![]()
![]()
![]()
W.G. Cazemier
Netherlands Institute for Fishery Investigations
IJmuiden, The Netherlands
ABSTRACT
Bream grows very slowly in some Dutch waterbodies. In normal practice, stimulation of growth rate is attempted by removing part of such populations. The results are mostly poor, not durable and this type of management is very expensive. Alternative management measures have to be found by research.
In some cases bream even seem to lack the ability to increase their growth rate. By transplantation of very slowly-grown bream from a poor to a rich environment, it was established that even old bream could raise their growth rate to a remarkable extent.
The influence of the environment on growth and population density are discussed as well as the implications for such regulation of bream populations as thinning and stocking.
RESUME
Dans le cadre de recherches sur la régulation des populations de brèmes on a prélevé un échantillon de ces poissons dans un petit lac situé près de Langeraar, où, depuis au moins 60 ans, leur croissance est très médiocre, pour les transplanter dans un autre petit lac, Oude Veer, où leur croissance est à l'inverse très rapide. Dans le premier lac, la longueur totale des brèmes à l'âge de neuf ans est de l'ordre de 30 cm en moyenne alors qu'elle est de 50 cm environ dans le second, les poids correspondants se situant aux alentours de 300 et 1 500 g.
En l'éspace de trois ans, on a recapturé, sur un total de 1 517 brèmes marquées et transplantées, 440 individus. On a constaté que leur croissance avait été très bonne durant cette période et même supérieure à celle des brèmes autochtones. Leur état s'était également amélioré de façon remarquable.
La faible croissance des brèmes dans le lac de Langeraar pendant une longue période n'avait donc en rien réduit leur potentiel de croissance rapide. Il semble même que les brèmes habituées à une nourriture insuffisante aient un métabolisme ou un comportement alimentaire qui leur permet, après transplantation dans un milieu plus favorable, de se développer mieux que les autres.
Il est presque certain que l'accélération de la croissance des brèmes transplantées est due essentiellement à des différences d'alimentation d'un lac à l'autre. A Oude Veer, la biomasse benthique est beaucoup plus importante que dans l'autre lac, dans lequel la population de brèmes est nettement plus dense.
L'exploitation ou la régulation artificielle des stocks de brèmes ne peut pas être à l'origine de la différence de densité observée d'une pièce d'eau à l'autre. Selon toute probabilité, la régulation est due à des mécanismes indépendants de la densité. L'auteur examine ces mécanismes et souligne qu'il est capital de bien les comprendre pour aménager correctement les nombreuses populations de brèmes des Pays-Bas.
Adjustments of the population of bream (Abramis brama) by removing a portion of such populations are more and more common in the Netherlands. Such measures are intended to cope with the problems of stunted growth of this species and to decrease competition for food (mainly benthos) with the eel (Anguilla anguilla). Sportfishermen are interested in fast growth of bream and professional fishermen in high yields of their eel fisheries. The ecological mechanisms regulating bream stocks are not well known at the moment. In some water bodies the population density stabilizes at a high level, while in others the density remains low. As far as is known, fishing mortality plays an unimportant role. Most probably the mortality of eggs, larvae and juveniles forms the key factor in this phenomenon.
Opinions of population reductions as a management tool are somewhat divergent because, among other things, results are disappointing in most cases; the effects are not lasting, and the evaluation is mostly rather poor.
An important problem was that in some populations bream appear unable to increase their growth rate, possibly through a lack of appropriate genetic strains in the population. If this does play a role in bream, it is not surprising that lowering the number of fish per hectare, which means an increase of the benthos available per fish, is not always followed by a significant improvement of the growth rate. Therefore an experiment was designed whereby bream from a water body (Langeraarse Plas) having a dense population of fish in which individual growth has been consistently poor for at least 60 years (Redeke, 1923) were transplanted to a water body in which bream growth rate is very high (Oude Veer - Fig. 1). The growth and condition of the transplanted bream were determined over a period of three years, from the autumn of 1978 until April 1981, by examining recaptured specimens.
