![]()
![]()
![]()
EIFAC SESSION 1/ENERGY/R/1
METHODS FOR DETERMINATION OF ENERGY REQUIREMENT IN FISH
C. Young Cho
Fisheries Policy Branch, Ontario Ministry of Natural Resources and Dept. of Nutritional Sciences,
University Guelph, Guelph, Ontario, Canada NIG 2WI
Determination of energy requirements for fish is no different than for other homeotherms. Procedures for energy balance (CN-method) and comparative carcass analysis (CCA-method) are the same for fish as for other animals. There are differences in techniques if respirometry is used because fish use dissolved oxygen from water.
Fish biologists have emphasized either measurement of oxygen consumption for physical activity (i.e. swimming) or the measurement of more questionable energy costs of specific functions such as “mechanical component of specific dynamic action” and “heat increment by diet and fish density” in studies of bioenergetics. Energy balance using the whole animal concept has been rarely studied for fish. This review emphasizes studies of nutritional energetics on a whole animal basis.
The largest portion of heat energy from the metabolizable energy (ME) intake is used to sustain life: the maintenance energy cost (HEm), in both poikilotherms and homeotherms. This requirement takes priority over other energy requirements. The widely accepted formula for calculating HEm in homeotherms is 70 BWkg0.75 kcal/day (Kleiber, 1961), however, there is much confusion for fish. NRC (1983) and Smith et al. (1978) reported 57.6 BWkg0.63 kcal/day (a) for 4–57 g rainbow trout at 15°C, 32.4 BWkg1.0 (b) for 1–4 g and 48.8 BWkg0.75 (c) for 1–57 g. Cho (1990) found HEm = (-1.04+3.26T-0.05T2) • BWkg0.824kJ/day where T = water temperature (°C) as shown in the table:
Approx. HEm Requirement of Rainbow Trout at 15°C and Homeotherms (KJ/day)
| Body Weight (g) | Poikilotherms | Homeotherms | |||
|---|---|---|---|---|---|
| Cho 1990 | NRC, 1983 | Smith, 1978 | Kleiber, 1961 | ||
| (a) | (b) | (c) | |||
| 1 | .1 | - | .1 | 1.1 | 1.6 |
| 5 | .5 | 8.6 | .7 | 3.8 | 5.5 |
| 10 | .8 | 13.2 | - | 6.5 | 9.3 |
| 50 | 3.1 | 36.5 | - | 21.6 | 31.0 |
| 100 | 5.5 | - | - | - | 52.1 |
| 1000 | 36.6 | - | - | - | 293.0 |
Paradoxically, the NRC (1983) data indicated that poikilotherms need more energy than homeotherms for HEm. Heat increment of feeding (HiE) can also be measured with the calorimetry techniques. Any excess of ME intake over HEm is used for recovered energy (RE) retained as tissue gain. It can be determined most accurately by the CCA-method. Data from direct and indirect calorimetry and from the CCA-method will be discussed.
EIFAC SESSION 1/ENERGY/E/1
MEASUREMENT OF ENERGY REQUIREMENTS FOR MAINTENANCE AND SUBSTRATE UTILIZATION IN RAINBOW TROUT USING INDIRECT CALORIMETRY AND GROWTH TRIALS
F. Médale*, F. Vallée and S. J. Kaushik
Laboratorie de Nutrition des Poissons, INRA, BP3, Station d'Hydrobiologie,
64310, Saint Pée-sur-Nivelle, France
Four experimental diets were formulated as follows:
| HPHE | LPHE | HPLE | LPLE | |
|---|---|---|---|---|
| Digestible protein (mg/g DM) | 371 | 326 | 367 | 328 |
| Digestible energy (kJ/g DM) | 20.4 | 18.3 | 16.2 | 14.1 |
| DP/DE (mg/kJ) | 18.2 | 17.8 | 22.7 | 23.2 |
Apparent digestibility of nutrients and energy was determined using the indicator method. Maintenance energy needs of rainbow trout (IBW: 30 g) at 17°C and substrate utilization were estimated both through growth trials and through indirect calorimetry.
Growth trials lasted 8 weeks. Duplicate groups of 50 fish each were fed one of the experimental diets at 0.4, 1 or 2% BW/day or near to satiation. Two other groups were fasted. Fish were weighed every two weeks and rations were re-evaluated. Maintenance energy needs estimated from weight gain using the model of Mercer (1982) were not significantly different between the diets. The average value was 56 kJ DE/kg showing that the maintenance requirements were affected neither by the levels of digestible protein nor by the dietary DP/DE ratios.
Indirect calorimetric measurements were conducted on duplicate groups of 20 fish each in a thermoregulated recirculated water system. O2 uptake, CO2 output, N-urea and N-ammonia excretion were measured during three consecutive 24-hour cycles after a one-week period of adaptation to the diet and to the feeding level. Energy expenditures were not significantly different between the diets. The values were slightly lower but close to those obtained from growth trials. Less than 30% of energy for maintenance was brought by proteins with the diet LPHE and above 40% with the diet HPLE. Lipids were the main energy source except for diet HPLE which contained a high level of digestible carbohydrate. Carbohydrates within this diet were efficiently used as energy sources at the expense of lipids.
EIFAC SESSION 1/ENERGY/E/2
A FACTORIAL APPROACH TO THE ESTIMATION OF METABOLIZABLE ENERGY REQUIREMENT OF GROWING TROUT (ONCORHYNCHUS MYKISS)
J.M. Gropp *, F. Beck and Annette Schuhmacher
Institut für Ernährungsphysiologie, Veterinärstr. 13, D-8000 München, Germany
Most recent data on nutrient or energy requirement of farm and domestic animals are calculated on a factorial basis. The net energy expenditure of starved trout was analyzed statistically by Beck and Gropp (1988) using the data of eight starvation experiments. Energy losses showed a distinct dependence on fish mass and water temperature. The nitrogen corrected energy loss [J * fish-1* d-1] was described as: 22.09* e(1.034T) *FM[g]0.833 (FM=fish mass:8–400g, T = water temperature [°C], p <0.001, r2 = 0.887). The basal metabolism of fish deprived of feed for several weeks is probably near but not equal to standard metabolism because of higher activity. For growing fish fed several times daily, maintenance net energy expenditure corresponds more to routine metabolism, which exceeds standard metabolism by 60% (Brett and Groves, 1979). The energy loss of trout after starvation should therefore be adapted to the higher level of routine metabolism by ca. 30% to derive net energy requirement for maintenance.
The net energy requirement for growth can be calculated from the energy content of fish mass gain. On the basis of published data, the exponential regression equations of Weatherly and Gill (1983) are used because they are based on energy determined by bomb calorimetry. Thus net energy requirement of growing trout can be calculated for several fish body masses, temperatures and growth rates by simple addition. The difference between net energy and the (higher) figures for metabolizable energy is caused by the “heat increment” energy losses. Heat increment for growing trout is reliably estimated by Brett and Groves (1979) and Schwalb-Bühling & Gropp (1982) with 6–14% of digestible energy, figures varying with feed composition and water temperature. For generalization, a heat increment amounting to 15% of net energy may be assumed.
REQUIREMENT FOR METABOLIZABLE ENERGY (ME) OF TROUT AT DIFFERENT BODY MASS, TEMPERATURE AND GROWTHRATE
| Fish mass [g] | 50 | 100 | 200 | |||||||||
| Water temp. [°C] | 7 | 12 | 7 | 12 | 7 | 12 | ||||||
| Growth rate [α as %] | 1,0 | 1,5 | 1,0 | 1,5 | 1,0 | 1,5 | 1,0 | 1,5 | 1,0 | 1,5 | 1,0 | 1,5 |
| ME [kJ * d-1] | 6,9 | 8,8 | 8,2 | 10,4 | 13,6 | 17,6 | 16,6 | 21,4 | 26,9 | 35,4 | 33,8 | 44,5 |
The data seem a reliable basis for estimating the energy requirements of growing trout. Metabolizable energy is preferred compared to digestible energy in expressing feed energy, because digestible energy varies remarkably with water temperature. The factorial approach described above for trout seems also applicable for calculating energy requirement of other fish species.
