
|
||||||

|
|
|||||
|
FAO. 2009. Porphyra spp. In Cultured aquatic species fact sheets. Text by Chen, J. & Pu Xu. Edited and compiled by Valerio Crespi and Michael New. CD-ROM (multilingual). |
||||||

 |
 |
Thalles de Porphyra et ses crampons (photo de MBARA sur internet) |
Unités de culture des filaments conchocelis |
 |
 |
Récolte manuelle |
Récolte avec machine spéciale |
 |
 |
Traitement de Porphyra |
Sushi cuisiné selon différentes formes |
En Chine, le Porphyra haitanensis est présent dans les zones méridionales, tandis que le P. yezoensis est trouvé dans le nord. Le cycle de vie qui s'appelle généralement nori ou laver, comporte les thalles des espèces (voir le n.1 dans la figure ci-dessous), qui émergent en automne ou en début d'hiver le long des côtes rocheuses. Ses graines s'appellent conchospores et sont libérées par le stade conchocelis filamenteux (voir le n. 8 dans la même figure), elles sont trouvées fixées sur des coquilles de mollusques en été et en automne. Avant 1949, le stade filamenteux était considéré comme une espèce séparée, Conchocelis rosea.
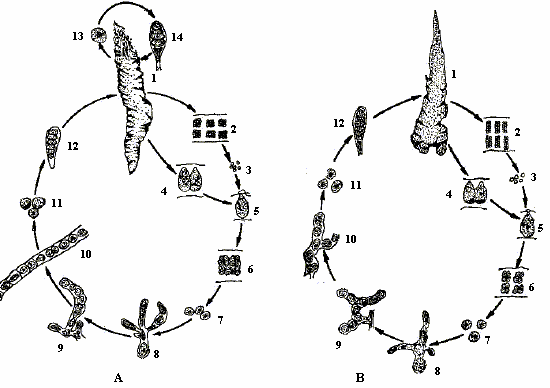
Schema du cycle de developpement du Porphyra spp. - A = Porphyra yezoensis; B = Porphyra haitanensis. 1 thalles; 2 spermagonie; 3 sperme; 4 carpogone; 5 oeuf fertilisé; 6 carposporange; 7 carpospore; 8 conchocelis filamenteux; 9 branche de sporanges des conchocelis; 10 libération des conchospores; 11 conchospore; 12 thalles jeunes; 13 monospore et thalles jeunes |
Le stade conchocelis est diploïde. Sous des conditions spécifiques d'intensité et de qualité de lumière, la longueur et la température du jour stimulent la production des gamètes. Ces filaments forment les branches gonflées («conchosporange») dans lesquelles les cellules développent les caractéristiques plastidiales des cellules du stade lame (plastides étoilées avec le pyrénoïde visible). Ces branches s'éloignent du substrat et libèrent, par la suite, leur contenu sous formes de cellules individuelles («conchospores»).
Les gamètes mâles (voir schéma, 3: «spermaties») sont produits en groupes (2) aux bords des lames et sont libérés par la dissolution du bord. Les gamètes femelles (4: «carpogones») sont formés d'une certaine manière derrière les bords. Une surface réceptive («trichogyne») déborde de chaque carpogone et à travers la matrice environnante, à laquelle les spermaties s'attachent et accomplissent la fécondation (5). Le zygote se divise alors pour former un groupe de cellules diploïdes (6: «carposporange»). Les «carpospores» diploïdes (7) sont libérés du carposporange par la dissolution des bords des lames. La germination des carpospores, pour former les filaments conchocelis diploïdes (8), ne dépend pas de la présence du carbonate de calcium solide; cependant, apparemment, dans des conditions naturelles seules ces carpospores parviennent à germer, et pénètrent plus tard, ce substrat (coquilles des mollusques) évitant ainsi d'être mangés par les escargots et d'autres petits brouteurs marins.
