![]()
![]()
![]()
Nelly Delgado
Humberto Rodríguez
* This study was co-financed by INIA-Venezuela and Project S1200000771 of the Fondo Nacional de Ciencia, Tecnología e Innovación (FONACIT), formerly CONICIT, and was supported by the DANAC Foundation.

Nelly Delgado
Centro de Investigaciones Agropecuarias, Instituto Nacional de Investigaciones
Agrícolas (INIA), Apartado Postal 102, Acarigua, Estado Portuguesa, Venezuela.
E-mail: [email protected]
Abstract
Intensive cropping has increased the incidence and severity of rice sheath blight, caused by the fungus Rhizoctonia solani Kühn, in Venezuelan rice-producing areas. Only moderate resistance to this pathogen has been found in cultivated rice. Because of the complexity and difficulty of selecting for resistance to R. solani, yield potential and grain quality, recurrent selection is the methodology of choice. To select parental materials for the base population, various elite lines and commercial cultivars were evaluated under pressure from the fungus, and the least susceptible and most distantly related were selected. Other lines and cultivars were also selected for their yield potential, and good milling and culinary qualities, regardless of their reaction to the pathogen. To create a base population, the selected parents were then crossed with male-sterile plants of ‘IR36’. The F1 generation obtained was backcrossed with the recurrent parent (BC1), thus reducing the influence of ‘IR36’ and increasing the contribution of the selected parents. To create a multicytoplasmic population, pollen from each F1 hybrid was used to fertilize the recurrent parent. Finally, a complete diallel, without reciprocals, between 10 selected parents was made, using F2 seeds resulting from the selfing of each BC1. The selection strategy included an evaluation of resistance to R. solani in the S0 and S0:1 generations, and evaluations of yield and white belly at the S0:2 family level. We expect this methodology to help increase the odds of obtaining improved lines for these traits while exploiting the segregation occurring in each recurrent cycle.
Resumen
La incidencia y severidad del añublo de la vaina (Rhizoctonia solani Kühn) ha aumentado en las zonas productoras de arroz de Venezuela por el uso de sistemas intensivos de cultivo. El arroz cultivado posee solamente una resistencia moderada. Debido a la complejidad y dificultad de seleccionar para resistencia a R. solani, potencial de rendimiento y calidad de grano, se utilizará la selección recurrente. Para escoger los progenitores de la población base, se evaluaron diversas líneas élite y variedades comerciales sometidas a presión del hongo R. solani y se escogieron las menos susceptibles y las menos emparentadas entre sí. Como progenitores también se seleccionaron otras líneas y variedades comerciales con base en su potencial de rendimiento, calidades molinera y culinaria sin importar el tipo de reacción al patógeno. Para crear la población base se cruzaron los progenitores seleccionados con plantas androestériles del ‘IR36’. Luego se retrocruzaron la F1 obtenida con el progenitor recurrente (RC1), disminuyendo la influencia de ‘IR36’ y aumentando el aporte de los progenitores seleccionados. El polen de cada híbrido F1 se utilizó para polinizar el progenitor recurrente creando así una población poli-citoplasmática. Finalmente, para sintetizar la población base se realizó un dialelo completo y sin recíprocos entre los 10 progenitores seleccionados, utilizando la semilla F2 proveniente de la autopolinización de cada RC1. La estrategia de selección incluye la evaluación de la resistencia a R. solani en las generaciones S0 y S0:1 y la evaluación de rendimiento y centro blanco a nivel de familias S0:2. Con esta metodología se espera obtener una población mejorada que concentre los genes de resistencia a esta enfermedad, y los de buena calidad y alto rendimiento de grano. Así se incrementa la probabilidad de obtener líneas mejoradas al explorar la segregación presente en cada ciclo de recurrencia.
Rice sheath blight (ShB), caused by the fungus Rhizoctonia solani Kühn, is a disease of global importance. It causes heavy losses of yield in susceptible varieties cultivated in intensive production systems, particularly when infection occurs during the initiation of panicle growth, booting or initiation of flowering (Sharma and Teng, 1990; Rush and Lee, 1992; Cu et al., 1996).
In recent years in Venezuela, the incidence of ShB in rice-growing regions has increased through the use of cropping systems characterized by high planting density, abundant applications of nitrogenous fertilizers and intensive monocropping. Such farming practices, together with the pathogen’s saprophytic nature and wide host range, and the planting of susceptible varieties, have led to the dissemination, establishment and persistence of the fungus in all rice-producing areas (Nass et al., 1995).
The disease causes general weakness in the plant, which leads to its lodging. It also affects grain filling and weight, which negatively influences yield and milling quality (Cu et al., 1996; Rodríguez et al., 1999).
Evaluations conducted recently on rice farms in the Venezuelan states of Portuguesa, Guárico and Barinas have demonstrated that the incidence and severity of ShB have reached significant levels, giving rise to the need to revise strategies for reducing its effects on the crop’s productivity (Montealegre, 1993; Cedeño et al., 1998). An efficient, safe and economic strategy for the farmer is to use resistant or less susceptible varieties. The problem from the view of genetic improvement is that, so far, complete resistance to ShB has not yet been detected; only differences in reaction of some cultivars to the pathogen R. solani, and which reactions could be considered only as moderate resistance.
