![]()
![]()
by
B.E. Marshall
Department of Biological Sciences
University of Zimbabwe
P.O. Box MP167, Mount Pleasant
Harare, Zimbabwe
ABSTRACTSmall body size is a feature of Limnothrissa miodon in the man-made Lakes Kariba and Cahora Bassa. Recent reliable determinations of their growth confirm the earlier very high estimates of their mortality. These high mortality rates are consistent with the small body size of the fish. A similar allometric relationship applies to the production/biomass ratios. Because most fish die after a few months there is no relationship between fishing effort and catch. Regulating effort may therefore contribute little to the management of this stock. The greatest risk to the fishery is a collapse of recruitment and there is an urgent need to understand the factors that regulate reproduction. RESUMELa petite taille est une caractéristique de Limnothrissa miodon dans les lacs artificiels de Kariba et de Cahora Bassa. Des mesures fiables récentes de leur croissance confirment de précédentes estimations selon lesquelles leur mortalité était très élevée. Cette mortalité élevée concorde avec la petite dimension du poisson. Une relation allométrique analogue s'applique aux ratios production/biomasse. Etant donné que la plupart des poissons meurent au bout de quelques mois, il n'y a aucune relation entre l'effort de pêche et les captures. La réglementation de l'effort ne peut donc guère contribuer à l'aménagement de ce stock. Le risque le plus grand pour la pêche réside dans un effondrement du recrutement et il est urgent de comprendre les facteurs qui régissent la reproduction. |
The great man-made lakes on the Zambezi River present many dramatic spectacles, not the least of which is their population of pelagic fish. In Lake Kariba, for example, this consists almost entirely of the introduced sardine Limnothrissa miodon which, with an average total length of 50–55mm, is probably smaller than any other important commercially exploited fish species. Although aspects of its ecology are now relatively well-known (e.g. Begg, 1974, 1976; Cochrane, 1978, 1984; Marshall, 1982, 1985, 1987a, 1987b, 1988a, 1988b, 1991, 1992; Mtada, 1987) we still do not fully understand how a tiny fish, living in an apparently oligotrophic and relatively variable lake, exposed to heavy predation and subjected to an immense and steadily increasing fishing effort has been able to sustain a major fishery.
Because of its small size it would not be surprising to find that its population dynamics differ considerably from those of larger species. Studies on the effect of scaling in animals (Peters, 1983) have shown that the rates of most physiological processes are determined by body size. This paper considers the proposition that the demographic parameters of Limnothrissa are predictable functions of the allometric relationship
Y = aWb
where Y is the biological characteristic and W is body weight (Peters, 1983). The implications for management and future research are briefly discussed. Only the Lake Kariba population is considered because there are insufficient data from Lake Cahora Bassa, but the discussion applies equally to the sardines in both lakes because of the similarity in the body size of the fish (Gliwicz, 1984).
The sardines in Lakes Kariba and Cahora Bassa are, on average, about half the length and sometimes only one-tenth of the weight of those in Lakes Tanganyika or Kivu. Most importantly, their reproductive characteristics have changed as well. They reach sexual maturity at 35 mm in length (about 3 months old) in Lake Kariba compared to 70–75 mm (about 9 months old) in Lakes Tanganyika and Kivu (Begg, 1974; Ellis, 1971; Spliethoff et al., 1983). This stunting is caused by a reduction in their average life-span from two years to one year (Marshall, 1993). Intensive commercial fishing can bring about a reduction in the body size of fish but it is unlikely to have done so in this case. There is no fishing in Lake Cahora Bassa and they have always been small in Lake Kariba. For example, in 1971, only 3–4 years after their introduction and before any fishing had begun, their average length was 55∙7 mm (Balon, 1974), much the same as it was 10 years later (Marshall, 1988a).
The length-frequency distribution of sardines caught at 10 stations in the eastern basin of Lake Kariba in 1981–1983 gives a good impression of the nature of the population (Fig. 1). The apparent lack of small fish is an artifact; most small Limnothrissa live in shallow water (Cochrane, 1984) and the sampling programme included only one station less than 5m deep. On the other hand, the rapid decline in the numbers of fish over 55mm long is a true reflection of the population. Some large fish do occur in the lake, of course, but their numbers are low (only 0.01% of the sample was over 70mm in length). The absence of large fish from the sample is almost certainly a consequence of their high mortality since there is no evidence that they are not selected by the fishing gear or that they migrate to areas where they cannot be caught.
