![]()
![]()
![]()
2. PEUPLEMENTS ICHTYOLOGIQUES
Du fait des échanges des populations ichtyologiques fluviales et lacustres la zone deltaïque, lieu de passage obligatoire des migrateurs, constitue un point stratégique pour l'étude des peuplements ichtyologiques.
Au cours d'une année hydrologique moyenne, la crue du Chari est suffisamment importante pour inonder les yaérés, permettre la mise en eau de l'El Beïd et équilibrer le bilan hydrique du lac Tchad. Les captures fournissent alors très probablement une bonne image des peuplements du bief en ce qui concerne leurs phases exploitées. En effet, les pêcheurs appartiennent à de nombreuses ethnies réparties dans plusieurs pays ayant pour la plupart d'entre elles une longue tradition dans le domaine de la pêche. Ils disposent en outre d'un matériel de bonne qualité. Enfin, beaucoup d'entre eux ont quitté leur région d'origine pour se consacrer à la pêche et sont à la recherche du rendement maximum. Dès qu'une concentration de poissons se forme, ce phénomène est aussitôt repéré et exploité.
Les engins de pêche du bief capturent les poissons présents à partir de 150 à 200 mm de longueur standard. C'est entre ces limites en effet que se situent approximativement les longueurs moyennes de sélection pour les espèces pêchées aux filets à petites mailles. Un biais peut parfois résulter de la façon dont l'effort de pêche se répartit, à un moment donné, entre les catégories d'engins. Quoi qu'il en soit la meilleure image du peuplement du bief dont nous puissions disposer est celle fournie par les captures totales faites dans ce bief avec l'ensemble des engins utilisés.
Pour la période qui nous intéresse ici, ces conclusions ne sont plus valables. En effet, ainsi que nous l'avons exposé précédemment le type d'exploitation du bief n'est pas resté constant. Il convient donc de dissocier les deux phases d'exploitation qui se sont succédée entre septembre 1972 et décembre 1973. Durant la première (septembre 1972 - avril 1973) les conditions de pêches sont restées comparables à celles observées les années précédentes, à savoir :
On peut donc considérer que l'échantillonnage des stocks exploitables est encore satisfaisant. Les enquêtes de pêches sont utilisables sous un angle biologique. Au cours de la seconde phase (août à décembre 1973) l'exploitation du bief devient très irrégulière et il est délicat de distinguer l'influence des facteurs humains et celle des facteurs biologiques.
La perte d'information est importante et entrave la bonne compréhension des phénomènes provoqués par la sécheresse durant cette période.
2.1. Choix des paramètres. Mode d'exploitation des données
Les enquêtes des pêches fournissent chaque mois des valeurs de production mensuelle et de p.u.e. en nombre et en poids — pour chacune des espèces capturées —. Lors des enquêtes, afin de réduire au maximum la perte de temps imposée aux pêcheurs par l'observation de leurs prises, seules les longueurs des poissons ont été mesurées. Les poids ont ensuite été estimés à partir de clefs longueur/poids calculées pour un coefficient de condition moyen. (Durand, Franc, Loubens, 1973). Or, la condition vraie des différentes espèces peut différer notablement des valeurs moyennes du fait d'une plus ou moins bonne résistance et/ou adaptation aux conditions écologiques provoquées par la sécheresse. Les productions mensuelles ont le désavantage de donner une grande importance aux efforts de pêche qui sont beaucoup moins liés aux phénomènes biologiques que l'on désire étudier qu'au rythme d'activité des pêcheurs. Nous avons donc écarté ces deux paramètres au profit des p.u.e. spécifiques en nombre.
Les phénomènes liés à l'assèchement du lac Tchad étant complexes et peu prévisibles il était souhaitable d'analyser les données recueillies à l'aide de la méthode nécessitant le minimum d'hypothèses préalables ou de transformations.
Nous avions le choix entre deux types de méthodes. D'une part, les classifications hiérarchiques; d'autre part, les analyses multifactorielles.
Les méthodes de classifications hiérarchiques ont rapidement été écartées. Elles nécessitent un trop grand nombre de choix arbitraires, ne permettent pas d'établir de relations directes entre les groupements de relevés et les groupements de variables qu'elles effectuent sur des tableaux du type P × V (R = {r} = ensemble des relevés, V = {v} = ensemble des variables) enfin et surtout elles s'appliquent mal à l'interprétation de phénomènes continus comme le sont les variations de structure d'un peuplement.
Nous avons donc étudié les méthodes de statistiques descriptives multimentionnelles. Nos premiers essais ont porté sur l'analyse en composantes principales, mais les projections se sont révélées difficiles à interpréter du fait de la complexité des relations entre les points de l'espace paramètre et ceux de l'espace observation. Par ailleurs, ce type d'analyse est basée sur des matrices de corrélations linéaires qui en toute rigueur ne sont valables qu'entre variables normalisées. Il est donc nécessaire d'effectuer une transformation préliminaire des données pour appliquer correctement cette méthode.
L'analyse factorielle des correspondances ne nécessite par contre aucune transformation préalable. Elle présente en outre l'avantage d'établir une relation directe entre l'analyse des espèces et celle des observations par rapport aux axes factoriels qu'elle définit. Elle permet ainsi une représentation synthétique des résultats, la position relative d'un point station se situant au barycentre des points espèces affectés des poids que celles-ci présentent pour le relevé considéré. Un relevé sera donc d'autant plus proche d'une espèce que celle-ci aura d'importance dans son profil. Inversement, les espèces se situeront à proximité des relevés où elles sont abondantes.
Nous avons complété l'étude des projections obtenues par analyse des correspondances, par le calcul des indices de diversité mensuel selon la formule de Shannon :
I = - ∑ Pi Log2 Pi
Pi = fréquence relative de l'espèce i.
à partir des fréquences relatives des 15 espèces les plus abondantes, ou de celles parmi les 15 qui étaient présentes pour le mois considéré. Les autres espèces dont les fréquences relatives sont toujours faibles influent peu sur la valeur de I.
Nous avons également calculé l'équitabilité qui se définit comme étant le rapport : I/IMAX (IMAX étant la diversité maximale pour le nombre n d'espèces présentes = Log2n), (Tabl. X).
2.2. Evolution du peuplement.
44 espèces ont été capturées entre septembre 1972 et décembre 1973, soit la quasi totalité des espèces capturables avec les filets utilisés (25 à 35 mm pour les petites mailles; 40 à 70 mm pour les grandes mailles).
Tableau X
Indices de diversité et équitabilité
| I | IMAX | I/IMAX | |
| SEP. 72 | 1,8912 | 3,1699 | 0,5966 |
| OCT. 72 | 1,9169 | 3,3219 | 0,5771 |
| NOV. 72 | 2,3076 | 3,7004 | 0,6236 |
| DÉC. 72 | 2,3493 | 3,7004 | 0,6349 |
| JAN. 73 | 1,4647 | 3,9069 | 0,3749 |
| FÉV. 73 | 2,2478 | 3,8074 | 0,5904 |
| MAR. 73 | 2,9396 | 3,8074 | 0,7721 |
| AVR. 73 | 2,8157 | 3,3219 | 0,8476 |
| AOU. 73 | 2,4261 | 3,8074 | 0,6372 |
| SEP. 73 | 2,2191 | 3,4594 | 0,6415 |
| OCT. 73 | 0,6246 | 2,5850 | 0,2416 |
| NOV. 73 | 0,3264 | 3,1699 | 0,1030 |
| DÉC. 73 | 2,0423 | 3,1699 | 0,6443 |
Toutes les espèces n'apparaissent pas avec des fréquences importantes dans les captures, loin s'en faut. Sur les 6980 poissons identifiés, 6 827 soit 97,81 % du total se répartissent entre 15 espèces, et, 6 600 soit 94,57 % entre les dix espèces les plus abondantes.
| Alestes baremoze | 27,87 % |
| Synodontis batensoda | 15,73 % |
| Synodontis membranaceus | 14,66 % |
| Alestes dentex | 12,82 % |
| Marcusenius cyprinoïdes | 8,95 % |
| Hyperopisus bebe | 4,58 % |
| Schilbe spp | 4,05 % |
| Eutropius niloticus | 2,12 % |
| Hydrocyon forskalii | 2,01 % |
| Synodontis cf. schall | 1,78 % |
L'analyse a été effectuée à partir des p.u.e. en nombre des 15 espèces les plus abondantes (97,81 % du total des captures) pour les 8 mois pendant lesquels la pêche a été effectuée de façon régulière par des professionnels. Les p.u.e. considérées sont des moyennes des p.u.e. observées pour les filets à grandes mailles et les filets à petites mailles pondérées par les efforts de pêche de ces deux types d'engins.
