![]()
![]()
![]()
por
NICOLAS CRUZ MOREJON
Jefe, Departamento Control de Capturas
y
EMYGDIO CADIMA
Biólogo Pesquero de la FAO
Centro de Investigaciones Pesqueras
Instituto Nacional de la Pesca
Playa Habana, Bauta, Cuba
Extracto
Se determinan algunas relaciones biológicas con interés práctico. Se presentan las relaciones largo total - largo cefalotórax, largo total - peso; y los factores de conversión de peso de cola de camarón y peso de camarón seco a peso de camarón entero fresco para las especies Penaeus duorarum y Penaeus schmitti.
LENGTH AND WEIGHT RELATIONSHIPS OF SHRIMP CAUGHT ON THE CONTINENTAL SHELF OF CUBA
Abstract
Certain biological relationships of practical interest are determined. The ratios for total length/size of cephalothorax and total length/weight are given; conversion factors for changing weight of shrimp tails and weight of dry shrimp into weight of whole fresh shrimp for the species Penaeus duorarum and Penaeus schmitti are given.
RAPPORT ENTRE LA LONGUEUR ET LE POIDS DES CREVETTES CAPTUREES SUR LE PLATEAU CONTINENTAL CUBAIN
Résumé
Les auteurs ont établi quelques relations biologiques présentant un intérêt pratique. Ils décrivent les rapports longueur totale/longueur du céphalothorax et longueur totale/poids et indiquent pour les espèces Penaeus duorarum et Penaeus schmitti, les coefficients de conversion pondérale des queues de crevettes et des crevettes sèches en crevettes entières fraîches.
En este trabajo se han determinado las relaciones: largo peso, largo cefalotórax -largo total, y peso cola - peso total de las especies de camarón Penaeus duorarum y P. schmitti.
Los datos que fueron utilizados para estas determinaciones provienen de muestras tomadas a bordo del barco “CAMARON I” por la Sección de Camarones del CIP durante el período de abril a diciembre de 1965 y también de ejemplares estudiados en julio de 1965 en la Cooperativa de Manzanillo, especialmente para las relaciones peso de camarón cola-peso de camarón entero fresco y peso de camarón seco - peso de camarón entero fresco.
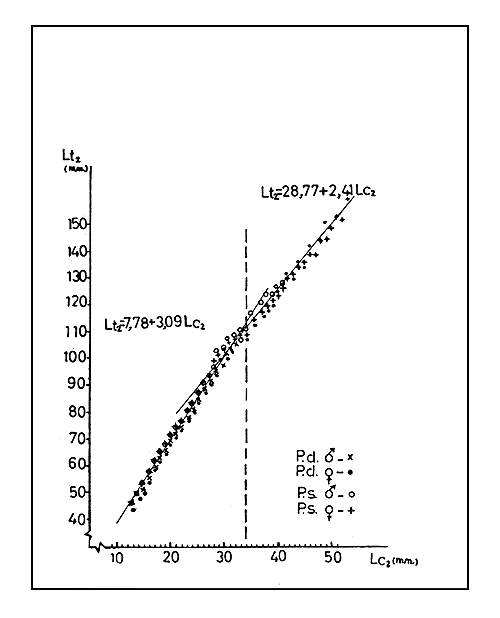
2.1 Relación largo cefalotórax - largo total
Los largos del cefalotórax (Lc2) son medidos a partir de la base de la escotadura post-orbital hásta el punto medio dorsal del borde exterior del cefalotórax.
Los largos totales (Lt2) son medidos desde la base de la escotadura post-orbital hasta el borde posterior del útlimo somito abdominal en su punto medio dorsal.
Las mediciones fueron realizadas con un pie de rey y aproximadas al milímetro más próximo.
Los datos fueron agrupados por especies y sexos y clasificados en clases de un milímetro según el largo de cefalotórax (Cuadro I).
Las distribuciones de los largos totales de cada sexo y especie en cada clase de Lo2 son presentados en la Fig. 1.