The growth of Limnothrissa has been determined three times in Lake Kariba. Cochrane (1984) and Marshall (1987a) used length-based methods which may be unreliable because overlapping cohorts make it difficult to separate the significant peaks whilst the lack of large fish means that older age groups are poorly represented. (These problems apply equally to computer-assisted interpretations and the results from programmes like ELEFAN should not be accepted uncritically). Chifamba (1992) used daily growth rings on the otoliths and was able to overcome the problem of under-representation by selecting larger fish from commercial catches. An estimate based on growth checks should be more accurate, even though there was considerable variation in the length of fish in relation to the number of daily rings.
The growth parameters vary considerably (Table 1) but the resultant growth curves are very similar, at least up to the age of 12 months (Fig.2). The differences between them are of little significance, given the difficulties of determining age, the range of individual variability and the year-to-year variations to be expected in a short-lived species that responds rapidly to environmental changes (Marshall, 1988c). Future research might improve the precision of the estimates but it is unlikely to change the general form of the growth curve. We can assume that the growth of Limnothrissa in Lake Kariba is now sufficiently well-known to allow a re-examination of its mortality rates.
The published instantaneous rates of total mortality (Z) for Limnothrissa in Lake Kariba range from 9∙6–13∙7 year-1 (Marshall, 1987a). Recent workers on Lake Kariba have suggested that these estimates are too high (Anonymous, 1992; Pearce, undated). This is important because estimates of mortality are necessary components of stock assessment and we have to be satisfied that they are accurate.
In order to assess the effect of growth determinations on mortality the samples collected in 1981–1983 (Fig. 1) were aged by means of the three growth curves shown in Fig. 2. The samples were then split into age groups and their total mortality rates were estimated by the construction of catch curves (Fig. 3). The lowest mortality rate was obtained with Cochrane's curve (Z = 6∙96 year-1) whilst the highest rate (Z = 17∙04), which exceeded any of the published estimates, resulted from Chifamba's curve. If the published mortality estimates (Marshall, 1987a) which were derived from a length-based growth curve are, in fact, too high then they should have been lowered by the application of a more accurate growth estimation. This was not the case, suggesting that the existing mortality estimates are not unrealistically high and can be used with confidence.
The mortality rates of Limnothrissa can be put into context by comparing them with those of the other fish in the lake. There was a close allometric relationship between mortality and size in these fish (Fig. 4) and the values for Limnothrissa lie within the 95% confidence band. They are, therefore, within the range that might be expected in a fish of its size in Lake Kariba. They lie close to the upper confidence limit, which is also to be expected because mortality rates tend to be higher in small, shoaling pelagic species. Most of the other fish species in Lake Kariba, with the exception of Hydrocynus, are bottom-dwellers. It should also be noted that some values are probably too low because of inaccurate ageing of the cichlid fish (Hustler and Marshall, 1990).
There is a similar relationship in a much wider range of both marine and freshwater tropical fish (Fig. 5). This should be treated with some caution, however, because total mortality rates can be increased considerably through fishing. On the other hand, this problem may be partly offset by a reduction in the average size of individuals in exploited fish stocks, which is a frequent consequence of intensive fishing. Despite these reservations, the similarity of the exponent in Figs. 4 and 5 is noteworthy and suggests a degree of universality in the processes that govern mortality in fish.
No population can maintain itself in the face of such high mortality without a high turnover, or production/biomass, ratio which is a difficult quantity to measure. The instantaneous growth coefficient (G) is approximately equal to it (Waters, 1977) but, when mortality is exponential and growth conforms to the von Bertalanffy equation (as in Limnothrissa), the mortality rate (Z) will also closely approximate the P/B ratio (Allen, 1971). Thus it is possible to obtain a range of P/B values for Limnothrissa in Lake Kariba (Table 2). There is an allometric relationship between P/B and size in the fish community in Lake Kariba (Fig. 6). The exponent of -0∙25 is similar to the one obtained by Banse and Mosher (1980). The same caveat applied to mortality is required because inaccuracies in the ageing of cichlid fish means that their P/B ratios may have been considerably underestimated (Hustler and Marshall, 1990).
