


El maíz (Zea mays L.) es una especie única: por la gran diversidad genética de la planta, de la mazorca y del grano; por su adaptación a gran rango de ambientes; por su resistencia a enfermedades e insectos; por su tolerancia a distintos estreses ambientales, por sus múlti-ples usos como alimento humano o animal y por la gran variedad de productos que se obtienen de esta especie. El maíz apareció entre los años 8 000 y 5 000 A.C. (capítulo Introduc-ción al maíz y su importancia). Ha evolucio-nado por selección natural, por la selección dirigida por los agricultores-mejoradores durante miles de años y por los mejoradores profesionales en los últimos 150 años. Existen aún una serie continua de tipos de plantas que van desde sus antecesores salvajes a razas mas avanzadas, cultivares mejorados y mantenidos durante generaciones por los agricultores y las variedades mejoradas de polinización abierta con una base genética amplia, obtenidas profesionalmente. Al final de todo este espectro de materiales están los distintos tipos de híbridos: desde híbridos intervarietales hasta cruzas simples con un preciso diseño genético y de base genética angosta para satisfacer propósitos y ambientes especiales.
Muchos mejoradores de maíz piensan que en esta planta se encuentra toda la variabilidad genética necesaria para mejorar cualquier característica que se desee. En realidad los programas de mejoramiento han hecho poco o ningún uso de la diversidad de los recursos genéticos del maíz disponibles fuera de los criaderos de mejoramiento. Ha habido una tendencia general a usar el mismo germoplasma limitado disponible en los trabajos experimentales. Nass, Pellicano y Valois (1994) llevaron a cabo encuestas entre los mejora-dores de maíz y soja en Brasil para evaluar el uso de las accesiones de los bancos de germoplasma y las fuentes de variabilidad que se están usando en los programas de mejora-miento. El estudio mostró que las accesiones de los bancos de germoplasma son normal-mente usadas por los fitomejoradores de soja, un cultivo autofecundo, pero que las acce-siones de los bancos de germoplasma eran escasamente usadas por los fitomejoradores de maíz. La principal fuente de variabilidad genética usada en los trabajos con maíz provenía en general de los materiales de que disponía el mejorador o de un intercambio de germoplasma con otros mejoradores. Esta situación no parece ser muy distinta de lo que ocurre en los Estados Unidos de América donde hay fuertes programas de mejoramiento de maíz. Goodman (1985) documentó que solo el 4% del área de maíz de ese país es sembrada con material que contiene una cantidad muy limitada -10-25%- de germoplasma extranjero en su composición genética. De cualquier manera, hay una creciente preocupación entre los mejoradores profesionales de maíz para expandir la búsqueda de genes útiles que aumenten la variabilidad genética de modo de incrementar la sostenibilidad de la producción. De la misma forma, hay una constante aprecia-ción de la necesidad de conservar los recursos genéticos para su uso en el siglo XXI y en el futuro mas lejano. Durante una discusión sobre el tema de la conservación y uso de los recursos fitogenéticos en un panel de la "Crop Science I Conference" llevada a cabo en Ames, Iowa, EUA, (Williams y Duvick, 1993), hubo consenso en que ahora había una mayor conciencia entre los fitomejoradores respecto a los problemas de los recursos fitogenéticos. Los programas de recursos fitogenéticos necesitan establecer relaciones mas estrechas entre los fitomejoradores y los biotecnólogos para un uso mas efectivo de los genes poco conocidos presentes en los recursos fito-genéticos disponibles. Un mecanismo para acelerar este desarrollo alrededor del mundo sería una mayor comprensión y conocimiento de los recursos fitogenéticos del maíz, su utilidad y su disponibilidad.
Los recursos genéticos que tienen un uso potencial en el mejoramiento del maíz se pueden agrupar en siete clases principales. El objetivo de este trabajo no es el de propor-cionar una lista exhaustiva de recursos de germoplasma sino el de dar una idea del amplio espectro de recursos fitogenéticos que pueden ser explotados para el mejoramiento del maíz; se darán unos pocos ejemplos en cada caso particular. Dowswell, Paliwal y Cantrell (1996) han tratado en detalle el tema de los recursos genéticos del maíz, incluyendo una lista de germoplasma y las fuentes de disponibilidad.