When combined with appropriate cultural practices, the use of this type of resistance reduces the pathological effects of the fungus on crop yield. Cutting down the density and viability of sclerotia in the soil will restrict their dissemination and incidence (Rush, 1983; Marchetti, 1991; Damicone et al., 1993).
This chapter presents the following aspects of our work in Venezuela:
The strategy we chose for developing a rice population with resistance to ShB without forgetting other traits important for the commercial acceptance of rice varieties
Stages for implementing the methodology of population improvement through recurrent selection
Expected outcomes of using this methodology to help solve the problem in farmers’ fields.
Before discussing the strategy and our choice of genetic improvement method to develop ShB-resistant germplasm, we will briefly discuss inheritance type and various factors involved in the crop’s resistance to R. solani. Some researchers have observed transgressive heredity in the crosses of certain varieties and have obtained recombinants or progeny that possess higher resistance than either parent. This suggests quantitative heredity (Marchetti, 1991; Groth and Nowick, 1992; Graterol et al., 1996). Other authors indicate that simple dominant genes control the intermediate resistance of some varieties. These genes segregate in an independent manner (not allelic), and could produce almost complete resistance on being reunited in a new variety (Pan et al., 1999).
Taking into account the number of factors involved in resistance in a cross of resistant with susceptible varieties, as many as six quantitative trait loci (QTLs) for resistance to R. solani have been found distributed throughout the genome. This fact supports the idea of genes for resistance as much from the resistant parent as from the susceptible (Li et al., 1995; Zou et al., 2000).
To work with complex traits-as the genetic control of ShB appears to be-some authors have suggested that rice population improvement is an efficient alternative (Fujimaki, 1979; Singh and Ikehashi, 1981; Châtel and Guimarães, 1998). Recurrent selection is a methodology of population improvement. It is designed to gradually and continually accumulate and increase the frequency of favourable genes for selected quantitative traits. This is done by frequently crossing superior individuals selected from segregating populations. The goals are to improve the population’s average performance, maintain genetic variation to continue selection and increase the probability of extracting improved populations and superior lines (Hull, 1945; Brim and Stuber, 1973; Poehlman and Sleper, 1996; Weyhrich et al., 1998). As such, it has been successively used in rice to improve oligogenic traits, as described by Borrero et al. (2000) for resistance to the rice ‘hoja blanca’ virus.
Until only very recently, the use of recurrent selection for population improvement was restricted to allogamous plants, partly because of the cost associated with manual emasculation and crossing among progeny of selected plants, and partly because of the low number of seeds produced by crossing (Brim and Stuber, 1973; Brim and Burton, 1979; Poehlman and Sleper, 1996). However, the presence of sterility in some autogamous species has provided an opportunity of adapting this method’s procedures to this type of crop (Brim and Stuber, 1973; Poehlman and Sleper, 1996; Châtel and Guimarães, 1998). In rice, genetic male sterility was produced and identified by Singh and Ikehashi in 1981, in a mutant ‘IR36’, which, in homozygous form, presented a recessive nuclear gene that produced sterility in pollen grains.
For this study, we chose population improvement through recurrent selection because the traits to improve - first, resistance to ShB and, second, yield potential, and milling and culinary qualities - are complex and difficult to select when using the classic methods of improving autogamous plants. The main objective is to create and accumulate in a population with a broad genetic base the major and/or minor genes of resistance to ShB from varieties or lines that are moderately resistant, as well as the genes for other traits that the selected lines may be carrying. Thus, to form the base population, parental materials with moderate resistance will be used, regardless of their potential for yield and/or milling and culinary qualities, with the expectation that they will carry genes favourable for this trait.
Moreover, we will seek parental materials that, although susceptible, will bring to the base population genes that are favourable for yield potential, and milling and culinary qualities. With this strategy, we expect to increase resistance to ShB in lines from each selection cycle, while guaranteeing the levels of grain quality and yield characteristic of a competitive commercial variety. Likewise, increase in resistance to the pathogen will contribute to stability of yield and grain quality as the stress provoked by the disease during grain filling is reduced.
When selecting parental materials for the base population, we took into account the following factors: their adaptation to the Venezuelan rice-producing regions, agronomic performance, yield, grain quality, the materials’ degree of resistance to ShB and relatedness between selected lines and cultivars. Such selection was carried out to reunite, within the population, favourable and complementary traits determined by genes of diverse origins.
To select parental materials that would bring genes for resistance to R. solani to the base population, 38 materials, including elite lines and varieties, were evaluated, using artificial inoculations of the fungus under conditions favourable for disease development. Two experiments for evaluation were conducted, one with 18 materials from commercial varieties and elite lines, and the other with 20 introductions. The variety Fundarroz PN-1, common to both experiments, was used as susceptible check. Of these materials, those presenting the least susceptibility to the pathogen in terms of lesion length, number of affected progeny per plant and number of affected sheaths were chosen.