The P/B ratio can also be examined in the context of the sardine's longevity which is itself related to size (Blueweiss et al., 1978). Lévêque et al. (1977) have shown that the two variables are closely related in a wide range of animals (Fig. 7). The values for Limnothrissa, using either G or Z, fit this relationship closely, thus providing further confirmation that the sardine's population parameters are a consequence of its small size.
The high P/B ratios of the sardines in Lake Kariba contribute to the high yields of the fishery. The average biomass is around 25–30 000 t (Marshall, 1988a) and so a P/B ratio of 5∙0 would result in an annual production of 125–150 000 t. Since the P/B ratio is probably higher than this the annual production could amount to 250 000 t or more per annum. Not all of this production can be taken up by the fishery but, nevertheless, a high proportion could be caught without risk to the stock.
In Lake Kariba Limnothrissa is essentially an annual species. Marshall (1987a) attributed increases in the mortality rate to increased fishing effort but the changes may equally have been due to environmental changes (Marshall, 1988c). It is more likely that fishing mortality substitutes for rather than adds to mortality from other sources. Those individuals that are not caught by the fishermen will soon die in any case. Under such circumstances conventional fisheries models are of little practical value in the management of the fishery and attempts to use them (e.g. Marshall, 1985) have been largely unsuccessful.
This applies especially to the relationship between fishing effort and catch per unit effort (CPUE). Apart from rapid changes that occurred in the early years of the fishery, following limnological changes in the lake after the decline of its Salvinia mats (Marshall, 1992), the massive increase in fishing effort has had no effect on the CPUE (Fig. 8). The CPUE is not a good measure of the abundance of small shoaling fish (Csirke, 1988) and some of the problems of using it may apply to the Lake Kariba fishery. Instead CPUE is most likely to be a reflection of the intrinsic catching power of the fishing gear, influenced to a small extent by changes in the abundance of the fish from year to year.
What happens to adult sardines is therefore of little significance because most of them die anyway in a short time. This applies irrespective of whether death is caused by fishing. The current focus on fishing effort is therefore largely irrelevant to management of the fishery. In spite of this, regulation of effort is, on the Zimbabwean side at least, a key management principle (Machena and Mabaye, 1987). Much of the current research is apparently intended to refine the estimation of effort and provide data for its further control.
The major threat to this fishery, as it is in other clupeid fisheries, is reproductive failure leading to a decrease in recruitment. Except for some preliminary work by Cochrane (1978) little is known about the breeding biology and early life history of Limnothrissa in Lake Kariba. Key questions relate to the environmental factors that determine reproductive success, to the effect of fishing on the sexually mature population and stock-recruitment relationships.
Many of these questions are difficult to answer but they are of vital importance if the fishery is to be managed successfully. This work could provide an opportunity to make a significant contribution to our understanding of fundamental biological processes in the tropics.
I am grateful to my colleague Peter Frost whose understanding of allometric relationships assisted me greatly in the preparation of this paper. My attendance at this symposium was made possible through sponsorship from the Technical Centre for Agriculture and Rural Cooperation (CTA) arranged by the staff of the FAO-IFIP project in Bujumbura.
Allen, K.R., 1971. The relation between production and biomass. J. Fish. Res. Bd. Can., 28: 1573–81.
Anonymous, 1992. Working group on assessment of kapenta (Limnothrissa miodon) stocks in Lake Kariba. SADCC Zambia/Zimbabwe Fish. Proj., Kariba 4–17 March 1992, Lake Kariba Fish. Res. Inst., 75pp.
Balon, E.K., 1974. Fish production of a tropical ecosystem. In E.K. Balon and A.G. Coche (eds) Lake Kariba; a Man-made Tropical Ecosystem in Central Africa p. 249–573. The Hague; Dr W Junk.
Banse, K. and S. Mosher, 1980. Adult body mass and annual production/biomass relationships of field populations. Ecol. Monogr., 50: 355–79.
Begg, G.W., 1974. Investigations into the biology and status of the Tanganyika sardine, Limnothrissa miodon (Boulenger), in Lake Kariba, Rhodesia. Lake Kariba. Fish. Res. Inst., Proj. Rept 17, 151pp.
Begg, G.W., 1976. The relationship between the diurnal movements of some of the zooplankton and the sardine Limnothrissa miodon in Lake Kariba, Rhodesia. Limnol. Oceanogr., 21: 529–39.
