitoring environmental variability are:
Maritime observations. As part of the global weather reporting network, commercial ships generally record sea-temperature, surface-wind, and other atmospheric measures. These observations provide basic data on the state of the marine environment. The cost to fishery programmes is low because their use for fisheries is a byproduct of meteorological programmes. In fact, the incorporation of these observations into large-scale analysed fields by the meteorological agencies offer a resource for fishery applications such as indices of wind-driven Ekman transport, wind-generated turbulent mixing, and locally wind-produced upwelling.
Shore stations. Observations taken regularly at coastal installations offer valuable and cost-effective means of monitoring the coastal marine environment. Sea-level measurements can be a particularly useful data resource, providing signals of important ocean processes including alongshore flow, water structure, and propagating ocean disturbances such as appear to be involved in El Niño-type anomalies. Longer-period sea-level fluctuations are samples of a fairly large spatial scale, which is an advantage in interpretation. Sea temperature measurements are also very cost-effective signals of ocean variability, but may sample a more local spatial scale. Coastal sampling of other ocean properties, such as salinity or dissolved oxygen (at depths below the surface), may offer very favourable cost-benefit ratios under certain circumstances. Measurements of wind at exposed coastal locations are also valuable data resources.
Ships of opportunity. It is sometimes possible to avoid some of the high cost of dedicated scientific vessels by utilising commercial ships in collection of specialised data. For example, expendable bathythermograph (XBT) systems allow a measurement of sub-surface temperature structure without impeding the travel of the ship in any way. XBT observations are collected and disseminated world-wide by the IGOSS programme of the Inter-governmental Oceanographic Commission.
Research vessels. Dedicated ships represent a rather high-cost mode of data collection. However, they may offer the only means to obtain high-quality data on the time- and space-scales required. Hydrographic surveys yield the quasi-synoptic geostrophic current pattern and three-dimensional coverage of ocean temperature, salinity, oxygen and density structure. Towed sensor systems such as undulators or batfish offer a means to measure short-scale vertical or horizontal pattern. A variety of special purpose measurements may be practical only from a dedicated vessel.
Aircraft. Wide areal coverage in a nearly synoptic time-scale can be achieved with instrumented aircraft. Measurements of atmospheric variables such as wind, air, temperature, and humidity as a function of altitude can be obtained. Techniques are available for remote sensing of SST (sea-surface “skin” temperature) and areal distribution of near-surface chlorophyll. Subsurface temperature structure can be observed from airborne XBT systems.
Satellites. Remote sensing of SST with infrared and microwave imagery has been developed intensively. The absolute accuracy is not presently adequate for defining long time-scale variability of temperature; however, satellite pictures are useful for detecting surface frontal formations and other spatial patterns. Development of descriptors and parameterisations of satellite-sensed spatial pattern that would illuminate longer time-scale variations in pattern-related processes would be very useful to fishery-environmental studies. Chlorophyll concentration can be sensed with high spatial resolution and is expected to prove useful as the technique is developed. Remote sensing of sea-surface properties is limited by availability of cloud-free skies; however, information on cloud cover may be useful for estimating solar radiation reaching the sea-surface in particular areas, although estimates of precision of indications of longer-scale variations are not presently available. Further development of satellite altimetry may yield important information on variations in sea-surface topography and related geostrophic flow. All remote-sensing applications require ground-truth data for absolute calibration; however, for many studies relative indices are adequate.
Historical techniques. As well as current day sampling of biological systems. Some data can be gained from dendrochronological studies (e.g. tree rings) which give a past history of climatic influence. Direct evidence of the response of fish populations is available from fish scale or otolith deposits in marine sediments.
Biological sampling programmes generally involve substantial resource outlays and therefore are not appropriate at all scales and under all circumstances. However, obtaining insights into the actual interactions of life-cycle events with food organisms, predation, cannibalism, etc., on the scales on which they actually occur, is certainly a crucial requirement for progress on the recruitment problem.
These provide measurements of the standing crop of phytoplankton and zooplankton, usually by means of various indices such as particle counts or chlorophyll a concentration. Because of the inherent generalizations of broad-scale sampling only major patterns will be detected and only simple or clear-cut relationships with recruitment or adult availability will be found. Concurrent measurement of appropriate simple physical and chemical variables (e.g. temperature, salinity, nutrients) should be made as these may provide more easily obtained analogues of the broad-scale biological distributions.