Géneros muy distantes que pudieran ser posibles fuentes de genes útiles contra los estreses naturales se podrían encontrar en las Maydeas orientales (ver capítulos Introduc-ción al maíz y su importancia y Origen, evolución y difusión del maíz) y en especies de la tribu de las Andropogoneas, p.ej. el sorgo. Solamente Harda et al., (1954) y Harda, Umekage y Nakazima (1955) informaron acerca de la hibridación entre Coix, un género de Maydeas orientales, y el maíz. También se ha informado acerca de nueva diversidad en alguno de los géneros de las Maydeas orien-tales, especialmente en el género Coix (Koul y Paliwal, 1964; Venkateswarlu y Chaganti, 1973; Venkateswarlu, Cahganti y Rao, 1976; Jain y Banerjee, 1974; Arora y Nayar, 1986). Las Maydeas orientales existen en sus hábitats naturales y ahora están siendo mantenidas ex situ en algunos bancos de germoplasma -por ejemplo, NBPGR, India; NJAS, Japón. Podría haber algunos genes extraños en las Maydeas orientales y en las Andropogoneas que pudieran ser transferidos al maíz con las herramientas biotecnológicas de que disponen ahora los fitomejoradores. Por medio del análisis de marcadores moleculares se ha encontrado evidencia de que hay considerable semejanza genética entre las diferentes especies de Gramíneas; el análisis genético molecular; ha puesto en evidencia las antiguas relaciones entre el maíz, el arroz, el sorgo y el trigo (Paterson et al., 1995). Los intercambios genéticos entre el maíz y otros géneros po-drían ser mas frecuentes en el futuro próximo.
Los genes del teosinte se han introducido en el maíz a través del proceso evolucionario y de domesticación (ver capítulo Origen, evolución y difusión del maíz); (Galinat, 1988, 1992, 1995; Wilkes, 1977, 1985, 1989). Por otro lado, hay ciertas razas de teosinte que pre-sentan una barrera para los cruzamientos con el maíz (Kermicle y Allen, 1990). Es un hecho generalmente aceptado que las especies anuales y perennes de teosinte contienen una reserva de características deseables que po-drían ser útiles para aumentar la producción sostenible actual y futura del maíz. Wilkes (1988, 1989, 1993), Benz, Sánchez Velásquez y Santana Michel (1990), Sánchez et. al.,(1995), y Taba (1995b) han descripto varias razas de teosinte y sus hábitats naturales. Todas las razas conocidas de teosinte todavía sobre-viven in situ en los campos de los agricultores junto con el maíz, en áreas sin cultivar y en algunos casos en áreas de reservas de bio-diversidad. Estos y otros autores han enfa-tizado la necesidad de fortalecer e intensificar la preservación y supervisión in situ del teosinte en México y Guatemala. Los bancos de germoplasma de INIFAP y CIMMYT en México coleccionan y mantienen semillas de diferentes razas de teosinte en sus bancos de germoplasma en condiciones de almacena-miento a mediano y largo plazo. Sin embargo, hay una urgente necesidad de crear sistemas para el incremento y el matenimiento rutinario de las accesiones de teosinte (Taba, 1995b).
En la historia de la evolución del maíz, del teosinte y del Tripsacum, no hay una evidencia clara del intercambio de germoplasma entre Tripsacum con maíz o teosinte. Sin embargo, Tripsacum es el único género con el cual se ha cruzado el maíz, en condiciones experi-mentales, y cuyos segmentos de DNA han sido transferidos al maíz, si bien en forma limitada (ver capítulo Origen, evolución y difusión del maíz). Un informe reciente indica que se han producido híbridos fértiles de Tripsacum dactyloides x Zea diploperennis (Eubanks, 1995). Es probable que con alguna ayuda de las nuevas herramientas tales como los marca-dores moleculares, la hibridación somática y las técnicas diferenciales de tinción para la identificación de la transferencia de segmentos cromosómicos, se pueda progresar rápida-mente en la transferencia de características deseables de Tripsacum a maíz. Reciente-mente, Tsanev y Todorova (1994) han in-formado que los modelos electroforéticos de esterasas pueden ser usados para controlar la presencia de material genético de teosinte y Tripsacum dactyloides en el genomio de maíz, mientras que modelos electroforéticos de zeína son mas adecuados para detectar la presencia de material genético de T. dactyloides. Galinat (1988) y Wilkes (1989) describieron los beneficios que pueden derivarse de la transferencia de genes de Tripsacum a maíz. Ejemplos de tales características son resis-tencia a los insectos, apomixis y resistencia a la maleza Striga sp. (Bertrand et al., 1995; Savidan, Grimanelli y Leblanc, 1995). La transferencia de genes apomícticos al maíz con la ayuda de marcadores RFLP (Leblanc et al., 1995) será el primer uso de un carácter de un antecesor salvaje en el mejoramiento del maíz.