Dans certaines conditions, les filaments conchocelis (9-10) se développent et forment les conchospores (11). En automne, ces conchospores sont libérées des sporanges développés dans les filaments conchocelis gonflés dans l'eau et se fixent (12) sur le substrat et commencent leur nouveau cycle de vie (1).
En plus de ce processus, les P. yezoensis (voir le schéma A ci-dessus) se reproduisent aussi par voie asexuée, en produisant des monospores sur les bords des thalles. Ceci ne se produit pas chez les P. haitanensis (voir le schéma B ci-dessus).
Les conditions optimales de température, salinité, et intensité lumineuse pour la croissance des thalles de Porphyra varient. Généralement, les jeunes thalles peuvent supporter une température plus élevée que les thalles adultes; les températures plus basses influenceront la croissance normale du P. yezoensis et du P. haitanensis (moins de 3 °C et 8 °C respectivement). Une intensité de la lumière élevée (5 000-8 000 lux) est idéale pour la croissance des deux espèces. Les thalles de Porphyra tolèrent la sécheresse; par exemple, le P. haitanensis peut survivre pendant une semaine, même s'il perd 70 pour cent de son humidité. Le Porphyra est un autophyte; les aliments tels que l'azote nitrique ou l'azote ammoniacal lui sont essentiels. Les expériences indiquent que 100-200 mg d'azote/m3 est nécessaire pour une croissance normale; la croissance sera perturbée si le niveau d'azote est inférieur à 50 mg d'azote/m3.
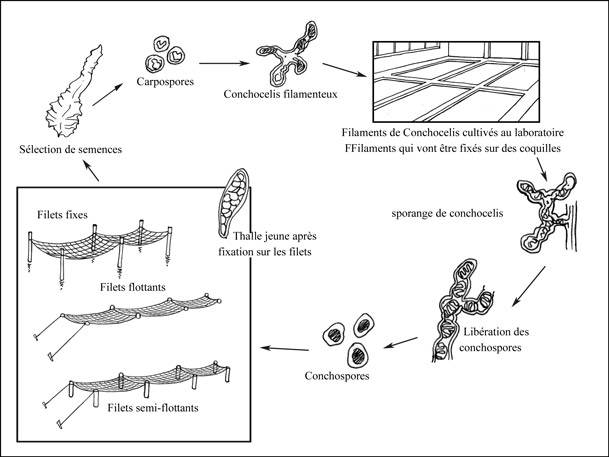
Élevage de conchocelis
Comme il a été précédemment mentionné dans cette fiche d'information, l'élevage du nori se produit en deux étapes. La première c'est la culture à l’intérieur, allant de mai jusqu'à octobre, où le stade conchocelis est cultivé et les conchospores se produisent. La deuxième étape a lieu à partir d'octobre jusqu'à avril ou mai; dans cette phase les algoculteurs se concentrent sur la culture des thalles sur le terrain.
Vers début et mi-mai, les cythérées communes (Meretrix meretrix) sont semées en desséchant les conchocelis pour libérer les conchospores; le substrat est alors aspergé par ces conchospores en suspension, ou bien il est submergé dans la suspension. La culture a lieu dans de grands réservoirs ovales peu profonds remplis de 20-30 cm d'eau de mer qui a été soumise à la sédimentation et à laquelle ont été ajoutés des aliments azotés et du phosphate. A ce stade, la température n'est pas contrôlée, elle varie avec la température ambiante. Cependant, la croissance optimale se produit à une température entre 20-25 °C. Les carpospores se développent dans la phase conchocelis.
De la mi-mai jusqu'au début juin, la température d'eau augmente à 22-23 °C, pour stimuler le développement végétatif de la phase conchocelis. En juillet, l'intensité de la lumière est réduite pour inciter la formation des conchosporanges, qui vont, par la suite, libérer les conchospores. A partir du début juillet jusqu’à fin août, la température d'eau augmente graduellement pour atteindre 27-28 °C à la mi-août. Après, la température d'eau commence à diminuer progressivement de nouveau.