EIFAC SESSION 1/ENERGY/E/3
A NEW METHOD FOR DETERMINING DIGESTIBLE ENERGY IN FISH
J.Opstvedt*, T.Hjertnes and A.Aksnes
Norwegian Herring Oil and Meal Research Institute,
N-5033 Fyllingsdalen - Bergen, Norway
A method for determining digestible energy in fish was developed in rainbow trout. The method can be used on single dietary components and complete diets and can also be applied to components which may only be included at low levels in the diet. Further, the method allows for testing effects of levels of inclusion on digestibility. The method uses a standard diet composed of casein (60%), fish oil (5%), soybean oil (5%), dextrinized maize starch (22%) and added vitamins and minerals. In the test diet casein is substituted for the test ingredients. Faeces collection is done by stripping and digestibility is determined indirectly using chromium oxide as an indicator. Digestible energy of casein was determined in three consecutive trials by substitution of 10 and 20 % casein for cellulose and ascertaining the covariance between levels of casein and digestible energy of the total diet. Tests for homogeneity showed no significant differences in coefficient of regression (b) between experiments with a mean value equivalent to 18.7 ± 0.021 MJ digestible energy per kg of casein. Subsequent trials tested the content of digestible energy in one sample of fish meal and two samples of soya products. At dietary inclusions of 15, 25, 30 and 50%, as substitution for casein, the fish meal was determined to contain (mean ± sd) 22.3 ± 0.4, 20.8 ± 0.2, 20.0 ± 0.4 and 20.0 ± 0.2 MJ per kg of dry matter. Similarly, the two samples of soy products had 12.9 ± 1.1 and 11.1 ± 0.95, 12.6 ± 0.3 and 10.2 ± 0.7 and 10.9 ± 0.1 and 10.2 ± 0.1 MJ per kg of DM when included at levels of 15, 30 and 45% respectively in the diets. Thus there was a tendency for the determined digestible energy to be higher when the test sample was present at low levels (i.e. about 15%) compared with at higher levels. Only for fish meal was this effect significant.
EIFAC SESSION 1/ENERGY/E/4
DETERMINING DIGESTIBLE ENERGY IN INDIVIDUAL COMPONENTS OF RAINBOW TROUT DIETS
M. Rodehutscord*, S.L. Kinzinger, H.G. Schulte and E. Pfeffer
Institut für Tierernährung, Endenicher Allee 15, D-53115 Bonn, Germany
Feed compounders aiming to produce mixtures of defined energetic value require corresponding information about individual ingredients to be used. As most ingredients cannot be fed alone, their respective energy value has to be estimated by using the difference method.
Digestibility trials with rainbow trout (Oncorhynchus mykiss) were carried out using the indirect method with Chromium oxide as a marker. Faeces were collected from anaesthetized trout by mildly pressing the body from the ventral fins to the anus. Groups of trout with a total body mass of about 60 kg were kept in plastic tanks of 2000 I capacity at a constant water temperature of 15°C. Each experimental diet was given to two or three replicate tanks twice a day to satiation.
Energy digestibility from lysine-supplemented wheat gluten was found to be constant irrespective of feeding intensity and the proportion of wheat gluten in the diet. Therefore, a basal diet was developed consisting exclusively of wheat gluten supplemented with L-Lysine HCl, vitamins, minerals and Cr2O3. Within the test diets, the basal diet was replaced by either 25 or 50% of a protein source or by 30% fat.
Energy digestibility in components (DQc) was calculated as follows:
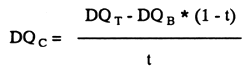 | with DQT: | digestibility of energy in the test diet |
| DQB: | digestibility of energy in the basal diet | |
| t: | proportion of component in test diet |
DE in individual components was (MJ/kg T):
| % component in test diet | 25 | 50 | 30 |
|---|---|---|---|
| soy beans, hydrothermical treated | 20.4 | 19.6 | |
| wheat gluten | 21.0 | 21.0 | |
| feather meal, hydrolysed | 20.9 | 19.3 | |
| grieves | 22.9 | 21.0 | |
| field beans, untreated | 7.7 | 5.7 | |
| field beans, autoclaved | 11.8 | 10.1 | |
| field peas, untreated | 8.6 | 7.1 | |
| field peas, autoclaved | 11.3 | 6.8 | |
| herring-, sunflower- and soy bean oil | 39.5 | ||
| beef tallow | 28.1 |
EIFAC SESSION 2/PROTEIN, AMINO ACIDS/R/2
PROTEIN AND AMINO ACID REQUIREMENTS - A CRITIQUE OF METHODS
C.B. Cowey
Department of Nutritional Sciences, University of Guelph, Ontario NI G 2WI Canada
Protein requirements have been determined by dose-response curves with weight gain the response measured. When expressed as a proportion of the dry matter in the diet, requirement values obtained have been high - even for herbivorous species. Questions centre around whether the diets used were isoenergetic, the rates of growth obtained and the method used (if any) to analyse the data. Kim et al. (1991; Aquaculture, 96:57–67) recently reevaluated the protein requirement of rainbow trout (Oncorhynchus mykiss) and drew attention to the fact that differences in estimated protein requirements between different laboratories may be due to the energy contents and energy sources in the diets.
Measurement of amino acid requirement has also depended primarily on growth experiments. Values obtained have indicated large differences between species and, even for a single species, values obtained from different laboratories show wide differences. These may have arisen from different laboratory conditions (differences in water quality, flow rate and biomass density), the use of different reference proteins as sources of amino acids, of diets containing large amounts of crystalline amino acids, and variation in the protein energy : total energy ratio in the diet. It is doubtful that any, or all, of these factors provide a satisfactory explanation for the observed discrepancies. Ogino (1980; Bull.Jap.Soc.Sci.Fish. 46:171) used the accretion of essential amino acids over a period of 14–28 days to estimate requirements. This method ignores the maintenance requirements of the fish and, in addition, it is difficult to see how it takes account of the fact that only 40 % or so of ingested protein is retained in the body. Oxidation methods, applied after fish have been fed diets containing graded levels of the amino acid under study for several weeks, have been used as a supplementary procedure to growth studies. Kim et al. (1992; Aquaculture, 107:89) question whether the direct oxidation method is appropriate because of variation in the specific activity of the amino acid in the precursor pool. The indirect oxidation method does not appear to suffer from this drawback. Plasma and tissue concentrations of amino acids have occasionally proved useful as confirmatory data to a growth curve but this is not invariably the case. The possibility that a method based on N retention (the concept that the removal of a non-limiting amino acid from the diet will have no effect on N retention while, conversely, reduction in the level of the first limiting amino acid will reduce N retention to the greatest extent Wang and Fuller, 1989; Brit.J.Nutr. 62:77) is feasible with fish will be explored.
EIFAC SESSION 2/PROTEIN, AMINO ACIDS/E/5
USE OF INFIC DATA BASE FOR AMINO ACID REQUIREMENT STUDIES
John E. Halver
School of Fisheries HF-15, University of Washington, Seattle, WA 98195, USA
The International Network of Feed Information Centers (INFIC) database contains amino acid profiles of over 15,000 feedstuffs prepared by various techniques by the countries of the world. Preliminary filtering of appropriate feedstuffs which might be used in fish feed formulations has generated a data base of over 100 commonly used ingredients. Many of these have digestibility coefficients listed. The data base is faulted by the variety of techniques used to measure amino acid profiles, and by many ingredients which have not had digestibility coefficients for the protein component determined. This data base is now being re-examined by the National Research Council to generate more exact values for the indispensable and dispensable amino acids present in feedstuffs. It is anticipated that the data base will be further improved by cooperating laboratories specifically interested in amino acid profiles of various fish feed formulas. Regardless of these deficiencies, the existing INFIC data base and the scheduled improved one, should provide fish feed formulae, chemical amino acid profiles, for least cost and for research diet functions. These data will form a first proximation for satisfying dietary requirements for various fish species used in nutritional requirement studies.
EIFAC SESSION 2/PROTEIN, AMINO ACIDS/E/6
USING WHOLE-BODY AMINO ACID PATTERNS AND QUANTITATIVE REQUIREMENTS TO RAPIDLY DEVELOP DIETS FOR NEW SPECIES
Paul B. Brown
Purdue University, Department of Forestry and Natural Resources,
1159-Forestry Building, West Lafayette, IN 47907-1159, USA
Correlation between whole-body essential amino acid patterns, particularly the A/E ratio, and quantitative dietary requirements for essential amino acids are typically good. The whole body pattern has also been identified as a positive initial step in dietary formulation for new aquaculture species. However, without a quantitative starting point (i.e., a relatively precise requirement for an individual essential amino acid), nutritionists are forced to use quantitative values developed for other species. We have conducted a series of studies that has been designed to decrease the time required to develop diets that contain the appropriate amounts and ratios of essential amino acids. The first study was the identification of an acceptable amino acid test diet. We then used that diet to quantitatively determine the requirements for lysine, methionine + cyst(e)ine, and arginine for hybrid striped bass (Morone saxatilis × M. chrysops). Using the concentrations of essential amino acids in whole fish and the established dietary lysine requirement, A/E ratios adequately predicted the total sulphur amino acid requirement, but underestimated the dietary arginine requirement. Based on our research and published research with channel catfish, both arginine and the total aromatic amino acid requirements will be underestimated when using whole body patterns to predict dietary requirements. However, the use of this series of studies appears to offer the potential of rapidly developing diets for new species.
