![]()
![]()
![]()
METHODOLOGIES
Ideally, impact studies should be conducted in areas that have been subjected to trawling on a spatial and temporal scale that is representative of commercial fishing activities. Quantitative sampling in replicate control sites is also a crucial prerequisite to allow firm conclusions.
The ideal impact study should be designed with the aim of detecting benthic habitat and community changes that can be attributed unambiguously to fishing disturbance on a spatial and temporal scale that is representative of commercial fishing activity. To date it has been difficult to conduct studies that fulfil these requirements (Kaiser et al., 2000), and most studies have been carried out within small experimental areas that have been trawled over a relatively short period. The potential effects in narrowly trawled corridors can be masked by rapid recolonization by organisms from adjacent untouched areas, or on the other hand disturbance effects may be enhanced by abundant scavengers feeding in a small area of exposed benthic fauna. The time aspect is also an important factor to consider, because changes relevant to the temporal scale of commercial fishing will not have evolved in samples taken immediately after a relatively short period of experimental trawling. Potential long-term impacts that may arise in areas subjected to commercial trawling could be more serious to the function of benthic communities than immediate effects.
Sampling in replicate control sites has been raised as a crucial prerequisite for impact studies because changes in community structures demonstrated in the area trawled cannot be attributed to trawling disturbance unless more than one control site is monitored (Hurlbert, 1984; Underwood, 1992; Lindegarth et al., 2000b). The potential consequences of insufficient replication on the interpretation of impact experiments are demonstrated in the study by Lindegarth et al. (2000b) in which significant differences between pairs of control sites were revealed. However, conclusions from studies on short-term effects may be less affected by this type of confounding because the abundance of benthic animals may not show natural variability at these temporal scales.
Strictly quantitative sampling tools have not been used in all studies on trawl impacts, and even the use of quantitative tools (corers, grabs, improved sledges) does not ensure fully comparable samples. Trawling has been shown to cause sediment resuspension and changes in surface sediment characteristics (e.g. topography and hardness; see the chapter on Physical impacts), thus such changes may affect the efficiency of benthic sampling devices (Currie and Parry, 1996; 1999; Prena et al., 1999; Kutti et al., in press.). Increased abundance of sedentary organisms shortly after trawling has been demonstrated (e.g. Eleftheriou and Robertson, 1992; Kaiser et al., 1998; Ball, Fox and Munday, 2000), and can be explained only by an increase in the efficiency of the sampling device. The catchability of a sampling sledge may also decrease owing to the burial of organisms by resuspended sediment (Prena et al., 1999).
The use of semi-quantitative sampling devices and changes in sampling efficiency after experimental trawling may introduce serious artefacts that ought to be considered when the results of impact studies are being interpreted. Furthermore, inappropriate sampling may increase the variability among samples, and thus decrease statistical power to detect true effects of trawling disturbance.
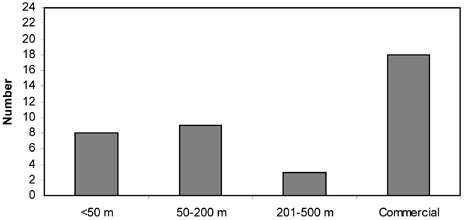
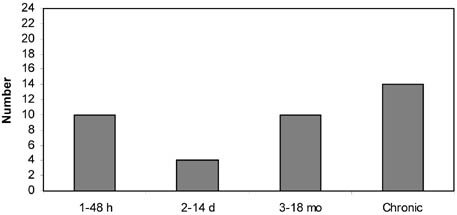
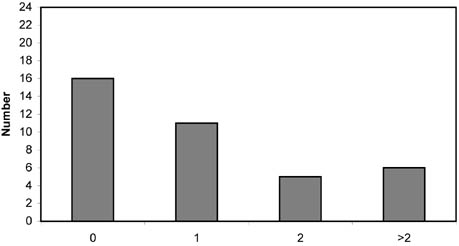
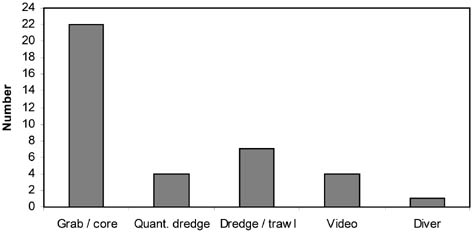
This chapter addresses three important methodological requirements for studying the impacts of trawling: disturbance at a spatial and temporal scale that is representative of commercial fishing, replicate controls, and quantitative sampling. Because the resources available (e.g. vessel time and labour cost for sample analyses) are limited, compromises have to be made, and most impact studies thus fail to meet one or more of these requirements (Table 5; Figure 8). These and other potential deficiencies have to be taken into consideration before firm conclusions can be drawn. Although the investigators of these studies often recognized that their experimental designs had limitations, less caution is seen in review papers on this topic.