La información sobre la sistemática del Tripsacum se encuentra en los trabajos de Randolph (1970) y de de Wet et al. (1981, 1982, 1983). La mayor biodiversidad se encuentra en México, Guatemala y en algunas partes de América del Sur. Berthaud et al. (1995) listaron 20 especies de Tripsacum, muchas de cuyas poblaciones continúan existiendo in situ. Sin embargo, son muy susceptibles al pastoreo y son muy vulnerables al peligro de extinción, si es que no están ya en peligro. La conservación ex situ ha sido llevada en la forma de un jardín de introducción de Tripsacum en el CIMMYT, en México, donde se han establecido mas de 1 000 colecciones; además semillas de Trip-sacum son mantenidas en bancos de germo-plasma en México. Los recursos genéticos de Tripsacum se pueden obtener en forma vegetativa como esquejes, los cuales corres-ponderán exactamente al tipo, o por semillas cuyas progenies serán siempre variables (Berthaud et al., 1995).
Una colección de diferentes razas de maíz, sobre todo del hemisferio occidental, es mantenida en diversos bancos de germo-plasma. Taba (1995a) ha hecho una lista de las razas de maíz almacenadas en los bancos nacionales de germoplasma de México, América Central y América del Sur y en el CIMMYT. Estas representan una enorme reserva de diversidad genética, mucha de la cual no ha sido aún explotada para el desarrollo de cultivares mejorados. Dowswell, Paliwal y Cantrell (1996) estimaron que menos del 10% de la diversidad genética del maíz acumulada en todas las razas de maíz está probablemente siendo usada como material para el mejora-miento de los cultivares. Un artículo extenso sobre las razas de maíz ha sido escrito por Goodman y Brown (1988). Muchos de los términos usados cuando se hace referencia a las reservas de diversidad genética son inter-cambiables: razas de maíz, razas locales, super razas, sub-razas, tipos de maíz primitivo, grupos raciales o geográficos y complejo de razas. Anderson y Cutler (1942) introdujeron el concepto de razas de maíz; cada raza repre-senta un grupo de individuos relacionados con suficientes características en común como para permitir su reconocimiento como grupo, teniendo un alto número de genes comunes. Mas adelante, Anderson y sus colaboradores desarrollaron y definieron el concepto de raza para que fuera mas útil para la descripción del maíz. La información sobre las razas de maíz recolectadas en varios países o regiones americanas ha sido publicada bajo la forma de boletines por los diversos autores; la informa-ción incluye la descripción de las medias de la raza para la planta, la panoja, la mazorca, el grano y, en algunos casos, características fisiológicas y datos de los nudos cromo-sómicos (para listas y referencias ver Goodman y Brown, 1988). Brandolini (1970) describió las razas de maíz a nivel universal e indicó los centros primarios y secundarios de diferencia-ción de las razas. Mangelsdorf (1974) dividió todas las razas de maíz de América Latina en seis grupos de linajes, cada grupo derivado de una raza salvaje de maíz. Estos grupos son:
Goodman y Bird (1977) revisaron la información sobre las razas de maíz en América Latina y describieron una clara relación entre las razas en ese hemisferio. McClintock, Kato y Blumenshein (1981) publicaron un estudio completo sobre la constitución cromosómica, incluyendo los cromosomas B y los nudos cromosómicos, en 1 352 colecciones de maíces en las Américas y el Caribe. Estas carac-terísticas proporcionan buena información, no influenciada por las condiciones ambientales, para determinar las relaciones entre las varias razas de maíz. Las variaciones alélicas de las isoenzimas han sido estudiadas en varias razas y proporcionan una herramienta adicional para una mejor comprensión de las relaciones raciales (Bretting, Goodman y Stuber, 1987).