EIFAC SESSION 2/PROTEIN, AMINO ACIDS/E/7
INDISPENSABLE AMINO ACID REQUIREMENTS OF FISH: CORRESPONDENCE BETWEEN QUANTITATIVE DATA AND AMINO ACID PROFILES OF TISSUE PROTEINS
Muriel Mambrini* and S. Kaushik
Laboratoire de Nutrition des Poissons, INRA, BP 3, Station d'Hydrobiologie,
64310 St Pée-sur-Nivelle, France
Currently available data on the indispensable amino acid (IAA) requirements of different fish species were analyzed and compared to IAA profiles of reference proteins. The requirements determined from dose/response curves show some degree of variability (table). This can be due to a) the kind of experimental diet (semi-purified, practical) b) the nature of the response monitored (growth, plasma amino acid concentration, amino acid oxidation) or c) the type of response analysis (variance analysis, broken line, modelization). It was impossible to detect any difference due to species.
It is advisable that experimental diets be formulated on IAA profiles of proteins of egg (EG), muscle (MU) or carcass (CA) of the species being tested. The IAA profiles of the three groups of proteins from different fish species (136 profiles) were characterized and then compared to the available data on requirements (27 references). A multiple factorial analysis of correspondence was performed, where each IAA was expressed as per cent of total IAA. Muscle proteins were characterized by a higher lysine (18.2%) content than EG (14.9%) and CA (16.6%). Compared to MU and CA, EG contained more leucine (17.4 vs 15.8 and 16.2% for MU and CA respectively). Carcass proteins had higher arginine content (13.8%) than MU (11.5%) and EG (11.0%). No variation associated with species was detected.
Carcass protein IAA profiles showed the best degree of correspondence with the IAA requirements suggesting that they reflect most the ideal IAA pattern. The main differences were for methionine and phenylalanine, the needs being systematically underestimated by the use of IAA profiles of CA, and leucine for which the need was overestimated.
Table: Minimum and maximum IAA requirements of different species of fish (g/16g d'N) found in the literature (n = number of studies)
| IAA | Min | Max | n |
| Arginine | 2.2 | 6.0 | 26 |
| Isoleucine | 1.2 | 4.4 | 16 |
| Lysine | 3.8 | 6.6 | 25 |
| Phenylalanine | 2.0 | 6.5 | 7 |
| Tryptophan | 0.2 | 1.1 | 18 |
| Histidine | 0.9 | 2.7 | 16 |
| Leucine | 2.7 | 8.4 | 16 |
| Methionine | 1.3 | 3.6 | 20 |
| Threonine | 0.6 | 5.0 | 19 |
| Valine | 0.6 | 5.5 | 16 |
EIFAC SESSION 2/PROTEIN, AMINO ACIDS/E/8
AMINO ACID REQUIREMENTS OF RAINBOW TROUT (ONCORHYNCHUS MYKISS)
I. DEVELOPMENT OF THE TEST DIET
M. Rodehutscord*1, M. Pack2, S. Jacobs1 and E.Pfeffer1
1 Institut für Tierernährung, Endenicher Allee 15, D-53115 Bonn, Germany
2 Degussa AG, Applied Technology Feed Additives, P.O. Box 1345, D-63403 Hanau, Germany
For studies on amino acid requirements a test diet was developed giving growth performance, protein deposition and feed/gain ratio similar to practical type diets.
Four consecutive trials with trout (initial body mass 30 – 50 g) were carried out in a recirculating system with 24 plastic tanks of 250 l. Each experimental treatment comprised of four replicate tanks of 20 fish. Diets were prepared as moist pellets using a mincer. Fish were fed twice a day to satiation reaching a final body mass of about threefold of the initial mass. Protein deposition was measured by the comparative slaughter method.
In trial A, the 54% protein (N × 6.25) control diet contained 30% fish meal, 30% wheat gluten and 4.5% glutamic acid. Reduction of protein was achieved by replacing wheat gluten on a DE-basis by a mixture of fat, binder and cellulose. Results show, that a reduction of dietary protein to 37% in dry matter (DM) is possible without a decrease in protein deposition or increase in feed/gain ratio.
Consequently, in trial B dietary protein was reduced in two steps from 38 to 30% in DM as described above. Diets with reduced protein content were tested with or without addition of all essential amino acids in crystalline form. Decrease in protein deposition and feed/gain ratio caused by reduction of dietary protein was restored by the addition of crystalline amino acids. In the resulting 30% protein diet complete replacement of fish meal by wheat gluten, inorganic phosphorus (P) and L-Lysine. HCl did not alter performance of trout (trial C). Results of trial D showed that the use of wheat gluten as sole source of protein can be reduced from 30 to 15% of the diet, if essential and non essential amino acids are added.
In summary, a standardized test diet is available, from which each essential amino acid can be removed individually to create a specific deficiency. Hence, it can be used as a basal diet to determine requirements for potentially limiting amino acids in dose response experiments. The diet consists of 27.5% fat, 23% extruded wheat, 15% wheat gluten, 22% crystalline amino acids, 6% binder and 6.5% premix.
EIFAC SESSION 2/PROTEIN, AMINO ACIDS/E/9
AMINO ACID REQUIREMENTS OF RAINBOW TROUT (ONCORHYNCHUS MYKISS)
II. PROTEIN DEPOSITION AS FUNCTION OF DIETARY METHIONINE,
THREONINE AND ARGININE
M. Pack*1, M. Rodehutscord2, S. Jacobs2 and E. Pfeffer2
1 Degussa AG, Applied Technology Feed Additives, P.O. Box 1345, D-63403 Hanau, Germany
2 Institut für Tieremährung, Endenicher Allee 15, D-53115 Bonn, Germany
The first communication described the development of the test diet. Based on these results, the second contribution reports a series of dose-response experiments to estimate requirements for individual amino acids.
Three growth trials with rainbow trout were performed to determine the influence of methionine, threonine or arginine on weight gain and protein deposition. In order to obtain sensitive data on the dose related effect of each individual amino acid, the respective amino acid under test was removed from the test diet and then added in 12 steps in its crystalline form.
The fish responded very well to increasing dietary content of the respective amino acid. Exponential curves were used to describe the non-linear growth and protein deposition responses. Data from one experiment are shown in Figure 1 as an example, and Table 1 details the exponential response functions determined from the three experiments.
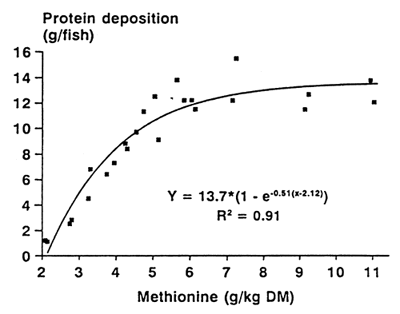 | Table 1: Protein deposition as influenced by dietary methionine, threonine and arginine | ||||
| Exponential function: y = a (1-e-b(x-c)) | |||||
| Amino acid | Estimated parameters | R2 | |||
| a | b | c | |||
| Methionine | 13.7 | 0.51 | 2.12 | 0.91 | |
| Threonine | 14.4 | 0.49 | 4.27 | 0.90 | |
| Arginine | 14.7 | 0.39 | 3.89 | 0.84 | |
| Figure 1: Protein deposition response of rainbow trout to increasing dietary methionine content | |||||
Giving a good fit to the original data, these curves deliver the basis to determine requirement figures depending on the level of fish performance and on the parameter to be optimized. They can also be used as a tool to determine most economical amino acid standards for trout diets
EIFAC SESSION 2/PROTEIN, AMINO ACIDS/E/10
N-TURNOVER OF CARP IN RELATION TO PROTEIN SUPPLY
K.-H. Meyer-Burgdorff and K.D. Günther
Institut für Tierphysiologie und Tierernährung,
Kellner-Weg 6, 3400 Göttingen, Germany
N-turnover of carp (30–60 g) was studied using a diet based on casein, starch and sunflower oil at different regimes of protein supply. Fish were fed on this diet with an optimized P/E ratio (20 mg dCP/kJ DE) at three different feeding levels (A, B, C) or at deviating P/E-ratios (D:17.6; E:23 mg dCP/kg DE). Under consideration of the assumptions made for a two-pool model after a single dose of an identical 15N-labelled casein diet, N-turnover of carp was monitored. Using groups of fishes, N-balance experiments combined with comparative salughter technique were carried out in triplicate at a water temperature of 23°C.
The expression N-flux, calculated as the flow rate through the metabolic amino acid pool, was chosen to characterize the N-turnover of fish. N-flux increased linearly according to the N-intake, when fish were supplied at a fixed P/E-ratio. In relation to N-intake restricted feeding seems to favour re-incorporation of N derived from protein breakdown, resulting in a slightly superior protein utilization efficiency.