|
FIGURE 8 Frequency distributions of impact studies at different spatial scales (width of trawled area), temporal scales (time between experimental trawling and sampling), numbers of control sites and types of sampling tools Spatial scale
Temporal scale
Control sites
Sampling tools
|
TABLE 5
Characteristics and design of studies to
investigate the impacts of trawling and dredging on benthic habitats and
communities
|
References |
Spatial scale |
Temporal scale |
Control sites |
Bottom mapping |
Quantitative sampling |
Trawling intensity |
Gear description |
|
Prena et al., 1999 |
81-264 m |
1-93 h |
2 |
Yes |
Yes |
12 tows |
Yes |
|
Kenchington et al., 2001 |
81-264 m |
8-132 h and 1 yr |
2 |
Yes |
Yes |
12 tows |
Yes |
|
Kutti et al., in press. |
200 m |
15-29 h |
No |
Yes |
Yes |
10 tows |
Yes |
|
McConnaughey, Mier & Dew, 2000 |
1 nm² |
Chronic |
42 |
No |
No |
No data |
No |
|
Engel & Kvitek, 1998 |
22 km² |
Chronic |
No |
No |
Yes |
4 times |
No |
|
Tuck et al., 1998 |
250 m |
6-18 mo |
1* |
Yes |
Yes |
16 (10 tows/mo) |
No |
|
Hansson et al., 2000 |
100 m |
7-10 mo |
3 |
No |
Yes |
40 (2 tows/wk) |
Yes |
|
Drabsch, Tanner & Connell, 2001 |
200 m |
1 wk |
3 |
No |
Yes |
2 times |
Yes |
|
Sparks-McConkey & Watling, 2001 |
A few metres |
0-6 mo |
1 |
No |
Yes |
4 times |
No |
|
Van Dolah, Wendt & Levisen, 1991 |
Commercial |
5 mo |
1 |
No |
Yes |
No data |
No |
|
Frid, Clark & Hall, 1999 |
Commercial |
Chronic |
No |
No |
Yes |
No data |
No |
|
Ball, Fox & Munday, 2000 |
40 m |
24 h |
No |
No |
Yes |
2 tows |
Partly |
|
Ball, Fox & Munday, 2000 |
Commercial |
Chronic |
Wreck |
No |
Yes |
No data |
No |
|
Hall et al., 1993 |
Commercial |
Chronic |
Wreck |
No |
Yes |
No data |
No |
|
Smith, Papadopoulou & Diliberto, 2000 |
500 m |
8 mo |
2* |
No |
Yes |
No data |
No |
|
Sanchez et al., 2000 |
150 m |
1-150 h |
2 |
No |
Yes |
1 and 2 tows |
No |
|
Simboura et al., 1998 |
Commercial |
Chronic |
1* |
No |
Yes |
No data |
No |
|
Freese et al., 1999 |
5 m |
2 h-5 d |
8 |
No |
Video |
1 tow |
Yes |
|
Moran & Stephenson, 2000 |
360 x 925 m |
1 d |
1 |
No |
Video |
4 times |
No |
|
Van Dolah, Wendt & Nicholson, 1987 |
30-35 m |
0 and 12 mo |
5 |
Divers |
Yes |
1 tow |
Yes |
|
Bergman & Hup, 1992 |
200 x 200 m |
16 h and 2 wk |
No |
No |
Yes |
2 times |
Yes |
|
Bergman & Santbrink, 2000 |
60 x 2000 m |
24-48 h |
No |
No |
Yes |
1.5 times |
Yes |
|
Kaiser & Spencer, 1996 |
30-40 m |
12 h |
3 |
Yes |
Yes |
10 and 20 times |
Yes |
|
Kaiser et al., 1998 |
30-40 m |
24 h and 6 mo |
4 |
Yes |
Yes |
10 and 20 times |
Yes |
|
Jennings et al., 2001 |
Commercial |
Chronic |
No |
No |
No |
Rel. indices |
No |
|
Currie & Parry, 1996 |
600 x 600 m |
0-14 mo |
1 |
No |
Yes |
2 times |
Yes |
|
Currie & Parry, 1999 |
600 x 600 m |
0-8 mo |
1 |
No |
No/Yes |
2 and 4 times |
Yes |
|
Thrush et al., 1995 |
20 x 35 m |