Blueweiss, L.H., H. Fox, V. Kudzma, D. Nakashima, R. Peters and S. Sams, 1978. Relationships between body size and some life history parameters. Oecologia, 37: 257–72.
Chapman, D.W. and P. van Well, 1978. Growth and mortality of Stolothrissa tanganicae. Trans. Am. Fish. Soc., 107: 26–35.
Chifamba, P.C., 1992. Daily rings on otoliths as a method for ageing the sardine Limnothrissa miodon in Lake Kariba. Trans. Zimbabwe Sci. Ass., 66 (in press).
Cochrane, K.L., 1978. Seasonal fluctuations in the catches of Limnothrissa miodon (Boulenger) in Lake Kariba. Lake Kariba Fish. Res. Inst., Proj. Rept 29, 163pp.
Cochrane, K.L., 1984. The influence of food availability, breeding seasons and growth on commercial catches of Limnothrissa miodon (Boulenger) in Lake Kariba. J. Fish Biol. 24: 623–35.
Csirke, J., 1988. Small shoaling pelagic fish stocks. In J.A. Gulland (ed.) Fish Population Dynamics, 2nd edn., pp. 271–302. New York; John Wiley.
Edwards, R.R.C. and S. Shaher, 1991. The biometrics of marine fishes from the Gulf of Aden. Fishbyte, 9: 27–9.
Ellis, C.M.A., 1971. The size at maturity and breeding seasons of sardines in southern Lake Tanganyika. Afr. J. Trop. Hydrobiol. Fish., 1: 59–66.
Gliwicz, Z.M., 1984. Limnological study of Cahora Bassa reservoir with special regard to sardine fishery expansion. FAO/GCP/MOZ/006/SWE Field Doc. 8, 71pp. Rome; FAO.
Garrod, D.J., 1959. The growth of Tilapia esculenta in Lake Victoria. Hydrobiologia, 12: 1959 268–98.
Garrod, D.J., 1963. An estimation of the mortality rates in a population of Tilapia esculenta Graham (Pisces; Cichlidae). J. Fish. Res. Bd. Can., 20: 195–227.
Hustler, K. and B.E. Marshall, 1990. Population dynamics of two small cichlid fish species in a ropical man-made lake (Lake Kariba). Hydrobiologia, 190, 253–62.
Lévêque, C., J.-R. Durand and J.-M. Écoutin, 1977. Relations entre le rapport P/B et la longévité des organismes. Cah. ORSTOM, sér. Hydrobiol., 11: 17–31.
Machena, C. and A.B.E. Mabaye, 1987. Some management aspects and constraints in the Lake Kariba fishery. NAGA, the ICLARM Quarterly, 10(4): 10–2.
Marshall, B.E., 1982. The influence of river flow on pelagic sardine catches in Lake Kariba. J. Fish. Biol., 20: 465–69.
Marshall, B.E., 1985. A Study of the Population Dynamics, Production and Potential Yield of the Sardine Limnothrissa miodon (Boulenger) in Lake Kariba. Ph.D. thesis, Rhodes University, Grahamstown.
Marshall, B.E., 1987a. Growth and mortality of the introduced Lake Tanganyika clupeid, Limnothrissa miodon, in Lake Kariba. J. Fish Biol., 31: 603–15.
Marshall, B.E., 1987b. On the relationship between tigerfish and sardines in Lake Kariba. J. Limnol. Soc. sth. Afr., 13: 78–9.
Marshall, B.E., 1988a. Seasonal and annual variations in the abundance of pelagic sardines in Lake Kariba, with special reference to the effects of drought. Arch. Hydrobiol., 112: 399–409.
Marshall, B.E., 1988b. A preliminary assessment of the biomass of the pelagic sardine, Limnothrissa miodon, in Lake Kariba. J. Fish Biol., 32: 515–24.
Marshall, B.E., 1988c. Why are the sardines in Kariba so small? Zimbabwe Sci. News, 22: 31–5.
Marshall, B.E., 1991. The impact of the introduced sardine Limnothrissa miodon on the ecology of Lake Kariba. Biol. Conserv., 55: 151–65.
Marshall, B.E., 1992. The relationship between catch and effort and its effect on predicted yields of the Lake Kariba sardine fishery. NAGA, the ICLARM Quarterly (in press).