Genes for adaptability, yield, and milling and culinary qualities would be brought to the population by commercial varieties and elite lines characterized over several years in commercial fields and experimental trials conducted in Venezuela. These varieties and elite lines were not selected for the type of reaction they showed later to inoculation with the pathogen R. solani, but for their pedigree, thus excluding from the population those materials closely related to each other or to progenitors likely to bring genes for resistance. These criteria were followed to obtain a population with a broad genetic base and with sufficient genetic variance to guarantee progress with selection, as proposed by Hallauer and Miranda (1995).
The gene for male sterility would be brought by ‘IR36’, developed at the International Rice Research Institute (IRRI), Philippines, and released in 1976. At least eight countries of Asia and Africa cultivated this variety in their rice-growing areas between 1977 and 1988 (Peng et al., 2000). This material presented a modern plant type with high yield, short cycle, good grain quality and resistance to the major insect pests and diseases of the rice crop (Peng et al., 2000). Its use in synthesizing irrigated rice populations has been beneficial, given that, besides possessing the male-sterility gene, it also introduces other agronomic traits favourable to populations developed for the irrigated ecosystem (Rangel et al., 2000).
Rice germplasm was evaluated in two separate experiments. In the first, commercial varieties with wide adaptability and good yield potential were evaluated, together with some elite lines that presented high yield potential and good milling and culinary qualities (Table 1). The second experiment involved the evaluation of lines and varieties from other countries (Table 1) introduced through the germplasm bank held at the Small Grains Collection of the United States Department of Agriculture (USDA), previously characterized for their reaction to R. solani in USA. Both experiments were carried out in the mesh-house of the Instituto Nacional de Investigaciones Agrícolas (INIA), State of Portuguesa, Venezuela.
The germplasm subjected to evaluation was planted in 25 cm high pots with 20 cm diameters. The pots were placed in a tank, filled to 10 cm deep with water. The tank’s height (1 m) was practical for inoculating and evaluating the materials. Each pot carried 5 plants arranged in the form of a cross. Sixty days after planting, the central plant in each pot was inoculated by using forceps to place a sclerotium on the main stem of the plant at the neck of the second basal leaf, and between the sheath and stem. Relative humidity was maintained at more than 70% during the critical hours, using a misting system to guarantee the micro-environment that would be favourable for disease development. The following evaluations were conducted:
Measuring lesion length in the inoculated and successive foliar sheaths and counting the number of sheaths affected by the disease
Ascertaining the number of lesions in the inoculated and successive foliar sheaths
Establishing the number of diseased tillers (NDT) in the inoculated plant
Determining the total number of tillers (TNT) in the inoculated plant
With the values of NDT and TNT, calculating the index of diseased tillers (IDT) according to the ratio NDT/TNT × 100, thus indicating the horizontal advance of the disease.
Table 1. Commercial varieties and elite lines of rice evaluated for their resistance to the fungus Rhizoctonia solani.
|
Material |
Origina |
Material |
Origina |
|
Materials already used in Venezuela |
|||
|
Araure 1 |
V |
Cimarrón |
V |
|
Araure 2 |
V |
Fundarroz PN-1 |
V |
|
Araure 3 |
V |
CT10184-2-1-M-1-MI (Danac-1) |
C |
|
Araure 4 |
V |
CNTBR82074-210-1-2-3 |
Br |
|
Fonaiap 1 |
V |
CT9807-3-5-1-2-M-MI-MC-1C |
C |
|
Fonaiap 2 |
V |
CNARR 4953-8B-BM70A-26-1P (Danac-4) |
Br |
|
Fonaiap 2000 |
V |
IR57514-PMI-5-8-1-2 (Danac-5) |
Ph |
|
Palmar |
V |
Oryzica 3 |
C |
|
Oryzica Llanos 5 |
C |
Barinas 1 |
V |
|
Materials introduced for evaluation under Venezuelan conditions |
|||
|
BR736-20-3-1 |
B |
P 5446-6-6-2-1 |
C |
|
Dal Kaisha |
B |
Ta Poo Cho Z |
Ch |
|
EMBRAPA Taim 7 |
Br |
Jefferson |
USA |
|
Fedearroz 50 |
C |
New Bonnet |
USA |
|
CT10179-13-5-M |
C |
Leah |
USA |
|
CT10195-52-1-M |
C |
Bluebelle |
USA |
|
CT10310-15-9-M |
C |
RU8703196 |
USA |
|
CT10323-3-2-M |
C |
Katy |
USA |
|
CT10323-8-2-M |
C |
B82-761 |
USA |
|
CT10323-18-3-M |
C |
San Martín 86 |
P |
a. Abbreviations are B for Bangladesh; Br for Brazil; C for Colombia; Ch for China; P for Peru; Ph for Philippines; and V for Venezuela.