A significantly higher level of N-turnover was observed when carps were supplied at suboptimal P/E-ratios. In the case of a raised P/E-ratio (E) this effect was the result of a distinct increase of body protein degradation and subsequent amino acid oxidation. When carps were supplied at a lower P/E-ratio (D) an extremely high N-turnover was due to both, a marked increase of synthesis and breakdown of body protein. A direct comparison at the same level of N-intake (B and D; C and E) confirmed that protein retention decreased as an effect of an increased N-turnover induced by a suboptimal P/E-ratio in the diet. These findings support the concept of an optimal P/E-ratio in fish nutrition.
| A | B | C | D | E | |
| Intake mg N/kg0.8d | 435c | 601b | 756a | 610b | 764a |
| Flux mg N/kg0.8d | 1075e | 1380d | 1679c | 2467a | 2094b |
| Retention mg N/kg0.8d | 200d | 270b | 327a | 238c | 264b |
EIFAC SESSION 2/PROTEIN, AMINO ACIDS/E/11
A NOTE ON THE USE OF PLASMA UREA LEVEL TO VALIDATE THE ARGININE REQUIREMENT ASSESSED BY GROWTH DATA IN SEA BASS (DICENTRARCHUS LABRAX)
E. Tibaldi, Francesca Tulli and D. Lanari
Dípartimento di Scienze della Produzione Animale, via San Mauro 2,33010 Pagnacco (UD), Italia
Responsiveness of urea synthesis to dietary arginine is believed to be predictable in the European Sea Bass (D. labrax) because of the high level of arginase activity in this species (Corti et al., 1985). In rainbow trout fry, Cho et al., (1992) have used post-prandial serum urea level to confirm the arginine requirement assessed by growth parameters. A similar approach was applied in the present study to fingerling sea bass by measuring plasma urea levels.
The experiment started at the end of a seven week trial carried out to assess the arginine requirement of fingerling sea bass (Tibaldi et al., unp. res.). Fish (12–18 g) were kept in 65 1 flow-through tanks (temperature 25°C and salinity 15 ‰) and fed seven isonitrogenous (47% N × 6.25) isolipidic (12% EE) diets containing graded levels of Arginine (L-ARG). A basal diet (diet 1) was formulated to be limiting in arginine (1% by weight). It contained maize gluten (300 g/kg), fish meal (100 g/kg) and mixtures of essential and dispensable AA to simulate the AA composition of sea bass muscle protein. Diets from 2 to 7 were obtained by adding 3, 6, 9, 12, 15 and 18 g/kg of crystalline L-ARG to the basal diet. Diet 4 was used in a preliminary study in order to determine the time pattern of post-prandial changes in plasma urea levels. Fish were withdrawn for blood sampling 1,3,5,7,9,12 and 24 hr after the morning meal. Peak urea level was found 5 hr post-prandial so that all the blood samples were taken 5 hr after the single meal. Blood was collected by severing the caudal peduncle and plasma was stored under liquid N and analysed for urea content within 24 hr. Three fish were sampled for each time period (preliminary study) and per diet.
The arginine requirement of fingerling sea bass estimated by the dose-growth relationship was found to be 4% of the dietary protein. The 5 hr post-prandial plasma urea level varying with the arginine content of the diet and increased from 0.40 ± 0.04 mM (diet 1, ARG content 2.2% of the protein) to 0.61 ± 0.08 mM (diet 2, ARG 2.9%) without any further change up to diet 4 (ARG 4.2%, plasma urea 0.63 ± 0.04 mM). A sharp increase was noted thereafter (± 180%) by elevating the dietary arginine up to 5.4 % of the protein (diet 6, plasma urea 1.11 ± 0.03 mM). The critical dietary arginine value of 4.2% based on plasma urea level was similar to the dietary requirement estimated by growth rate (4%).
EIFAC SESSION 2/PROTEIN, AMINO ACIDS/E/12
PROTEIN AND AMINO ACID REQUIREMENTS IN GOLDFISH LARVAE
(CARASSIUS AURATUS)
Emile D. Fiogbé* and Patrick Kestemont
Unité d'Ecologie des Eaux Douces, Facultés Universitaires ND, de la Paix,
61 rue de Bruxelles, B-5000 Namur, Belgium
Protein and amino acid requirements of goldfish (Carassius auratus) larvae of 8.3 mg initial body weight were estimated based on growth response over 20 days to graded levels of protein ranging from 31 to 60 %. Based on growth performances, the optimum dietary protein requirement of goldfish larvae was estimated to be 51 % of the dry diet. Maintenance requirement in proteins based on endogenous losses during the 20 day feeding trial with the 1.65 % protein diet were about 500 μg/100 mg body weight/day. Amino acid composition of the whole body of goldfish larvae were determined at the start and at the end of the experiment. The profiles of essential amino acid (EAA) requirements were estimated based on daily whole body EAA losses or increments in larvae fed semi-purified dry diets containing 1.65, 31, 39, 45, 53 and 60% protein. As shown in the table, EAA requirements for goldfish larvae are higher than those reported for Cyprinus carpio juveniles. These high values may result from the observed growth of goldfish larvae during the short rearing period (20 day experiment). However, owing to their similarity to body EAA composition of the initial larvae, these results appear to be satisfactory. Indeed, previous studies established a high correlation between whole body amino acid patterns and dietary amino acid requirement patterns for several fish species (Wilson and Poe, 1985; Comp.Biochem.Physiol., 80 B, 385–388).
Table: Essential amino acid (EAA) composition and requirements in goldfish larvae compared to the EAA requirements in common carp C. carpio juveniles
| E.A.A. (% Prot.) | Body composition of goldfish larvae | Requirements of goldfish larvae | Requirements of C.carpio juveniles (Ogino, 1980) |
|---|---|---|---|
| Arg | 8.22 | 8.25 | 3.3 |
| Lys | 7.31 | 8.17 | 4.7 |
| His | 10.61 | 15.88 | 1.2 |
| Ile | 4.18 | 4.69 | 2.0 |
| Leu | 8.45 | 8.49 | 3.7 |
| Val | 5.14 | 5.95 | 2.6 |
| Phe | 4.13 | 5.15 | 2.9 |
| Thr | - | - | 1.8 |
| Met | 2.76 | 3.10 | - |
| Trp | 0.97 | 6.46 | - |
EIFAC SESSION 2/PROTEIN, AMINO ACIDS/E/13
NON-ESSENTIAL AMINO ACID SOURCES IN CRYSTALLINE AMINO ACID DIETS FOR TROUT (ONCORHYNCHUS MYKISS)
J.M. Gropp*1, Annette Schuhmacher1 and M. Münch2
1) Institut für Tierernährung, Gustav-Kühn-Straβe, D-04159 Leipzig, Germany
2) Institut für Ernährungsphysiologie, Veterinärstr. I 3, D-8000 München, Germany
Diets containing a mixture of crystalline amino acids either as the sole source or as a part of dietary protein are an important tool in nutrition experiments. If applied in trials concerning amino acid or vitamin requirements, fish growth is often not satisfactory.
Although the essential amino acids (EAA) and their relations are known and some work has been done concerning the ratio of EAA to the non essential amino acids (NEAA), only poor information is available on the optimal source of NEAA. Therefore a series of experiments with trout fingerlings was conducted in which the source of NEAA was varied. The results of one trial are given below. The composition of the EAA mixture was 10.9% l-lysine, 6.0% dl-methionine, 3.9% l-cystine, 7.4% l-threonine, 1.9% l-tryptophan, 8.3% l-isoleucine, 14.0% l-leucine, 8.3% l-phenylalanine, 7.0% l-tyrosine, 8.4% l-valine, 13.0% l-arginine and 4.0% l-histidine. NEAA were added isonitrogenously by glycine, l-glutamic acid, l-glutamine or mixtures of all of them. The diets containing 15–45% cooked wheat starch, 4–13% fishoil and micro-cellulose were calculated isocalorically (poultry tables).
| Cumulative results after 12 weeks feeding 6 replicates/group, 20 fish/replicate | ||||||||||
| AA-CP [%] | 27 | 45 | 45 | 45 | 45 | 18 | 45 | 45 | 45 | 45 |
| EAA-CP [%] | 27 | 27 | 27 | 27 | 27 | 18 | 18 | 18 | 18 | 18 |
| NEEA-CP [%] | - | 18 | 18 | 18 | 18 | - | 27 | 27 | 27 | 27 |
| NEEA-source | - | gly | glu | gln | ggg* | - | gly | glu | gln | ggg* |
| Initial BM [g] | 47.4 | 47.5 | 48.0 | 47.4 | 47.4 | 47.4 | 47.4 | 47.4 | 47.4 | 47.5 |
| Final BM [g] | 60.1 | 68.7 | 63.3 | 77.1 | 83.2 | 53.4 | 66.8 | 63.7 | 83.6 | 84.1 |
| α [%] | 0.28 | 0.46 | 0.29 | 0.58 | 0.70 | 0.13 | 0.41 | 0.35 | 0.67 | 0.68 |
| Feed intake [g] | 37 | 44 | 40 | 50 | 46 | 33 | 37 | 41 | 56 | 59 |
| Gain/feed | 334 | 522 | 394 | 598 | 811 | 168 | 522 | 393 | 654 | 626 |
| * ggg = gly + glu + gln | ||||||||||
The addition of NEAA to EAA improved growth and feed efficiency evidently. Glutamine was superior to glycine and glycine superior to glutamic acid. However, best results were obtained by adding a mixture of all three NEAA. The obvious changes in trout performance by glutamine could hypothetically be traced back to a better amino acid absorption by a less acidic intestinal milieu. In consequence, the optimal EAA to NEAA ratio should be newly defined.