Dès fin août jusqu'à fin septembre, la température d'eau baisse à 23 °C; c’est la période de formation des conchosporanges. L'intensité de la lumière est tenue au même niveau pour encourager la formation des conchospores dans le conchosporange et la longueur du jour est réduite à 8-10 heures. Les conchospores se forment en fin septembre mais le phénomène en masse se produit au début et à la mi-octobre.
Une fois les conchospores sont libérés, ils sont semés sur des filets de culture spéciaux qui sont placés dans des bassins. Quand 50 000 spores sont libérées par jour, l'intensité de la lumière est augmentée légèrement pour stimuler la germination des thalles. De même, l'eau dans les bassins est agitée pour une distribution uniforme des spores et pour assurer leur contact avec les filets de culture. Cette opération s'appelle «collecte des graines». Environ 50 pour cent des fermes de laver procèdent, eux-mêmes, à l'élevage des semences, alors que le reste achète des filets de culture chez les fournisseurs de graines.
Il existe trois principales méthodes: (a) structures flottantes, (b) semi-flottantes; et (c) filets fixes. Une quatrième technique connue sous le nom de "freezing nets" (voir ci-dessous).
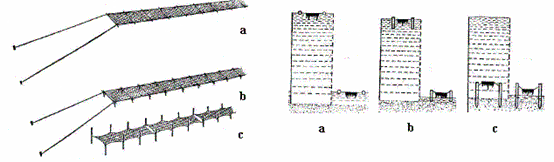
Trois méthodes d'élevage du Porphyra spp.
Système flottant
Cette méthode était pratiquée au Japon et récemment elle a été largement adoptée par des algoculteurs chinois. Les filets sont fixés aux bouées flottantes sur la surface de la mer, ainsi les thalles nori sont constamment imbibés d'eau. Avec cette méthode il est possible de cultiver les thalles même dans les zones en dehors des baies peu profondes, dans des profondeurs de 10-20 m.
Système semi-flottant
Cette méthode est largement répandue en Chine, c’est un mélange de systèmes flottant et filet fixe. Pendant la marée haute les filets flottent sur la surface, durant les marées basses (reflux), le système se pose par terre. Cette méthode combine les avantages des systèmes flottants et ceux des filets fixes.
Système de filets fixes
Dans ce système, également connu sous le nom de «filets fixés sur des pieux», les filets de nori sont accrochés entre des pieux. Pendant les marées basses, les filets sont exposés à l'air et deviennent secs. La culture intertidale sur pieux prévale souvent sur la culture flottante ou semi-flottante dans l'eau profonde, parce qu'elle assure l'exposition périodique, ce qui aide à réduire l'éruption des maladies et la croissance des espèces concurrentielles (plantes aquatiques), particulièrement les diatomées épiphytiques. Cependant, ce type de culture est limité aux parties intérieures des baies, avec les fonds peu profonds et sableux.
Filets de congélation ou Freezing nets
Cette méthode a deux avantages: la prévention des maladies chez les nori, et l'amélioration de la qualité du produit final. La procédure à suivre pour le traitement des filets de congélation est comme suit: �
- Quand les jeunes thalles atteignent 1 à 3 cm de longueur, ils sont rapportés des zones d'élevage, ainsi que les filets.
- Les filets sont séchés à l'air jusqu'à ce que la teneur en humidité des thalles diminue de 20-40 pour cent, d'habitue cette opération dure 2 à 3 heures.
- Les filets séchés sont mis dans des sacs en vinyle.
- Les filets séchés sont stockés dans un congélateur à une température de -20 ºC.
- En cas de besoin, les filets sont pris et rapportés de nouveau vers les zones d'élevage de nori.