Results from the first year of evaluation indicated that differences existed among the materials for their reaction to R. solani. Some presented moderate resistance to the pathogen and were chosen as parental materials carrying genes for resistance. The varieties and elite lines that were moderately resistant in terms of their scores for lesion length and number of lesions (data not presented) were the following:
Table 2. Means of severity (SEV) of sheath blight for 20 introductions of rice and the susceptible check (Fundarroz PN-1) evaluated for selection as parental materials for work on population improvement.
|
Introduction |
SEV |
Groupa |
Introduction |
SEV |
Groupa |
||
|
Susceptible: |
|
|
Susceptible (cont´d): |
|
|
||
| |
RU8703196 |
8.3 |
a |
|
Bluebelle |
6.0 |
abcd |
|
New Bonnet |
7.6 |
ab |
CT10323-8-2-M |
5.3 |
abcd |
||
|
Leah |
7.6 |
ab |
Fedearroz 50 |
5.0 |
bcd |
||
|
B82-761 |
7.6 |
ab |
CT10195-52-1-M |
4.6 |
bcd |
||
|
Dal Kaisha |
7.6 |
ab |
CT10323-18-3-M |
4.6 |
bcd |
||
|
Ta Poo Cho Z |
7.5 |
abc |
CT10310-15-9-M |
4.5 |
cd |
||
|
Katy |
7.3 |
abc |
|
|
|
||
|
BR736-20-3-1 |
6.8 |
abcd |
Moderately resistant: |
|
|
||
|
CT10179-13-5-M |
6.5 |
abcd |
|
P 5446-6-6-2-1 |
4.0 |
de |
|
|
San Martín 86 |
6.0 |
abcd |
CT10323-3-2-M |
3.8 |
de |
||
|
EMBRAPA Taim 7 |
6.0 |
abcd |
Jefferson |
1.3 |
e |
||
|
Susceptible check: |
|
|
|
|
|
||
| |
Fundarroz PN-1 |
5.3 |
abcd |
|
|
|
|
|
Average mean |
5.9 |
|
|
|
|
||
a. LSD = 3.0
Likewise, the introductions that had moderate resistance in terms of their values for severity (Table 2) were, in descending order:
Varieties Palmar and Oryzica 3, and Fonaiap 2 and Oryzica Llanos 5, sister lines deriving from the same cross, performed similarly in their reaction to ShB, which indicates that they must have identical genes for resistance by descendency. Hence, of the two pairs of sister lines only those that had been commercial varieties recommended for Venezuelan conditions were selected, that is, Fonaiap 2 and Palmar. Despite these two selected varieties sharing CICA 7 as a common ancestor, both will be used to obtain the base population because Palmar also offers genes for milling and culinary qualities that Fonaiap 2 does not have. It should be noted that their genes differ from those carried by D-Sativa (Table 3).
Table 3. Parents and grandparents of rice varieties and elite lines selected to form part of the base population.
|
Variety or line |
Parents |
Grandparents |
|
Fonaiap 1 |
P 1386/Camponi/Tapuripa |
P 1221; P 1238 |
|
Fonaiap 2 |
P 5003/P 1274//P 2060 |
Colombia 1; P 1274; P 1217; |
|
Palmar |
CICA 7//CICA 8/Pelita I-1 |
IR22; P 809; CICA 4; IR665; Tetep |
|
Cimarrón |
HEBI G11330// |
IR579; IR747 |
|
CT10184-2-1-M-1-MI (Danac-1) |
CT8154//CT7415/P 4743 |
P 5601; P 4711; CT6889; |
|
CNARR 4953-8B-BM70A-26-1P (Danac-4) |
IR13146/IR15314// |
IR10776; IR46; IR11738; |
|
CT10323-3-2-M |
P 3084/P 3844//CT8154 |
P 1491; P 2624; Oryzica 1; |
|
IR57514-PMI-5-8-1-2 |
IR21836//IR43581 |
IR18197; IR4570; IR41289; |
|
Jefferson |
B82-761/Rosemont |
Vista; Lebonnet; CI9881; |
|
D-Primera |
IR2823/IR5533//IR43 |
CR 94; IR1529; IR1818; |
|
D-Sativa |
P 3050/Oryzica 1//IR21015 |
BG90-2; P 2672; Batatais; |
|
IR36 |
IR1561/IR1737//CR 94 |
IR579; IR747; IR24; IR1721; |
The variety Jefferson and lines:
CT10184-2-1-M-1-MI (Danac-1)
CNARR 4953-8B-BM70A-26-1P (Danac-4)
IR57514-PMI-5-8-1-2 (Danac-5)
CT10323-3-2-M
were selected for their resistance to ShB. Except for CT10323-3-2-M, none had ancestors in common with the rest of the selected materials.