EIFAC SESSION 2/PROTEIN, AMINO ACIDS/E/14
PLASMA AMINO ACID LEVELS IN RAINBOW TROUT (ONCORHYNCHUS MYKISS)
Annette Schuhmacher*1, M. Goldberg2, J. Schön 2, Chr. Wax 2 and J.M. Gropp1
1 Institut für Tierernährung, Gustav-Kühn-Str. 8, D-7022 Leipzig, Germany
2 Institut für Tierphysiologie, Veterinärstr. 13, D-8000 München 22, Germany
A large number of in vivo as well as in vitro studies to evaluate amino acid absorption and utilization from different sources in fish is published. A multitude of criteria have been applied. Among these the post-prandial plasma amino acid level (PAAL) reflects intestinal passage and digestion of protein, amino acid (AA) availability as well as satisfactory or unsatisfactory supply with AA. But only a few studies deal with plasma amino acid increase (PAAI) as a parameter (Murai et al., 1982, 1984, 1987; Ogata et al., 1986, Walton and Wilson, 1986, and Ash et al., 1989). As a first step, PAAI was determined after force feeding certain amounts of casein, fish protein concentrate (FPC), wheat gluten meal (WGM) and a mixture of crystalline AA to trout (Oncorhynchus mykiss) fasted for two and seven days, respectively. PAAL was measured at least for 48 hours after force feeding as well as on fasting trout. Blood samples were obtained by heart puncture from 3 – 6 trout each at intervals given in the graph below. Plasma AA were determined by HPLC.
Generally a marked PAAI occurred during the first 12 hours after force feeding followed by a rather slow decrease for further 36 hours. Thereafter, PAAL did not significantly differ from fasted fish. In trouts fasted for two days maximum PAAL of the essential AA was reached for casein, FPC and WGM after 18, 6–12 and 18 hours, respectively; peaks of PAAL occurred ca. 6 h later after a fasting period of 7 days. Individual PAAL between 6 and 18 hours after ingestion was closely correlated to AA uptake (p < 0.05). The differences in time, after which PAAL peak was reached, indicate velocity of absorption. They may also reflect a negative correlation between PAAI and the suitability of protein sources. Remarkable differences in PAAL and PAAI were observed for the different essential AA. If individual PAAI are ranged an estimation of limited AA will be possible. It is concluded that for determination of AA availability and utilization, PAAL should be measured at least three times (0, 12 and 18 hours after feeding) with small variations for different protein sources.
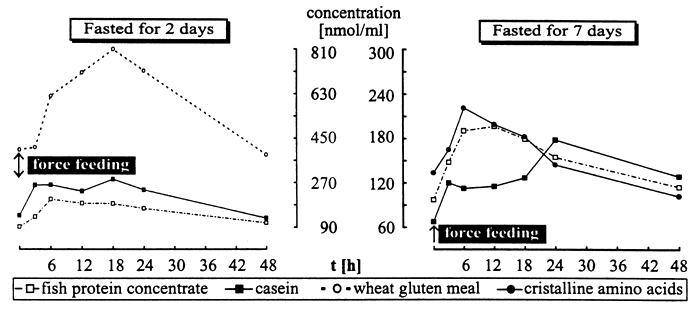
Plasma amino acids (total essential amino acids) after force feeding trout with 0,32 % protein (of body weight) from casein, fish protein concentrate, wheat gluten and cristalline AA, respectively.
EIFAC SESSION 3/DIGESTION, UTILIZATION OF PROTEIN AND ENERGY/E/15
ESTIMATION OF PROTEIN AND ENERGY NEEDS FOR MAINTENANCE AND GROWTH OF NILE TILAPIA (OREOCHROMIS NILOTICUS) USING DIFFERENT CRITERIA
S. J. Kaushik*, T. Doudet1, F. Médale, P. Aguirre and Denise Blanc
Laboratoire de Nutrition des Poissons, INRA, BP 3, Station d'Hydrobiologie,
64310 Saint Pée-sur-Nivelle, France
1 present address: Sepia International, 16, Ave Gustave-Eiffel
78182 St. Quentin-Yvelines cedex, France
Triplicate groups of 40 nile tilapia (Oreochromis niloticus) each (IBW: 8 g) were fed one of six experimental diets containing practical ingredients over seven weeks at a water temperature of 28°C in a recirculated culture system. Another duplicate batch was fasted over the experimental period. The diets were formulated to be isoenergetic (DE: ca. 16 kJ/g DM) and to contain graded levels of digestible protein (0 to 350 g DP/kg DM) in order to vary the dietary DP/DE ratios. Apparent digestibility of the major nutrients (protein, energy, starch, phosphorus) and diets were determined using the indicator method. At the end of the growth trial, ammonia excretion and oxygen uptake rates were also measured.
Endogenous body nitrogen loss in fasted fish was higher than in those fed a protein-free diet (148 mg N/kg BW/d vs 73 mg N/kg BW/d). Similarly, endogenous (ammonia + urea) nitrogen excretion (ENE) was lower in protein-free diet fed fish (below 60 mg N/kg BW/d) than in fasted fish (190 mg N/kg BW/d). Analyses of data on growth and nitrogen retention showed that the maintenance protein requirements for nitrogen balance was 1.9 to 2.2 g/kg BW/d. Maximum growth rates and best protein and energy utilization were observed with tilapia fed diets containing 32 % crude protein. Although this value is apparently lower than some of the literature data, the optimal DP/DE ratio found in this study (18 mg DP/kJ DE) is comparable to values found by earlier works. The daily absolute protein requirement for maximum growth, nitrogen and energy retention was estimated to be 11 ± 1 g/kg BW/day.
Mobilization of body energy stores in fasted fish was 70 kJ/kg BW/d while a slight positive energy balance was achieved in tilapia fed the protein-free diet. The contribution of protein oxidation to energy expenditure in fasted fish represented above 2 mg N/kJ. Estimation of maintenance energy requirement based on oxygen uptake under fasting however showed values at least twice lower than those based on comparative carcass analyses. Oxygen uptake was linearly related to protein intake. Retained energy was maximal (40 % of DE) in tilapia fed diets having CP levels of 24 to 32 %. Protein energy retained per unit body weight increased with increasing protein intake. Fat energy gain represented a major part of energy increment and showed a tendency to decline with high protein intakes.
EIFAC SESSION 3/DIGESTION, UTILIZATION OF PROTEIN AND ENERGY/E/16
ENERGY AND NITROGEN BALANCE STUDIES AND EXCRETION PATTERNS IN RAINBOW TROUT UNDER DIFFERENT FEEDING REGIMES
J.H. van Weerd*, A.M. Verástegui and P.A.T. Tijssen
Department of Fish Culture and Fisheries, Wageningen Agricultural University,
P.O. Box 338, 6700 AH Wageningen, The Netherlands
Energy and nitrogen as well as excretion patterns of nitrogenous compounds were determined for rainbow trout (Oncorhynchus mykiss) of 110 g average weight, maintained for five weeks in the experimental facilities (respiration chambers with continuous flow analyzer) described earlier (Heinsbroek et al, in press). After a two-week adaptation period a three-week experimental period followed in which four feeding frequencies were employed, namely 1x, 2x, 4x and continuous feeding. A fasting group (no feeding) constituted the fifth treatment. During each week of the three week experimental period gaseous exchange (O2-consumption and CO2-production) and production of NH4- and NOx was continuously measured during two 24-hr runs.
Additional measurements on selected samples of in- and outflowing water were made in an attempt to elucidate the nature of the missing elements of the nitrogen budgets and to overcome difficulties in determination of RQ-values, which had been observed in earlier experiments (Heinsbroek et al, in press).
Heinsbroek, L.T.N., Tijssen, P.A.T., Flach, R.B. and De Jong, G.D.C., in press. In: Fish Nutrition in Practice. Proceedings IV International Symposium on Fish Nutrition and Feeding, Biarritz, France, June 24–27, 1991.
EIFAC SESSION 3/DIGESTION, UTILIZATION OF PROTEIN AND ENERGY/E/17
EFFECT OF DIFFERENT FEEDING INTENSITY ON RETENTION AND EFFICIENCY OF UTILIZATION OF ENERGY AND PROTEIN IN CARP (CYPRINUS CARPIO L.)
F.J. Schwarz* and M. Kirchgelβner
Institut für Ernährungsphysiologie, Technische Universität München-Weihenstephan,
8050 Freising, Germany
Feeding intensity was modified either by daily restricted feed amount or by limited daily protein or energy supply (restriction period) followed by an adequate supply (realimentation period) for carp (Cyprinus carpio L.) over a live weight range from 150 to 500 g. The mean daily energy retention of I up to 57 kJ per carp strictly depended on energy supply and live weight. The retained energy consisted of differing fat and protein gains, which was also reflected in the varying energy content of 6.4 to 13.0 kJ per g growth. The relative proportion of fat and protein had a strong influence on the utilization of the food energy. The total utilization of the gross energy and the partial utilization of the metabolizable energy for growth (kg) varied between 30 and 50 % and 0.50 and 0.70, respectively. The mean daily protein gain ranged from 0.3 to 1.1 g per carp and the protein content per g gain from 0.13 and 0.18 g depending on feeding (= energy) intensity and live weight. On the average there was a protein productive value (PPV) of 32 %.