Marshall, B.E., 1993. The biology of the African clupeid Limnothrissa miodon with special reference to its small size in artificial lakes. Rev. Fish Biol. Fish., 3 (in press).
Moreau, J., J. Munyandorero and B. Nyakageni, 1991. Evaluation des paramètres démographiques chez Stolothrissa tanganicae et Limnothrissa miodon du lac Tanganyika. Verh. Internat. Verein. Limnol., 24: 2552–8.
Mtada, O.S.M., 1987. The influence of thermal stratification on pelagic fish yields in Lake Kariba, Zimbabwe. J. Fish Biol., 30: 127–33.
Pauly, D., 1984. Studying single-species dynamics in a multispecies system. In D. Pauly & G.I. Murphy (eds) Theory and Management of Tropical Fisheries, p. 33–70. Manila; ICLARM.
Pauly, D. and D. Martosubroto, P., 1980. The population dynamics of Nemipterus marginatus (Cuvier & Val.) off western Kalimantan, South China Sea. J. Fish Biol., 17: 263–73.
Pauly, D. and J.L. Munro, 1984. Once more on the comparison of growth in fish and invertebrates. Fishbyte, 2(1):21. Pearce, M.J., A review of the stock assessment of kapenta in Lake Kariba. Lake undated Kariba Fish. Res. Inst., unpublished report, 12pp.
Peters, R.H., 1983. The Ecological Implications of Body Size. Cambridge University Press.
Spliethoff, P.C., H.H. de longh and V. Frank, 1983. Success of the introduction of the freshwater clupeid Limnothrissa miodon (Boulenger) in Lake Kivu. Fish. Mgmt., 14: 17–31.
Venema, C. and N.P. van Zalinge (eds), 1989. Contributions to tropical fish stock assessment in India. Papers presented by the participants at the FAO/DANIDA/ICAR National Follow-up Training Course in Fish Stock Assessment, Cochin, India, 2–28 November 1987. FI:GCP/INT/392/DEN/1, 157pp. Rome;FAO.
Venema, S.C., J.M. Christensen and D. Pauly, D. (eds), 1988. Contributions to tropical fish biology. Papers prepared by the participants at the FAO/DANIDA follow-up Training Courses in Fish Stock Assessment in the Tropics, Hirtshals, Denmark, 5–30 May 1986 and Manila, Philippines, 12 January – 6 February 1987. FAO Fish. Rept 389, 519pp.
Waters, T.F., 1977. Secondary production in inland waters. Adv. Ecol. Res., 10: 91–164.
Table 1. Growth parameters of Limnothrissa miodon in Lake Kariba. L∞ and K (annual rate) are components of the von Bertalanffy growth equation whilst Φ' is an index of growth performance = logK + 2logL∞ (Pauly and Munro, 1984).
| Source | L∞ (cm) | K | Φ' |
| Cochrane (1984) | 8.10 | 1.74 | 2.06 |
| Marshall (1987a) | 7.42 | 3.05 | 2.23 |
| Chifamba (1992) | 13.58 | 0.95 | 2.24 |
Table 2. Estimates of P/B based on the instantaneous growth coefficient G) and total mortality rate (Z) of Limnothrissa miodon in Lake Kariba. Values of G are derived from published growth curves and length-weight relationships. Both G and Z are expressed as annual rates.
| Source | G | Z |
| Cochrane (1984) | 4.13 | |
| Marshall (1987a) | 4.20 | 9.6 – 13.7 |
| Chifamba (1992) | 5.61 |
There are relatively few fisheries that depend on very small species of fish (i.e. <10 cm in length) and so their growth parameters are seldom reported. Tuna baitfish are small clupeids and engraulids which have population parameters very similar to those of Limnothrissa miodon in Lake Kariba. The following data could not be included in the paper since length-weight relationships were not given and estimates of asymptotic weight could not be made (K and Z are annual rates). PNG = Papua New Guinea, SI = Solomon Islands.