CT10323-3-2-M shared one ancestor with D-Sativa (Oryzica 1) and another with CT10184-2-1-M-1-MI (CT8154) (Table 3). The latter, however, was the most resistant after cv. Jefferson, with an average score for severity at 3.8. It contributed half of the genetic conformation of both lines, and may be the source of resistance to ShB, given that these lines do not share progenitors in the other half of their genetic constitution. Even so, we decided to include both in forming the base population. From line CT10323-3-2-M, the four most resistant plants were selected and, a mass selection was obtained with the same number of seeds from each plant.
Table 4. Rice varieties and lines selected as parental materials for the base population.
|
Variety or line |
Criterion for selectiona |
|
Fonaiap 1 |
Yield potential, resistance to blast, culinary qualities |
|
Cimarrón |
Yield potential |
|
D-Primera |
Yield potential, resistance to blast |
|
D-Sativa |
Yield potential, milling and culinary qualities, resistance to blast |
|
Palmar |
Resistance to ShB, milling and culinary qualities |
|
Fonaiap 2 |
Resistance to ShB, resistance to blast |
|
Jefferson |
Resistance to ShB, milling and culinary qualities |
|
CT10323 |
Resistance to ShB |
|
CT10184 |
Resistance to ShB |
|
IR57514 |
Resistance to ShB |
|
IR36 |
Donor of the male-sterility gene, resistance to blast |
a. ShB refers to rice sheath blight.
The seed will be used in crosses to obtain the base population.
The materials selected as donors of genes for yield and/or milling and culinary qualities were Fonaiap 1, Cimarrón, D-Primera, D-Sativa and Palmar (Table 4). These varieties were not related to each other at the grandparent level and will contribute in broadening the base population’s genetic base. Moreover, they will bring genes for adaptability to Venezuelan agroclimatic conditions, particularly Cimarrón and Fonaiap 1, which have been widely planted for 14 and 10 years, respectively, in Venezuela’s rice-producing zones.
In summary, 10 materials were selected as parental materials for the base population. Eight corresponded to varieties or elite lines, previously evaluated for their reaction to R. solani. The other two (D-Primera and D-Sativa), whose reaction to the pathogen was unknown, were selected for the potential contribution of their genes to yield and quality. So that we would know the reaction of all the selected materials to this pathogen, we are currently evaluating the last two varieties.
At the end of this project, we expect to have a population with sufficient genetic variability to concentrate diverse genes for resistance to R. solani, yield, and milling and culinary qualities. Such variability should permit genetic gain with selection and thus the acquisition of improved populations and lines for the desired traits.
Different ways exist to synthesize a base population that will be subjected to population improvement through recurrent selection. The process of creating is characterized by a series of crosses between the parental materials and the source of the male-sterility gene, followed by some backcrosses to reduce the contribution of the male-sterility source (Rangel et al., 2000). The strategy to be used in creating our base population is presented in Figure 1. The selected varieties or lines will be crossed with male-sterile plants of ‘IR36’ to obtain an F1 generation that will be backcrossed (BC1) with the recurrent parent (variety or line) to increase its contribution to the population. Each F1 will be used as a male parent, with the variety or line as the female parent or pollen receiver, thus diversifying the source of cytoplasm (Brim and Stuber, 1973; Borrero et al., 1997; Rangel and Neves, 1997). The BC1 seeds obtained will be multiplied by selfing and the BC1S1 seeds will be used in recombination.
Figure 1. Steps for creating a rice base population for improving through recurrent selection. Each step is carried out over one semester, that is, 2 years are needed to complete one recurrent selection cycle.
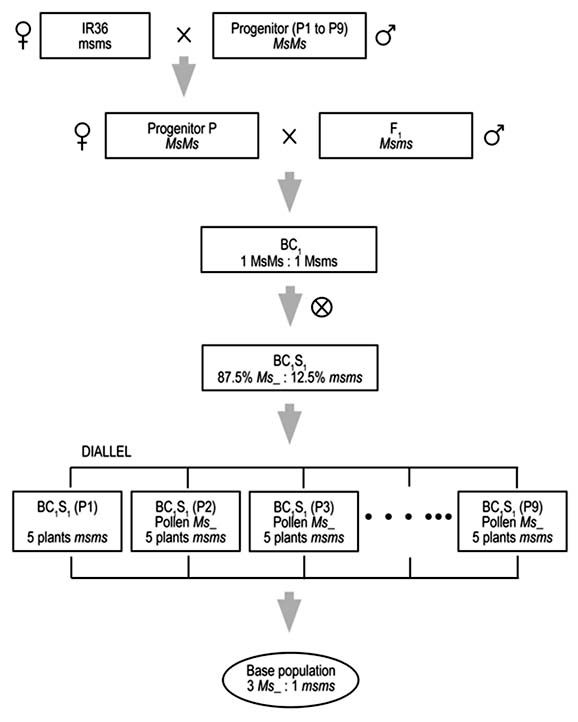
Table 5. Timetable for obtaining the rice base population INDA-Rs 1\0\0\0 carrying genes for resistance to the fungus Rhizoctonia solani.