EIFAC SESSION 3/DIGESTION, UTILIZATION OF PROTEIN AND ENERGY/E/18
IN VIVO AND IN VITRO DIGESTIBILITY OF DIETS AND FEEDSTUFFS FOR RAINBOW TROUT (ONCORHYNCHUS MYKISS)
E.F. Gomes1, A. Oliva Teles2, A. Gouveia2 and P. Rema3
1 Instituto Ciências Biomédicas, Universidade do Porto, Largo Prof. Abel Salazar 2,
P-4000, Porto, Portugal
2 Instituto de Zoologia, Fac. Ciencias, Universidade do Porto,
P-4000, Porto, Portugal
3 Universidade de Trás-os-Montese, Alto Douro, Dep. Zootécnia,
P-5001, Vila Real, Portugal
Knowledge of feedstuffs and diet digestiblity is a basic requirement for the elaboration of well balanced diets and for the evaluation of diet quality. Digestibility studies with fish have a lot of methodological problems, as fish live in water and use it as a vehicle for feed and for the elimination of faeces and end products of metabolism. As a consequence, a variety of different methods for faeces collection have been used, with variable results. Among them, the Choubert apparatus is one of the most sophisticated faeces collection methods and gives more reliable results. Besides the methodological difficulties of digestibility trials, they are time consuming, expensive and tedious. In vitro methods for the estimation of diet and feedstuff digestibility are frequently used with terrestrial land animals, however, their application to fish still needs confirmation.
The aim of this work was to compare the apparent protein digestibility coefficients of animal and plant protein feedstuffs and of compounded diets determined by an in vitro method with the values obtained in vivo with rainbow trout (Oncorhynchus mykiss) using the Choubert apparatus for faeces collection. In vitro protein digestibility was determined according to the technique described in AOAC (1980) using mucose porcine pepsin for digesting the samples.
Overall results showed that differences in the digestion coefficients obtained with the two methods were statistically significant. These differences were also significant in each group of feeds tested separately, namely, plant protein feeds, animal protein feeds, and compounded diets. The overall correlation coefficient between the in vivo and in vitro digestibility was also very low. The correlation coefficient between in vitro and in vivo digestibility of feedstuffs was considerably higher than that of the compounded diets, although it was not statistically significant.
It is concluded that the in vitro digestibility method used in this study was not a good criteria for the estimation of the apparent digestibility of protein for rainbow trout.
EIFAC SESSION 3/DIGESTION, UTILIZATION OF PROTEIN AND ENERGY/E/19
A CAUTIONARY NOTE FOR DIGESTIBILITY STUDIES BASED ON FAECAL SAMPLES OBTAINED BY STRIPPING THE FISH
Ruohonen K. and J. Vielma
Finnish Game and Fisheries Research Institute,
Evo State Fisheries and Aquaculture Research Station, 16970 Evo, Finland
A potential problem resulting from incorrect apparent digestibility coefficients (ADC) within studies using moist and semi-moist based feeds in Baltic herring was observed. Two year-old rainbow trouts (initially weighing about 450 grams) were raised in net cages with four different experimental diets having varying fish meal:minced herring ratios (isocaloric and isonitrogenous diets). Guar gum was used as a binder in the diets (3% of dry matter). In addition, groups of fish were also raised with a dry commercial feed and plain chopped herring. After seven weeks feeding with the experimental diets, the method of ADCs for protein, fat and phosphorus were determined using chromic oxide as a marker. For chopped herring, acid insoluble ash (AIA) was used as a marker. Faeces samples were collected by stripping the fish according to the method of Austreng (1978) under mild anaesthesia (tricaine 0.1 g/l).
The ADCs decreased with increasing herring content within the four experimental moist and semi-moist diets. The highest ADCs were observed with the commercial dry feed. The AIA based ADCs for chopped herring were higher than ADCs for moist diets and slightly lower than for commercial dry feed. The major differences between fish meal and minced Baltic herring are (1) the greater water content and (2) the lower fat content of herring. The experimental diets differed in water content, but were designed to be isonitrogenous and isocaloric. The fish ate approximately the same amount of dry matter despite of the diet's water content. The fish meal and minced herring differed also in their grading level, the fish meal being more finely graded than herring minced with a meat mincer.
The decreasing ADCs with the increasing dietary water content and fish meal:minced herring ratio may be a result of an interaction between water and guar gum, but more likely the results indicate that having coarsely graded raw material in the diet presumably leads to poor and inhomogenous mixing of the exogenous marker (as chromic oxide) within the diet. This may result in biased faeces samples when collecting them by one-time stripping. It is known that indigestible solids are emptied slower from the stomach and gut than the digestible part of the diet. Minced herring includes quite large particles (as pieces of bones and eye lenses) compared to the fine graded fish meal. Use of endogenous markers (as with chopped herring in this study) or preferably, collecting faeces over longer periods of time or on several occasions are probable ways to avoid the possible error.
Austreng, E., 1978: Digestibility determination in fish using chromic oxide marking and analysis of contents from different segments of the gastrointestinal tract. Aquaculture, 13: 265–272
EIFAC SESSION 3/DIGESTION, UTILIZATION OF PROTEIN AND ENERGY/E/20
THE USE OF N-ALKANES AS MARKERS FOR DETERMINATION OF INTAKE AND DIGESTIBILITY OF FISH FEED
Olafur Gudmundsson and Kristín Halldorsdottir
Agricultural Research Institute, Keldnaholt, IS-I 12 Reykjavik, Iceland
Measurements of intake and digestibility are among the most critical estimates needed to determine the nutrient requirements in fish. Many methods have been devised to study these parameters, as direct measurements are not applicable, but all have their shortcomings and disadvantages. Therefore indirect marker based approaches have become popular to determine digestibility, for example using chromium sesquioxide (Cr2O3), whereas accurate measurements of feed intake have been more difficult. Use of n-alkanes has acquired some popularity in measuring intake and digestibility in grazing animals. Therefore it has become interesting to study these compounds in fish as well. To evaluate the method, several feedstuffs commonly used in fish farming were analyzed for n-alkanes. Fish-and soybean meal only contain minimal amounts of n-alkanes (less than 1 mg/kg DM), whereas wheat and some other grains contain somewhat higher amounts of odd-chain n-alkanes, giving a limited increase in the quantity of these n-alkanes in compound feeds for fishes. Consequently digestibility and intake estimations were done in Arctic charr by using octacosane (C28) and/or dotriacontane (C32) as markers, either mixed in the feedstuff before pelleting or coated on the pellets. Celite and Cr2O3 were used for comparison in the digestibility study. During the intake study C32 was mixed into a small amount of the experimental feedstuff to be fed in limited amount at the first feeding of each day so as to know the exact consumption of C32, followed by feeding of the rest of the experimental feedstuff containing only C28 as a marker. The results indicate that in the 10 feedstuffs tested the n-alkanes tended on the average to give lower, but only in a few cases significantly (P<0.05) different digestibility, than celite and Cr2O3, for dry matter, protein, fat and energy, making them very prominent markers in digestibility studies. The n-alkanes also look promising as dosed markers for intake studies in fishes, provided that the chain length of the n-alkanes mixed in the total feed and the n-alkanes fed in known amount are of similar chain length and with the same faecal recoveries.
1 Supported in part by Icelandic Council of Science Grant No. 91NO76 and NATO Collaborative Research Grant No. 910484
EIFAC SESSION 3/DIGESTION, UTILIZATION OF PROTEIN AND ENERGY/E/21
METHODS TO ASSESS OPTIMAL NUTRIENT COMPOSITION OF DIETS FOR ATLANTIC SALMON
Barbara Grisdale-Helland* and Ståle J. Helland
AKVAFORSK, Institute of Aquaculture Research Ltd., 6600 Sunndalsora, Norway
To determine optimal diet compositions for Atlantic salmon, we use a combination of comparative slaughter analysis and metabolic measurements. The comparative slaughter technique is used to determine retention of nutrients such as nitrogen and energy from the diet in a group of fish. This requires knowledge of the total feed intake of each group, throughout the period of study. To allow for optimal growth in our studies, the fish are fed ad libitum using automatic feeders and disturbances from external sources are reduced to a minimum. Each tank (150 l) is equipped with a small light source directly above, preventing the fish from registering activity around the tank. Daily feed intake is calculated by measuring waste feed which is continuously sieved from the effluent water of each tank. While good feed stability is an important factor when using this method of waste feed collection, commercially produced diets have been used successfully with this technique. In a group of 12 tanks, water inflow, level and exit are automatically controlled giving 20 minute cycles of water recirculation every two hours. These semi-open, semi-closed respiration tanks are used for automatic measurements of oxygen consumption. A bleeder line can also be connected to an automatic analyzer for determinations of production rates of excreted metabolites.