La première phase de récolte commence après l’élevage en mer ouverte pendant 40 ou 50 jours dans le cas du Porphyra haitanensis et P. yezoensis respectivement. Les activités de récolte peuvent durer 5 mois. L'intervalle entre les récoltes est de10 à 15 jours; ainsi, la production est récoltée 10 à 12 fois annuellement. Deux méthodes sont utilisées, à savoir la récolte manuelle et mécanique, comme illustré dans la galerie d'images.
Les thalles récoltés sont coupés en morceaux et traités en les séchant dans des flocons traditionnels minces et rectangulaires, d'une taille de 21 x 19 cm chacun et pesant environ 3 g. Le procédé consiste en le choix de nori humide; son lavage avec eau de mer puis avec eau douce; coupage; ramollissement; transformation en flocons; déshydratation; séchage; triage; et emballage.
Le nori séché peut être raffiné en produits assaisonnés qui peuvent être directement consommés; ces articles sont devenus aussi populaires que les sucreries.
Les coûts de production varient selon les régions; aucune information n'est disponible.
| MALADIE | AGENT | TYPE | SYNDROME | MESURES |
| Chytridiose | Olpidiopsis sp. | Champignon | Moisissure; produits de mauvaise qualité; production réduite | Réduction de la densité des filets de culture; amélioration de la gestion des premiers stages |
| Moisissure | Pythium sp. | Champignon | Moisissure; produits de qualité inférieure; production réduite | Séchage des filets d'élevage; réduire la température |
| Bactérie filamenteuse | Leucothrix mucor | Bactérie | aucune information | Séchage des filets d'élevage |
| Atrophie | N'est pas connu | N'est pas connu | Les thalles infectés sont agaric (comme des champignons) et couleur sombre | Aucune mesure connue |
| Maladie des pores | N'est pas connu | Bactéries Gram-négatif | Les microbes pathogènes absorbent les aliments des thalles, faisant apparaître des trous | Réduction de la salinité en cas de culture dans les laboratoires; aucune mesure en cas d'élevage dans la nature |
| Pourriture verte | Pseudomonas sp. | Bactéries Gram-négatif | Moisissure | Séchage des filets de culture; réduire la densité de plantation |
| Décoloration | Stress environnemental | La couleur des thalles change de pourpre à verte jaunâtre et puis à blanche | Réduire la densité de stockage |
En général, le bon choix du site potentiel, les bonnes densités de plantation et la bonne gestion sont considérés comme les mesures les plus efficaces pour la prévention des maladies.
Fournisseurs d'expertise en pathologie
-
Jiangsu Marine Fisheries Research Institute
Dr. Xu Pu ([email protected]).
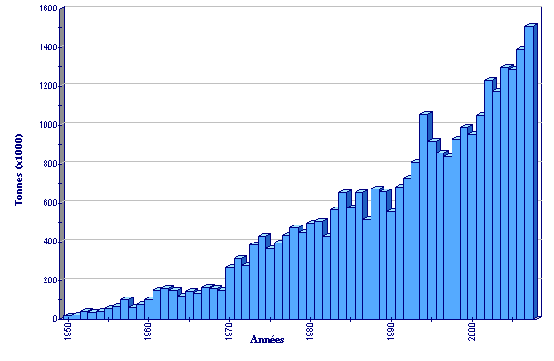
(FAO Statistiques des pêches)
Le nori est riche en protéines et acides aminés libres, de même il a un bon goût; c'est un aliment conventionnel de la région asiatique en particulier en Chine, Corée et Japon. Le nori est un aliment favori dans ces pays, notamment au Japon. Dans les buffets des gares de train, des hôtels et des restaurants, il remplace le sandwich, et il est présenté au public sous le nom de «sushi». Ce dernier est préparé en plaçant le riz et les bandes bouillis de la viande ou de poisson sur une feuille de nori, qui est alors enroulée et coupée en tranches (voir photo dans la galerie d'images).
