![]()
![]()
![]()
All the large Latin American catchment basins contain fish species which migrate over fairly long distances for breeding or feeding purposes (Welcomme, 1985; Petrere, 1985). These movements are longitudinal, through the main channel and tributaries. In most cases, fish migrate upstream in order to spawn. Sometimes, upstream migrations occur on a massive scale, especially in upper reaches. In others, e.g. the lower Plata basin of the Paraná river, only a few individuals undertake long-distance upstream migrations. Most of them stay in the tagging area or undertake lateral migrations to the floodplains or the river's branch channels (Bonetto, Canon Verón and Roldán, 1981). Migrations may involve distances of up to 1 500 km (Petrere, 1985) and usually exceed 200 km (Welcomme, 1985). Mean speeds, between tagging and recapture, may exceed 20 km/day. Speeds vary depending on the species, the stage of the river's hydrological cycle and the reach in which the fish are tagged.
Poddubnyi, Espinach Ros and Oldani (1981) determined rates of movement and daily activity in the Paraná river during reproductive migrations. The fish were tagged with ultrasound capsules and monitored for several days. The mean river flow during that time was estimated at 0.8 to 1.0 m/sec. Of the five tagged Prochilodus platensis individuals which moved through the arms of the floodplain, one drifted downstream and another remained in the restricted main channel area for two days, moving short distances. The fifth moved upstream following the gradient of the slower flowing left bank and often frequented the littoral zone resting close to clumps of submersed trees on the slopes of bottom elevations. It travelled only during the day, showing three peaks of activity (Figure 3). Maximum speed against the current was 1.0 km/h, maintained for one and a half hours. The individuals moving through the branch channels and lagoons of the floodplain travelled at a speed of 0.6 km/h. In the branch channels, they moved along the edge of the aquatic plant zone and often stopped to penetrate the zone. In the lagoons, they stayed in areas of dense vegetation most of the time. In similar experiments conducted at Salto Grande, a river-type reservoir, the fish were active both day and night and the maximum speed recorded was 0.45 km/h. Their course was a very sinuous one which followed the slope of the former channel and took them close to the bank (Poddubnyi, Espinach Ros and Oldani, 1981).
An individual of the species Salminus maxillosus, monitored for nine days, moved through a very small area of the river. It showed three peaks of diurnal activity and did not travel at night. Maximum speed against the current was 2 km/h. In the Salto Grande reservoir, Salminus travelled upstream at a maximum speed of 1.3 km/h. The maximum speeds at which Pseudoplatystoma coruscans and Luciopimelodus pati travelled against the current were 2.6 and 1.0 km/h respectively. When moving against the current, they followed the contours of the bottom and travelled in the evening and at night (Poddubnyi, Espinach Ros and Oldani, 1981).
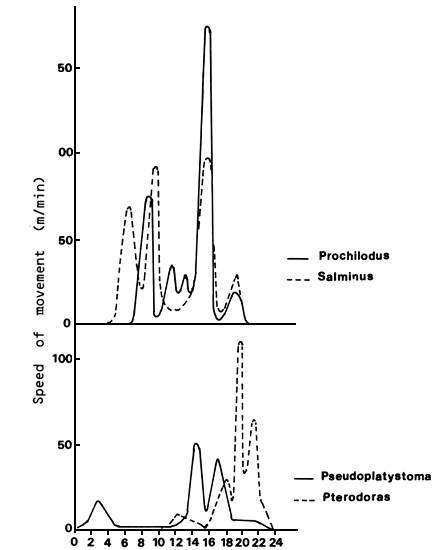
Figure 3 Daily movements and speed distributions for Salminus maxillosus, Prochilodus platensis, Pseudoplatystoma coruscans and Pterodoras granulosus (Poddubnyi, Espinach Ros and Oldani, 1981)
The fish were active for 8–12 hours a day with 1 to 3 peaks of activity during that time. Prochilodus and Salminus followed the gradients of flow velocity and Pseudoplatystoma and Luciopimelodus, the contours of the bottom. Generally speaking, they tend to avoid the zones where flow velocity is greatest. Mean daily migration rates upstream varied between 5 and 17 km/day, depending on the species (Poddubnyi, Espinach Ros and Oldani, 1981). These coincide with the mean rates obtained through tagging and recapture experiments (Poddubnyi, Espinach Ros and Oldani, 1981; Welcomme, 1985; Petrere, 1985).
Delfino, Baigún and Quirós (1986) reported that fish approach the Salto Grande dam (see Section 6.2) following the zones where water flow is slowest.
Virtually nothing is known about the “burst” speeds of Latin American fish and still less about how speed is related to fish size and functional morphology or water temperature. Current knowledge only allows us to set lower limits for maximum speeds. For Salminus and Prochilodus, over 30 cm in length, these speeds would exceed 1.3 m/sec at 11°C (Delfino, Baigún and Quirós, 1986), and for Salminus, Leporinus, Prochilodus and Triurobrycon, over 30 cm in length, they would exceed 2.3 m/sec at temperatures of over 20°C (Godoy, 1985; Castello, 1982).