Paterniani y Goodman (1977) estimaron que cerca del 40% de las razas de maíz en las Américas tienen endospermo harinoso, 30% son duros, 20% son dentados y 3% tienen tipos de granos dulces. Además informaron que muchas de las razas están adaptadas a los trópicos, 50% adaptadas a los ambientes de las tierras bajas, 40% a las tierras altas y 10% a los ambientes intermedios. Pandey y Gardner (1992) mencionaron que en el hemisferio occidental se han descripto 285 razas de maíz, de las cuales 265 están presentes en América Latina, la mayoría de ellas en América del Sur. Anteriormente, Hallauer y Miranda (1988) habían informado que podrían haber solo 130 razas distintas de maíz en el hemisferio occidental. Dowswell, Paliwal y Cantrell (1996), indicaron que cerca de 300 razas de maíz involucrando a miles de cultivares diferentes habían sido descriptas e identificadas en todo el mundo y que esas colecciones represen-taban del 90 al 95% de la diversidad genética del maíz. Las listas de las razas que están siendo usadas en los programas de mejora-miento de maíz en México y países de América Central y del Sur han sido presentadas por Taba (1995a).
Casi todos los cultivares que se usan hoy día en los trópicos pueden ser reconducidos a una raza que todavía se puede encontrar en el campo y/o está depositada en un banco de germoplasma. Algunas razas muestran una amplia adaptabilidad y han proporcionado germoplasma que se encuentra en casi todos los ambientes tropicales. Una de ellas es el Tuxpeño; varios cultivares mejorados que se originan directamente en esta raza o en una combinación con otros materiales, están siendo cultivados desde México a China. La genealogía de los cultivares mejorados que se originan en el Tuxpeño se presenta en la Figura 5. Esta raza ha sido usada como germoplasma con fuente de resistencia a las enfermedades foliares, especialmente al marchitamiento de la hoja, por su buena calidad del tallo, por su resistencia a la sequía, por la tolerancia a la toxicidad de aluminio, como fuente para un tipo de planta mas bajo con un mejor índice de cosecha, para variedades que pueden tolerar un cierto grado de inundación y también para el transplante. El germoplasma Tuxpeño muestra una alta habilidad combinatoria con la raza Tuscón y con los maíces duros de la costa y con los maíces duros de Cuba. El híbrido Tuxpeño x ETO está probablemente presente en los parentales de un gran número de híbridos en los trópicos. Muchas otras razas también son fuentes útiles de germoplasma. Goodman y Brown (1988) y Pandey y Gardner (1992) han hecho una lista de razas que han sido efectivamente usadas para mejorar características específicas.
Árbol genealógico del complejo del germoplasma Tuxpeño
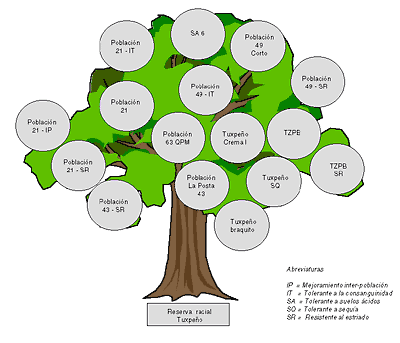
Fuente: adaptado de Dowswell, Paliwal and Cantrell, 1996.
La información sobre las razas de maíz y la biodiversidad en Asia y África es muy limitada. Alguna información ha sido publicada sobre la biodiversidad del maíz en Asia; los estudios mas importantes son: Stonor y Anderson (1949); Anderson y Brown (1953); Suto y Yoshida (1956); Mochizuki (1968); Singh (1977a, 1977b) y Jotshi (1982).