La Figura sugiere que existe una relación entre el largo total y el largo del cefalotórax independientemente del sexo o especie. La relación parece ser curvilínea.
Con todo, las observaciones pueden ser ajustadas por dos líneas rectas, una para los puntos correspondientes a los valores menores de Lo2, por ejemplo, inferiores o igual a 34 mm, y otra para los valores superiores.
Para obtener las ecuaciones de esas rectas utilizamos las técnicas de regresión lineal.
Se calcularon las dos rectas de regresión para cada grupo de sexos y especies y no se verificaron diferencias significativas entre sexos y especies.
Las rectas que por tanto se ajustan a los datos son:
| Para Lc2 menores o igual a 34 mm | Lt2 = 7,78 + 3,09 Lc2 |
| Para Lc2 mayores de 34 mm | Lt2 = 28,77 + 2,41 Lc2 |
La relación largo cefalotórax - largo total, puede tener la importancia práctica de poder facilitar la medición de los largos del camarón. Así, se hace más rápida la medición del largo del cefalotórax que la del largo total; largos del cefalotórax pueden ser, siempre que sea necesario, convertidos a largo total por medio de las ecuaciones dadas anteriormente.
CUADRO I
Distribución de largos de cefalotórax y largo total
| Penaeus duorarum | Penaeus schmitti | |||||||
| Lc2 P. medio | Lt2 Media | Frec. | Lt2 Media | Frec. | Lt2 Media | Frec. | Lt2 Media | Frec. |
| ♂ | ♀ | ♂ | ♀ | |||||
| 13 | 46,5 | 4 | 44,3 | 7 | 47,1 | 1 | ---- | -- |
| 14 | 50,1 | 8 | 48,2 | 5 | 50,6 | 3 | ---- | -- |
| 15 | 52,2 | 24 | 51,5 | 17 | 55,0 | 7 | 53,9 | 17 |
| 16 | 54,9 | 18 | 54,7 | 25 | 58,9 | 17 | 57,8 | 32 |
| 17 | 59,9 | 19 | 58,7 | 23 | 61,6 | 31 | 61,4 | 36 |
| 18 | 62,9 | 52 | 61,8 | 42 | 65,0 | 67 | 64,8 | 85 |
| 19 | 65,7 | 100 | 64,9 | 72 | 68,3 | 74 | 67,8 | 100 |
| 20 | 69,0 | 168 | 68,3 | 101 | 71,7 | 98 | 71,4 | 108 |
| 21 | 72,0 | 157 | 71,2 | 126 | 74,9 | 73 | 74,2 | 112 |
| 22 | 74,4 | 148 | 74,3 | 140 | 77,9 | 68 | 77,0 | 73 |
| 23 | 78,5 | 131 | 78,8 | 122 | 81,1 | 35 | 80,1 | 81 |
| 24 | 81,8 | 102 | 80,8 | 117 | 83,9 | 38 | 83,3 | 32 |
| 25 | 84,0 | 61 | 83,4 | 103 | 87,0 | 21 | 86,2 | 28 |
| 26 | 87,6 | 40 | 86,2 | 65 | 91,9 | 13 | 90,0 | 26 |
| 27 | 90,7 | 28 | 89,1 | 74 | 93,7 | 18 | 92,4 | 20 |