Les mois d'août à décembre 1973 pour lesquels les méthodes de pêche sont très changeantes n'ont pas été inclus dans l'analyse mais ont été replacés sur les graphiques après calcul de leurs coordonnées sur les axes factoriels.
Il est évident que pour un tableau de données aussi restreint, l'utilisation d'une méthode d'analyse sophistiquée n'était pas indispensable. Une étude soigneuse du tableau initial aurait conduit aux mêmes conclusions que celles exposées ci-dessous. L'application de cette méthode descriptive au cas des enquêtes de pêche n'en demeure pas moins un progrès méthodologique par la clarté qu'elle apporte et la rapidité avec laquelle elle permet l'exploitation de données parfois confuses.
Les pourcentages d'inertie extraits par les cinq premiers axes donnés par l'analyse sont les suïvants :
| - 1er axe | 46,74 |
| - 2e axe | 28,88 |
| - 3e axe | 13,66 |
| - 4e axe | 8,35 |
| - 5e axe | 1,89 |
soit 89,28 % pour les 3 premiers axes et 99,52 % pour les cinq premiers.
La signification des axes peut s'expliquer par les contributions des espèces. Les espèces présentant les plus fortes contributions à un axe factoriel sont davantage responsables de la répartition des relevés sur cet axe. A l'inverse les espèces de contribution nulles n'influent pas sur cette répartition.
C'est ainsi que l'on peut interpréter la répartition des prélèvements mensuels sur les 3 premiers axes en fonction de leur abondance respectivement en Synodontis membranaceus, Alestes baremoze et Alestes dentex, et Schilbe spp.
Synodontis membranaceus contribue très fortement au premier axe, 68,6 % et masque ainsi l'influence des autres espèces. Cette forte contribution est liée à une grande abondance à une période précise de l'année. Comme les variations de fréquences de cette espèce sont bien marquées et nettement repérables, il est possible de la soustraire de l'analyse sans perte d'information.
Une seconde analyse portant sur les 14 espèces restantes a été entreprise. Les % d'inertie extraits par les cinq premiers axes sont :
| - 1er axe | 48,23 % |
| - 2e axe | 23,24 % |
| - 3e axe | 15,39 % |
| - 4e axe | 7,57 % |
| - 5e axe | 3,59 % |
soit 86,86 % pour les 3 premiers axes et 98,0 % pour les cinq premiers. La condensation de l'information est du même ordre que dans la première analyse. Les contributions au premier axe sont par contre beaucoup plus équilibrées puisque 80 % des contributions à cet axe sont réparties entre Synodontis frontosus, Synodontis schall, Alestes dentex et Polypterus bichir.
Les espèces contribuant le plus aux deuxièmes et troisièmes axes sont respectivement : A. baremoze, A. dentex, Schilbe spp. (72,4 %), Eutropius niloticus, et Hydrocyon forskalii (59,10 %) (tabl. XI).
Après élimination de Synodontis membranaceus, 6 espèces au lieu de 3 contribuent à définir les 2 premiers axes et 8 espèces au lieu de 4 contribuent à définir les 3 premiers axes. La description que les premiers plans offrent de l'évolution du peuplement est ainsi plus complète et plus fidèle. Le premier plan restituant 71,47 % de l'inertie totale est suffisant pour permettre de comprendre cette évolution (fig. 7). Les mois de septembre, octobre, novembre et décembre 1972 forment un noyau assez lâche autour des espèces dominantes durant cette période : Alestes baremoze, Alestes dentex, Marcusenius cyprinoïdes, Synodontis batensoda auxquelles vient se joindre Synodontis membranaceus en octobre et novembre. La richesse spécifique s'accroît régulièrement durant cette période mais l'équitabilité se maintient constante (tabl. X).
Vient ensuite le mois de janvier qui se différencie nettement des précédents du fait d'une grande abondance en Alestes baremoze (78,1 % des captures du mois). L'équitabilité diminue malgré un accroissement de la richesse spécifique.
Les mois de février, mars et avril 1973 montrent l'évolution du peuplement jusqu'à l'arrêt de la pêche fin avril. On observe un glissement très net du peuplement à A. dentex et A. baremoze dominant décrit précédement vers un peuplement à Synodontis dominant où les A. baremoze et A. dentex sont en nette régression. Synodontis clarias, Synodontis frontosus, Polypterus bichir, Synodontis schall et gambiensis. L'équitabilité ne cesse de croître durant cette période pour atteindre 84,8 % en avril du fait de l'accroissement des fréquences d'apparitions de nombreuses espèces.
Ce phénomène n'est pas lié au développement de ces espèces mais simplement à la régression des A. baremoze et A. dentex dont les fréquences importantes éclipsaient ces espèces les mois précédents. Aprèse la reprise de la pêche en août-septembre 1973, on retrouve une diversité comparable à septembre 1972 mais la richesse spécifique est plus importante et surtout les A. baremoze et A. dentex sont très rares. Ils disparaissent complètement des captures à partir d'octobre. La diversité subit alors de fortes variations du fait du passage de migrations de Synodontis membranaceus en novembre et curieusement de Synodontis batensoda en octobre. L'utilisation pour la pêche d'un même type d'engin renforce l'effet apparent de ces migrations sur le peuplement.
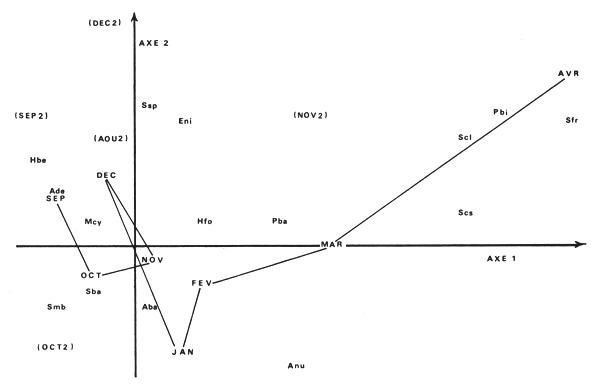
Fig. 7. — Projection, sur le plan des axes 1 et 2, des espèces et des mois entre septembre 1972 et avril 1973.
Tableau XI
Cotes, corrélations et contributions des espèces aux trois premiers axes factoriels définis par la seconde analyse
| 1er Facteur | 2e Facteur | 3e Facteur | |||||||
| Espèces | Cote | Cor | Ctr | Cote | Cor | Ctr | Cote | Cor | Ctr |
| A. baremoze | -81 | 442 | 9 | 381 | 896 | 390 | 57 | 20 | 13 |
| S. batensoda | 302 | 111 | 14 | 254 | 78 | 21 | -425 | 221 | 91 |
| A. dentex | 444 | 598 | 172 | -342 | 356 | 212 | -109 | 37 | 33 |
| M. cyprinoïdes | 216 | 92 | 3 | -174 | 60 | 4 | -101 | 20 | 2 |
| M. bebe | 655 | 249 | 34 | -538 | 168 | 48 | -619 | 223 | 96 |
| Schilbe spp | -64 | 2 | 2 | -879 | 357 | 122 | 1131 | 589 | 305 |
| E. niloticus | -312 | 58 | 5 | -778 | 362 | 59 | 889 | 471 | 116 |
| H. forskalii | -384 | 159 | 12 | -154 | 26 | 4 | 813 | 711 | 170 |
| S. cf. chall | -2022 | 948 | 190 | -229 | 12 | 5 | -356 | 30 | 19 |
| S. frontosus | -2692 | 849 | 341 | -778 | 71 | 59 | -761 | 68 | 86 |
| S. clarias | -2014 | 696 | 85 | -654 | 74 | 19 | -467 | 38 | 14 |
| A. nurse | -990 | 359 | 21 | 772 | 219 | 27 | 308 | 35 | 6 |
| P. bichir | -2235 | 844 | 102 | -816 | 113 | 28 | -371 | 23 | 9 |
| P. bane | -871 | 237 | 13 | -172 | 9 | 1 | 879 | 241 | 40 |
| 1 000 | 1 000 | 1 000 | |||||||
La comparaison de l'état du peuplement deltaïque en 1972–1973 avec celui décrit pour les 13 mois précédents permet de dégager les principaux effets de la sécheresse.
Il faut noter tout d'abord que la liste des 17 espèces abondantes retenues par Loubens est assez différente de celle retenue ici. Si A. baremoze demeure prépondérant bien qu'il n'ait été capturé que 8 mois sur 13, Schilbe spp. passe de la seconde place à la septième sans doute du fait de la faible utilisation des filets dormants où il constitue habituellement le quart des captures. Alestes dentex se situe après Synodontis membranaceus et Synodontis batensoda alors qu'il précédait ces deux espèces.