En todos los ambientes en los que se ha sido cultivado el maíz durante siglos -comparado con aquellas áreas donde el maíz ha sido introducido hace relativamente poco tiempo- los cultivares de maíz han sido mantenidos, desarrollados y mejorados por agricultores-mejoradores durante muchas generaciones y son cultivados aún hoy día. Estos cultivares reciben varios nombres tales como variedades primitivas, variedades de los agricultores o variedades locales. Han sido mantenidos y mejorados in situ por los agricultores, basados en la percepción de sus necesidades y su experiencia y su capacidad naturales y no han sido sometidos a los procesos de selección y mejoramiento por mejoradores profesionales de maíz. Estas variedades locales pueden no representar una fuente de diversidad genética como aquella de que se dispone en los bancos de germo-plasma, pero son fuentes de características que son importantes para la adaptación local, la estabilidad económica y la sostenibilidad del agricultor. Son características que pueden no estar disponibles en las variedades mejoradas mejoradas desarrolladas por los mejoradores profesionales donde el rendi-miento es el objetivo principal y dirigidos a ambientes favorables con prácticas especiales de manejo y cultivo. Algunos de los cultivares locales han sido excelentes fuentes de genes para resistencia a hongos o insectos y podrían ser fuente de genes para otros estreses. Por ejemplo, los genes para resistencia al mildio lanuginoso en Asia, para resistencia al estriado en África y al marchitamiento tardío en Medio Oriente e India, provienen de variedades de los agricultores. Estas variedades locales están siendo reemplazadas lenta pero constan-temente por cultivares mejorados. Se cree que muchas de estas variedades locales han sido coleccionadas y conservadas ex situ en bancos de germoplasma de maíz para su uso futuro. Sin embargo, la conservación ex situ debería, por lo menos, suplementar la con-servación in situ de estas variedades y no ser un sustituto. Soleri y Smith (1995) analizaron los datos morfológicos y fenológicos de dos variedades de maíz Hopi conservadas in situ y ex situ. Las conservaciones ex situ sufrieron cambios y derivaciones genéticas, y el maíz Hopi mantenido ex situ fue diferente de aquel mantenido in situ. Hay una diferencia respecto a la utilidad de la diversidad genética en el concepto de los mejoradores profesionales y en el concepto de los agricultores usuarios del producto. Los agricultores buscan y pueden usar la máxima diversidad que se encuentra en gran número de las colecciones y de los bancos de germoplasma ex situ y están interesados en acumular y conservar tal diversidad y estructura de la población que proporciona adaptación a las condiciones locales. Existe hoy día una mayor apreciación de la necesidad de la conservación in situ de las variedades locales y de los cultivares primitivos en las condiciones en que estos pueden continuar a evolucionar junto con los estreses de los cultivos y las necesidades de los agricultores: una conservación evolucio-nista (Worede, 1993; Brush, 1995).
Otro recurso genético en esta categoría son los cultivares obsoletos, que hace algunas décadas pudieron ser los mejores cultivares disponibles pero que han sido reemplazados por nuevas generaciones de cultivares. Los genes de esos cultivares obsoletos se pueden encontrar en las líneas parentales de muchos cultivares actuales; sin embargo, esos culti-vares obsoletos han sido cultivados en los campos de los agricultores, en algunos casos por muchos años, se han cruzado con otros maíces cultivados en la vecindad y han sido, voluntaria o involuntariamente, sujetos a alguna forma de presión de selección por parte de los agricultores para algunos caracteres que a su criterio eran importantes, en especial para estabilidad y sostenibilidad económica. Estos cultivares obsoletos fueron en su momento nuevas variedades para los agricultores y deben ser conservados tanto ex situ como in situ. Podrían ser una fuente de características útiles cuando el énfasis del mejoramiento se dirige a los ambientes margi-nales y cuando se buscan cultivares para aumento de la producción sostenible. Bellon y Brush (1994) informaron que en Vicente Guerrero, Chiapas, México, generaciones avanzadas de cultivares mejorados que habían sido introducidas varias décadas antes, se mezclaron con las variedades locales y hoy día son manejadas como tales por los agricultores. De este modo, es posible que la erosión genética ocurra en forma mas lenta que en otras especies autofecundas. Los agricultores por lo general siembran varie-dades mejoradas y sus propias variedades de maíz en terrenos adyacentes lo que da lugar a la formación de nuevas variedades de los agricultores con las combinaciones deseables de genes de ambas fuentes.
Los cultivares mejorados actuales -varie-dades, variedades sintéticas, híbridos, líneas puras- representan las fuentes de materiales genéticos mas usados en casi todos los programas de mejoramiento de maíz. Sin embargo, tales programas abarcan solamente una minúscula fracción de la diversidad genética. De cualquier manera, las ganancias inmediatas de los esfuerzos en el mejoramiento del maíz para aumentar los rendimientos son mucho mayores que el uso de los mejores cultivares existentes. Es por lo tanto dable esperar que los mismos continuarán siendo recursos genéticos importantes para el mejora-miento del maíz, especialmente si los campos de cría de los mejoradores tienen una buena colección de tal germoplasma de distintas fuentes.