Biological sampling should be designed to properly integrate the patchy distribution of planktonic organisms. Analysis of the samples to species level is often not required but some estimate of zooplankton size structure and feeding mode (herbivorous/carnivorous) is beneficial; such information allows the calculation of predation pressure on larvae and availability of food for juvenile and adult fish. In this context attention should be paid to coelenterate tissue which is generally indicative of predatory organisms.
For the purpose of broad-scale biological sampling, conventional plankton net tows are usually suitable. However, some savings in both ship and analysis time can be made by using continuous plankton filtering or size-particle analysis using undulating towed vehicles. These instruments offer the additional considerable advantage of taking a concurrent range of physical measurements (also chlorphyll a) at a fine spatial scale. In areas of coherent hydrographic structure it may be fruitful to use some estimate of larval morphometric and histological conditions to determine favourable biological and physical conditions for feeding.
Sampling problems are more involved at fine-scales but should receive significant attention in view of the potential importance of the processes taking place. Of particular importance is the vertical stratification of plankton and fish larvae since variability is typically several orders of magnitude greater in the vertical than in the horizontal dimension; thus vertical migratory behaviour of fish larvae may increase their access to favourable food concentrations.
As well as obtaining a finer scale of standing crop measurements fine scale sampling can be integrated with measurements of primary production (e.g., C14 uptake) and some quantification of secondary production (e.g. copepod growth rates). Additionally larval fish experiments can be used as direct quantifications of the suitability of the environment for larval survival.
Fine-scale sampling techniques may involve conventional net systems but depth-stratified systems such as pumped samplers or Longhurst-Hardy Plankton Recorders offer great advantages. Particle counter analysis can provide a rapid means of measuring fine-scale vertical distribution; however, some attention should be paid to species or type composition of the plankton to allow for prey selectivity of fish larvae.
Sampling for eggs and larvae is sometimes routinely carried out for stock estimation. Valuable interpretive information can be obtained if relatively simple concurrent measurements are made of other biological and physical properties. Normally, a representative sample of plankton other than fish eggs and larvae can be obtained from the same tow taken for surveying ichthyoplankton. This provides information on broad-scale plankton abundance and distribution for general feeding and predation assessment.
In view of a number of reports of juvenile fish being abundant in the neuston, some attempt to refine neustonic sampling techniques is desirable where significant mortality of the juvenile stages is proposed as a mechanism determining recruitment.
It will nearly always be possible to assemble some amount of data on environmental variability. This amount will always be limited in quantity and quality by the realities of feasibility and cost. The key problem is to find ways to derive the type of information needed (e.g. for fishery management) from the data that may be available (monitored environmental properties, etc.).
One crucial aspect is the integration of data corresponding to various time and space scales. For example, it is generally useful to combine long time-scale, low-spatial resolution information with short term, high resolution information. As an example we can think of recruitment as the outcome of biological processes occurring at short time-scales and small space-scales, i.e. egg, larvae and juvenile survival rates are controlled by the availability of food which is distributed in patches or micro-layers and by the density of predators. However, what might be important for a successful year-class to develop could actually be the relative availability of local high quality pieces of environment at a scale commensurable with the areal distribution of the spawning of the species. For the prediction of recruitment success, therefore, the definition of the quality of the small-scale environment assimilates to an intensive property and their relative availability to an extensive property of the system. As mentioned before, the properties to be measured are likely to be system-specific, although some analogy between similar systems in different geographic localities is expected.
The conversion of these measured properties to information relevant to fishery management, depends on identification of the nature of the causal linkages involved. This will either be on a rational basis, where sufficient mechanistic understanding is available, or on an empirical basis. In general, some combination of rational and empirical approaches will allow the greatest amount of available knowledge to be utilized.
When employing an empirical approach it is essential to recognise the difference between exploratory data analysis and hypothesis testing. In an exploratory data analysis the available data is used to formulate a “model” which describes the observed data; the resulting model is at that point an untested hypothesis, requiring additional data for verification. Hypothesis testing, on the other hand, is directed toward describing the ability of the model to predict outcomes that may not have been observed; it depends on an a priori definition of the model which is independent of the test data. Thus a large portion of published fish-stock-environmental relationships are really untested hypotheses and it is not surprising that they may fail when subjected to new data. This is not to say that exploratory data analysis is not a useful research activity; however, it should not be mistaken for hypothesis testing even though the statistical descriptors may appear to be similar.