Sampling fish food-organisms and measuring some chemical and physical parameters was carried out from September 1973 until March 1976 in five water bodies. Fish stock assessments were made in these in March 1975 and in March 1977. This work was done prior to the transplantation of bream, in March 1978. Langeraar and Oude Veer are two of the five water bodies but only the results from these two are discussed in this paper.
Information from the other water bodies as well was published by Cazemier (1975), Cazemier and Van der Heul (1979) and Cazemier (1982).
In both water bodies a number of physical and chemical parameters were measured during 30 months (14 times) by using normal standard methods. The characteristics of the bottom were determined by direct observations, completed with information from soil maps. Plankton and other small organisms were sampled with nets.
Two surveys were carried out to estimate the population of bream, using a bottom trawl (3.5 m wide), towed by two boats equipped with 25 hp outboard motors (see Backiel and Welcomme, 1980). A number of short hauls (five minutes) were made over measured distances. The efficiency of the trawl was considered equal in both waters, because their characteristics for trawling were to a large extent similar. The actual efficiency was not known but could be estimated as rather high - about 75–90 percent.
To determine the rate of growth, ageing was carried out by counting annual marks in scales and finrays. Back calculations of lengths in successive years were carried out by measuring the distance between the nucleus of scales and the rostral part of the annuli. These data were substituted in a fish length-rostral scale radius relationship, expressed by the following expressions:
r = 0.0012 l 1.4104 (bream shorter than 9.7 cm)
r = -0.5970 + 0.0142 l (bream longer than 9.7 cm total length)
(r = rostral scale radius in mm; l = total fish length in mm)
The condition factor for bream is expressed as a quotient:
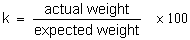
The expected weight corresponds to the mean weight determined for bream from a number of Dutch waters, which can be expressed by the formula:
g = 0.0053 l 3.1997
(g = weight in g; l = total length in cm)
An estimate of the average standing crop of benthos was made by regular sampling, 13 times during 26 months. The fresh weights of chironomid larvae (pupae) and oligochaetes only are given here because these organisms are the main food for bream bigger than 20 cm.
The transplanted bream were all caught on one day with a seine net in the Langeraarse Plas. Immediately thereafter they were transported and marked (1 March 1978) by injecting a small quantity of alcian blue in aqueous solution into the skin, with a Panjet Injector. A proportion of these fish was tagged by putting a small (1.4 × 0.3 cm) stainless steel numbered tag through an incision into the body cavity. Both marking methods had been successfully used in former experiments.
In Langeraar the turbidity is caused by a permanent bloom of Oscillatoria agardhii and in Oude Veer by flagellates, green algae and diatoms.
Chlorinity and total hardness in Oude Veer are very high for fresh water, due to seepage of marine water into the lake.
A reduced oxygen content was found only once when emanations fell to 61 percent saturation on a hot day near the bottom of Langeraar. Otherwise measurements always showed at least 80 percent saturation, and supersaturation was frequently observed.
Langeraar: very soft, muddy peat bottom, with a high content of dead plant material. The waterbody originates from peat-cutting activities.
Oude Veer: rather firm clayey mud with only a small amount of vegetable remains, made up of calcareous young sea claysoil. The water body lies in a polder reclaimed from the sea some 200 years ago.
Neomysis integer a small brackishwater shrimp, was estimated to be about five times as numerous in Oude Veer than in Langeraar. The species mostly swims about in dense shoals. Data on the abundance of other planktonic organisms are considered to be less relevant to the problem and therefore omitted here.
Fishing with bottom trawls were carried out in the spring of 1975 and 1977 in both water bodies. The figures in Table 3 refer to bream of over 20 cm (total length) and should be considered as minimum estimates of the stocks (see also Figs. 2, 3 and 4).
The number of juveniles, especially one-year old bream, was much higher in Langeraar than in Oude Veer. In 1977, for instance, 562 individuals were caught per 100 m trawl in Langeraar, against only 0.6 per 100 m in Oude Veer.