Distichodus rostratus, Citharinus citharus, Labeo sengalensis, Hydrocyon brevis, Bagrus bayad, Citharinus latus ne font plus partie des 15 espèces les plus abondantes et sont remplacées par des espèces autrefois plus rares comme Marcusenius cyprinoides, Synodontis frontosus, Synodontis clarias, Petrocephalus bane.
Il est à noter qu'il s'agit surtout de poissons de grande taille et que leur variation d'abondance ne peut être imputée à un relâchement de l'effort de pêche fournit pour les filets de grandes mailles puisque ces derniers sont les seuls à être utilisés à partir d'octobre 1973.
Une seconde différence importante entre les 2 séries de relevés est due à la disparition progressive des A. baremoze et A. dentex à partir de la mi 1973.
Disparition significative malgré le changement de mode de pêche. La grande valeur marchande de ces poissons étant le garant de leur exploitation. Les causes de cette disparition sont liées à l'évolution du Lac. Les individus capturés au delta proviennent de l'Archipel Sud-Est et surtout de la cuvette Nord dont l'Archipel constitue le principal réservoir en Alestes spp. (Durand, Loubens, 1970).
L'isolement de l'ancienne zone d'eau libre de la cuvette Sud a donc coupé le delta des régions assurant sont approvisionnement en grands Alestes d'où leur disparition rapide des captures.
Il importe enfin de noter le développement des Mochocidae durant la 2e moitié de 1973. Ces espèces par leurs nombres et leurs bonnes conditions semblent montrer une bonne adaptation aux milieux turbides et peu profonds.
Peu de renseignements complémentaires à la biologie des espèces peuvent être apportés ici. L'arrêt de la pêche en avril et août 1973 rend impossibles les comparaisons avec la période précédente quand aux cycles migratoires en particulier pour A. baremoze, Schilbe spp., Synodontis batensoda, Synodontis schall, Eutropius niloticus, et Polypterus bichir. Pour toutes ces espèces les maximums de passage dans le bief étudié ont habituellement lieu entre les mois de mai et août.
Il est probable que s'il y a eu migration à la période normale pour ces espèces elle dut être peu importante. Car on peut supposer qu'alors les p.u.e. n'auraient pas pris les valeurs aussi faibles que celles qui ont été observées en avril. Mais faute de données entre mai et juillet, il est difficile de conclure. Il est à noter cependant que si les migrations n'ont pu se produire à la période normale pour ces espèces un certain nombre d'entre elles semblent abondantes à d'autres périodes. C'est ainsi que S. batensoda apparaît avec des fréquences très importantes en octobre 1973.
Hyperopisus bebe migre également entre mai et août. Sa grande abondance dans les captures de septembre 1973 (45% des captures) laisse à penser que son cycle migratoire aurait pu être décalé dans le temps.
Pour les espèces dont les migrations ne se situent pas entre mai et août. On observe deux cas extrêmes: d'une part A. dentex dont les fréquences sont nulles au plus fort de sa période habituelle de migration (août à décembre); d'autre part S. membranaceus qui représente 96,4% du peuplement au moment de sa migration anadrome. Hydrocyon forskalii migre habituellement de novembre à mars. Ce passage est très discret en 1972, et absolument invisible en 1973 du fait de l'utilisation des grands filets où il se prend peu pendant cette période. Cette espèce supporte assez mal l'accroissement de turbidité lié à la sécheresse.
Polypterus bichir a été signalé par Loubens comme ayant deux périodes de fréquence maximale. La première en mi-juin; la seconde en novembre-décembre. S'il nous est ici impossible de parler de la première période, la seconde est plutôt caractéristique ici de la plus grande rareté en P. bichir les fréquences maxima ayant été observées en février, mars et avril. Ces différences peuvent n'être liées qu'à l'absence de lignes de fond, qui capturent mieux cette espèce que les autres engins.
CONCLUSIONS
La sécheresse qui sévit sur le Sahel au moment de l'étude influe de façon importante sur le peuplement ichtyologique du bassin Tchadien.
En provoquant un accroissement de la mortalité (du fait de l'assèchement d'une grande partie du Lac ainsi que de l'accroissement de la turbidité des eaux mal supportée par de nombreuses espèces), en entravant le développement des jeunes dans les zones d'inondations et en perturbant le cycle migratoire de nombreuses espèces, la sécheresse a contribué à diminuer fortement l'importance du peuplement deltaïque. Par ailleurs, la disparition de grandes espèces telles que Distichodus spp., Citharinus spp., Labeo spp., etc., la disparition des Alestes dentex et A. baremoze, l'importance croissante des Synodontis membranaceus, S. batensoda et autres Mochocidae indiquent des modifications importantes dès à présent visibles dans la composition du peuplement deltaïque.
Les conséquences en ont été pour les pêcheries deltaïques une diminution importante des p.u.e. globales et spécifiques pour les espèces recherchées qui provoquèrent l'abandon de la région deltaïque par les pêcheurs professionnels qui assurent normalement son exploitation.
Manuscrit reçu au S.C.D. de l'O.R.S.T.O.M. le 31 Octobre 1975.
BIBLIOGRAPHIE
Benzecri (J.P.), 1973. — L'analyse des données, Tome I et II. Dunod, Paris.
Blache (J.), 1964. — Les poissons du bassin du Lac Tchad et du bassin adjacent du Mayo Kebbi. Mém. O.R.S.T.O.M., no 4, Paris, 483 p.
Carmouze (J.P.) et al., 1972 — Grandes zones écologiques du Lac Tchad. Cah. O.R.S.T.O.M., sér. Hydrobiol., VI, 2: 103–165.
Durand (J.R.), 1970. — Les peuplements ichtyologiques de l'El Beïd. 1re note: présentation du milieu et résultats généraux. Cah. O.R.S.T.O.M., sér. Hydrobiol., VI, 1 : 3–26.
Durand (J.R.), 1971. — Les peuplements ichtyologiques de l'El Beïd. 2e note: Variations inter et intraspécifiques Cah. O.R.S.T.O.M., sér. Hydrobiol., V, 2 : 147–159.
Durand (J.R.), Franc (J.), Loubens (G.), 1973. — Clefs Longueur-Poids pour 58 espèces de poissons du Bassin du Lac Tchad. O.R.S.T.O.M. N'Djamena, 37 p. multigr.
Durand (J.R.), Loubens (G.), 1970. — Observations sur la sexualité et la reproduction des Alestes baremoze du bas Chari et du Lac Tchad. Cah. O.R.S.T.O.M., sér. Hydrobiol., IV, 2 : 61–81.
Lebart (C.), Fénelon (J.P.), 1973 — Statistique et informatique appliquées. 2e édition. Dunod Paris.
Lemoalle (J.), 1974. — Bilan des apports en fer au Lac Tchad. (1970–1973). Cah. O.R.S.T.O.M., sér. Hydrobiol., Vol. VIII, no 1 : 35–39.
Loubens (G.), Franc (J.), 1972. — Étude méthodologique pour la récolte de statistiques de pêches basées sur l'observation de pêcheries d'un bief du delta du Chari. O.R.S.T.O.M., N'Djamena. 44 p. multigr.
Loubens (G.), 1973. — Production de la pêche et peuplements ichtyologiques d'un bief du delta du Chari. Cah. O.R.S.T.O.M., sér. Hydrobiol., VII, 3/4 : 209–233.
Lauzanne (L.), 1972. — Régimes alimentaires des principales espèces de poissons de l'Archipel oriental du Lac Tchad. O.R.S.T.O.M. N'Djamena, 19 p. multigr.
par/by
Vincent BENECH - Jacques LEMOALLE - Jacques QUENSIÈRE
Hydrobiologistes de l'O.R.S.T.O.M., B.P. 65, N'Djamena (Tchad).
Cah. O.R.S.T.O.M., sér. Hydrobiol., vol. X, no 2, 1976: 119–130.
Résumé
Au cours de la période de sécheresse 1972–74 le lac Tchad s'est scindé en plusieurs bassins qui ont évolué séparément. Des mortalités de poissons sont décrites en fonction des conditions de milieu (profondeur, oxygène, transparence, salinité) dans les différentes régions.
La turbulence au fond, associée à la faible profondeur est le facteur essentiel des mortalités au cours de la baisse du lac. Dans l'archipel de l'est envahi par la végétation, la décomposition des macrophytes cause des mortalités importantes lors de la crue annuelle du lac. Un niveau critique, en dessous duquel les mortalités deviennent probables, a été défini pour la cuvette nord et l'archipel de l'est.
Abstract
Fish mortalities in Lake Chad during the sahelian drought.