In the absence of well-defined causal relationships the simplest exploratory approach is that of correlation studies. These have proven useful for identifying potential causes of changes in recruitment, however, they are limited in scope for several reasons. First, in order to obtain significant results, long-term series of data are needed; unfortunately fishery data series are characteristically short. Second, despite the fact that correlation techniques provided an objective basis on which to judge the results, unrestricted testing over the same set of data actually restricts very severely the actual number of degrees of freedom available for comparisons (multiple testing). This problem has often gone unrecognised and has given “significant” results that are formally incorrect.
Because it is likely that the variables being monitored will have some effect at different scales and may not have immediate effect on recruitment, it is advisable to employ cross-correlation methods that preserve their functional relationship with time, making allowances for the expression of time-lagged responses. For example, intense coastal upwelling upstream of, and preceding, spawning activity could be a favourable larval survival factor in its effect on abundance of appropriate food organisms; however, it could be an unfavourable factor if it occurs during the peak of spawning where it could result in lack of proper food organisms and in offshore dispersion of the larvae themselves.
At times different environmental or biological processes exhibit variability on distinctly different scales; if these are not separated (by filtering, etc.) the analysis may be hampered by interference and smearing among different processes. Certainly, failure to separate obvious scale differences that appear in data sets will result in loss of valuable degrees of freedom in the shorter scale components due to autocorrelation imposed by longer-scale components. One useful exploratory data analysis technique is spectral analysis, whereby historical information can be used to explore how total variability is partitioned among different time or space-scales.
It should be noted that it is not necessary that a causative process has the same dominant period of variation as the affected variable. (For example, one need not necessarily look for an environmental property with a 40-year period of variation as an explanatory variable for the two periods of high Japanese sardine catches.) Effects tend to “cascade” from shorter to longer scales. One or several short-lived storms during peak spawning periods might disrupt a year's recruitment; several strong or weak recruitments might determine long-term viability of an exploited stock.
Various exploratory data analysis techniques are available. The main point here is that they basically involve searching for relationships and structures within a set of observed data and exhibiting them in a compact and recognisable manner. In order to draw well-founded inferences about situations not in the observed data set, or those which have not yet occurred, it is necessary to shift the point of view toward hypothesis testing. Thus it is necessary to define a priori the functional relationships to be tested, hopefully based on fundamental principles or at least following a well-defined, explicit model or rationale layed out beforehand. In this sense it is more important to have at hand a well-defined rationale on how different processes are related, even if some of the details have been deliberately simplified, than to try every plausible relationship or alternatively to formulate complex models which have parameters that are difficult or impossible to measure accurately.
A goal of environmental studies is to provide a basis for the formulation of predictive models. At this stage, robustness of models must be given high priority. The systematic application of the simplest functional relationship, almost conceived as a null hypothesis on which to contrast results, will provide the foundation on which to build upon. First order approximations and linear theory should be applied and their explanatory and heuristic possibilities exhausted before proceding to more sophisticated formulations. This recommendation follows from the fact that neither the relationships linking the different system components, nor the variability (stochasticity) of variables are well enough understood to warrant other approaches.
The distinction between the dependence of small-scale variability of the environment on larger scales as distinct from the effect of both upon the biosystem (or subsystems), is of paramount importance. Parameterisation on a larger scale is often used to describe variability of a smaller scale process. The selection of variables to use will very often depend on system-specific characteristics: i.e. coral reefs, shelf systems and eastern boundary current systems, might require different sets of variables. Parameterisation of environmental variables should be based whenever possible on realistic physical or biological mechanisms. This is particularly important with variables where a single measurement, such as wind, could represent completely different processes depending on the choice of functional relationships parameterised. For example, characteristic Ekman transport over a given time period would be properly formulated as being proportional to a vector average of the wind velocity raised to the second power, whereas turbulent mixing energy added to the water column would be formulated as a scalar average of the wind speed raised to the third power.
Given the nature of the problem, with important processes taking place on different scales, development of parameters that could integrate extensive properties (large scale patterns of relevant variables) with intensive properties measured at a local scale (“quality” of local environmental) may be crucial to effective modelling efforts.