2 h and 3 mo |
1 |
Yes |
Yes |
1 time |
Yes |
|
Hall-Spencer et al., 1999 |
60 m |
15 h |
No |
Yes |
Video |
7 times |
Yes |
|
Pranovi et al., 2000 |
3 m |
1 and 7 d |
1 |
Yes |
Yes |
1 tow |
Yes |
|
Eleftheriou & Robertson, 1992 |
A few metres |
1 d |
1 |
Yes |
Yes |
2-25 tows |
Yes |
|
Hoffmann & Dolmer, 2000 |
Commercial |
Chronic |
2* |
No |
Yes |
No data |
No |
|
Kaiser et al., 2000 |
Commercial |
Chronic |
No |
No |
No |
Rel. indices |
No |
|
Kaiser, Spencer & Hart, 2000 |
Commercial |
Chronic |
No |
Yes |
No |
No data |
No |
|
Thrush et al., 1998 |
Commercial |
Chronic |
No |
Yes |
Yes |
No data |
Yes |
|
Collie, Escanero & Valentine, 1997 |
Commercial |
Chronic |
No |
Yes |
No |
No data |
No |
|
Collie, Escanero & Valentine, 2000 |
Commercial |
Chronic |
No |
Yes |
Video |
No data |
No |
|
Hill et al., 1999 |
Commercial |
Chronic |
No |
No |
No |
Rel. indices |
No |
* Differences between the control sites or between the control site and the trawled site before the experimental trawling, i.e. not true controls.
Spatial scale: the size or width of the trawled area.
Temporal scale: the time between experimental trawling and sampling; "chronic" means that grounds subjected to commercial fishing have been investigated.
Quantitative sampling: core, grab, camera-equipped dredges and counting by divers are regarded as quantitative.
Trawling intensity: the number of tows along the experimental corridor, or the estimated number of times that the area has been touched by the trawl.
Flattening of the bottom topography is the main physical impact of beam trawling and scallop dredging, whereas the doors of otter trawls create furrows. The consequences of these physical changes for the benthic community are poorly understood, and physical impacts should be determined at a fine scale of resolution that is relevant to benthic organisms.
The chains of a beam trawl and the teeth of a scallop dredge are designed to penetrate the upper few centimetres of the sediment, and these parts cover the whole width of the gear. The trawl doors are the only part of an otter trawl that is rigged to penetrate into the sediment (with the exception of a few types of otter trawls rigged with chains). As a result of these differences in the rigging and catching principle, the physical impacts on the sea bed caused by otter trawling are likely to be different from those caused by beam trawling and scallop dredging. As the latter two gear types penetrate into the sediment, the most conspicuous physical impact is flattening of bottom features such as ripples and irregular topography (e.g. Thrush et al., 1995; Kaiser et al., 1996). In addition, features such as bioturbation mounds and polychaete tubes are shown to be eliminated in the tracks of beam trawls and scallop dredges (Currie and Parry, 1996; Hall-Spencer et al., 1999).