| Species | L∞ (cm) | K | Z | Source |
| Spratelloides lewisi (PNG) | 6.3 | 5.4 | 14.2 | 1 |
| Spratelloides delicatulus (SI) | 6.8 | 4.5 | 12.5 | 2 |
| Stolephorus devisi (PNG) | 7.5 | 2.2 | 8.6 | 1 |
| S. devisi (SI) | 7.2 | 2.1 | 5.8 | 2 |
| Spratelloides gracilis (PNG) | 7.6 | 4.3 | 14.4 | 1 |
| Stolephorus heterolobus (PNG) | 8.3 | 2.5 | 9.8 | 1 |
| S. heterolobus (SI) | 8.1 | 2.5 | 5.4 | 2 |
| Stolephorus waitei (PNG) | 10.9 | 1.7 | 3.8 | 1 |
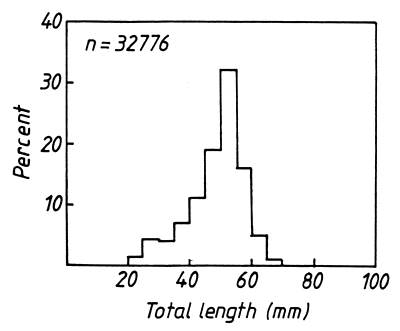
Figure 1. The length-frequency distribution of Limnothrissa miodon collected at 10 sampling stations in the Sanyati Basin of Lake Kariba, 1981–1983 (see Marshall, 1985, 1987a, 1988a).
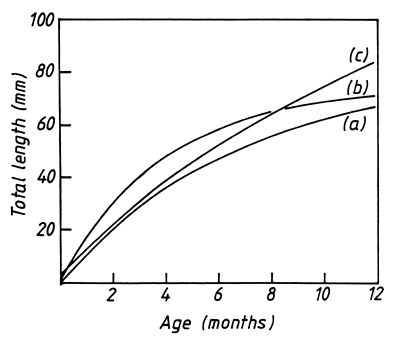
Figure 2. The growth of Limnothrissa miodon in Lake Kariba according to (a) Cochrane (1984), (b) Marshall (1987a) and (c) Chifamba (1992). Data for (a) and (b) were obtained from length-frequency analysis whilst data for (c) came from daily rings on otoliths; all three curves were fitted by the von Bertalanffy equation.
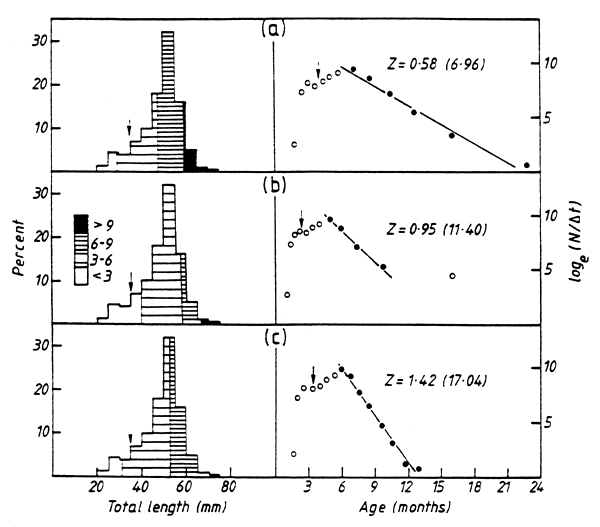
Figure 3. The age distribution (left) and resultant catch curves (right) of the sample of Limnothrissa miodon shown in Fig. 1 determined by using the growth curves of (a) Cochrane (1984), (b) Marshall (1987a) and (c) Chifamba (1992). The small arrow in each figure denotes the length (35 mm) and the estimated age of the smallest sexually mature fish (data from Begg, 1974). The catch curves were determined according to the methods in Pauly (1982); the solid circles denote the points that were used in calculating the total mortality rate (Z). Values of Z are monthly rates with the annual rates in brackets.
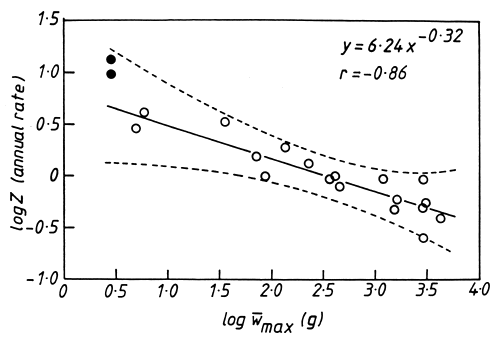
Figure 4. The relationship between the total mortality rate and the mean maximum weight, i.e. the mean weight of the oldest size class, of fish species in Lake Kariba, based on data in Balon (1974) and Mahon and Balon (1977). The broken lines show the 95% prediction limits while the solid circles represent the highest and lowest values for Limnothrissa miodon (Marshall, 1987a). They are not included in the regression.