| 28 | 92,7 | 21 | 92,7 | 63 | 97,1 | 18 | 99,4 | 16 |
| 29 | 96,5 | 14 | 95,7 | 65 | 101,4 | 17 | 97,9 | 11 |
| 30 | 97,7 | 9 | 99,3 | 50 | 104,5 | 21 | 100,7 | 13 |
| 31 | 104,0 | 1 | 101,4 | 43 | 105,8 | 40 | 104,9 | 13 |
| 32 | 105,7 | 3 | 104,3 | 40 | 109,2 | 69 | 107,9 | 16 |
| 33 | --- | -- | 106,7 | 42 | 110,9 | 85 | 107,9 | 16 |
| 34 | 107,9 | 35 | 110,9 | 16 | 113,8 | 63 | 111,6 | 20 |
| 35 | --- | -- | 112,7 | 51 | 116,7 | 58 | 112,4 | 31 |
| 36 | --- | -- | 113,9 | 23 | 117,9 | 27 | 114,9 | 25 |
| 37 | --- | -- | 117,2 | 23 | 121,7 | 18 | 117,0 | 32 |
| 38 | --- | -- | 119,3 | 24 | 124,5 | 10 | 119,2 | 43 |
| 39 | --- | -- | 121,3 | 8 | 124,0 | 2 | 121,9 | 28 |
| 40 | --- | -- | 125,2 | 11 | --- | -- | 123,7 | 33 |
| 41 | --- | -- | 127,3 | 9 | 128,3 | 3 | 126,3 | 22 |
| 42 | --- | -- | 130,3 | 10 | --- | -- | 129,7 | 24 |
| 43 | --- | -- | 131,0 | 2 | --- | -- | 131,4 | 17 |
| 44 | --- | -- | 135,0 | 4 | --- | -- | 134,4 | 15 |
| 45 | --- | -- | 134,0 | 1 | --- | -- | 135,9 | 12 |
| 46 | --- | -- | 142,0 | 2 | --- | -- | 139,1 | 7 |
| 47 | --- | -- | --- | -- | --- | -- | 138,8 | 4 |
| 48 | --- | -- | --- | -- | --- | -- | 144,0 | 9 |
| 49 | --- | -- | 151,0 | 1 | --- | -- | 144,4 | 9 |
| 50 | --- | -- | --- | -- | --- | -- | 149,0 | 1 |
| 51 | --- | -- | --- | -- | --- | -- | 153,0 | 1 |
| 52 | --- | -- | --- | -- | --- | -- | 152,0 | 2 |
| 53 | --- | -- | --- | -- | --- | -- | 160,0 | 1 |
 |  |
| Fig. 1 Relación largo total-largo cefalotórax. | Fig. 2 Relación logaritmo peso - logaritmo largo. |
2.2 Relación largo total - peso camarón entero
Los camarones en las muestras fueron clasificados por sus largos totales en clases de 5 mm, con puntos medios de 43 mm, 48 mm, 53 mm, etc. En cada muestra los camarones de cada clase fueron pesados en conjunto (Cuadro II).
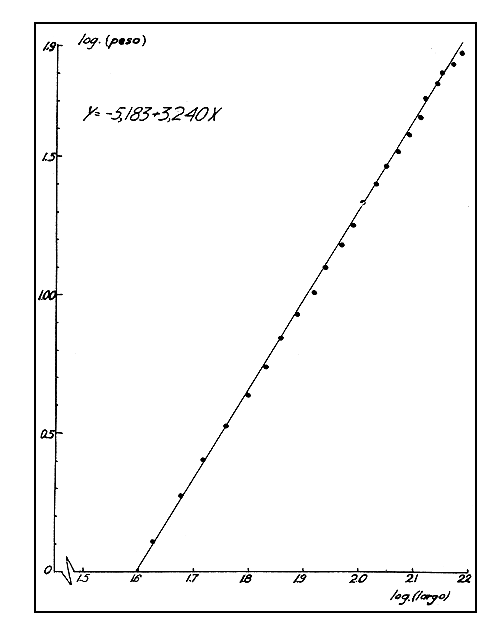
La representación gráfica de los logaritmos de los pesos medios de cada clase contra los logaritmos de los puntos medios de las clases correspondientes sugiere que una línea recta se ajusta bien a los puntos observados (véase Fig. 2).
La ecuación de la recta calculada por los métodos de regresión lineal ponderada fue:
Y = -5,183 + 3,240 X
donde, “Y” es el logaritmo del peso y “X” es el logaritmo del largo.