In studies with Atlantic salmon (50–250 g) in fresh and saltwater, growth in these tanks has been similar to that expected in commercial units. Feed conversion rates as low as 0.6 kg dry feed per kg growth have been registered. Our results indicate large day to day variations in feed intake emphasizing the need for continuous measurements during the period of study. Nitrogen retention has been highest (66%) for smolt fed a diet containing 39% protein, 34% fat and 14% carbohydrate (18.5 MJ digestible energy/kg). Energy retention was 62% on this diet. In an experiment comparing diets containing from 41 to 63% protein and 30 to 16% fat, differences in oxygen consumption per kg fish were registered after only 3 weeks. Average oxygen consumption ranged from 3.9 to 5.1 g/kg body weight/day and was highest on the high protein diet. In conclusion, this system functions well as a model for optimalizing diet composition. This is emphasized by the low variation between replicates for the parameters measured.
EIFAC SESSION 3/DIGESTIBILITY, UTILIZATION OF PROTEIN AND ENERGY/E/22
CHANGES IN THE NUTRITIONAL QUALITY OF FISH MEAL BY HEAT TREATMENT
G. Andorsdottir1, T. Storebakken* and E. Austreng
AKVAFORSK, Institute of Aquaculture Research, Ltd., N-6600 Sunndalsora, Norway
1 Present Address: Faroe Seafood, P.O. Box 68, FR-110 Tórshavn, The Faroe Islands
During the 1980's, new low temperature dried qualities of fish meal were made available for fish feed production. In connection with this we wanted to evaluate the effects of heat in the production process. It is logistically difficult to obtain different qualities of meal, produced by full scale industrial processes from the same batch of fish. Thus, we compared a low temperature dried meal with an “artifically” over-dried meal (autoclaved after drying) from the same batch in four experiments:
Experiment 1. The two qualities of fish meal with and without supplementation of 1% DL
methionine in diets for Atlantic salmon fry and fingerlings.
Experiment 2. Digestibility of crude protein and individual amino acids from the same diets fed to
0.5 kg salmon.
Experiment 3. Effects of the diets on short-term feed intake (measured by a radioisotope method)
in 0.5 kg salmon.
Experiment 4. Biological protein evaluation of the fish feeds incorporated into standard diets to rat.
Autoclaving did not alter the chemical composition of the fish meal. The chemical score for the protein was 72 with histidine as the first limiting amino acid. Autoclaving the fish meal, however, resulted in reduced growth in Experiment 1. This was accompanied by an overall reduction in digestibility in Experiment 2. No effects of autoclaving the meal were seen on feed intake in Experiment 3. Digestibility of protein by rats in Experiment 4 was reduced by autoclaving the fish meal. The biological value (BV) was 81 for the low temperature meal and 72 for the autoclaved fish meal,
Methionine supplementation resulted in reduced growth in Experiment 1, but did not affect digestibility nor feed intake in Experiments 2 and 3. In Experiment 4, extra methionine resulted in increased BV of the protein.
In conclusion, autoclaving fish meal resulted in reductions in biological value, which were not detectable by chemical score.
EIFAC SESSION 3/DIGESTION, UTILIZATION OF PROTEIN AND ENERGY/E/23
ABSOLUTE FEEDING DESIGN, A REALISTIC WAY FOR FISH NUTRIENTS REQUIREMENTS DETERMINATION
Yann Moreau, Adou Cisse and Pierre Luquet
ORSTOM CRO, OI BP V 18,
Abidjan 01, Côte d'Ivoire
A feeding experiment is designed on the basis of food allowance rather than feed formulation. Food allowances are fixed in an attempt to provide the desired amount of nutrients per body mass of fish per day (in terms of g.kg-l. d-1 or J.Kg1.d-l). By this way, previous determination of optimum feeding ration is not required, and utilization of neutral feed complement (e.g. cellulose) is unnecessary. Experiment interpretation of this design is presented for a sample protein-energy requirement experiment for tilapia, (Sarotherodon melanotheron). Interrelationship of commonly used indices for growth and nutrition efficiency is discussed.
Table 1. Sample of absolute feeding formulation design. Feed compositions (in parts) and rations (% body weight) are calculated to provide 15 g protein per kg body weight per day (68%CP fish meal) associated to increasing amount of starch (cassava meal) from 0 to 40 g.kg-1.d-1.
| Feed name | AM0/15 | AM10/15 | AM20/15 | AM30/15 | AM40/15 |
| Fish meal | 21.95 | 21.95 | 21.95 | 21.95 | 21.95 |
| Cassava meal | 0.00 | 10.53 | 21.05 | 31.58 | 42.11 |
| Soyabean Oil | 0.90 | 0.90 | 0.90 | 0.90 | 0.90 |
| Vitamin & Mineral | 0.15 | 0.15 | 0.15 | 0.15 | 0.15 |
| Premix | |||||
| Digestibility marker | 0.48 | 0.71 | 0.93 | 1.15 | 1.37 |
| Binder | 0.73 | 1.06 | 1.39 | 1.72 | 2.06 |
| Total | 24.21 | 35.29 | 46.37 | 57.45 | 68.53 |
| Ration | 2.4% | 3.5% | 4.6% | 5.7% | 6.9% |
EIFAC SESSION 4/LIPIDS, ESSENTIAL FATTY ACID/R/3
REQUIREMENT CRITERIA FOR ESSENTIAL FATTY ACIDS
John R. Sargent
NERC Unit of Aquatic Biochemistry, University of Stirling, Stirling FK9 4LA, UK
All animals including fish require both (n-6) and (n-3) polyunsaturated fatty acids (PUFA) in their diets. However, in no species including fish are the dietary requirements for both (n-6) and (n-3) PUFA known with certainty. This reflects first the complexity and range of essential functions performed by PUFA in animals and second the interactions that exist not only between the (n-6) and (n-3) series of PUFA in metabolic functions in animals, but also between these fatty acids and their monounsaturated and saturated counterparts. In addition, essential fatty acid requirements can differ markedly at different development stages, a situation with particular relevance to fish where rapidly developing, free - feeding early larval stages may have high and exacting essential fatty acid requirements. Criteria for essential fatty acid requirements must be based on knowledge of first, the extent to which a species is capable of converting 18:2(n-6) to 20:4(n-6) and 22:4(n-6), and of 18:3(n-3) to 20:5(n-3) and 22:6(n-3). The extent to which these conversions occur for a given species is not related to environmental salinity, (i.e. whether the species is freshwater or marine), but it is related to the species' natural diet which can change markedly during the life cycle. Second, consideration must be given to the roles of (n-3) and (n-6) PUFA in maintaining cell membrane “structural integrity” in which context the different requirements for different tissues at different developmental stages are of paramount importance. Examples are the very high requirements of developing neural tissues for 22:6(n-3) in early life stages and the generally high requirements of endovascular tissue for 20:4(n-6). Third, consideration must be given to the importance of the continuing biosyntheses of eicosanoids from the essential fatty acids and the marked interactions that can exist between (n-6) and (n-3) PUFA in this area. Specific examples from each of these areas will be used to illustrate and validate criteria of essential fatty acid deficiency and imbalance in marine and freshwater fish encompassing both omnivores and extreme carnivores.
EIFAC SESSION 4/LIPIDS, ESSENTIAL FATTY ACID/E/24
STUDIES ON ESSENTIAL FATTY ACID REQUIREMENTS IN ATLANTIC SALMON
M. Thomassen
Institute of Aquaculture Research, P.O. Box 5010, N-1432 Aas, Norway
The results of experimental cage feeding trials conducted on the essential fatty acid requirements of Atlantic salmon are reported. The experiment was performed using purified experimental diets based on the use of casein as a protein source and coconut oil as a saturated lipid source; the latter was found to have a very poor palatability for Atlantic salmon. As with rainbow trout studies, growth in Atlantic salmon was positively correlated to (n-3) fatty acid content and thus indicated an essential dietary requirement for (n-3) fatty acids; better growth rates being observed with the highly unsaturated fatty acids (HUFA). However, at high dietary EFA inclusion levels a negative effect on growth was observed. Although EFA requirement was estimated to be about 1% (i.e. using growth as the requirement criteria), the requirement was estimated to be about 2% when histologial (i.e. pyloric caeca) and phospholipid (i.e. EFA, HUFA, 20:3 (n-9) samples are considered. The need for more studies concerning the EFA requirements of salmon was emphasized, and in particular concerning the optimum balance of (n-3) and (n-6) fatty acids.
(The above abstract is based on the oral presentation of Dr. Thomassen since no written abstract was provided prior to the workshop).
