![]()
![]()
![]()
Because relatively little is known about the productivity of small water bodies, some degree of predictive capability is required for management purposes. Numerous attempts have been made to produce models to predict fishery yields in larger water bodies. These predictions are very variable and are often associated with wide errors (Marshall, 1984; Cochrane and Robarts, 1986). One reason for this is that the catch statistics on which these models are based are often of low quality because of the inherent difficulties of collecting them. Most also require a set of relatively uniform conditions, which is an especially difficult criterion to fulfil where small water bodies are concerned. These factors need to be considered when attempting to use these relationships.
Rivers are characterized by flowing water, typically with low retention times and low productivity. The populations of primary producers are generally small and much of the organic matter in river systems is allochthonous, i.e., derived from outside sources (Dunn, 1989). Welcomme (1985) found that their fish yield was linked to the area of their drainage basins according to the relationship
C = 0.03A0.97 (r = 0.91)
where C = fish catch (tonnes) and A = area of the drainage basin (km2). This relationship can be transformed into one based on the length of the main channel, and becomes
C = 0.0032L1.98 (r = 0.90)
where L = the length of the main channel (km). These relationships were based on data from large rivers and their applicability to smaller ones has not been tested. Nevertheless, they do offer a means of making some estimate of the potential catch from small rivers, but the results should be treated with caution.
Swamps and floodplains are seasonally flooded areas with some of the characteristics of both rivers and lakes. They can be highly productive, since they derive their productivity from both the nutrients contributed by the water and organic matter produced on the floodplain during the dry season (Dunn, 1989; Hildrew, 1981). Welcomme (1985) found that the annaul productivity of various floodplains ranged from 5 to 143 kg/ha and developed a predictive relationship in which
C = 4.23A1.005
where C = catch (kg/ha) and A = area of the floodplain (km2). Welcomme points out that since the exponent is close to unity, the relationship is nearly linear, although larger floodplains are marginally more productive than smaller ones. Thus, a one-hectare plain would be expected to produce 42.8 kg/ha, compared to 44.6 kg/ha for one measuring 5 million ha. Again, this relationship was based on data from large floodplains, so the applicability to smaller ones has yet to be tested.
Reservoirs and, to a lesser extent, small lakes are the most common type of small water body in the region. They represent the resource in which the greatest increase in fisheries production is likely to take place. The two kinds of water body are very similar in most respects, except that in artificial lakes the inflow is generally large in relation to their volume and they have a lower water retention time than the natural ones (Dunn, 1989). These waters vary considerably because of climate, nutrient status and other factors, and they are correspondingly variable in relation to their fishing intensity. Henderson and Welcomme (1974) suggested that large lakes and reservoirs might be fully exploited when they support more than five fishers per square kilometre. Small ones seem to be able to support a higher density of fishers (Tables 18 and 19) because of their greater fish biomass (Figures 14 and 15).
The water quality of small, shallow reservoirs may not be significantly different from that of the river flowing into it, but they are frequently warmer and more eutrophic. Most artificial lakes tend to be most fertile immediately after their construction, sometimes reaching a peak after 3–4 years, followed by a drop, after which they stabilize at a lower level of productivity, but one which is still greater than that of the original river (Coche, 1974; McLachlan, 1974; Mitchell and Marshall, 1974). The final level of fertility can be greatly influenced by outside factors, especially nutrients added to the inflowing rivers, and the rate at which nutrients accumulate in the sediment. Blooms of toxic blue-green algae can occur in eutrophic systems (Marshall and Falconer, 1973; Scott et al., 1980). These blooms can also develop in small dams in arid areas when nutrients become concentrated as water levels fall. Animals drinking the water at these times are known to have been killed, but the effects on fish are currently unknown (various sources, cited in Toerien, Scott and Pitout, 1976). They have also occurred in waters in humid areas, such as Lake Muhazi, Rwanda, during the 1970s (M. Maes, unpublished data).
The productivity of lakes will, in the first instance, depend simply on their size. Youngs and Heimbuch (1982) developed a simple relationship between the fish catch and the area of North American lakes. Their approach was followed by Marshall (1984) for some African lakes, and fish yield could be estimated from
logeY = 3.57 + 0.76logeA
in which Y = fish yield (t) and A = lake area (km2).