Bream contributed between 70 and 86 percent of the total catch and compared with the stock of bream the quantity of other benthivorous species was negligible. The only predatory species of importance for the bream population was pike-perch (zander) but is considered to be of minor importance as a factor in bream population-density regulation in these lakes.
The length frequency distribution for cm-classes above 20 cm, of bream caught during the trawl assessment fishery in March 1977, is presented in Figs. 2 and 3.
The growth of bream in Langeraar and Oude Veer was calculated by using data from a number of year-classes (Table 4).
The condition of bream over 20 cm was at the start of the transplantation experiment at an average of 101 in Langeraar and 109 in Oude Veer.
Estimates of average standing crop of chironomid larvae and oligochaetes in the period January 1974–March 1976 are presented in Fig. 4, together with a minimum estimate of that portion of the stocks of bream feeding on benthos.
The mean length of the transplanted bream was 30.8 cm (s = 3.5) on the day of stocking.
Within the three years after stocking 29.6 percent were recaptured in professional fishing gear, mainly gill nets.
Eighteen bream belonging to the original length group of 30 ± 2 cm (average age nine years) were recaptured in the period between 1 October 1980 and 1 April 1981 having completed three years in Oude Veer. By means of back calculation, the lengths at the end of 1978 and 1979 were determined.
The growth curve of this select group from their ninth year onward, is compared with the growth curves of both autochthonous groups in Fig. 5.
The ecology of the two bream lakes studied is very different. The water in Oude Veer is more alkaline, contains much more chloride and is much harder than Langeraar water (Table 2). The bottom structure and soil composition are also completely different. Both waters are eutrophic, however, have a low transparency, little aquatic vegetation, the same depth, and surface areas which are of the same magnitude (Table 1). The bream population in Langeraar is very dense and stunted as compared with that of Oude Veer (see Tables 3 and 4 and Figs. 2, 3, 4 and 5). The average standing crop of bottom fauna per kg of bream in Langeraar is only 10 percent of that in Oude Veer (see Fig. 4). The Langeraar bream population has already lived for at least 60 years under circumstances which though apparently favourable for a high density of the stock are not favourable for a high growth rate.
The main purpose of the investigation was to determine whether it would be possible for poorly grown bream to increase their growth rate and condition under more favourable circumstances.
The results show this to occur (see Tables 6 and 7 and Fig. 5). Enhancement of growth rate and condition were very striking and introduced bream, in fact, scored better in Oude Veer than the autochthonous ones in both factors, indicating that introduced fish adapted to poor conditions, could exploit the food supply more effectively than local ones.
The only way to achieve such growth rates and condition in Langeraar itself, would most probably be to reduce the bream population drastically and to keep it on a very low level of density. Increases in growth rate as a result of removing a part of a fish population are described by many authors (e.g., Walter, 1934; Schaperclaus, 1961; Weatherley, 1972; Hofstede, 1974; Bagenal, 1978). On the other hand, bream is a species which flourishes very well in most eutrophic waters (Willemsen, 1980) and managing dense, stunted populations by such adjustment fisheries seems to be rather problematic. Else Nielsen (1977) reports an experiment in Lake Arreso in Denmark in which a bream population was heavily exploited over many years. The growth did not become faster and the number of recruits rose, probably only because cannibalism became less. The total weight of bream per ha was only slightly reduced.
It looks as if there are mechanisms at work, which tend to stabilize the bream population density at a certain level, which is specific for each water body. The food supply seems to be of minor importance for this regulation. It was observed rather frequently, however, that the gonads of lean female bream out of a dense population were poorly developed, with a high percentage of small, white eggs (atretic eggs). In such extreme cases availability of food obviously plays a role in limiting populations.
Possibly, the natural survival rate of bream (especially in its first year) and the productivity of benthos (e.g., Schaperclaus, 1961) should be considered more or less fixed properties for water bodies. The latter is normally higher when the total hardness is higher as established by the author (Cazemier, 1982). Kennedy and Fitzmaurice (1968) also determined that alkalinity was positively correlated with bream growth rate in Irish waters.