During the 1972–74 drought, lake Chad was split into three separate basins as a result of the water level decrease (Fig. 1). The variations of some chemical and physical parameters are described for the northern basin (Fig. 2, 3) and for the eastern archiplago at Bol (Fig. 4,5). Mass fish mortalities have been observed in both basins. They are associated with water turbulence during the decrease of the water level but anoxia due to macrophyte decomposition is the main cause for mortalities during the annual flood at Bol (Fig. 7). Relative catch per unit effort of some species are given (Fig. 6, 8). A critical level, below which mortalities occured, has been determined for both basins. (See also figure legends in english).
* Reproduit avec l'autorisation de l'ORSTOM (Paris)
Au course de la période 1972–1974, des mortalités importantes de poissons ont eu lieu dans le lac Tchad. Ces phénomènes qui n'avaient pas été observés au cours des années précédentes doivent donc être considérés comme une conséquence de la baisse du niveau du lac depuis 1964 et plus particulièrement de 1972 à 1975.
Le lac Tchad est un lac plat de faible profondeur, dont la superficie et la morphologie sont très sensibles aux variations de niveau (Carmouze et al 1972). Depuis Tilho (1928) on distingue un état « Tchad normal », correspondant à une altitude du plan d'eau de l'ordre de 282 m, et un état « Petit Tchad» du même lac où l'altitude du plan d'eau est inférieure à 279 m. Le Tchad normal présente un plan d'eau unique, bordé d'un archipel au nord et à l'est tandis que dans l'état Petit Tchad deux ou trois bassins, séparés par l'exondation de hauts fonds, voient leurs niveaux varier individuellement suivant qu'ils sont plus ou moins alimentés par les eaux du Chari. Le dernier état Petit Tchad apparu vers avril 1973, s'est prolongé en 1974 et 1975 (Chouret-Lemoalle, 1974). Les niveaux dans la cuvette nord, dans la région de Bol et dans les eaux libres du sud-est ont évolué séparément (fig. 1), entraînant des variations de conditions de milieu qui ont influé de façon notable sur les peuplements de poissons, en particulier au cours de mortalités en masse.
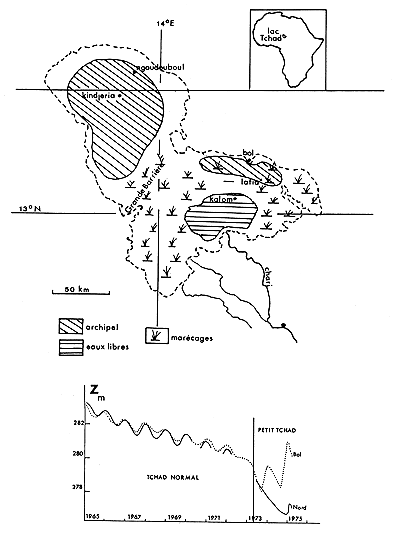
Fig. 1. — Carte schématique du lac Tchad en juillet 1974. Le tireté indique les limites du Tchad normal. Évolution approximative de l'altitude du plan d'eau dans la cuvette nord et à Bol depuis 1965.
Fig. 1. — Lake Chad, july 1974, schematic map. (----) indicates the shoreline during « Normal Chad » periods. The level decrease (absolute altitude) is shown since 1965.
On distingue deux types de mortalités en masse, l'un associé à la turbulence créée par des tornades, l'autre consécutif à la crue annuelle dans l'archipel de l'est. A ces mortalités spectaculaires, mais limitées dans le temps et dans l'espace, se superpose une modification continue des peuplements avec disparitions progressives d'espèces mises en évidence par nos pêches expérimentales.
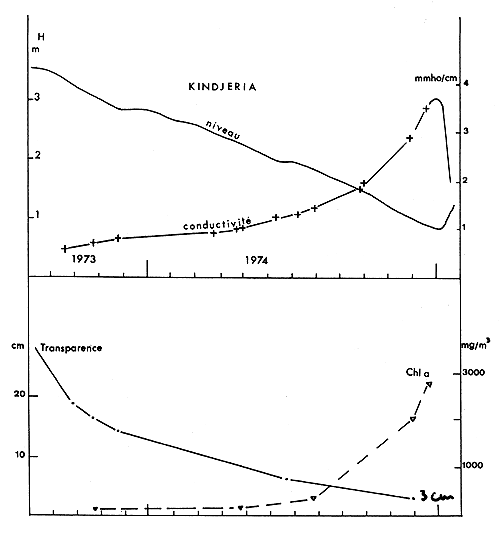
Fig. 2. — Conditions de milieu dans la cuvette nord. En haut: niveau à l'échelle de Kindjéria (d'après le service hydrologique O.R.S.T.O.M.) et conductivité. En dessous: transparence au disque de Secchi et chlorophylle.
Fig. 2. — Northern basin: level and conductivity (above) Secchi disc transparency and chlorphyll a (under).
1. LES CONDITIONS DE MILIEU
1.1. Les paramètres étudiés
Le facteur principal, auquel nous rapporterons les autres paramètres étudiés, est le niveau dont les variations depuis 1966 sont représentées figure 1.
Les facteurs secondaires étudiés sont :
La diminution de la transparence des eaux du lac est une conséquence directe de l'influence croissante, sur les sédiments, de la turbulence de l'eau due au vent. La remise en suspension du sédiment superficiel, de plus en plus importante au cours de la diminution de la hauteur d'eau, agit sur les particules minérales et sur le phytoplancton en voie de sédimentation. On sait (Ganf, 1974 a, 1974 b) qu'une fraction importante du peuplement algal sédimenté, et notamment des cyanophycées, reste en vie assez longtemps. Il s'ensuit donc une augmentation des concentrations en phytoplancton et de la production primaire dans la zone euphotique et de la respiration dans toute la colonne d'eau. Alors que la transparence était auparavant principalement limitée par des particules minérales fines (Lemoalle, 1973), la plus grande turbulence des périodes de bas niveau a maintenu en suspension des particules plus grosses et des quantités plus importantes de phytoplancton qui ont joué un rôle non négligeable dans l'absorption de la lumière et dans les variations des concentrations de l'oxygène dissous.
En 1973 le passage du lac à l'état de Petit Tchad et l'abaissement plus rapide du niveau dans les différentes cuvettes a en outre fait apparaître, notamment dans la cuvette nord, une concentration notable des sels dissous jusqu'alors moins prononcée. Par la suite, la cuvette nord et la région de Bol ont suivi une évolution faisant intervenir les mêmes phénomènes mais à des degrés divers, leur évolution hydrologique ayant été différente.
1.2. Les conditions de milieu dans la cuvette nord
— Niveau et transparence
En avril 1973, l'exondation de la Grande Barrière a privé la cuvette nord de ses apports, la transformant en bassin fermé où l'évaporation était de l'ordre de 2,2 m par an. Le niveau a baissé de façon importante, pour une profondeur moyenne de la zone étudiée passant de 3,3 m à 0,5 m entre septembre 1973 et décembre 1974.
Au cours de la même période, la transparence mesurée au disque de Secchi est passée de 20 à 3 cm (fig. 2) marquant la phase finale d'une diminution constante depuis les valeurs supérieures à 1 m relevées en 1967. La valeur mesurée de 3 cm correspond à 2,1 g/l, de seston (poids sec), la concentration en chlorophylle a (non corrigée pour les phéopigments) étant alors supérieure à 2,5 mg/l.
— L'oxygène dissous
Les variations de concentrations en oxygène dissous sont, en partie, une résultante des conditions de turbulence. En période de Tchad normal, et encore en 1973, la cuvette nord était du type polymictique chaud avec stratification diurne occasionnelle, la tension d'oxygène étant généralement de 90 à 95 % de la saturation le matin, à un moment où la colonne d'eau était homogène (Lemoalle, 1976). En 1974, le régime nycthéméral des vents, la photosynthèse d'un phytoplancton de plus en plus dense et la consommation d'oxygène par un seston abondant, se sont combinés de façons diverses. Il en est résulté une variabilité très grande des teneurs en oxygène, aussi bien dans le temps que dans l'espace, même à courte échelle.
On a notamment observé à N'Goudouboul, en juin 1974, au nord-est de Kindjéria, l'influence d'une forte concentration de phytoplancton en période de vents faibles, avec des variations de 20 mg O2/l en 12 heures, à 10 cm sous la surface (fig. 3). La consommation d'oxygène par le milieu, de 3,9 mg O2/l. h, créait une anoxie pendant une partie de la nuit, les Tilapia respirant à la surface de l'eau, près du rivage. Un ciel couvert pendant une seule journée pouvait alors suffire à créer des conditions encore plus drastiques dues à une diminution de la photosynthèse.