En las últimas tres o cuatro décadas varios programas nacionales de mejoramiento de maíz tropical y los centros regionales e internacionales de mejoramiento han desarro-llado complejos de germoplasma de base amplia, combinaciones y poblaciones que han demostrado ser fuentes excelentes para el desarrollo de variedades y de variedades sintéticas superiores de polinización abierta y también líneas puras e híbridos. Los recursos genéticos del maíz disponibles para los mejora-dores en los trópicos son hoy día mejores y mas diversos que los que estaban disponibles en los años 1950 y 1960. Paliwal y Sprague (1981), Paterniani (1985, 1990), Pandey y Gardner (1992) y Dowswell, Paliwal y Cantrell (1996) han listado y descripto tales recursos genéticos superiores. Es deseable que estos recursos continuen aumentando y que sean mas intensamente utilizados para el mejora-miento del maíz en los trópicos.
Los recursos genéticos especiales, inclu-yendo los genes y los cromosomas mutantes inducidos natural o artificialmente, son otra fuente importante de recursos genéticos. Los maíces azucarados y cerosos, el opaco-2 o el maíz de proteínas de calidad y el maíz folioso son unos pocos ejemplos de mutantes naturales que han tenido un amplio uso. Los ejemplos de otros tipos de materiales son los cromosomas mutantes, los marcadores gené-ticos, los marcadores moleculares y los maíces RFLP para pruebas. Organizaciones tales como el GRIN -Red de Información de Recursos Genéticos- de la Universidad de Missouri, el USDA, Beltsville, las Existencias de Base de Illinois y la Organización para las Semillas de Base de Hawaii, Honolulu, todas ellas en los EEUUA, son algunas de las fuentes que proporcionan información y materiales. Parte de esta información está también disponible en forma electrónica en los sistemas de información Internet y World Wide Web.
Anderson, E. & Brown, W.L. 1953. The pop corns of Turkey. Ann. Mo. Bot. Gard., 40: 33-48.
Anderson, E. & Cutler, H.C. 1942. Races of Zea mays. I. Their recognition and classifi-cation. Ann. Mo. Bot. Gard., 29: 69-89.
Arora, R.K. & Nayar, E.R. 1986. Wild relatives and related species of crop plants in India - their diversity and distribution. Bull. Bot. Surv. Ind., 25: 35-45.
Bellon, M.R. & Brush, S.B. 1994. Keepers of maize in Chiapas, Mexico. Econ. Bot., 48: 196-209.
Benz, B.F., Sánchez-Velásquez, L.R. & Santana-Michel, F.J. 1990. Ecology and ethnobotany of Zea diploperennis: preliminary investigations. Maydica, 35: 85-98.
Berthaud, J., Savidan, Y., Barre, M. & Leblanc, O. 1995. Tripsacum: its diversity and conservation. In S. Taba, ed. Maize genetic resources, p. 74-85. Mexico, DF, CIMMYT.
Brandolini, A. 1970. Maize. In O.H. Frankel & E. Bennett, eds. Genetic resources in plants - their exploitation and conserva-tion, p. 273-309. Philadelphia, PA, USA, F.A. Davis.
Bretting, P.K., Goodman, M.M. & Stuber, C.W. 1987. Karyological and isozyme variation in west Indian and allied main land Ameri-can races of maize. Am. J. Bot., 74: 1601-1613.
Brush, S.B. 1995. In situ conservation of landraces in centers of crop diversity. Crop Sci., 35: 346-354.
de Wet, J.M.J., Brink, D.E. & Cohen, C.E. 1983. Systematics of Tripsacum section Fasciculata (Gramineae). Am. J. Bot., 70: 1139-1146.
de Wet, J.M.J., Harlan, J.R. & Brink, D.E. 1982. Systematics of Tripsacum dactyloides. Am. J. Bot., 69: 125-127.
de Wet, J.M.J., Timothy, D.H., Hilu, K.W. & Fletcher, G.B. 1981. Systematics of South American Tripsacum (Gramineae). Am. J. Bot., 68: 269-276.