A rather more complex approach was used in Thailand, where data on the fish yields and physical variables of 17 reservoirs (> 1 000 ha in area) were collected (Chookajorn, 1988). The following relationship was obtained by means of a multiple regression analysis:
Y = 327.65 + 0.03A + 0.05C - 30.78z
where Y = fish yield (kg/ha), A = the surface area of the lake (ha), C = the area of the lake's catchment (km2) and z = the mean depth of the lake (m). The main drawback to this approach is that it is based on water bodies that differ climatically, edaphically and in their species composition from any in southern Africa. Consequently, any predictions made with it should be treated conservatively. A similar approach has not yet been applied to African lakes, but it may be a rewarding route to follow, since it takes into account the relative supply of nutrients (using the lake's catchment area as an index) and their mean depth.
The effect of depth becomes important because of the assumption that shallow lakes are intrinsically more productive than deep ones, provided that they have the same chemical composition. This forms the basis of the well-known morpho-edaphic index, which is used as a means of predicting fisheries yield. It was originally calculated as total dissolved solids/mean depth, but in Africa it is more convenient to use conductivity/mean depth since this variable is much more widely measured and is equivalent to total dissolved solids. Henderson and Welcomme (1974) were the first to apply it to African lakes and found that, in 17 intensively-fished lakes and reservoirs, yield could be predicted from the relationship:
Y = 14.3136 * MEI0.4681
Where Y = fish yield (kg/ha/yr) and MEI = the morpho-edaphic index.
In a later study, Marshall (1984) found that there were differences between natural and artificial lakes, and obtained the following relationships:
Y = 23.281 * MEI0.447 (artificial lakes)
and
Y = 7.889 * MEI0.595 (natural lakes).
These relationships suggest that reservoirs were generally more productive, perhaps because of the greater interaction between the water and sediments brought about by fluctuating water levels.
Most of the data used to calculate these relationships were based on relatively large lakes or reservoirs, and may not apply particularly well to small ones. However, the relationship obtained from reservoirs in Sri Lanka (de Silva, 1988) in which:
Y = 6.83465 * MEI0.9005
might be more useful. This relationship has similar drawbacks to that produced by Chookajorn (1988), although it may be more applicable, since fisheries in the reservoirs of Sri Lanka are dominated by tilapias introduced from Africa, which increases their similarities to those in southern Africa. On the other hand, reservoirs in Sri Lanka tend to be shallow, have a high draw-down and are often highly eutrophic (de Silva, 1988), which may affect the applicability of this relationship.
Table 21. Comparison between fish yields recorded in some southern African water bodies and predictions made by the relationships described in the text.
| Savory Dam | Lake Chivero | Hartbeesoort Dam | |
|---|---|---|---|
| Lake area (ha) | 11.5 | 2 630 | 2 000 |
| Catchment area (km2) | 4.2 | 2 230 | 4 144 |
| Mean depth (m) | 2.8 | 9.4 | 9.6 |
| Conductivity (μS/cm) | 285 | 172 | 670 |
| Morpho-edaphic index | 101.8 | 18.3 | 69.8 |
| Recorded fish yield (kg/ha) | 256 | 120 | 248 |
| Predicted annual fish yields (kg/ha) | |||
| [1] area | 596.8 | 104.5 | 346.1 |
| [2] areas and depth | 242.1 | 228.8 | 299.4 |
| [3] MEI | 124.7 | 55.8 | 104.4 |
| [4] MEI | 183.8 | 85.4 | 155.3 |
| [5] MEI | 439.2 | 93.7 | 312.7 |
| Mean predicted yield | 317.3 | 113.6 | 243.6 |
Notes: [1] Lake area relationship in Marshall, 1984.
[2] Lake area, catchment area and mean depth relationship in Chookajorn, 1988.
[3] Morpho-edaphic index (Henderson and Welcomme, 1974).
[4] Morpho-edaphic index for reservoirs (Marshall, 1984).
[5] Morpho-edaphic index in Sri Lanka (de Silva, 1988).
Data from Evans, 1982; Marshall, 1978a; Marshall and Falconer, 1973; and Cochrane, 1984, 1987.
The relative imprecision of these estimates was discussed at the beginning of this section, and this needs to be stressed. The degree of variability and possible usefulness of these relationships can be assessed by comparing the predictions with some real data (Table 21). The results of this approach are highly variable, but the predictions based on the morpho-edaphic index tend to be underestimates whilst those based on the area relationships tend to overestimate it. The mean of all the estimates were 24% above the recorded yield of Savory Dam, but only 8% and 2% below the recorded yields for Lake Chivero and Hartbeespoort Dam respectively. This suggests that using a suite of these models will permit a reasonably accurate first estimate of potential yield from a water body.