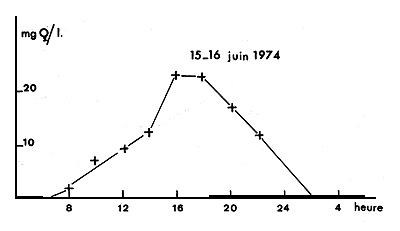
Fig. 3. — Exemple de variation nycthémérale de l'oxygène de l'eau de surface à N'Goudouboul (cuvette nord).
Fig. 3. — An example of diel oxygen concentration in surface waters in N'Goudouboul (northern basin).
A Kindjéria, en décembre 1974, la consommation d'oxygène par le milieu était, à une température de 22 °C, de 1,24 mgO2/l. h. La concentration en oxygène, in situ, était de l'ordre de 1,0 à 3 mg/l au lever du jour et très variable d'un point à un autre au cours de la journée (de 5 à 12 mg/l à 16 h. pour une température de l'eau de 22 °C).
— Les sels dissous
A ces conditions d'oxygénation difficiles s'est ajoutée, dans la cuvette nord du lac Tchad, une augmentation importante de l'alcalinité. Une certaine régulation saline existe, en effet, en période de Tchad normal, qui fait que grâce à des précipitations chimiques et des infiltrations (Carmouze, 1969 - Roche, 1973) la salinité de ce lac endoréique reste faible. Avec l'état Petit Tchad est apparue, dans la cuvette nord, une phase de concentration des sels qui suit approximativement les lois d'un bassin d'évaporation. La conductivité et l'alcalinité étant étroitement liées, nous n'avons représenté que les variations de la conductivité (fig. 2). Celle-ci a augmenté de 900 à 3 500 μmhos/cm entre mars et décembre 1974, pour un pH de 8,5 à 9. A la fin de cette période les ions carbonate, bicarbonate et sodium sont de très loin dominants, formant plus de 90 % des sels dissous (alcalinité 41 mé/l, pH 9). Le CO2 libre dissous est resté négligeable.
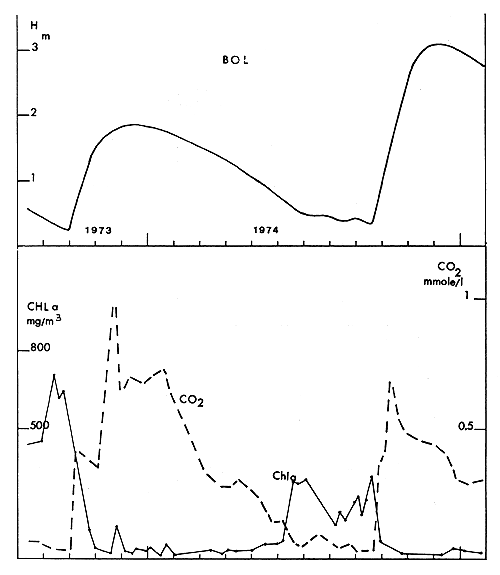
Fig. 4. — Conditions de milieu à Bol (d'après le service hydrologique O.R.S.T.O.M.), chlorophylle et CO2 libre dissous.
Fig. 4. — Level at Bol gauge (above) free dissolved CO2 and chlorophyll a (under) at Bol.
1.3. Les conditions de milieu dans la région de Bol
— Niveau et transparence
La baisse de niveau à Bol a suivi une évolution semblable à celle de la cuvette nord, avec exondation du seuil des îlots bancs à la limite de l'archipel et isolement de la masse d'eau considérée en avril 1973. L'assèchement par évaporation en bassin fermé ne s'est prolongé que jusqu'en septembre pour reprendre ensuite d'avril à septembre 1974 après une remise en eau correspondant à la crue du Chari de 1973 (fig. 4).
L'évolution générale de la transparence et du phytoplancton s'est manifestée à Bol jusqu'en septembre 1973 où l'on mesurait 8 à 10 cm au disque de Secchi (Lemoalle, 1974). Durant la période de basses eaux de 1973, une végétation semi-aquatique très dense (Cyperus papyrus, Aeschynomene, Vossia), a recouvert tous les sédiments exondés. Au cours des remises en eau postérieures, l'eau du Chari a dû traverser cette barrière végétale et est arrivée à Bol dépourvue de particules minérales ou organiques. Les masses d'eau circulant encore dans l'archipel une partie de l'année, la filtration est restée effective et la transparence a conservé des valeurs comprises entre 35 et 80 cm, avec des concentrations en chlorophylle inférieures à 300 mg/m3 (fig. 4).
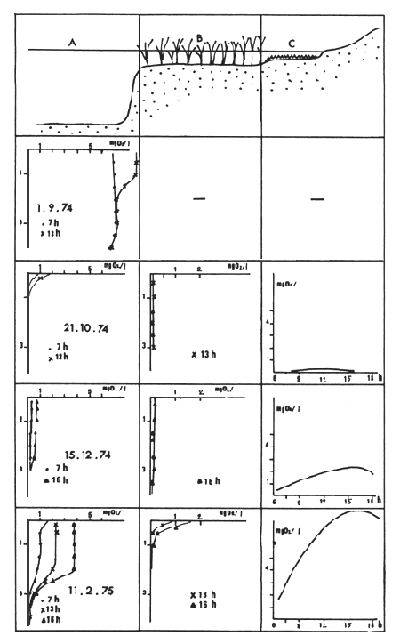
Fig. 5. — Profils d'oxygène à Bérim en pleine eau (milieu A), dans les macrophytes (milieu B) et variations diurnes d'oxygène dans la zone de bordure peu profonde (milieu C).
Fig. 5. — Oxygen profiles at Berim in open waters area (A) in the macrophytes (B) and diel variations near the shore (C).
— L'oxygène dissous
La surface d'eau réduite, abritée par de hautes îles ceinturées de macrophytes, est moins sensible au vent que les vastes étendues d'eaux libres de la cuvette nord, parsemées d'îles plates et nues. Au cours des périodes de bas niveau de la région de Bol le phytoplancton et la turbulence n'ont pas atteint les fortes valeurs constatées dans la cuvette nord. Cependant des tornades ont pu recréer en 1973 et 1974 des conditions proches de celles observées après la tornade du 8 août 1972 décrite plus loin.
Au début d'octobre 1973 les eaux de crue du Chari, avant de parvenir à Bol, ont traversé une barrière de végétation jeune, en pleine croissance et de densité moyenne. De plus, la crue n'a pas été très forte et peu de plantes ont été totalement submergées. Au contraire, en 1974, une crue plus forte (fig. 4) a traversé la même barrière, beaucoup plus dense, dont certains éléments étaient déjà morts. La remontée du niveau a aussi été plus importante et a submergé un grand nombre de plantes, notamment les papyrus. Les macrophytes morts et ceux qui ont été submergés sont entrés dans un cycle de décomposition dont le premier effet a été une absorption quasi totale de l'oxygène dissous dans l'eau. Ce déficit en oxygène a été longtemps dominant et ce n'est qu'en février, soit 4 mois après la crue, qu'ont été mesurées des quantités d'oxygène atteignant le minimum considéré comme nécessaire pour beaucoup de poissons : 3 mg O2/l en surface dans les zones dégagées (notées A sur la figure 5). Dans les régions colonisées par la végétation (notées B sur la figure) l'oxygène est resté, en moyenne, très faible. Il existait toutefois un micro-milieu (noté C) moins défavorable à la vie aquatique : entre les franges végétales et le rivage des îles, on a trouvé fréquemment une zone de quelques mètres de largeur, de 5 à 30 cm de profondeur, colonisée par des algues benthiques dont la photosynthèse active produisait de l'oxygène. La nuit, le fort rapport entre la surface de l'eau et son volume favorisait une légère oxygénation. De tels milieux abritaient, entre autres, des quantités de très jeunes Tilapia.
— Les sels dissous et le CO2 libre
L'isolement de la région de Bol-Bérim en 1973 et 1974 ayant été beaucoup moins long que dans le nord et entrecoupé de remises en eau (fig. 4), le facteur de concentration a donc été moins fort et la conductivité n'a pas dépassé 570 μmhos. L'alcalinité est ainsi restée modérée et le seul fait nouveau est l'apparition de CO2 libre dissous en concentration notable, due à la décomposition des macrophytes lors des remises en eau de 1973 et 1974.
2. LES MORTALITÉS DE POISSONS
Trois types principaux de mortalités peuvent être définis. Les deux premiers sont spectaculaires et concernent les mortalités massives, limitées dans l'espace et dans le temps. Le troisième, plus discret, mis en évidence par des pêches régulières aux filets maillants (Benech, 1975) correspond à une disparition progressive de différentes espèces résultant de l'évolution continue de certains paramètres de milieu dont les fluctuations particulièrement brusques et accidentelles causent les mortalités des premiers types. La diminution de certains effectifs se produisant le plus souvent en milieu fermé (cuvette nord ou zone de Bol isolées du reste du lac par des seuils exondés), une migration ne peut être invoquée pour expliquer les variations.