The ecological impacts of eliminating natural bottom features on the benthic community are not clear and have not been adequately addressed in the studies on trawling disturbance published to date.
Furrows and berms created by the trawl doors are the most conspicuous physical impacts from otter trawls. The trawl doors probably penetrate deeper into the sediments than scallop dredges and beam trawls, and create an irregular bottom topography rather than flattening natural features.
The area disturbed by the trawl doors comprises only a small proportion of the total area swept by the trawl, e.g. door spreads of 60 m and 140 m, respectively, for the cod trawls used in the studies by Prena et al. (1999) and Kutti et al. (in press). Because no or only faint marks are created by the other parts of an otter trawl, the physical impacts on the sea bed are likely to be marginal in most otter trawl fisheries. An exception may be intensively trawled fishing grounds in sheltered areas or in deep water, where trawl marks may last for a long time. In such areas, trawl furrows may accumulate during the fishing season and create long-lasting, irregular bottom topography.
Thus, the main physical impact of beam trawling and scallop dredging seems to be a flattening of the bottom topography, whereas the doors of otter trawls create irregular features on the sea bed. The longevity of these effects is determined by sediment type and natural disturbances (tidal current, wave actions and biological activities), and has been shown to last from a few hours to more than a year. However, data are too scarce to allow a clear relationship between persistence of trawl marks and bottom type/natural disturbance to be made. The consequences of physical disturbance of the sea bed topography for the benthic community structure are poorly understood, and have not been subject to much investigation (Gislason, 1994).
It should be noted however that the tools and methods used to determine physical impacts (RoxAnn, side-scan sonar, video) are rough and crude ways of describing sea bed characteristics. Schwinghamer, Guigné and Siu (1996), using very high-resolution acoustics, were able to determine small-scale structural changes in the upper 4.5 cm of the sediment at a scale of resolution that is relevant to the benthic biota. This is the scale at which the physical impacts of trawling should be investigated.
It is difficult to conduct impact studies leading to clear and unambiguous conclusions because knowledge of the complexity and natural variability of benthic communities is rudimentary.
Several studies on the impacts of trawling on benthic communities state that trawling is the most disruptive and widespread anthropogenic disturbance on benthic habitats and may substantially alter benthic communities (e.g. Rumohr and Krost, 1991; Watling and Norse, 1998; Koslow et al., 2001). This review has taken into account the fact that several studies of this topic have serious weaknesses and deficits, and has shown that the evidence for such statements is not well-documented or convincing. Other authors have stated that the impacts of trawling on benthic communities have been poorly studied and that such research has been limited to a few regions (Freese et al., 1999; Smith, Papadopoulou and Diliberto, 2000). Furthermore, it has been concluded that few of the impacts of fishing have been well-documented (Currie and Parry, 1996), and that it has been difficult to attribute changes in the benthic community to fishing effort at a spatial scale that is representative of commercial fishing activities (Kaiser et al., 2000). Benthic communities are complex and show large temporal and spatial variability. It is therefore difficult to conduct impact studies that give clear and unambiguous results, and general statements such as the one cited above should be avoided. This does not mean that trawling causes only subtle effects on benthic communities, but simply that it is difficult to demonstrate such impacts because knowledge of these complex communities is rather scarce, and effects may often be minor compared with the natural variability in space and time.
The complexity and natural variability of benthic communities raise two important issues related to the conclusions that can be drawn from trawl impact studies. Where effects have been demonstrated, caution must be taken when relating such effects to trawl disturbance, in particular in studies that lack appropriate replicate controls. Second, when effects have not been detected, it still cannot be concluded that there is no impact from fishing. The chance of detecting potential changes caused by trawling can be low because the power of the statistical tests in some studies has been shown to be very low (e.g. McConnaughey, Mier and Dew, 2000). Despite these deficiencies, and the fact that most studies have limitations, the growing literature on trawling impacts allows some conclusions to be drawn.