EIFAC SESSION 4/LIPIDS, ESSENTIAL FATTY ACID/E/25
EFFECT OF DIETARY LIPID ON GROWTH, FOOD CONVERSION AND BODY COMPOSITION OF INDIAN MAJOR CARP (LABEO ROHITA) FRY
M.R. Hasan*, A.M. Akand and M.A. Ali1
Department of Fisheries Biology and Limnology,
1 Department of Aquaculture
and Management, Bangladesh Agricultural University,
Mymensingh 2202, Bangladesh
A laboratory growth trial was conducted to determine the quantitative dietary lipid requirement of Indian major carp (Labeo rohita, Hamilton) fry (mean weight 1.56 ± 0.09 g). Seven purified diets containing graded levels (0, 3, 6, 9, 12, 15 and 18 %) of dietary lipid were used to determine the optimum dietary lipid and energy for the growth of major carp fry. Soybean and cod liver oil (2: 1) were used as the lipid source. The level of lipid in the diet was adjusted by the addition of a-cellulose. Casein was used as the source of dietary protein and dextrin and soluble starch (1.5: 1.0) were used as the source of carbohydrate. The levels of casein (35 %) and carbohydrate (25 %) were maintained the same for all diets. The experiment was conducted for 65 days in a static rearing system with three replications for each treatment. The effect of varying lipid level was measured on survivability, growth, food conversion, body composition and histological changes. Survivability of the experiment fish was high and ranged between 93.3 % to 100%. Analysis of variance showed that weight gain (%) and specific growth rate (SGR) were significantly (P<0.05) affected by the dietary lipid level. The weight gain (95.0 %) and SGR (1.03) were found to be highest in the fish fed the diet containing 12 % lipid and 3.66 kcal GE/g diet. The values for food conversion ratios (FCRs) were good in fish where the weight gain (%) and SGR were high. The FCRs values varied between 2.88 and 3.79. Carcass protein and lipid content increased with increasing level of dietary lipid up to 12 % and decreased thereafter. However, the carcass composition of fish fed the different levels of dietary lipid was within the normal health condition. Histological examination revealed intracellular lipid deposition in the liver of fish fed the diet containing 18 % dietary lipid. Based on the weight gain (%), SGR, FCR and histological changes, it may be concluded that the dietary lipid requirement of Indian major carp fry at an average water temperature of 27 ± 2°C was 12 % when soybean and cod liver oil were used as the sources of dietary lipid.
EIFAC SESSION 4/LIPIDS, ESSENTIAL FATTY ACID/E/26
DIETARY INFLUENCES ON PRODUCT QUALITY IN AFRICAN CATFISH (CLARIAS GARIEPINUS)
Helmut Wedekind
Forschungs-und Studienzentrum für Veredelungswirtschaft Weser-Ems, Driverstrassβe 22,
2848 Vechta (University of Gottingen), Germany
Present address:
Institut für Binnenfischerei, Potsdam-Sacrow, Jägerhof, 0-1501 Gross-Glienicke, Germany
In this trial a full-sib family of Clarias gariepinus was raised under standard conditions in a warmwater recirculation system. The fish were divided into two experimental groups of 1200 individuals each and were raised in separate tanks. The groups were fed two diets differing in crude protein (group I 40 % and group II 52 % d.m.) and crude fat (group I 8 % and group II 13 % d.m.). Gross energy was 21.3 and 22.3 MJ/kg d.m. Slaughtering of 100 fish per group (50 males, 50 females) took place after two experimental periods.
Besides the expected significant effect of the experimental diets on body weight and size a higher degree of corpulence was observed in group II. Tissue and organ percentages were not influenced, except a higher intestinal fat deposition in intensive fed fish. This group also showed a better net yield indicated by lower slaughtering-losses. Even more expressed differences between both feeding alternatives were observed in physical and chemical flesh quality criteria. Whereas in group I a tendency to a better texture of the flesh was found, the water binding capacity was significantly increased in group II. Moreover the appearance of the fillet was effected by the diet. Intensive fed catfish showed a visible higher degree of lightness with a tendency to a more yellowish colour. The chemical composition of the flesh indicated a significant increase of dry matter caused by higher protein and fat contents in this group. Further sensorial tests supported the measurements and showed marked differences in meat firmness between both groups with a better texture of group I. As an additional effect of the study a strong influence of sex and fattening age on inner and outer product quality was evident.
It can be seen that a high fat and protein content of the diet has a certain influence on product quality in African Catfish. It can be concluded that there is a clear effect of diet on quantitative criteria with a tendency to higher slaughtering yields in intensive fed catfish. This fact is connected with a positive effect on some physical flesh quality criteria which might be of importance for storage ability and processing operations. The chemical flesh composition does reflect the composition of the feeds but this effect, however, was rather small and not of practical importance.
EIFAC SESSION 4/LIPIDS, ESSENTIAL FATTY ACID/E/27
MEASUREMENT OF DIGESTIBILITIES OF CHOLESTEROL AND FATTY ACIDS USING AN INDICATOR 5α-CHOLESTANE
Manabu Ishikawa, Shin-ichi Teshima*, Akio Kanazawa and Shunsuke Koshio
Faculty of Fisheries, Kagoshima University, Shimoarata, Kagoshima 890, Japan
For the measurement of nutrient digestibilities in fish, an indirect method using chromic oxide (Cr2O3-method) as an indicator has been widely used by many workers. However, this method requires relatively large amounts of faecal samples and requires considerable time for the collection of faeces. In addition, it is difficult to recover aliquots of samples for the purpose of other analysis, in addition to the trouble of the treatment of chromium waste so as to avoid water pollution. In the present study, therefore, an attempt to measure the digestibilities of cholesterol and fatty acids using 5α-cholestane as an indicator (cholestane-method) was conducted by using the tilapia, Oreochromis niloticus and fresh-water prawn, Macrobrachium rosenbergii, as a test animal. The results were compared with those obtained by the Cr2O3-method.
A casein-based test diet containing 0.3 % levels of cholestane and chromic oxide, as the indicators, were prepared for determination of digestibilities of dietary cholesterol and fatty acids. The test animals, tilapia (3.0 g body weight) and freshwater prawn (0.5 g body weight) were pre-cultivated with the test diet for two weeks, and then faecal samples collected for six successive days. Lipids in the diets and faeces were extracted with chloroform-methanol and saponified with 10 % KOH in methanol. Gas-liquid chromatography (GLC) on 1.5 % OV-17 of the unsaponifiable matters afforded the quantities of 5α-cholestane and cholesterol, whereas that on 5 % Shinchrom E-71 of the saponifiable matters gave the quantities of fatty acids. Chromic oxide was determined by a wet digestion method.
In tilapia the absorption rates (%) of cholesterol as determined by the cholestane- and Cr2O3-methods were 72–75 % and 76–79 %, respectively. The digestibilities of individual fatty acids determined by the cholestane-method ranged from 65.2 to 96.4 %, whereas those determined by the Cr2O3-method varied from 69.9 to 96.9 %. Thus, the digestibilities of dietary fatty acids differed with the kinds of fatty acids. The digestibilities determined by the two methods were similar to each other, except for a slightly lower value for monoene-fatty acids using the cholestane-method compared with the Cr2O3-method. This indicates that the cholestane-method as well as the Cr2O3-method can be conveniently used for the measurement of digestibilities (or absorption) of fatty acids and cholesterol. The cholestane-method has the advantage that it is possible to determine cholesterol (or fatty acids) and 5α-cholestane simultaneously by GLC using small amounts of faecal samples (about 10 mg). As for the freshwater prawn, the cholestane-method was found to be useful for the determination of fatty acid digestibility. However, it is not clear whether this method is applicable for the measurement of absorption rate of dietary cholesterol.
EIFAC SESSION 5/CARBOHYDRATES/R/4
CARBOHYDRATE UTILIZATION AND ITS DETERMINATION
E. Pfeffer
Institut für Tierernährung der Universität Bonn,
Endenicher Allee 15, D-53115 Bonn, Germany
Carbohydrates which supply most of the digestible energy (DE) to terrestrial farm animals have long been thought to be of no nutritional value to salmonid fish. It has, however, been found that starch is not always indigestible and that absorbed glucose may contribute to meeting metabolic energy demands of these fish.
Within the range of practical relevance, digestion is the limiting step for the utilization of carbohydrates. Therefore, measuring digestibility is the adequate way of determining the degree to which carbohydrates are utilized by fish.
Alpha-linked glucose polymers and free glucose appear to be the only carbohydrates digestible to rainbow trout. Differentiation between starch and non-starch carbohydrates may, therefore, be a valuable nutritional information. Digestibility of starch may be increased substantially by hydrothermical treatments, such as extrusion. The extent of this influence may be monitored by measuring the amyloglucosidase degradability of starch, whereas the pancreatin test is not suited for such a differentiation.
Independent of the degree of gelatinization, however, passage rate seems to affect digestibility of starch substantially. Therefore, varying feeding intensity has been found to result in changes in starch digestibility of up to 23 %-units. When trout are fed to satiation, the concentration of starch of identical origin in the diet is negatively correlated with its digestibility.
These factors limit the utilization of carbohydrates and it appears unlikely that under conditions relevant to production that carbohydrates can contribute more than about 20 % to the total supply of DE to salmonids. Non-starch carbohydrates limit the use of potential plant sources of dietary protein such as legume seeds.
![]()
![]()
![]()