It is very difficult to increase the food available to bream in natural waters, thus there is hardly any alternative other than trying to adjust the stock of bream to the food resources to achieve a better growth rate. However, when the density tends to stabilize at a certain level, the removal of part of a dense population (or conversely stocking to make a sparse population numerically denser) has to be repeated at regular intervals to be effective. This conclusion is supported by Backiel and Le Cren (1967), Bennett (1970) and Nielsen (1977) as well as the results of many adjustment fisheries and stockings of fry or older fish in Dutch waters.
The costs of such a regulatory practice (removing dozens of kg of bream per ha each year by seining in winter) are very high and especially so in the case of bream and are generally not acceptable.
In some water bodies there are natural regulatory mechanisms at work, which seem to be lacking in overcrowded waters. It is the aim of further investigations to find out the nature of these mechanisms. Most obviously, the survival rate (and thus mortality) in the first months of life is decisive for the year-class strength (see also the remark in 3.4). Predation by some organisms probably plays an important role. Because Neomysis integer is so numerous in Oude Veer, experiments were carried out to reveal if this small shrimp could be a predator on bream larvae. We found this was indeed the case; the shrimp seems not to chase this prey but predation can follow when it hits a larva. Without any doubt there are other important causes as well (Zuromska, 1966, 1967; Blaxter, 1974). Revealing and quantifying these causes of mortality will be of great use for solving the problem of stunted bream populations in an effective and durable way. In some cases, changing the environment by, for instance, lowering the eutrophication, will obviously be more effective than manipulating the population itself.
I should like to thank Dr. J. Willemsen for the advice he was always ready to give for the experimental work and for preparing this paper.
Backiel, T. and E.D. Le Cren, 1967 Some density relationships for fish population parameters. In The biological basis of freshwater fish production, edited by S.D. Gerking. Oxford, Blackwell Scientific Publications, pp. 261–93
Backiel, T. and J. Zawisza, 1968 synopsis of biological data on the bream, Abramis brama (Linnaeus, 1758). FAO Fish.Synop., (36):pag.var.
Backiel, T. and R.L. Welcomme (eds.), 1980 Guidelines for sampling fish in inland waters. FIFAC Tech.Pap., (33):176 p.
Bagenal, T., 1978 Methods for assessment of fish production in fresh waters. IBP Handb., (3):365 p. 3rd ed.
Bennett, G.W., 1970 Management of lakes and ponds. New York, Van Nostrand Reinhold Company, 375 p. 2nd ed.
Blaxter, J.H.S. (ed.), 1974 The early life history of fish. Berlin, Springer Verlag, 765 p.
Cazemier, W.G., 1975 Investigation about the main causes of different growth rates of the bream, Abramis brama. Visserij, 28(4):197–208 (Preliminary report in Dutch, with English summary and discussion)
Cazemier, W.G., 1982 The growth of bream, Abramis brama L., in relation to habitat and population density. Hydrobiol.Bull., 16(2/3):269–77
Cazemier, W.G. and J.W. van der Heul, 1979 Groeiverbetering van brasem door overplaatsing. Visserij, 32 (7/8):471–81
Hofstede, A.E., 1974 The application of age determination in fishing management. In Ageing of fish, edited by T.B. Bagenal. Old Woking, Surrey, Unwin Brothers Limited, pp. 206–20
Kennedy, M. and P. Fitzmaurice, 1968 The biology of the bream Abramis brama (L.) in Irish waters. Proc.R.Ir.Acad., 67(B5)
Le Cren, E.D., 1972 Fish production in freshwaters. Symp.Zool.Soc.Lond., 29:115–33
Nielsen, E., 1977 The development of the fishery in Lake Arreso with special reference to the bream population (Abramis brama L.). Folia Limnol.Scand., 17:27–35
Redeke, H.C., 1923 Rapport omtrent onderzoekingen aangaande den groei van den brasem in verschillende wateren. Verh.Rapp.Rijksinst.Vissch., 1(2):221–53
Schaperclaus, W., 1961 Lehrbuch der Teichwirtschaft. Berlin, Paul Parey, 582 p.