Habitats exposed to natural disturbances and showing large interannual variability in their benthic assemblage seem to be resistant to intensive otter trawling. The impacts demonstrated on soft bottom communities subjected to shrimp trawling were also subtle compared with natural seasonal changes. Habitats dominated by large sessile fauna may be severely affected by trawling.
The comprehensive experiment conducted on the Grand Banks showed a 24 percent decrease in total biomass of megabenthic species (Prena et al., 1999). This decrease in biomass was explained by predation by scavengers migrating into the narrow trawled corridors and a change in catchability of the sampling sledge after the trawling. It may therefore represent an overestimate of the real effect of the disturbance (D.C. Gordon, personal communication). For the macrofauna, total numbers of individuals decreased by 25 percent (mainly owing to declines in polychaetes) immediately after trawling in one of the three years of the experiment (Kenchington et al., 2001). This decline seemed to recover within a year, and very few community indices or taxa showed any long-term effects from trawling. The most prominent feature of the Grand Banks study was considerable interannual variability in the mega- and macrofaunal assemblage, which indicates that the benthic community at the study site is dynamic and exhibits natural changes (Kenchington et al., 2001). Similar conclusions can be drawn from the Barents Sea experiment by Kutti et al. (in press). This study was conducted in an area exposed to natural disturbances, and the benthic assemblage seemed to be resistant to intensive trawling.
Studies investigating the long-term effects of commercial trawling are few because comparable unfished control areas seldom exist. One exception is the study in the eastern Bering Sea by McConnaughey, Mier and Dew (2000), who showed that trawling caused reduced biomass of erect sessile organisms and mixed responses within motile groups and bivalves. Long-term effects of trawling were also indicated in the study of Engel and Kvitek (1998), but this study had serious shortcomings (lack of an unfished control site and no site replication), and should be interpreted with caution.
The four experiments on shrimp trawling provide no clear evidence of disturbance effects on benthic soft bottom communities, with the exception of a decrease in the abundance of echinoderms (mainly brittle stars) demonstrated in the Gullmarsfjord experiment (Van Dolah, Wendt and Levisen, 1991; Hansson et al., 2000/Lindegarth et al., 2000a; Drabsch, Tanner and Connell, 2001; Sparks-McConkey and Watling, 2001). Natural seasonal changes in community structures were demonstrated in these studies, whereas the changes that were attributed to trawling disturbances were regarded to be subtle. The two experiments on Nephrops trawling were also conducted on fine sediment bottoms (clay-sand), and one of them showed that trawling caused short-term effects in abundance and community composition (Frid, Clark and Hall, 1999; Ball, Fox and Munday, 2000). However, these studies lacked true control sites, and comparisons were made between sites displaying different physical characteristics. The study by Tuck et al. (1998) on a silty bottom also had several shortcomings, and most of the effects demonstrated were shown to recover after trawling ceased and were relatively subtle compared with the spatial and temporal variability in the area studied.
Mediterranean ecosystems may be less robust to disturbance than others owing to their special chemical and physical characteristics (oligotrophic, high salinity and temperature, negligible tidal currents). However, the three otter trawl studies conducted in the Mediterranean did not indicate that trawling disturbance caused severe changes in benthic community structures (Simboura et al., 1998; Sanchez et al., 2000; Smith, Papadopoulou and Diliberto, 2000). One of the studies showed a higher number of species and abundance on the control sites (Smith, Papadopoulou and Diliberto, 2000), while another showed that these figures were higher on the trawled site (Sanchez et al., 2000). Furthermore, these Mediterranean studies demonstrated considerable differences in community structures that were attributed to natural variability in time and space.