Le nori est également coupé en petits morceaux et utilisé dans la fabrication des biscuits. Avec le développement de l'économie chinoise, récemment, les consommateurs locaux ont nettement augmenté. Comme mentionné ci-dessus, les « fast food » à base de nori sont des produits plus populaires que les sucreries. Les produits du P. haitanensis sont transformés en nori séché pour la préparation de potage et d'autres nourritures traditionnelles en Chine. Le nori est reconnu en tant qu'aliment salubre. Il est également accepté par des personnes vivant dans d'autres continents y compris l'Amérique et l'Europe. Il n'y a aucun doute que la culture de Porphyra est devenue un secteur rentable de l'industrie aquacole.
Bien que des essais expérimentaux de culture de nori soient entrepris aux États-Unis et au Royaume Uni, ces pays sont loin de devenir des producteurs principaux vu que cette activité demande une main d'œuvre importante.
Dans un proche avenir, les pays de la région asiatique demeureront le principal marché de nori.
| Chen, G.Y. 1980. Study on Breeding of free-conchocelis filament of Porphyra haitanensis and its seeding. Journal of Fisheries of China, 4(1):19-30. |
| Drew K.M. 1949. Conchocelis phase in the life history of Porphyra umbilicali (L) Kutz. Nature,164:748-751. |
| Jia, J. & Chen, J. 2001. Sea Farming and Sea Ranching in China. FAO Fisheries Technical Paper No. 418. FAO, Rome, Italy. 71 pp. |
| Liu, T. J., Wang, S.P., Zhang, D.R., Cui, G.F., Li, W.L., Zhu. Z.J., Wu, T.M., Li, X.G., You, J.X., Huang, F.L., Wang, X.B. and Lin, D.H. 1981. Study on Enhancement of Resources of Porphyra haitanensis. Marine Fisheries Resecrh, 3: 1-67. |
| Liu, T.J., Zhang, Y.J., Yang Y.X. and Wang, S.R. 1982. Prevention and Treatment of Diseases of Porphyra yezoensis. Marine Fisheries Research,4:1-8. |
| Ma, J.H. 1992. Elementary Report on Chytridiosis of Porphyra yezoensis. In Proceedings of 1991's National Mariculture Forum, Hold in Dalian City, Liaoning Province, China from 9 - 11 January 1992; Edited by Mariculture Special Committee, Chinese Fisheries Society, Published by Qingdao Publication Bureau. pp. 265-267. |
| Ma, J.H. & Cai, S.Q. 1996. Farming and Processing of Porphyra yezoensis. Science Press in Beijing, China. pp. 236. |
| McHugh, D. J. 2003. A guide to the seaweed industry. FAO Fisheries Technical Paper No. 441. FAO, Rome, Italy. 116pp. |
| Tseng, C.K. & Zhang, D.R. 1954. Study on Porphyra I: Life cycle of Porphyra tenera. ACTA BOTANICA SINICA. 3(1):287-302. |
| Tseng, C.K. & Zhang, D.R. 1955. Study on Porphyra II: Filament stage of Porphyra tenera and its Conchospores. ACTA BOTANICA SINICA. 4(1):27-46. |
| Tseng, C.K.,Wang, S.J., Liu, S.J., Guo, X.K., Zhang X.C. and Miao G.R. 1978. Phycoculture. Shanghai Science & Technique Press, Shanghai, China. Editor: Tseng C.K, Pp. 135-211. |
| Tseng, C.K., Wu, C.Y., Fei, X.G., Zhang X.C., Qin, S. and Zhou, H.E. 1999. Biology on Economic Seaweed Seeds. In Biological Basics on Porphyra Seed. Shandong Science & Technique Press, pp. 154. Shanghai, China. Editor: Tseng C.K, pp. 50-90. |
| Tseng, C.K., & Borowitzka, M. 2003. Algae culture. In J.S. Lucas & P.C. Southgate (eds.), Aquaculture: Farming Aquatic Animals and Plants, pp. 253-275. Blackwell Publishing, Oxford, England. |