Walter, E., 1934 Grundlagen der allgemeinen fischereilichen Produktionslehre. Hand.Binnenfisch., 4:483–663
Weatherley, A.H., 1972 Growth and ecology of fish populations. London, Academic Press, 293 p.
Willemsen, J., 1980 Fishery aspects of eutrophication. Hydrobiol.Bull., 14(½):12–21
Zuromska, H., 1966 An assessment of levels and causes of mortality among eggs and fry of Rutilus rutilus. Doctor's thesis, Faculty of Fisheries, College of Agriculture, Olztyn, Poland, 62 p.
Zuromska, H., 1967 Mortality estimation of roach (Rutilus rutilus L.) eggs and larvae on lacustrine spawning grounds. Rocz.Nauk Rol., 90(H-3):539–56
Zuromska, H., 1967a Some causes of mortality of roach (Rutilus rutilus L.) eggs and larvae on lake spawning grounds. Rocz.Nauk Rol., 90(H-3):557–79
Table 1 Area, depth and transparency in Langeraar and Oude Veer
| Transparency in cm (Secchi-disk) | ||||
| Water body | Area (ha) | Average depth (m) | Spring | Summer and autumn |
| Langeraar | 70 | 2.2 | 35 – 60 | 20 – 25 |
| Oude Veer | 55 | 2.1 | 50 – 55 | 25 – 30 |
Table 2 Range and mean of some chemical parameters from 14 samplings during the period September 1973 to February 1976
| Langeraar | Oude Veer | |||||
| Parameter | min. | max. | mean | min. | max. | mean |
| Acidity (pH) | 7.6 | 8.9 | 8.4 | 7.9 | 8.8 | 8.4 |
| Alkalinity (m. eq) | 2.9 | 4.6 | 3.5 | 4.2 | 6.9 | 6.0 |
| Chlorinity (mg/l) | 131 | 187 | 156 | 536. | 2 030 | 1 472 |
| Total hardness (Ca2+ + Mg2+; °D) | 15 | 20 | 16 | 40 | 72 | 57 |
Table 3 Results of assessments of bream stock (specimens longer than 20 cm) on two occasions; estimated by numbers and weights (kg) per ha
| March 1975 | March 1977 | mean | ||||
| Water | number | weight | number | weight | number | weight |
| Langeraar | 1 751 | 391 | 1 011 | 329 | 1 381 | 360 |
| Oude Veer | 203 | 251 | 166 | 206 | 185 | 229 |
Table 4 Growth rate expressed as mean total length at age (rounded to nearest 0.5 cm)
| Age | |||||||||
| Water | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 |