A final, possibly rather crude, approach to estimating yield would be to make an estimate of biomass using the relationships between the size of the water body and its fish biomass (Figures 14 and 15). It should be possible to harvest at least half this biomass annually, and so a rough initial estimate of yield could be obtained. Such an estimate is probably conservative, as most tropical fish species have production: biomass ratios that exceed 1.0. It increases in smaller fish, which may have ratios of up to 5.0 or 6.0 (Marshall, 1993) and these are the kind of fish which are likely to be most abundant in small water bodies.
The predictive relationships discussed in the previous section may be of some value in planning a fishery either when none exists or when it operates at a very low level. They are of less value in monitoring an existing fishery, and alternative approaches are needed. In an ideal situation it would be possible to determine potential yield by having sufficient data on the limnology of the water body and the biology of its fish. This is not possible, and the usual approach would involve some kind of stock assessment, which would enable managers to determine the effect of fishing indirectly through changes in mortality rates, growth and other demographic characteristics.
There is now a wide variety of suitable stock assessment methods, many of them relatively easy to use as they are based on length-frequency relationships and backed up by simple computer programmes, which allows the data to be processed rapidly (e.g., Pauly, 1983; Sparre, Ursin and Venema, 1989). They have not been widely applied in small water bodies, however, largely because there are so many of them and too few people available to collect the data that would be needed. Since the basic data cannot be collected, there is a clear need for simpler methods, which are based on readily available data and can be easily applied.
In theory, the size of a fish stock can be estimated by using a standardized fishing method, such as gillnets, and equations such as
C/f = qN, and therefore N = (C/f)/q
where C = the catch, f = the fishing effort, q = catchability coefficient, and N = the number of fish in the population. The data are not easy to obtain because gillnets (like other fishing gear) are highly selective. It is particularly difficult to investigate a stock made up of many different species, since the catchability of each will be different. Catchability will also be influenced by season, weather, visibility and other factors. This makes it difficult to compare data from different dams since they vary so extensively.
One solution might be to compare the length frequency distributions of the same species in different waters. Such a comparison could indicate the state of the stock, as intensive fishing tends to eliminate both the larger species and the larger individuals in any one species (Welcomme, 1985). This idea can be demonstrated by a comparison of the length frequency distributions of Oreochromis mossambicus in two small dams in Zimbabwe (Figure 21, taken from van der Mheen, 1993). The fish in the heavily-exploited Kudzwe dam were around 170 mm in length and only one length-class was apparent. In the less heavily exploited Nyamsizi dam, the fish were larger and two, possibly three, length classes were evident. By contrast, Oreochromis andersonii from an unexploited dam in Botswana were much larger and at least three length classes could be detected. Although the growth of tilapias is very flexible, this method seems to offer a rapid means of comparing fish stocks in different water bodies. The advantage of methods like this is that length is easy to measure and it is only necessary that the sample of fish should be large enough to be representative of the population.
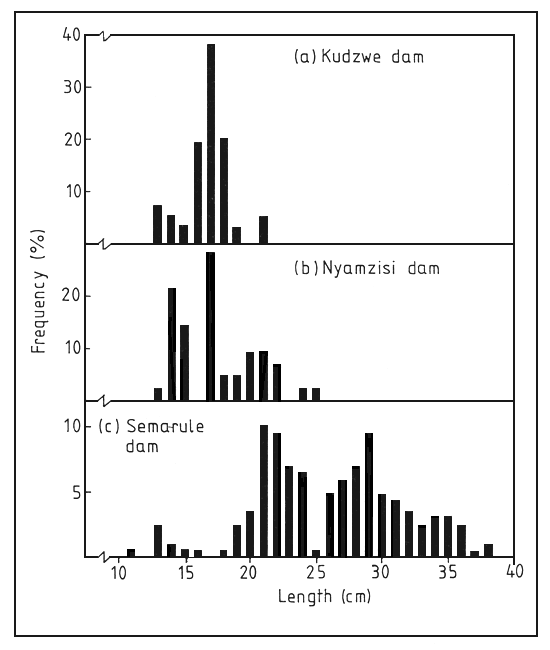
Figure 21. Length-frequency distributions of Oreochromis mossambicus in Kudzwe and Nyamzisi dams (Zimbabwe) compared with O. andersonii in Semarule dam (Botswana)
![]()
![]()
![]()