|
Activity |
Period of execution |
|||
|
Semester II 2002 |
Semester I 2003 |
Semester II 2003 |
Semester I 2004 |
|
|
Obtaining F1 |
X |
|
|
|
|
Obtaining BC1 |
|
X |
|
|
|
Obtaining BC1S1 |
|
|
X |
|
|
Synthesis of the base population (diallel) |
|
|
|
X |
Some authors suggest that several cycles of recombination be carried out before beginning selection to break the linkage blocks and negative genetic associations between traits (Meredith and Bridge, 1971; Brim and Stuber, 1973; Fehr, 1987). Other authors, however, suggest that allelic recombination does not increase with the number of cycles of intra-population crosses. They observe that the most productive lines of the advanced crosses in wheat and soybean are similar in yield to the best-performing lines derived from populations with only one cycle of recombination (Altman and Busch, 1984; Guimarães and Fehr, 1989).
Likewise, in rice, no changes in genetic variability occurred on increasing the recombination cycles. Hence, it is not necessary to go further than one cycle of recombination before beginning selection (Ospina et al., 1997; Chapter 17, this volume). In this study, only one cycle of recombination will be made before initiating population improvement. Accordingly, the cross will be made, using a complete diallel without reciprocals, of all the BC1S1 (10 × 9), which will give rise to 45 crosses. From these, seed will be taken in equal proportions to create the population INDA-Rs 1\0\0\0. The manual crosses will be made in three blocks of 15 BC1S1 each, planting two rows, each 10 m long, for each BC1S1. The sterile plants of one BC1S1 will receive pollen from fertile plants of the remaining BC1S1. The timetable for obtaining the base population is given in Table 5.
Figure 2. Steps of the planned recurrent selection for resistance to the fungus Rhizoctonia solani. Each step is carried out in one semester, that is, 2 years are needed to complete one recurrent selection cycle.
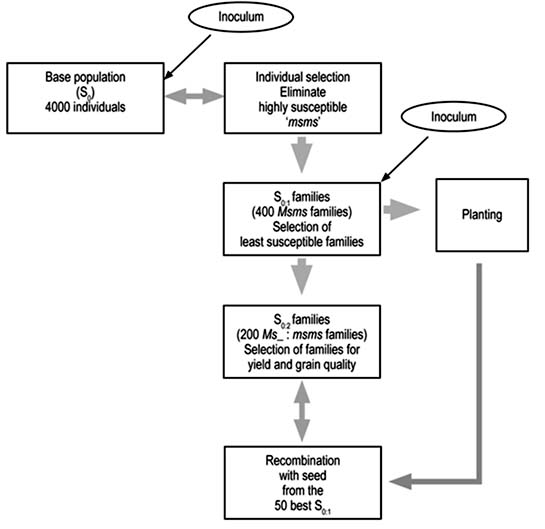
To preserve the genetic properties of the base and improved populations, a sample of at least 200 plants will be taken, as recommended by Geraldi and Souza (2000). The strategy of recurrent selection that will be used is described below and illustrated in Figure 2.
Male-sterile plants that are highly susceptible will be eliminated from the base population (S0) while the rest will be harvested. Inoculation will be done 60 days after planting by broadcasting 25 cm3 of fungus-infested rice grains per square metre of soil. This method was chosen on the basis of experimental results obtained in plots of irrigated rice in the experimental field of INIA-Portuguesa, carried out in parallel with the germplasm evaluation already discussed. The objective was to obtain a useful method of inoculation for evaluating and selecting plants or families in recurrent selection populations while minimizing the risk of escapes. With this inoculation methodology - which, moreover, was very practical - 94% of the inoculated plants showed typical symptoms of the disease 35 days after inoculation, that is, 5% more incidence than for inoculations made with sclerotia.
In the S0:1 generation, the seed of plants selected from the base population (S0) will be multiplied and, at the same time, the incidence and vertical advance of the disease will be evaluated to select those that have the least frequency of highly susceptible individuals. One part of the seed from the selected S0:1 families will be kept for later recombination, and the other part will be used to evaluate S0:2 families.
The S0:2 families will be planted in replicated experiments to evaluate their yield, and milling and culinary qualities, which are traits of least heritability and whose genetic gain with selection increases by using families constituted by the most closely related individuals (Morais, 1997).
Sheath blight appears in the crop at about 60 days after planting, thus permitting selection in both sexes before flowering. Moreover, as transplanting selected plants to crossing blocks in this phase of the cycle could limit seed production, the strategy of using selected pollen over selected male-sterile plants would imply a greater use of resources and could be impractical. With the proposed scheme, mass selection would be done only for the female parent, as the source of pollen is not controlled.
However, with the aim of increasing genetic gain with selection, advantage would be taken of the generation for seed multiplication (S0:1) to make a new evaluation of resistance to R. solani. This evaluation would not be postponed to S0:2 because the presence of disease would interfere with the other objectives of selection, such as yield, and milling and culinary qualities. These traits would not be able to express themselves in their best environment, that is, free of the pathogen, and would thus reduce capacity to select.