2.1. Les mortalités en masse consécutives à des tornades
Le 7 août 1972, nous avons observé une tornade à Bol. Sous l'effet d'un vent très violent, le sédiment a été perturbé, les masses d'eau se sont déplacées. Du jour au lendemain la concentration en phyto-plancton a presque doublé, la transparence a diminuée de 14 à 9 cm au disque de Secchi, la concentration en oxygène dissous, au matin du 8 août, était inférieure à 0,5 mg/l (5 % de la saturation). Les poissons sont morts en quantité, les plus petits mourant les premiers au cours de la nuit, les plus grands mourant dans la matinée du 8. La possibilité de telles mortalités était bien connue des populations riveraines qui ont ramassé les poissons tout au long de la nuit et de la matinée du lendemain.
Le même phénomène s'est répété dans l'archipel en 1973 et 1974. En juin 1973, des mortalités de Lates niloticus, Hydrocynus brevis et H. forskalii ont été observées dans les mêmes conditions à Lafia, à 15 km au sud-est de Bol. Le mois suivant, sur les berges du lac à Bol, de nombreuses carcasses de Synodontis spp, espèces dédaignées par les riverains, attestaient d'une autre mortalité en masse.
Dans la cuvette nord, des hécatombes de Tilapia ont été remarquées à la suite d'une tornade fin juin 1974 dans une zone de 0,8 à 1 m de profondeur, à 18 km au sud de Kindjéria. Ce phénomène se répétant plusieurs fois au cours de la saison des pluies en des endroits différents, on pouvait observer en septembre, sur les rives au vent de nombreuses îles, des franges de poissons morts qui atteignaient 0,5 à 1 m de largeur sur plusieurs centaines de mètres de longueur.
2.2. Les mortalités au cours de la crue annuelle du lac
Alors que le niveau de la cuvette nord a été constamment décroissant en 1973 et 1974, la région de Bol a subi, dans la même période, deux phases d'assèchement suivies chacune d'une remise en eau brutale. A la différence de celle de 1973, la crue de 1974 s'est accompagnée de mortalités importantes de nombreuses espèces qui venaient respirer près de la surface avant de mourir. Les Synodontis (en majorité S. balensoda), qui représentaient 45 % des captures le mois précédent ont alors totalement disparu en quelques jours (fig. 6) tandis que le nombre d'espèces péchées, déjà réduit à la suite de la saison des pluies, passait brutalement de 23 à 7 (fig. 7).
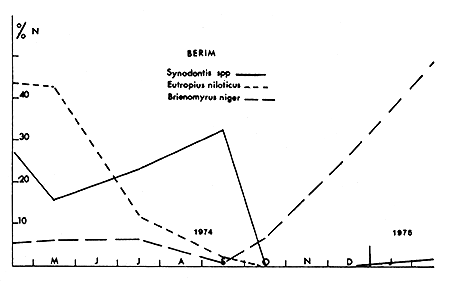
Fig. 6. — Variations des prises par unité d'effort pour quelques espèces à Bol-Bérim (N est la somme des prises, pour chaque genre, au cours de la période décrite).
Fig. 6. — Variations in catch per unit effort for some species at Bol-Berim. N is the sum of the catches, for each genus, during the period under study.
2.3. Disparition progressive de certaines espèces
Aux mortalités brutales, localisées et apparentes, se superpose une mortalité plus diffuse, mise en évidence par les pêches expérimentales. Ainsi certaines espèces bien représentées dans les peuplements de hautes eaux à Lafia au sud de Bol (Grandes zones écologiques, 1972), ont disparu des prises après 1973. Ce sont :
Lates niloticus
Hydrocynus forskalii
Hydrocynus brevis
Alestes dentex
Citharinus citharus
Synodontis membranaceus
Dans la cuvette nord, à Kindjéria, les espèces qui ont disparu de nos relevés de pêche entre décembre 1973 et décembre 1974 sont :
Heterotis niloticus
Hydrocynus forskalii
Hydrocynus brevis
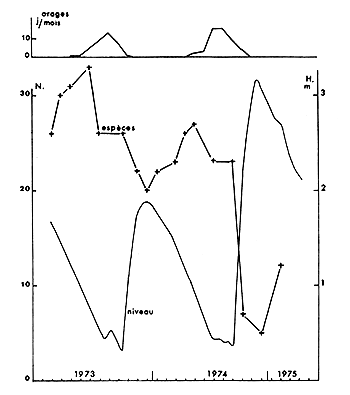
Fig. 7. — Nombre de jours d'orage (en haut), niveau de l'eau et nombre d'espèces capturées au cours des pêches expérimentales à Bol-Bérim.
Fig. 7. — Monthly number of rainy days (above), water level and number of species caught by our experimental fisheries at Bol-Berim.
Citharinus citharus
Tetraodon fahaka
Pollimyrus isidori
Mormyrus rume
On a assisté d'autre part à la diminution progressive d'espèces initialement importantes; c'est notamment le cas de Alestes baremose et Hydrocynus forskalii (ce dernier disparaissant totalement en décembre 1974) dont les prises par unité d'effort sont représentées figure 8. La disparition de ces deux espèces d'intérêt économique s'explique par les conditions de milieu et aussi, par des pêches très actives.
Dans l'archipel de Bol, l'évolution d'Eutropius niloticus, du même type au cours de la baisse du niveau en 1974, se termine par une disparition quasi-totale avant l'arrivée de la crue (fig. 6).
3. DISCUSSION
Les mortalités sont liées en premier chef à la baisse générale du niveau de l'eau dans le lac. Nous pouvons donc essayer de définir un niveau critique et examiner comment réagissent certaines espèces pour dégager les facteurs du milieu qui ont une importance.
3.1. Influence de la profondeur. Niveau critique
A Kindjéria, les premières mortalités ont été observées en avril 1974 puis pendant la saison des pluies, mais il est probable, étant donnée la composition spécifique du peuplement à cette époque, que des mortalités ont eu lieu avant cette date soit dans le centre de la cuvette, soit dans les régions moins profondes, plus sensibles au vent, de la périphérie. Une côte de 2,5 m à Kindjéria, ou 6,5 m à Malamfatori (échelle O.R.S.T.O.M.) peut donc être considérée comme critique en moyenne dans l'année. Bien que la bathymétrie du lac soit mal connue, nous pouvons estimer à environ 2 m la profondeur correspondante dans la majeure partie de la surface en eau. Ce niveau critique définit la cote en-dessous de laquelle des mortalités ont toutes chances de se produire au cours de l'année et notamment pendant la saison des pluies. Il ne tient pas compte des tornades exceptionnelles dont l'effet peut être notable pour des profondeurs supérieures. Quand le niveau continue de décroitre, les fortes turbulences deviennent de plus en plus fréquentes et le milieu devient peu à peu hostile à la plupart des espèces.
En 1972, les fortes tornades qui ont causé les premières mortalités dans l'archipel de Bol ont agi sur un plan d'eau encore assez étendu, dépourvu de bordure végétale, à une cote de 2,2 m (échelle de Bol), ce qui correspond à une profondeur générale de 1,5 m. A cette profondeur correspondent des mortalités accidentelles et localisées. Une cote de l'ordre de 2 m à l'échelle, soit une profondeur de 1,3 m peut donc être considérée comme critique pour la région en 1972, c'est à dire en période de lac normal avec un développement réduit de macrophytes. Par la suite la majorité des fonds correspondants a été colonisée par les macrophytes et ne sont restés en eau libre que les endroits les plus profonds, protégés par une importante végétation de bordure.
Les mortalités qui ont eu lieu au cours des saisons des pluies de 1973 et 1974 ont nettement affecté le nombre d'espèces présentes dans le milieu (fig. 7). Elles correspondent à une même cote du plan d'eau pour les deux années : 0,6 m à l'échelle de Bol. Il apparait donc des niveaux critiques différents, pour une même région, suivant l'état morphologique du lac.
En période de « Tchad normal », avant l'assèchement, les profondeurs critiques sont de 2 m dans la région de Kindjéria et environ 1,3 m près de Bol. Dans les deux régions le sédiment est constitué de vases organiques plus ou moins argileuses (Dupont, 1970), et on peut penser que le niveau critique est atteint dans chacune de ces zones quand la turbulence au fond est du même ordre. Celle-ci dépend de la force du vent, de sa zone d'action sur la surface de l'eau et de la profondeur (Smith, Sinclair, 1972). La force du vent pouvant être considérée comme égale, la différence de profondeur critique s'explique par la différence du rayon d'action du vent à la surface de l'eau. L'archipel autour de Kindjéria est en effet constitué d'îles beaucoup plus basses, plus espacées et dénudées que celui de Bol.