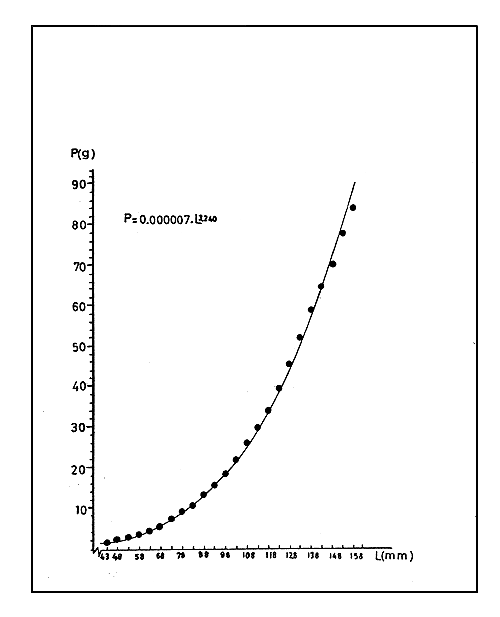
La ecuación puede ser presentada en términos del peso (P) y del largo (L):
P = 0,000007 L3,240
donde “P” es expresado en gramos y “L” en mm (véase Fig. 3).
La importancia práctica de la relación largo - peso es evitar el pesaje de camarones, operación siempre diffcil de realizar especialmente a bordo de los barcos. Con el uso de esta relación se pueden convertir en peso los largos observados y, por tanto, transformar, por ejemplo, una composición por largos en una composición por pesos.
En muchas situaciones no será necesario obterner los pesos, ya que bastará conocer cantidades proporcionales a los mismos. En tales casos la conversión será simplificada, siendo suficiente calcular las cantidades L3,240 de la ecuación de la sección 2.2.
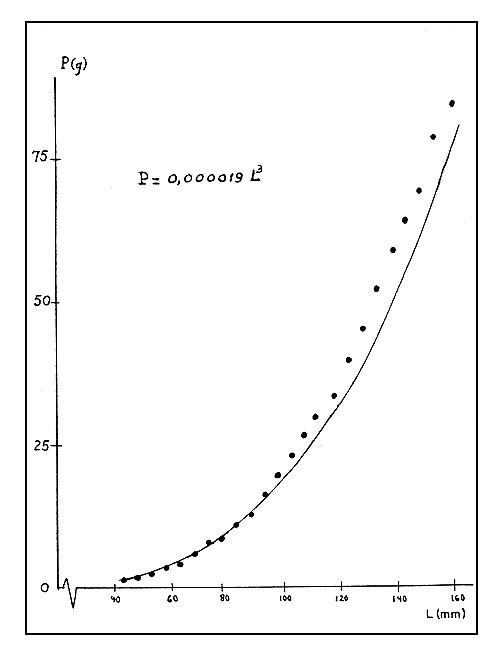
Para el análisis de las poblaciones de camarones puede ser útil tener una relación entre el peso y el largo, similar a la ecuación dada anteriormente, pero con el exponente de “L” igual a 3. Nos referimos por ejemplo a la posibilidad de utilizar la ecuación de crecimiento de Bertalanffy y los métodos y tablas existentes para los cálculos sobre capturas máximas sostenibles, etc. Así, determinamos también la ecuación:
P = 0,000019 L3
Para apreciar el ajuste de esta ecuación a los datos observados presentamos la Fig. 4. Esta ecuación subestima los pesos de los camarones menores y sobrestima los pesos de los camarones mayores. La utilización de esta ecuación deberá ser hecha teniendo en consideración estas diferencias.