Studies on the impacts of trawling on hard bottoms are few, but the three studies reviewed here all showed effects on large erect sessile invertebrates (Van Dolah, Wendt and Levisen, 1987; Freese et al., 1999; Moran and Stephenson, 2000). Large proportions (15 to 67 percent) of the tall sessile fauna were damaged by one pass of the ground gear. These results were based on observations in the path of the ground gear, and the proportion of animals damaged averaged over the whole area fished by the commercial fleet will be considerably lower (see Moran and Stephenson, 2000). Reduced biomass of erect sessile organisms was also demonstrated in the eastern Bering Sea (McConnaughey, Mier and Dew, 2000). Thus it can be concluded that tall sessile invertebrates such as sponges are damaged to a large extent when hit by the ground gear and, depending on the proportion of the fishing ground that is touched by this part of the trawl, habitats dominated by large sessile fauna may be severely affected by trawling.
Several studies provide clear evidence of the short-term effects of beam trawling and scallop dredging, with considerable decreases in abundance of several species. Trawling disturbance showed few effects in areas exposed to natural disturbances, and the long-term effects have not been studied.
Studies on the impacts of beam trawling have mainly been conducted in the North Sea and Irish Sea, and owing to the lack of appropriate control areas most of them investigated the immediate effects by comparing samples taken before and shortly after experimental trawling. These studies provide clear evidence of the short-term effects of beam trawling (Bergman and Hup, 1992; Kaiser and Spencer, 1996; Kaiser et al., 1998; Bergman and Santbrink, 2000). Some species of several taxa (echinoderms, polychaetes, bivalves, crustaceans) were shown to decrease considerably in abundance (by up to 68 percent) immediately after trawling. This decrease was explained by damage from the passage of the trawl, exposure in the trawl track and removal by the trawl. The immediate effects demonstrated in these studies cannot be extrapolated to long-term effects of beam trawling on the benthic community (Bergman and Hup, 1992), and the reduction in abundances of megafaunal species observed immediately after trawling was shown to recover six months later (Kaiser et al., 1998). On the other hand, a study based on fishing effort data showed that the total biomass of infauna decreased considerably with a 27-fold increase in trawling disturbance in an intensively trawled area of the North Sea (Jennings et al., 2001). The two studies on beam trawling that were conducted in an area of mobile sediments subjected to natural disturbance showed no effects attributed to trawling disturbance (Kaiser and Spencer, 1996; Kaiser et al., 1998).
Most of the studies on the impacts of scallop dredging are also based either on samples taken immediately after experimental trawling to demonstrate short-term effects or on comparison between areas subjected to different levels of commercial trawling. Effects demonstrated in the latter type of studies cannot be related unambiguously to dredging disturbance, as these experiments may involve the comparison of different habitat types. However, with the exception of two studies (Eleftheriou and Robertson, 1992; Hoffmann and Dolmer, 2000), effects on community structures were demonstrated in all the studies on scallop dredging reviewed here. The most common effects demonstrated were a decrease in number of species and reduced abundance for certain species (by 20 to 80 percent). These effects can be explained by mechanical damage and removal of benthos by the dredge. Recovery of the benthic community after dredging disturbance has been investigated in only one study, which showed that few effects lasted beyond eight months after dredging (Currie and Parry, 1996). Interestingly, the two studies in which no effects from dredging were observed were conducted at sites that were exposed to strong wave and tidal actions (Eleftheriou and Robertson, 1992) or that were subjected to oxygen depletion and large fluctuations in other environmental factors (Hoffmann and Dolmer, 2000). Several of the studies on the impacts of dredging demonstrated that natural temporal and spatial variability in community structure may be more pronounced than changes caused by dredging disturbance (Currie and Parry, 1996, 1999; Thrush et al., 1998; Kaiser, Spencer and Hart, 2000).
Despite the high number of impact studies conducted to date, this review has shown that few general conclusions can be drawn. As few studies incorporate factors such as habitat type, depth, level of disturbance, gear type and geographical region, the results of any single study are highly specific. In order to overcome this lack of progress, Collie et al. (2000) extracted results from 39 fishing impact studies and undertook an interesting meta-analysis. Although some consistent patterns and general relationships emerged from their study, the authors stated that their analyses had obvious limitations and the approach should be regarded as illustrative because of its paucity of data. Results from studies that incorporate factors such as habitat type, disturbance regime and gear type are needed before general conclusions can be drawn.