Godoy (1985) reports that in the Urubupungá Falls on the Paraná river and below the Ilha Solteira structure, tagged specimens of Prochilodus scrofa, Salminus maxillosus, Leporinus copelandii and Pimelodus clarias were recaptured above 18 m high falls where flow velocity is 13.8 m/sec. In a similar experiment in Cachoeira Dorada on the Paranaíba river, Prochilodus scrofa and Triurobrycon lundii were recovered both in and above the rapids (Cachoeira). The rapids are 12 m high and water velocity is 9 to 12 m/sec. At the Sao Simal channel in the Paranaíba river, tagged Salminus and Prochilodus specimens ascended the 8 km of the channel where water velocity is 15 m/sec and, in the Rio Grande, individuals of the same species succeeded in moving against channel curents of 15 m/sec. In this latter experiment, the fish were recaptured at various points on the margins of the channel. In Cochoeira de Emas on the Mogi Guassu river, Prochilodus, Salminus, Leporinus and Pimelodus, as well as others, succeeded in ascending currents of 6 to 12 m/sec. They leapt more than 1.80 m to enter the fishway. At the Itaipava dam in the Parado river, Godoy and his team observed Prochilodus and Salminus moving against flow velocities of 20 m/sec, leaping 4 m high and 5 m in a forward direction (Godoy, 1985).
The speeds mentioned by Godoy (1985) appear to be a little on the high side for fish (Wardle, 1975). The flow speeds reported are mean maximum speeds at high velocity points (Godoy, pers. comm.). It is to be expected that some species follow courses which take them close to rocks or the walls of the channels, while others follow the relief of the river bed. In both these cases, flow velocities would be much lower (Godoy, pers. comm.).
Bonetto (1980) reports having observed Salminus maxillosus, Salminus hilarii and Prochilodus platensis leap over obstacles at the Cachoeira de Emas fish pass, while Leporinus copelandii swam through the layer of water.
At the foot of the Los Quiroga dam on the Dulce river, Salminus and Prochilodus were seen to leap 2 m high and 3–4 m in a forward direction in attempts to surmount the flow deflectors below the weir (Quirós, pers. obs.).
Although more precise data on the swimming capacity of Latin American fish are not available, the empirical formulae obtained by Zhou (1982), based on the contraction time for white, anaerobic muscle in six species, could be used for fish travelling in the column of water (Wardle, 1975). Maximum swimming speed is determined by both the length of the fish and water temperature (Beach, 1984). With these data, maximum swimming capacity may be predicted (Figure 4) and attraction water speeds at pass entrances, as well as auxiliary water supply and flow velocities in the installations may be determined (Beach, 1984). The time during which a fish can maintain speed (endurance) also depends on its body length and the water temperature as well as on the amount of glycogen stored in white muscle cells. The endurance of a fish swimming at maximum speed can be predicted using another empirical formula, which takes account of the quantity of energy released through anaerobic glycolysis. Figure 5 shows the relationship between the time a fish can maintain maximum speed and its body length at different water temperatures. Endurance times shown in Figure 5 should be taken as absolute maxima to incur complete exhaustion. It is unlikely that a fish will reach that limit. however, fish tend to leap over obstacles during their reproductive migrations and this must make severe demands on their energy reserves and may well use some of their store of anaerobic glycogen (Beach, 1984).
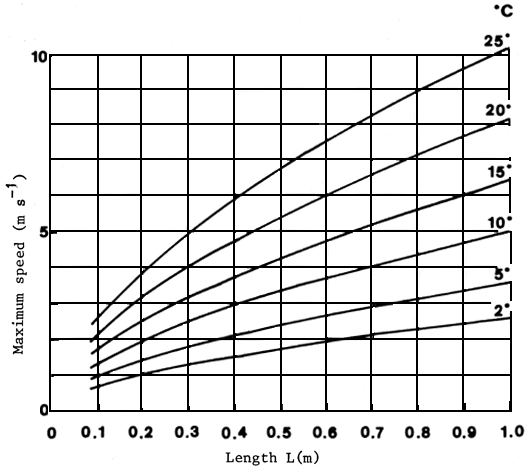
Figure 4 Maximum swimming speeds against fish length over a temperature range of 2° to 25°C (Beach, 1984)
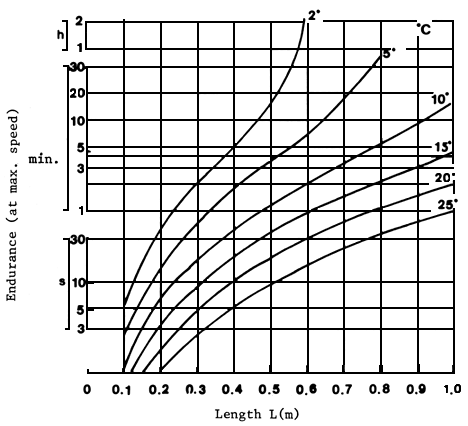
Figure 5 Endurance at maximum swimming speeds at various lengths of fish over a temperature range of 2° to 25°C (Beach, 1984)
![]()
![]()
![]()