Para su comparación presentamos enseguida algunas relaciones obtenidas por otros autores:
Relación largo - peso de la forma
P = cLn :
| Especie | Valor de n | Autor | |
| Leander styliferus = Palaemon styliferus | 2,87 | Kunju, M.M., | 1956 |
| Leander serrifer = Palaemon serrifer | 3,25 | Ynowye, A., | 1949 |
| Pandalus kessleri | 3,14 | Kubo, I., | 1951 |
| Metapenaeus joyneri | 2,95 | Ykematsu, W., | 1955 |
| Hymenopenaeus muelleri | 3,20 | Angelescu, V. y E.E. Boschi, | 1959 |
 |  |
| Fig. 3 Relación largo-peso. | Fig. 4 Relación largo - peso con n = 3. |
CUADRO II
Distribución de largos totales y pesos
| Largos P. Medios | Pesos Frec. Medias | |||
| 43 | 7 | 1,3 | ||
| 48 | 23 | 1,9 | ||
| 53 | 92 | 2,6 | ||
| 58 | 118 | 3,4 | ||
| 63 | 290 | 4,5 | ||
| 68 | 595 | 5,6 | ||
| 73 | 670 | 7,1 | ||
| 78 | 495 | 8,8 | ||
| 83 | 404 | 10,6 | ||
| 88 | 223 | 12,8 | ||
| 93 | 171 | 15,3 | ||
| 98 | 149 | 18,1 | ||
| 103 | 154 | 21,7 | ||
| 108 | 229 | 25,6 | ||
| 113 | 260 | 29,6 | ||
| 118 | 183 | 33,6 | ||
| 123 | 102 | 39,1 | ||
| 128 | 68 | 45,0 | ||
| 133 | 42 | 52,0 | ||
| 138 | 25 | 58,7 | ||
| 143 | 16 | 64,0 | ||
| 148 | 6 | 69,8 | ||
| 153 | 3 | 77,3 | ||
| 158 | 1 | 84,0 | ||
2.3 Factores de conversión de peso de camarón cola y camarón seco a peso de camarón entero fresco
Para la obtención de estas relaciones más que una muestra al azar nos interesa que la muestra cubra la diversidad de pesos posibles. Así, los datos fueron obtenidos tomando camarones de diversas tallas. Los métodos de separar las colas y de secar los camarones fueron usados por los pescadores. El camarón seco se obtiene cociéndolo entero en agua con sal, y posteriormente secándolo al sol; después de seco se saca el carapacho.
Los camarones fueron agrupados por especies y sexos y clasificados por los largos totales en clases de 5 mm. Los pesajes se realizaron en grupos de 10 camarones y cuando su número era inferior a 10 se pesaron todos ellos.
La representación gráfica de los pesos enteros contra los pesos de camarón cola y camarón seco muestra una tendencia lineal (Figs. 5 y 6). Las ecuaciones de regresión lineal pasando por el origen fueron calculadas y se presentan a continuación:
| P. duorarum | P. schmitti | |||
| Camarón cola (X) a Camarón entero (Y) | ♂ Y=1,59X | ♀ Y=1,73X | ♂ Y=1,61X | ♀ Y=1,69X |
| Camarón seco (X) a Camarón entero (Y) | Y=4,53X | Y=3,92X | Y=4,78X | Y=4,04X |
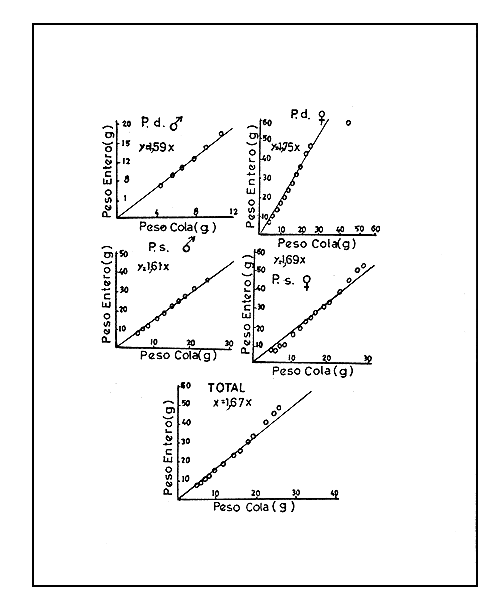 | 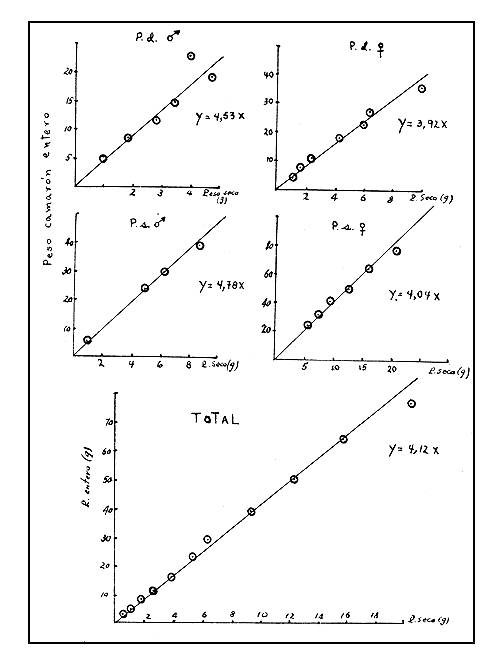 |
| Fig. 5 Relación peso camarón cola-peso entero. | Fig. 6 Relación peso camarón seco - peso entero fresco. |