La surface des zones en eau intervient aussi dans l'influence des tornades sur les peuplements : les orages étant localisés, leurs effets immédiats le sont aussi. Si la surface en eau est grande relativement à la perturbation (cuvette nord du lac), une recolonisation est possible à partir des zones non touchées, ce qui a peu de chances de se produire à Bol-Bérim, de surface réduite, lorsque la poche d'eau est isolée du reste du lac.
3.2. Réaction des espèces aux variations du milieu
Si l'on compare la composition des peuplements de Kindjéria et de Bérim en fin du cycle d'assèchement, on constate que l'influence de l'assèchement et de la turbulence a privilégié les mêmes groupes d'espèces : Polypterus, Synodontis et Tilapia qui représentent environ 80 % des captures. Dans les deux biotopes, S. batensoda représente les trois quarts des Synodontis et P. senegalus la quasi-totalité des Polypterus. Ces trois groupes qui persistent ne réagissent cependant pas de la même façon aux conditions de milieu.
Les Synodontis ne se sont pas reproduits ; bien qu'ils aient subi des mortalités notables, leur résistance relative et la concentration des eaux ont entraîné une augmentation de leur densité plus importante que pour d'autres espèces. Les Polypterus senegalus, fréquents dans les marécages (Blache, 1964) ont la possibilité de respirer l'oxygène de l'air et ont donc peu souffert des anoxies consécutives aux orages. Faute de données sur leur reproduction (les filets maillants capturent peu de jeunes Polypterus), nous ne pouvons que constater leur importance croissante au cours de cette période.
Dans les importantes quantités de poissons morts remarquées en 1974 dans la cuvette nord, les Tilapia étaient largement dominants. Leur capacité de reproduction leur a néanmoins permis de se maintenir et de proliférer, les jeunes devenant progressivement dominants en juin 1974, avant de grandir après la saison des pluies, comme le montre l'évolution des prises suivant la maille des filets (fig. 9). Les mortalités massives notées n'étaient donc pas une indication de disparition de ce groupe comme on aurait pu initialement le croire. Par contre le maintien de Tilapia après la disparition de Lates correspond bien aux résultats de Fish (1956) sur la tolérance relative de ces deux genres à un déficit d'oxygène.
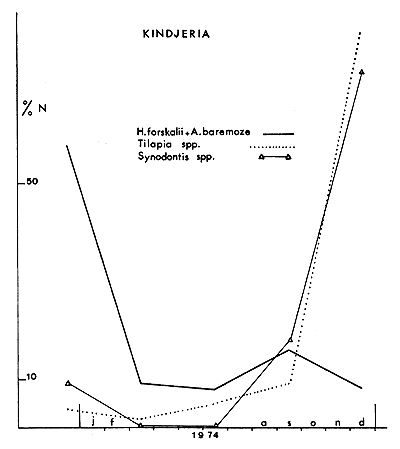
Fig. 8. — Variations des prises par unité d'effort pour quelques espèces à Kindjéria (N est la somme des prises, en nombre, pour les espèces ou les genres considérés, au cours de la périodé décrite).
Fig. 8. — Variations in catch per unit effort for some species at Kindjeria. N is the sum of the catches, for each genus or species association, during the period under study.
Au cours de l'assèchement, la progression de T. galilaea est beaucoup moins sensible que celle de T. nilotica et surtout T. aurea qui ont un développement considérable (tableau I). La première espèce a des besoins en oxygène plus élevés que T. nilotica (Johnson, 1974) et d'après Welcomme (1964) les besoins des jeunes T. aurea ne représentent que 30 % de ceux de T. galilaea. Les variations dans ce groupe soulignent donc l'importance de l'oxygène dissous dans l'évolution des peuplements.
La mortalité en masse observée à Bol sur le peuplement résiduel au début de la crue de 1974 est un phénomène général qui s'est produit dans tout l'archipel du sud-est avec l'avancée de la crue à travers la végétation. L'absence d'oxygène et une teneur inhabituelle en CO2 libre dissous (fig. 4 et fig. 5) ont causé la mort de la plupart des espèces présentes (fig. 7). Seuls Clarias, Polyplerus, Gymnarchus niloticus et Brienomyrus niger ont subsisté. Les trois premiers ont une possibilité de respiration aérienne (cf. Carter in Brown, 1957) ; le dernier cité, fréquent dans les mares en voie d'assèchement, est particulièrement rustique (Blache, 1964).
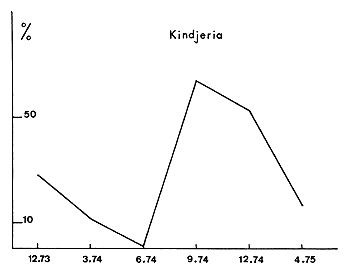
Fig. 9. — Pêches expérimentales à Kindjéria. Pourcentages en nombre des prises de Tilapia dans les filets à petites mailles (11 et 15 mm) par rapport aux prises dans les filets de 11, 15, 35 et 40 de mailles. Les jeunes apparaissent pendant la saison des pluies et se développent ensuite.
Fig. 9. — Experimental fisheries at Kindjéria. Percent ratio of Tilapia caught in small gillnets (11 and 15 mm meshbar) to the catches of Tilapia in gillnets of 11, 15, 35 and 40 mm mesh size. The young fishes appear at the beginning of the rainy season.
Tableau I
Pourcentage en nombre des Tilapia par rapport à l'ensemble des poissons capturés dans une même pêche. Pêches effectuées à Kindjéria à l'aide d'une batterie de filets maillants.
| Espèces | Dates | |||||
| 12/3 | 3/74 | 6/74 | 9/74 | 12/74 | 4/75 | |
| Tilapia nilotica | 1 | 3 | 3 | 5 | 13 | 7 |
| Tilapia aurea | ε | ε | 3 | 15 | 17 | 50 |
| Tilapia galilaea | 2 | 1 | 1 | ε | 5 | 0 |
Ce phénomène, observé en 1974 dans l'archipel, s'est vérifié début 1975 dans la Grande Barrière. Il avait déjà été décrit par Freydenberg (1908) au cours d'une crue semblable, dans les mêmes régions.
Les mortalités dues à la dégradation de la végétation ont été rapportées par plusieurs auteurs dans les pays tropicaux lors de la pénétration d'eau de marécage dans une rivière (Tait, 1965); c'est également ce qui peut se passer dans les régions tempérées lors de la décomposition des amas de feuilles mortes (Larimore, Childers, 1959).
Le déficit en oxygène peut en outre être rendu plus efficient du fait d'autres conditions de milieu telles que la salinité, l'alcalinité, la turbidité ou la présence de substances toxiques. Tous les facteurs défavorables ont un effet cumulatif et, si le seuil léthal n'est pas atteint, des effets néfastes à long terme peuvent se faire sentir sur le métabolisme, la croissance et la reproduction. On sait en effet que les poissons sont capables d'adaptation progressive à des conditions défavorables et que le seuil léthal est différent suivant que les variations d'oxygène (Farnier, Beanish, 1969) de salinité (Erichsen-Jones, 1964) ou de température sont plus ou moins brutales.
Parmi ces facteurs autres que le manque d'oxygène, nous avons noté en particulier deux exemples spécifiques sur Alestes nurse et sur les Mormyridae. En décembre 1974 un examen des Alestes nurse capturés à Kindjéria mettait en évidence des ouïes irritées, couvertes d'un épais mucus dont la présence, qui gêne fortement la respiration, est attribuée à l'action conjuguée de l'alcalinité élevée et de la forte teneur en suspensions des eaux (Morgan, 1972). En période de Tchad normal, l'extension des Mormyridae se limitait aux eaux de conductivité inférieure à 400 μmhos, évitant les régions les plus concentrées (Grandes zones écologiques du lac Tchad, 1972). Au cours de l'année 1974, la conductivité à Kindjéria a varié de 800 à 3 500 μmhos, plaçant ainsi les Mormyridae loin de leur préférendum.
3.3. Comparaison avec le lac Chilwa
Les milieux lacustres à assèchement périodique plus ou moins accentué possèdent en général une ichtyofaune peu diversifiée, adaptée aux variations des conditions de milieu. C'est le cas du lac Chilwa qui abrite 13 espèces dont trois importantes: Clarias mossambicus, Tilapia shirana chilwae, endémique et particulièrement tolérante vis à vis de la concentration ionique et de l'oxygène, et Barbus paludinosus qui vient au lac par les affluents (Morgan et Kalk, 1970).