The requirements of experimental studies are of a different nature than those of environmental monitoring. They are specifically designed to test a hypothesis explicitly formulated. Consideration of the accuracy and precision of the variables to be measured with respect to the expected changes of the dependent variable is a prerequisite. Very often this implies the development of new sampling gear or laboratory techniques. For example our limited knowledge on juvenile or pre-recruit survival rates is basically due to the inadequacy of available sampling gear. Progress in this area will undoubtedly result in an increased explanatory ability.
Because of restrictions imposed by practicality and precision, field experiments will be focussed on small to medium-scale processes affecting individual survival (i.e. egg, larvae and juvenile survival), and on how these are influenced by short-term environmental changes. The research goal is to recognise and predict the quality of the environment on a local scale on some quantitative basis. The effects of pattern formation and destruction, mixing, and stratification are areas that can be studied in the field making full use of current technology.
A special area on which field experiments can be of great use, refers to the calibration of “proxy” environmental variables. Short term, high resolution field studies are particularly useful for understanding the mechanisms by which a global variable, that may in itself incorporate different types of information, will be manifested at a certain scale. For example, observations of the variation of the depth of the mixed layer could help to calibrate time series of the wind to describe turbulent mixing. Detailed hydrographical surveys could serve to intercalibrate time series of sea-level observations from tidal stations at the coast. Ground truth measurements of chlorophyll are needed to calibrate synthetic chlorophyll estimates from satellites.
A necessary complement to field studies, and sometimes a required pre-requisite, are laboratory studies. Here the emphasis is upon the explanation of causal mechanisms through the manipulation and control of environmental variables. In particular, measurements of rates is a goal frequently reserved for laboratory studies. The definition of the physiological requirements of the different stages of the early life history can be studied in the laboratory. This in turn provides meaningful information for identifying areas of the environment suitable for the organisms. Rates of feeding, the effect of the quality of food (particle size, specific composition) are well suited for laboratory experimentation. Other rate processes such as rate of predation and cannibalism are potentially suitable for laboratory experiments.
Intercomparisons among different regional systems exhibiting similar environmental dynamics and similar fish population assemblages may be sources of additional information. The costs of such studies may be limited to acquiring or exchanging data that may already have been taken in the various regions. Such comparisons may provide a firmer basis for evaluating the validity of transferring experience of fishery events in one region in order to predict outcomes of actions taken in another.
In addition, to the extent that closely-related species function as analogues, pattern recognition among seasonality and geography of spawning habits and environmental processes may yield information on the crucial mechanisms controlling reproductive success Since natural selection implies accommodation of reproduction strategies to the most crucial environmental factors, compelling patterns of correspondence suggest important causal linkages. Such information might serve to aid in choosing a strictly limited set of explanatory variables so as to make best use of limited degrees of freedom available in recruitment time series.
Finally, these studies may be useful in building statistical confidence in weak empirical relationships. We have noted that high correlations are not expected between recruitment which is affected by many environmental mechanisms acting on various scales at various life-cycle stages, and any one environmental variable. Empirical relationships which have the same general form in several independent regional systems would, on the assumption of analogues, acquire increased statistical significance. In fact, subjecting any empirical relationship to the discipline of inter-regional comparison would appear to offer some useful guidance as to validity and generality.
Implicit in the requirement for recruitment information for fisheries management is that there is a relationship between spawning stock size and eventual recruitment such that at some population level recruitment overfishing may occur. Identification of the form of this relationship, particularly the point at which recruitment is reduced, is, therefore, of great interest. However, there is considerable dissatisfaction with the concept of fitting of a single time-invariant “stock-recruitment curve” to a pattern of data points which exhibits a great amount of “environmental” scatter and may contain substantial serial correlation.
It has been suggested that a family of stock-recruitment curves, representing a range of environmental situations, may be a more satisfactory way to approach the issue. This family of curves may look like those in Figure 3, where R represents recruitment due to spawning of a parental stock size, S, and ei represent various environmental states.
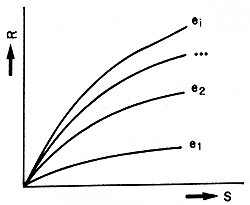
Figure 3. A family of stock-recruitment curves
Heuristically, this makes very good sense; however, the problems in properly fitting a family of multi-parameter curves to a very limited number of actual data points, in a purely empirical manner, are formidable. There is substantial danger of generating pure nonsense by non-judicious use of such a procedure.