Las rectas de regresión, que pasan por el origen y que fueron obtenidas sin distinción entre sexos y especies, son:
Y = 1,67 X para camarón cola-camarón entero
Y = 4,12 X para camarón seco-camarón entero fresco
Las relaciones peso cola - peso entero fresco y peso camarón seco - peso entero fresco son útiles ya que permiten obtener los factores de conversión correspondientes, que deberán ser aplicados a los desembarcos de camarón en cola o seco a fin de convertir esos totales en camarón entero fresco y poder adicionarlos a los desembarques de camarón entero.
Los factores de conversión son los coeficientes de regresión cuando las rectas pasan por el origen. Así, en Cuba se adopta como factor de conversión de peso cola a peso entero fresco el valor 1,67 y el factor de conversión de peso de camarón seco a peso entero fresco el valor 4,12.
Las relaciones entre largo total y largo de cefalotorax de las dos especies de camarones - P. duorarum y P. schmitti - fueron calculadas obteniéndose:
Lt2 = 7,78 + 3,09 Lc2 para valores de Lc2 inferiores o iguales a 34 mm
y
Lt2 = 28,77 + 2,41 Lc2 para valores de Lc2 superiores a 34 mm
El peso de los camarones en el Golfo de Guacanayabo puede ser obtenido a partir del largo total por medio de la relación:
P = 0,000007.L3,240
El factor para convertir el peso de cola de camarón en peso de camarón entero fue determinado obteniéndose el valor 1,67.
El factor para convertir el peso de camarón seco en peso de camarón entero fue calculado obteniéndose el valor 4,12.
Las ecuaciones y factores obtenidos en este trabajo no son muy diferentes a las relaciones que han sido determinadas por otros autores para especies similares.
Ninguno de estos resultados debe ser tomado como definitivo. Sería conveniente en el futuro preparar experimentos apropiados para la obtención de una información que permita perfeccionar estas conclusiones.
Angeleseu, V, y E.E. Boschi, 1959 Estudio biológico pesquero del langostino del Mar del Plata en conexión con la operación nivel medio. Buenos Aires, Servicio de Hidrografía Naval, H. 1017:135 p.
Kubo, I., 1951 Bionomics of the prawn Pandalus kessleri Cserniavski. J.Tokyo Univ.Fish. 38(1):1–26
Kunju, M.M., 1956 Preliminary studies on the biology of the Palaemonid prawn, Leander styliferus Milne-Edwards. Proc.Indo-Pacif.Fish.Coun., 6(3):404–16
Ykematsu, W., 1955 On the life-history of Metapenaeus joyneri (Miers) in Ariake Sea. Bull. Jap.Soc.scient.Fish., 20(11):969–78
Ynowye, A., 1949 A note on the ecology of a common shoreshrimp Leander serrifer. Bull.Jap. Soc.scient.Fish., 15(7):318–22
![]()
![]()
![]()