| Langeraar | 6.5 | 11.0 | 15.0 | 18.5 | 22 | 25 | 27.0 | 28.5 | 30.0 |
| Oude Veer | 7.0 | 13.5 | 20.5 | 27.5 | 34 | 39 | 43.5 | 48.0 | 50.5 |
Table 5 The numbers of bream stocked (1/3/78) and recaptures (till 1/4/81) in Oude Veer
| Mark | Numbers marked and stocked | Numbers recaptured 1/3/78 to 1/4/81 | ||
| Blue spots and metal tag | 928 | 61.2 % | 267 | 59.5 % |
| Blue spots only | 589 | 38.8 % | 182 | 40.5 % |
| Total | 1 517 | 100.0 % | 449 | 100.0 % |
Table 6 The average condition (k) of bream of the three different groups
| Time of sampling | ||||
| 1/3/78 | Winter 1978 | Winter 1979 | Winter 1980 | |
| Langeraar bream | 101 | 97 | 95 | 87 |
| Oude Veer bream | 109 | 110 | 114 | 113 |
| Transplanted bream | 101 | 116 | 116 | 118 |
Table 7 Mean length (∆1) and weight (∆w) increments per original length-group, during three years after the transplantation from Langeraar to Oude Veer
| Length group (cm) | Langeraar autochthonous | Oude Veer autochthonous | Transplanted bream | |||
| Feb. 1978 | ∆1(cm) | ∆w(%) | ∆1(cm) | ∆w(%) | ∆1(cm) | ∆w(%) |
| 21 – 25 | 8.5 | 132 | 15.6 | 428 | 16.3 | 537 |
| 26 – 30 | 6.0 | 69 | 14.2 | 292 | 14.0 | 309 |
| 31 – 35 | 5.3 | 47 | 11.4 | 147 | 13.0 | 249 |
| 36 – 40 | 5.7 | 61 | 7.2 | 77 | 13.7 | 212 |
Fig. 1 Map of the Netherlands with the locations of the two lakes mentioned in this contribution
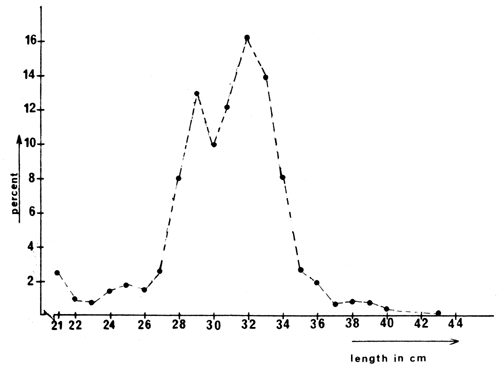
Fig. 2 Length frequency distribution of bream longer than 20 cm caught with bottom trawl in Langeraar, March 1977
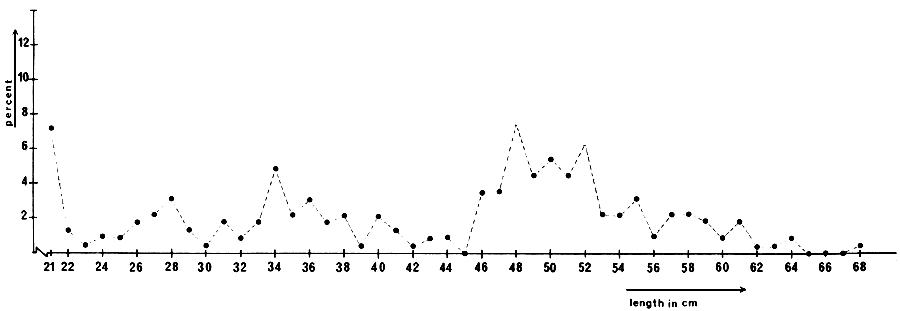
Fig. 3 Length frequency distribution of bream longer than 20 cm caught with the bottom trawl in Oude Veer, March 1977
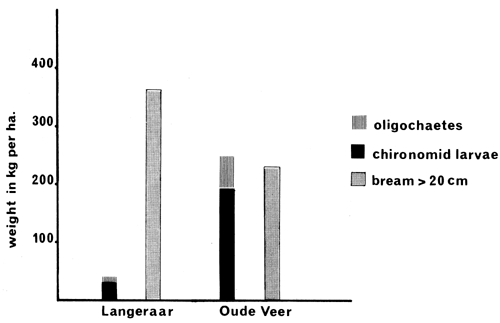
Fig. 4 Average standing crop (fresh weight) of chironomid larvae and oligochaetes (January 1974 – February 1976) and of bream (see Table 3)
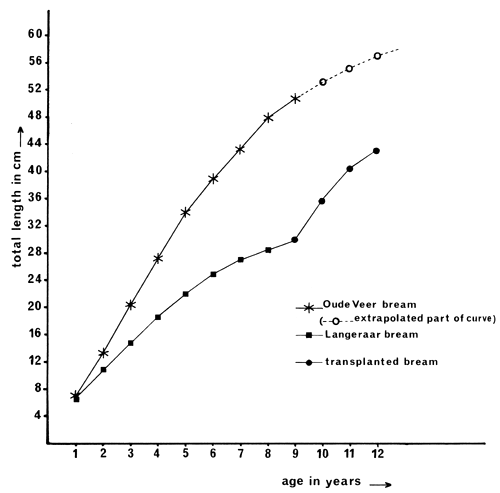
Fig. 5 Growth curves of bream in Langeraar and in Oude Veer and of a group that was transplanted at the average age of nine years from Langeraar to Oude Veer
![]()
![]()
![]()