Dowswell, C.D., Paliwal, R.L. & Cantrell, R.P. 1996. Maize in the third world. Boulder, CO, USA, Westview Press.
Eubanks, M. 1995. A cross between two maize relatives: Tripsacum dactyloides and Zea diploperennis (Poaceae). Econ. Bot., 49: 172-182.
Galinat, W.C. 1988. The origin of corn. In G.F. Sprague & J.W. Dudley, eds. Corn and corn improvement, 3rd ed., p. 1-31. Madison, WI, USA, American Society of Agronomy.
Galinat, W.C. 1992. Evolution of corn. Adv. Agron., 47: 203-229.
Galinat, W.C. 1995. The origin of maize: grain of humanity. Econ. Bot., 49: 3-12.
Goodman, M.M. 1985. Exotic maize germplasm: status, prospects, and remedies. Iowa State J. Res., 59: 497-527.
Goodman, M.M. & Bird, R.McK. 1977. The races of maize. IV. Tentative grouping of 219 Latin American races. Econ. Bot., 31: 204-221.
Goodman, M.M. & Brown, W.L. 1988. Races of corn. In G.F. Sprague & J.W. Dudley, eds. Corn and corn improvement, 3rd ed., p. 33-79. Madison, WI, USA, American Society of Agronomy.
Hallauer, A.R. & Miranda, J.B. 1988. Quantitative genetics in maize breeding, 2nd ed. Ames, IA, USA, Iowa State University Press.
Harda, K.M., Murakami, M., Fukushina, A. & Nakazima, M. 1954. Breeding study of the forage crops: studies on the inter-generic hybridization between the genus Zea and Coix (Maydeae). I. Kyoto Pref. Univ. Fac. Agric. Sci. Rep., 6: 139-145.
Harda, K.M., Umekage, O. & Nakazima, M. 1955. Studies on the intergeneric hybridization between the genus Zea and Coix (Maydeae). Jap. J. Breed., 4: 288.
Jain, S.K. & Banerjee, D.K. 1974. Preliminary observations on the ethnobotany of the genus Coix. Econ. Bot., 28: 38-42.
Jotshi, P.N. 1982. Knobs in Kashmir maize II. Nucleus, 25: 152-161.
Kermicle, J.O. & Allen, J.O. 1990. Cross-incompatibility between maize and teosinte. Maydica, 35: 399-408.
Koul, A.K. & Paliwal, R.L. 1964. Morphology and cytology of a new species of Coix with 32 chromosomes. Cytologia, 29: 375-386.
Leblanc, O., Grimanelli, D., González de León, D. & Savidan, Y. 1995. Detection of the apomixis mode of reproduction in maize - tripsacum hybrids using maize RFLP markers. Theor. Appl. Genet., 90: 1198-1203.
Mangelsdorf, P.C. 1974. Corn. Its origin, evolution and improvement. Cambridge, MA, USA, Harvard University Press.
McClintock, B., Kato-Y, T.A. & Blumenshein, A. 1981. Chromosome constitution of races of maize. Chapingo, Mexico, Colegio de Postgraduados.
Mochizuki, N. 1968. Classification of local strains of maize in Japan and selection of breeding materials by application of principal component analysis. In Symp. Maize Production in Southeast Asia, p. 173-178. Kasumigaseki, Chiyoda-ku, Tokyo, Japan, Agriculture, Forestry and Fisheries Res. Counc., Ministry of Agriculture and Forestry.
Nass, L.L., Pellicano, I.J. & Valois, A.C.C. 1994. Utilization of genetic resources for maize and soybean breeding in Brazil. Revista Brasileira de Genetica, 16: 983-988.
Paliwal, R.L. & Sprague, E.W. 1981. Improving adaptation and yield dependability in maize in the developing world. Mexico, DF, CIMMYT.
Pandey, S. & Gardner, C.O. 1992. Recurrent selection for population, variety, and hybrid improvement in tropical maize. Adv. Agron., 48: 2-79.
Paterniani, E. 1985. State of maize breeding in tropical areas of South America. In A. Brandolini & F. Salamani, eds. Breeding strategies for maize production improvement in the tropics, p. 329-339. Rome, FAO, Florence, Italy, Istituto Agronomico per L'Oltremare.
Paterniani, E. 1990. Maize breeding in the tropics. Crit. Rev. Plant Sci., 9: 125-154.