One way to objectively fit such a family of curves is by means of a multiple regression in which stock size may appear as one explanatory variable with one or more environmental variables. For example, the formulation:
In(R/S) = ao + a1S + a2ej + …
will generate a family of “Ricker-type” curves for constant values of the environmental variable(s), e. Note that ej could also represent a multispecies effect, e.g. the size of a predator stock, etc. Note also that this particular formulation represents a multiplicative model; i.e. the explanatory variables impact recruitment in linear proportion to the potential recruits present, rather than affecting absolute numbers as such. Such a multiplicative model would be appropriate for processes such as adult cannibalism on larvae by non-selective filtration of a portion of the water volume occupied, loss of a fraction of potential recruits by offshore dispersion, starvation or growth-dependent predation loss due to inadequate food particle availability, etc. Where other types of mechanisms operate, e.g. where limited habitat space may be filled by potential recruits or larvae may be abundant enough to substantially alter their ambient food supply, different (e.g. additive rather than multiplicative) formulations may be more appropriate.
Note that this example is only one of a range of possible formulations, the determination of which depends on the particular mechanisms involved. Further, it should be emphasised that, before much credence is assigned to the form of any apparent density dependence which may emerge from a fitting procedure of this type, two conditions must be fulfilled. First, the relationship to stock size must represent a substantial portion of the explained variance. Second, there must be a high degree of confidence that the environmental portion of the relationship is realistically modelled, thereby ensuring that the form of the stock dependence is not controlled by possible uncertainties in the environmental dependence.
Of course, situations where there are only two, or at most several, well-defined and separate environmental states (e.g. El Niño vs. non-El Niño vs. Anti-El Niño etc.) with corresponding separate clusters of data points on the stock recruitment diagram, it may be possible to directly fit separate curves.
However, it should be re-emphasised that models produced by purely empirical fitting procedures constitute exploratory data analyses, which are merely descriptions of the data already at hand. Their ability to yield information on possible situations not yet observed should be regarded as an untested hypothesis. Such an analysis can be regarded as a useful predictor only when the variables incorporated do indeed represent the dominant causative mechanisms.
There is a requirement for a more rigorous approach to the problem of environmental effects on fishery problems. Because of the complexity and number of possible interrelationships between the physical and biological aspects of an ecosystem, it is essential to have a sound conceptual basis for research activities. Simple searches for correlations with available environmental data series are likely to be unrewarding in the long run.
The benefits of supporting observational programs with experimental efforts and laboratory studies may be substantial. Interregional sharing of results and comparative studies may also be beneficial.
The ability to forecast effects of environmental anomalies on recruitment is an important goal. However, lacking this ability, the simple knowledge that an environmental anomaly may be present is useful in itself as a possible motivation for conservative management strategies or preparation for short lead-time adaptive management actions.
Valuable information on environmental variations may be obtainable at relatively low cost from maritime reports and from regular measurement of sea level, sea temperature, and wind at exposed coastal locations.
Understanding of processes operating at a scale commensurate with feeding and survival of fish larvae may be a key to understanding recruitment variation. Important advances may accrue from development of plankton sampling capable of resolving fine horizontal scales and the even finer scales associated with vertical structure of fish larvae, and planktonic food and predator organisms.
Care should be taken that empirical analysis procedures are appropriate; high value should be placed on ensuring statistical robustness of conclusions. Processes should be addressed on the scales at which they are acting, or dependence on larger scales should be parameterised according to a well-defined model of the linkage between scales.
Focus should be kept on integration of the various levels of knowledge which may be developed so as to bear directly on real world management concerns.
Bakun, A. et al., 1982 Ocean sciences in relation to living resources. Can.J.Fish.Aquat.Sci., 39(7):1059-70
SCOR Working Group 67, 1982 Report of the SCOR Working Group 67 on oceanography, marine ecology and living resources. Proc.Sci.Comm.Ocean Res., (18):57–67
Sharp, G.D. (Rapporteur), 1981 Workshop on the effects of environmental variation on survival of larval pelagic fishes, organised by FAO as a contribution to the IOC Programme of Ocean Sciences and Living Resources. Lima, 20 April - 5 June 1980. IOC Workshop Rep., (28):323 p.
![]()
![]()
![]()