Resistance to R. solani is a complex trait that is affected by both environment and low heritability. Although complete resistance to this pathogen has not been found, partial resistance has. By using population improvement through recurrent selection as a methodology of genetic improvement for this trait, we expect to concentrate favourable genes from different sources of a population. We therefore should increase the possibility of obtaining or deriving, after some cycles of population improvement, lines that are less susceptible than those currently existing on the market.
We do not know of any study that has evaluated the existence or absence of negative correlations between resistance to R. solani and traits such as yield and grain quality. Such studies may not exist because varieties such as Palmar and Jefferson combine excellent grain quality and good yield with least susceptibility to this pathogen. We therefore expect to be able to combine this resistance with traits of high yield potential and good grain quality, depending on these correlations to ensure that all traits of interest are obtained.
The genetic gain that could be obtained with selection cycles in terms of reaction to R. solani will depend principally on the skill of managing the selection environment and the selection of genuinely less susceptible individuals (genotypes). The first aspect can be managed to a certain point by subjecting the populations to artificial inoculations during the rainy season under conditions of high population density and good supplies of nitrogenous fertilizer.
For the second aspect, appropriate management requires knowing the dosage and form of application of inoculum to guarantee the best sampling of population and thus prevent the escape of plants that become included in the improved population for not showing symptoms of the disease but, if they had been inoculated, would have shown moderate susceptibility. In this sense, previous data from a trial evaluating inoculation methods will be validated in a second opportunity before beginning the first recurrent selection cycle.
Success in being able to combine resistance to R. solani with high yield potential and good grain quality will depend on the efficiency of selection in tandem. As already mentioned, we propose to select for resistance at the level of the base (S0) and S0:1 populations for grain quality and yield in S0:2. Selection will be more successful if no negative correlation exists between the said traits.
With adequate management of these aspects and with the security of relying on diverse genes for resistance, as well as on genes for high yield potential and grain quality that are not correlated with the first, we expect good progress with selection in the next few years.
References
Altman, D.W. & Busch, R.H. 1984. Random intermating before selection in spring wheat. Crop Sci., 24: 1085-1089.
Borrero, J.; Châtel, M. & Triana, M. 2000. Mejoramiento poblacional del arroz irrigado con énfasis en el virus de la hoja blanca. In E.P. Guimarães, ed. Avances en el mejoramiento poblacional del arroz, pp. 105-118. San Antônio de Goiás, Brazil, Embrapa Arroz e Feijão.
Borrero, J.; Ospina, Y.; Guimarães, E.P. & Châtel, M. 1997. Ampliación de la base genética de los acervos de arroz, mediante la introducción de variabilidad. In E.P. Guimarães, ed. Selección recurrente en arroz, pp. 55-66. Cali, Colombia, CIAT.
Brim, C.A. & Burton, J.W. 1979. Recurrent selection in soybeans, II: Selection for increased percent protein in seeds. Crop Sci., 19: 494-498.
Brim, C.A. & Stuber, C.W. 1973. Application of genetic male sterility to recurrent selection schemes in soybeans. Crop Sci., 13: 528-530.
Cedeño, L.; Nass, H.; Carrero, C.; Cardona, R.; Rodríguez, H. & Alemán, L. 1998. Rhizoctonia oryzae-sativae agente causal de la mancha agregada del arroz en Venezuela. Interciencia, 24(4): 248-251.
Châtel, M. & Guimarães, E.P. 1998. Selección recurrente con androesterilidad en arroz. Cali, Colombia, CIRAD-CA & CIAT. 70 pp.
Cu, R.M.; Mew, T.W.; Cassman, K.G. & Teng, R.S. 1996. Effect of sheath blight on yield in tropical, intensive rice production system. Plant Dis., 80: 1103-1108.
Damicone, J.P.; Patel, M.V. & Moore, W.F. 1993. Density of sclerotia of Rhizoctonia solani and incidence of sheath blight on rice fields in Mississippi. Plant Dis., 77: 257-260.
Fehr, W.R. 1987. Principles of cultivar development, Vol. 1: Theory and technique. Ames, IA, Iowa State University Press. 536 pp. (1993 reprint)
Fujimaki, H. 1979. Recurrent selection by using genetic male sterility for rice improvement. Jpn. Agric. Res. Q., 13(3): 153-156.
Geraldi, I.O. & Souza, Jr, C.L. de. 2000. Muestreo genético para programas de mejoramiento poblacional. In E.P. Guimarães, ed. Avances en el mejoramiento poblacional en arroz, pp. 9-20. San Antônio de Goiás, Brazil, Embrapa Arroz e Feijão.
Graterol, E.; Borges, O.; Nass, H. & Salih, A. 1996. Herencia transgresiva para la resistencia a Rhizoctonia spp. en poblaciones segregantes de arroz (Oryza sativa L.). Investigación agrícola. Vol. 1. 8 pp. (available at http://www.redpavfpolar.info.ve/danac/volumen1/art3/index.html).