Paterniani, E. & Goodman, M.M. 1977. Races of maize in Brazil and adjacent areas. Mexico, DF, CIMMYT.
Paterson, A.H., Lin, Y.R., Li, Z., Schertz, K.F., Doebley, J.F., Pinson, S.R.N., Liu, S.C., Stansel, J.W. & Irvine, J.E. 1995. Convergent domestication of cereal crops by independent mutations at corresponding genetic loci. Science, 269(5231): 1714-1718.
Randolph, L.F. 1970. Variation among tripsacum populations of Mexico and Guatemala. Brittonia, 22: 305-337.
Sánchez González, J.J. & Chuela, M. 1995. Systematic and ecogeographic studies on crop genepools: distribucion y caracterizacion del teocintle. Montecillo, Mexico, INIFAP and Colegio de Postgraduados.
Savidan, Y., Grimanelli, D. & Leblanc, O. 1995. Transferring apomixis from tripsacum to maize: progress and challenges. In S. Taba, ed. Maize genetic resources, p. 86-92. Mexico, DF, CIMMYT.
Singh, B. 1977a. Races of maize in India. New Delhi, Indian Council of Agri. Res.
Singh, B. 1977b. Evolution of primitive cultivars from north eastern Himalayan region in relation to lineage. Indian J. Genet., 37: 103-113.
Soleri, D. & Smith, S.E. 1995. Morphological and phenological comparisons of two Hopi maize varieties conserved in situ and ex situ. Econ. Bot., 49: 56-77.
Stonor, C.R. & Anderson, E. 1949. Maize among the hill peoples of Assam. Ann. Mo. Bot. Gard., 36: 355-404.
Suto, T. & Yoshida, Y. 1956. Characteristics of the oriental maize. In H. Kihara, ed. Land and crops of Nepal Himalaya, vol. 2, p. 375-530. Kyoto, Japan, Fuana and Flora Res. Soc., Kyoto University.
Taba, S. 1995a. Maize germplasm: its spread, use and strategies for conservation. In S. Taba, ed. Maize genetic resources, p. 7-58. Mexico, DF, CIMMYT.
Taba, S. 1995b. Teosinte: geographic variations and conservation. In S. Taba, ed. Maize genetic resources, p. 59-72. Mexico, DF, CIMMYT.
Tsanev, V. & Todorova, L. 1994. Use of electrophoretic patterns of esterase and prolamins for proving genetic material from teosinte and tripsacum in maize genome. Dokl. Bulg. Akad. Nauk., 47: 89-92.
Venkateswarlu, J. & Chaganti, R.S.K. 1973. Job's tears (Coix lacryma-jobi L.). New Delhi, ICAR.
Venkateswarlu, J., Chaganti, R.S.K. & Rao, P.N. 1976. Pachytene chromosome morphology and its bearing on inter-specific and intergeneric relationship of Coix. Bot. Mus. Leafl. Harv. Univ., 24: 205-224.
Wilkes, H.G. 1977. Hybridization of maize and teosinte in Mexico and Guatemala and the improvement of maize. Econ. Bot., 31: 254-293.
Wilkes, H.G. 1985. Teosinte: the closest relative of maize revisited. Maydica, 30: 209-223.
Wilkes, H.G. 1988. Teosinte and the other wild relatives of maize. In Recent Advances in the Conservation and Utilization of Genetic Resources. Proc. Global Maize Germplasm Workshop, p. 70-80. Mexico, DF, CIMMYT.
Wilkes, H.G. 1989. Maize: domestication, racial evolution and spread. In D.R. Harris & G.C. Hillman, eds. Forage and farming, p. 440-454. London, Unwin Hyman.
Wilkes, H.G. 1993. Conservation of maize crop relatives in Guatemala. In Perspectives on biodiversity: case studies of genetic resource conservation and development, AAAS Publication 93-10S, p. 75-88. Washington, DC.
Williams, J.T. & Duvick, D.N. 1993. Conservation and use of plant genetic resources - discussion. In International crop science I, p. 451. Madison, WI, USA, Crop Science Society of America.
Worede, M. 1993. The role of Ethiopian farmers in the conservation and utilization of crop genetic resources. In International crop science I, p. 395-399. Madison, WI, USA, Crop Science Society of America.