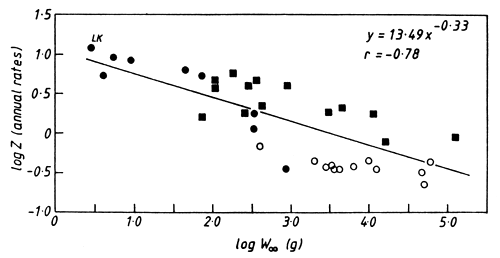
Figure 5. The relationship between the total mortality rate and asymptotic weight (W∞) in tropical fishes. ■ = data from papers in Venema and van Zalinge (1988) and Venema et al. (1989); ○ = data from Edwards and Shaher (1991); • = data from other sources (Garrod (1959, 1963), Chapman & van Well (1978), Pauly & Martosubroto (1980), Marshall (1987a), Hustler and Marshall (1990), Moreau et al. (1991). LK indicates the mean value for Limnothrissa in Lake Kariba; the point is included in the regression.
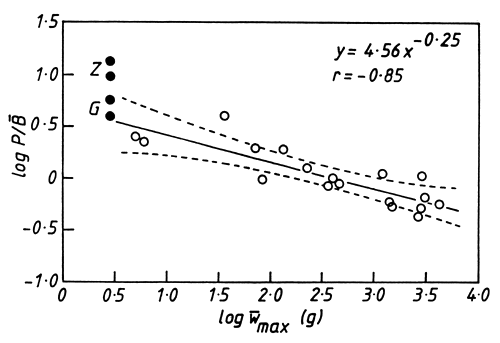
Figure 6. The relationship between the P/B ratio and the mean maximum weight of fish species in Lake Kariba, based on data in Balon (1974) and Mahon and Balon (1977). The broken lines show the 95% prediction limits while the solid circles represent the highest and lowest values for Limnothrissa miodon, based on its total mortality rate (Z) and growth coefficient (G). They are not included in the regression.
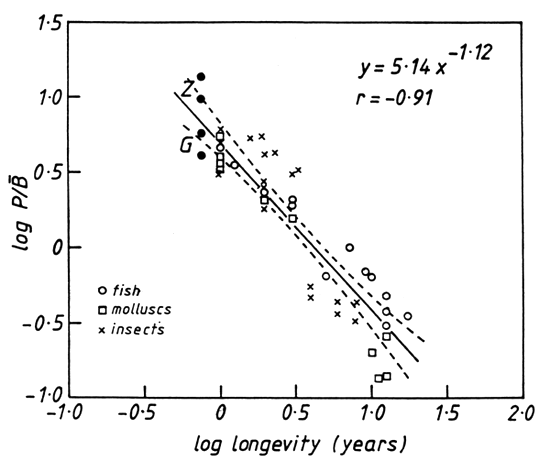
Figure 7. The relationship between the P/B ratio and longevity in a variety of animal species (data from Lévêque et al., 1977). The solid circles represent the highest and lowest values for Limnothrissa miodon based on their total mortality rates (Z) and growth coefficients (G). They are not included in the regression.
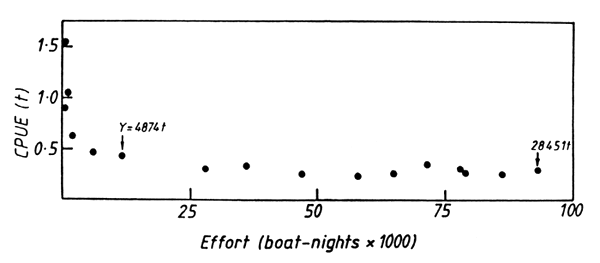
Figure 8. The relationship between catch per unit effort (t boat-night-1) and fishing effort (boat-nights × 1000) in the Lake Kariba sardine fishery, 1974–1989. Data are for both Zimbabwean and Zambian fisheries, redrawn from Marshall (1992). The total yields in 1979 and 1989 are included to illustrate the increase in catch that took place over this period.
![]()
![]()