Groth, D.E. & Nowick, E.M. 1992. Selection for resistance to sheath blight through number of infection cushions and lesion type. Plant Dis., 76: 721-723.
Guimarães, E.P. & Fehr, W.R. 1989. Alternative strategies of recurrent selection for seed yield of soybean. Euphytica, 40: 111-120.
Hallauer, A.R. & Miranda, J.B. 1995. Quantitative genetics in maize breeding. 2nd ed. Ames, IA, Iowa State University Press. 468 pp.
Hull, F. 1945. Recurrent selection for specific combining ability in corn. J. Am. Soc. Agron., 37(2): 134-145.
Li, Z.K.; Pinson, S.R.M; Marchetti, M.A.; Stansel, J.W. & Park, W.D. 1995. Characterization of quantitative trait loci (QTLs) in cultivated rice contributing to field resistance to sheath blight (Rhizoctonia solani). Theor. Appl. Genet., 91: 382-388.
Marchetti, M.A. 1991. Quantification of the relationship between sheath blight severity and yield loss in rice. Plant Dis., 75: 773-775.
Meredith, W.R., Jr & Bridge, R.R. 1971. Breakup of linkage blocks in cotton, Gossypium hirsutum L. Crop Sci., 11: 695-698.
Montealegre, F.A. 1993. Presencia del añublo de la vaina en Colombia y el exterior. Arroz, 42: 34-48.
Morais, O.P. de. 1997. Tamaño efectivo de la población. In E.P. Guimarães, ed. Selección recurrente en arroz, pp. 25-44. Cali, Colombia, CIAT.
Nass, H.A.; Cedeño, L.; Carrero, C.; Cardona, R.; Rodríguez, H. & Alemán, L. 1995. Rhizoctonia solani AG1-IA importante patógeno del arroz (Oryza sativa) en Venezuela. Rev. Forest. Venez., 1: 142-143. (Abstract.)
Ospina, Y.; Borrero, J.; Guimarães, E.P. & Châtel, M. 1997. Ciclos de intercruzamiento y variabilidad genética en poblaciones de arroz. In E.P. Guimarães, ed. Selección recurrente en arroz, pp. 45-53. Cali, Colombia, CIAT.
Pan, X.B.; Rush, M.C.; Sha, X.Y.; Xie, Q.J.; Liscombe, S.D.; Stenina, S.R. & Orad, J.H. 1999. Major gene non-allelic sheath blight resistance from the rice cultivars Jasmine 85 and Teqing. Crop Sci., 39: 338-346.
Peng, S.; Laza, R.C.; Visperas, R.M.; Sanico, A.L.; Cassman, K.G. & Khush, G.S. 2000. Grain yield of rice cultivars and lines developed in the Philippines since 1966. Crop Sci., 40: 307-314.
Poehlman, J.M. & Sleper, D.A. 1996. Breeding field crops. 4th ed. Ames, IA, Iowa State University Press. 494 pp.
Rangel, P.H.N. & Neves, P.C.F. 1997. Selección recurrente aplicada al arroz de riego en Brasil. In E.P. Guimarães, ed. Selección recurrente en arroz, pp. 79-97. Cali, Colombia, CIAT.
Rangel, P.H.N.; Zimmermann, F.J.P. & Fagundes, P.R.R. 2000. Mejoramiento poblacional del arroz de riego en Brasil. In E.P. Guimarães, ed. Avances en el mejoramiento poblacional del arroz, pp. 65-86. San Antônio de Goiás, Brazil, Embrapa Arroz e Feijão.
Rodríguez, H.; Nass, H.; Cardona, R. & Alemán, L. 1999. Alternativas para controlar el añublo de la vaina causado por Rhizoctonia solani en arroz. Fitopatol. Venez., 12(1): 18-21.
Rush, M.C. 1983. Rice sheath blight: a major rice disease. Plant Dis., 67(7): 829-832.
Rush, M.C. & Lee, F.N. 1992. Sheath blight. In F. Webster & P. Gunnell, eds. Compendium of rice diseases, pp. 22-23. St. Paul, MN, USA, APS Press.
Sharma, N.R. & Teng, P.S. 1990. Effect of rice growth stage on sheath blight (ShB) development and yield loss. Int. Rice Res. Newsl., 15(6): 20.
Singh, R.J. & Ikehashi, H.I. 1981. Monogenic male sterility in rice: induction, identification and inheritance. Crop Sci., 21(2): 286-289.
Weyhrich, R.A.; Lamkey, K.R. & Hallauer, A.R. 1998. Effective population size and response to S1-progeny selection in the BS11 maize population. Crop Sci., 38: 1149-1158.
Zou, J.H.; Pan, X.B.; Chen, Z.X.; Xu, J.Y.; Lu, J.F.; Zhai, W.X. & Zhu, L.H. 2000. Mapping quantitative trait loci controlling sheath blight resistance in two rice cultivars (Oryza sativa L.). Theor. Appl. Genet., 101(4): 569-573.
![]()
![]()
![]()