Les premières mortalités en masse au lac Chilwa ont été observées au cours de tornades en période de niveau décroissant, et sont devenues ensuite de plus en plus fréquentes au cours de la baisse de niveau du lac. Une concentration en oxygène de 0,3 à 0,4 mg/l apparaît léthale pour T. s. chilwae qui est d'autre part résistant à la concentration en Na Cl mais beaucoup moins à l'alcalinité (qui domine dans le lac Tchad). D'une façon générale, l'évolution des conditions de milieu a été du même type dans le lac Tchad et le lac Chilwa, où la cause première des mortalités est la baisse du niveau entraînant une turbulence inhabituelle au niveau du fond. Celle-ci entraîne à son tour une diminution de l'oxygène dissous dont les effets sont aggravés par une turbidité importante.
Les mortalités importantes liées au bas niveau de l'eau sont plus fréquentes pour le lac Chilwa que pour le lac Tchad; les dernières ont été reportées en 1955, 1960 et 1966 au lac Chilwa alors que les bas niveaux correspondants au lac Tchad ont eu lieu vers 1908, 1942 et 1974. Le lac Tchad est de plus relié à un vaste réseau fluvial et offre des milieux plus variés que le lac Chilwa, aussi abrite-t-il en temps normal une faune variée comprenant plus de 80 espèces (Blache, 1964).
Cette diversité favorise la recolonisation des différents milieux du moment, les remplacements d'espèces dominantes et la prolifération de nouvelles espèces se faisant très rapidement en des endroits divers du lac. Un fait tout à fait remarquable est la très grande mobilité des pêcheurs qui parviennent dans une certaine mesure à exploiter cette plasticité du peuplement malgré l'évolution rapide du milieu.
CONCLUSION
En milieu peu profond les mortalités de poissons liées à la turbulence sont un phénomène bien connu, parfois naturel à la suite d'orages, parfois provoqué par l'homme au cours de pêches d'épuisement dans des mares résiduelles. L'observation de mortalités massives et les pêches expérimentales ont permis de définir approximativement des niveaux critiques en-dessous desquels les mortalités se produisent. Le niveau de l'eau étant le facteur essentiel, les paramètres associés, qui traduisent l'instabilité écologique du milieu, permettent de comprendre, dans une certaine mesure, les mécanismes de ces mortalités. Certaines espèces étant plus sensibles que d'autres, une étude détaillée de l'évolution des peuplements pourra éventuellement conduire à la définition de niveaux critiques différents suivant les populations concernées.
Manuscrit reçu au S.C.D. de l'O.R.S.T.O.M., le 22 juillet 1976.
RÉFÉRENCES
Benech (V.), 1975.— Croissance, mortalité et production de Brachysynodontis batensoda (Pisces, Mochocidae) dans l'archipel sud-est du lac Tchad. Cah. O.R.S.T.O.M., sér. Hydrobiol., vol. IX, no 2, 91–103.
Blache (J.), 1964. — Les poissons du bassin du Tchad et du bassin adjacent du Mayo Kebbi. Étude systématique et biologique. O.R.S.T.O.M. Paris, 483 p.
Carter (G.S.), 1957. — Air breathing, in The physiology of fishes, Vol I, M. E. Brown ed., Academic Press New York, 447 p.
Carmouze (J. P.), 1969. — La salure globale et les salures spécifiques des eaux du lac Tchad. Cah. O.R.S.T.O.M., sér. Hydrobiol. vol. III, no, 2, 3–14.
Carmouze (J. P.) et al, 1972. — Grandes zones écologiques du lac Tchad. Cah. O.R.S.T.O.M., sér. Hydrobiol., vol. VI, no 2, 103–169.
Chouret (A.), Lemoalle (J.), 1974 — Évolution hydrologique du lac Tchad durant la sécheresse (1972 à 1974). O.R.S.T.O.M., N'Djaména, 12 p. multigr.
Dupont (B.), 1970. — Distribution et nature des fonds du lac Tchad (nouvelles données). Cah. O.R.S.T.O.M., sér. Géol., vol. II, no 1, 9–42.
Erichsen-Jones (J. R.), 1964. — Fish and river pollution. Butterwork and Co. London.
Farnier (G. J.), Beanish (F. W. H.), 1969. — Oxygen consumption of Tilapia nilotica in relation to swimming speed and salinity. J. Fish. Res. Bd Canada, 26, 2807–2821.
Fish (G. R.), 1955. — Some aspects of the respiration of six species of fish from Uganda. J. Exp. Biol., 33, 1, 186–195.
Freydenberg (H.), 1908. — Le Tchad et le bassin du Chari, F. Schmidt, Paris; 187 p.
Ganf (G. G.), 1974. — Incident solar irradiance and under-water light penetration as factors controlling the chlorophyll a content of a shallow equatorial lake (Lake George, Uganda). J. Ecol., 62, 593–609.
Ganf (G. G.), 1974. — Diurnal mixing and the vertical distribution of phytoplancton in a shallow equatorial lake (Lake George, Uganda). J. Ecol., 62, 611–629.
Johnson (R. P.), 1974. — Synopsis on biological data on Sarotherodon galilaeus. F.A.O. Fisheries Sinopsis No 90.
Larimore (R. W.), Childers (W. F.), 1959. — Destruction and reestablishment of stream fish and invertebrates affected by drought. Trans. Amer. Fish, Soc. 88, 261–285.
Lemoalle (J.), 1973. — L'énergie lumineuse et l'activité photosynthétique du phytoplancton dans le lac Tchad. Cah. O.R.S.T.O.M., sér. Hydrobiol., vol. VII, no 2, 95–116.
Lemoalle (J.), 1974. — L'activité photosynthétique du phytoplancton en relation avec le niveau des eaux du lac Tchad (Afrique). Verh. Internat. Verein. Limnol.
Lemoalle (J.), 1976. — Éléments sur la température des eaux du lac Tchad à Bol. O.R.S.T.O.M., N'Djaména 24 p. Multigr.
Morgan (A.), Kalk (M.), 1970. — Seasonal changes in the waters of lake Chilwa (Malawi) in a drying phase 1966–68. Hydrobiologia, 361, 81–103.
Morgan (P. R.), 1972. — Causes of mortality in the endemic Tilapia of lake Chilwa. Hydrobiologia, 40, 1, 101–119.
Roche (M. A.), 1973. — Traçage naturel salin et isotopique des eaux du système hydrologique du lac Tchad. O.R.S.T.O.M., Paris, 398 p.
Smith (I. R.), Sinclair (I. J.), 1972. — Deep water waves in lakes. Freshwat. Biol., 2, 4, 387–399.
Tait (C.), 1965. — Mass fish mortalities. Fish, Res. Bull. Sept. Game and Fisheries, Zambia, 3, 28–30.
Tilho (J.), 1928. — Variations et disparition possible du lac Tchad. Ann. Géographie, 37, 238–260.
Welcomme (R. L.), 1964. — The habitats and habitat preferences of the young of the lake Victoria Tilapia (Pisces, Cichlidae). Revue Zool. Bot. Afr., 70, 1–28.
The Committee devoted much of its time to the study of the dramatic situation which had developed in Lake Chad area and the repercussions on future development plans of that basin. Aware that a multi-donor fishery project formulation mission was scheduled to visit Chad in the Spring of 1978, the Sub-Committee, on the basis of the documentation and expertise available at its Third Meeting, prepared the following guidelines for the mission:
On the basis of available information and trends it is likely that a situation of “small Chad” will prevail for some time. Consequently, the opening of the Great Barrier to increase the water surface would only increase the evaporation rate with resulting damage to fish production.
In view of the important reduction in water area and the uncertainty of the future of the lake in the short and medium term, increase in the fishing effort should be encouraged only after very careful study of specific situations. Measures to improve fish processing, conservation and marketing should be given priority as well as efforts to develop fisheries in unexploited zones such as the Salamat.
The crucial role of floodplains for the breeding and hence the recruitment of fish to stock both Lake Chad and the Chari-Logone rivers is emphasized. Any intervention, human or natural, that would lead to the flood regime to be altered in such a way as to produce a reduction of the floodplain area or change in the timing of the flood would be detrimental to fisheries. This makes it essential that research on the Yaérés floodplains be undertaken without delay to ascertain this importance and role in the future of the fisheries of the basin.
It is evident that the lack of regulation to govern the exploitation of the basin gives rise to serious problems which continue to hamper fishery development in the area. Priority should therefore be given to harmonize fishery regulation and strengthen national and regional organizations interested in the fishery resources of the basin. This will require important socio-economic studies of the fishermen populations, the movement of the fishermen and the products and the marketing systems.
The important amount of research already done on the fisheries hydrology of Lake Chad points to the need of bringing together this wealth of documentation in a synthesis of the evolution of lake Chad and the fisheries stocks during the last ten years with scientific guidelines for its future exploitation and development. Such a study would constitute an important background document to the multi-donor project formulation mission.
![]()
![]()
![]()