This review has demonstrated the importance of including evaluation and consideration of the methodology applied in studies of the impacts of trawling disturbances. Most impact studies fail to fulfil one or more of the important requirements of a proper assessment of trawl impacts. An evaluation of the impacts that trawling activities may have on benthic communities may lead to biased or even undocumented conclusions if limitations in the methodology applied are not considered.
Benthic hard bottom habitats dominated by sessile erect fauna are most severely affected by otter trawling, whereas only subtle effects have been demonstrated on soft bottoms without tall sessile invertebrates. However, most impact studies have been on sandy sea beds in relatively shallow water (at shelf depths, i.e. above the wave base), and little is known about the impacts in deep muddy habitats. Furthermore, potential damage in unique, sensitive habitats such as deep-water coral reefs is likely to be severe (Koslow et al., 2001; Hall-Spencer, Allain and Fosså, 2002).
The level of impact on hard bottom communities will depend on the proportion of the habitat to be swept by the ground gear and the extent to which parts of the ground gear hit organisms. The full width or part of the ground gear may lose bottom contact, particularly on rough bottom (J.W. Valdemarsen, personal communication), and rock hopper and bobbins gear have large open spaces between the discs and bobbins (see Figure 5e). On the other hand, some fishing grounds are intensively fished and, owing to the pronounced patchiness of trawling effort, targeted fishing areas can be trawled several times per year (Rijnsdorp et al., 1998; Prena et al., 1999). If tall sessile fauna dominates such areas, the impacts are likely to be severe.
Documentation of the impacts of trawling in certain benthic habitats gives rise to the interesting management issue as to whether the associated fish populations and other exploited marine resources are affected by changes to habitat and benthic community structure. Although current knowledge of the linkage between benthic habitat complexity and the dynamics of fish populations is rudimentary (Auster and Langton, 1999), some effects on the fish community have been demonstrated, e.g. higher juvenile survivorship in more complex habitats (Tupper and Boutilier, 1995) and changes in the abundance of different fish genera following alteration in the abundance of epibenthic fauna (Sainsbury, 1988; Sainsbury et al., 1997). The ultimate impacts of trawling on exploited marine resources cannot be fully understood until more knowledge of this linkage has been obtained.
Beam trawling and scallop dredging have also been shown to affect benthic communities, although only short-term effects have been studied. More emphasis should be given to investigations aimed at studying the long-term effects because fish populations will be affected more seriously by potential long-term effects. Several studies have demonstrated that anthropogenic impacts have a negative effect on longer-lived benthic species, but a positive effect on small opportunistic species (see Thrush et al., 1998), and sole and plaice in the North Sea, which feed mainly on opportunistic species, have shown increasing growth rate over the last decades (Rijnsdorp and van Beek, 1991; Rijnsdorp and van Leeuwen, 1996). This increase in growth rate may be a result of increased productivity of suitable benthic food in heavily trawled areas of the North Sea, as these fishing grounds still provide profitable trawl catches (Rijnsdorp et al., 1998).
Although an increasing number of impact studies have been conducted over the last two decades, it can be concluded that the knowledge of how towed fishing gears affect different habitat types is still rather rudimentary. The main reasons for this lack of scientifically based knowledge is that such studies are very complicated and demanding to conduct and that benthic communities show large natural variability that is not well understood. Knowledge of seasonal, annual and spatial variations in the benthic assemblage studied is therefore a prerequisite for assessing the impacts of trawling disturbances. Furthermore, there is a need to define the degree to which it is accepted that fishing activities cause changes in marine community structures. The degree of anthropogenic impacts must be evaluated in relation to the magnitude of natural disturbances (e.g. current and wave actions, fluctuations in temperature and salinity), as well as temporal and spatial variations that can be both substantial and unpredictable (Auster and Langton, 1999). Gear impacts need to be balanced against the need to harvest resources (Auster et al., 1996).
![]()
![]()
![]()