![]()
![]()
![]()
JONATHAN W. WRIGHT
JONATHAN W. WRIGHT is Professor of Forest Genetics, Forestry Department, Michigan State University, East Lansing, Michigan, U.S.A. Other members of the drafting team were C. Heimburger (Canada), E. W. Jones (United Kingdom) and O. Fugalli (FAO).
The role of species hybridization in practical tree improvement programs varies. In Castanea and Populus species (other than the aspens) it has been given top priority because of the ease with which crosses can be made and because hybrids exhibit properties not obtainable by selection within a species. In some species of Pinus and Picea, hybridization will be used in conjunction with provenance testing and individual-tree selection the combination promising much more than any single approach. And there are some species that have resisted all attempts at crossing or are clearly superior to the hybrids produced and tested thus far.
Natural hybrids are common but probably represent only a small proportion of the total number of combinations which can be made. A study of the available data on natural crossing is always a desirable starting point for a serious hybridization program. Sometimes a breeder may have to rely wholly on natural crosses because controlled pollinations are difficult to make in reasonable quantity. But in other cases the controlled pollinations are so easily made that extensive artificial hybridizations are clearly indicated.
Extensive natural introgression is relatively uncommon but interesting when it occurs. From this it is possible to obtain a theoretical insight into the nature of genetic differentiation and of natural selection. Also, the great genetic variability in a hybrid swarm makes a desirable starting point for selective breeding programs.
Few intergeneric hybrids are known, the unqualified term "hybrid" usually meaning a cross between two species. In Populus and Castanea the crossability pattern is relatively simple and almost any species can be crossed with other species in the same genus. The pattern is a little more complex in Picea, where morphologically similar and neighboring species can be crossed much more easily than those which are geographically separated or are unlike. At the other extreme is Pinus, which is broken up into a large number of crossability groups and these groups correspond very well with taxonomic series and sections. Some groups contain several species whereas others contain only a single species which is so well differentiated that it does not cross with anything else. Knowledge of the crossability pattern is valuable because it permits a breeder to concentrate on those crosses with the greatest chance of success.
Recent work with mutation-inducing agents and mixed pollinations shows promise sin extending the number of crosses which can be made successfully.
Hybrid vigor in the F1 generation is of relatively common occurrence, especially when dealing with exotic species or when testing hybrids on disturbed habitats. The occurrence of hybrid vigor usually cannot be forecast beforehand but must be determined empirically by testing plants on a number of habitats. So far little is known as to whether the superior performance of the crosses is due to dominance, overdominance, or interaction between genepairs which have additive effects in themselves. That knowledge is not necessary when making practical use of the F1's, but will govern whether or not the extra vigor can be fixed in later generations.
Some hybrid combinations offer promise even though they show intermediacy in the F1. The genus Castanea provides an example. Castanea dentata is desirable in most respects but lacks resistance to the chestnut blight. This can be obtained by crossing with Asiatic species which do not have desirable timber form or growth rate. Backcrossing for a number of generations offers a way to incorporate disease resistance and timber quality in the same trees.
In a program designed for the production of an heterotic F1 cultivar, emphasis will be placed on crossability, mass production techniques, and long-term tests in the localities of probable use. Somewhat different emphasis is needed if the god is a late-generation segregate. Theoretical considerations from population genetics indicate that the goal can best be reached by starting with family selection in the F3. Hence, hastening through the F1 and F2 generations would be desirable.
To obtain hybrid vigor or a late-generation segregate it is not necessary to start with distinct species, but only to start with different races or portions of clines within a species. Swedish tree breeders have been the most active in this field, crossing native races with continental races. Their preliminary results indicate that a great expansion of this within-species hybridization would be productive.
Species hybridization is a very interesting subject. The successful hybridizer who produces a previously unknown combination feels that he has contributed something new, which in all likelihood would not have been produced in nature. And there is a very good chance that this new combination is also useful. Many of our most important crop plants are of hybrid origin, and new hybrid trees have definite promise.
A complete hybridization study for a single genus is a lifetime undertaking. Work by taxonomists on rare natural crosses or on introgression will provide clues as to which crosses can be made; for example, natural hybrids indicate the crossability pattern in the genus Eucalyptus. These clues should be supplemented by a good deal of controlled pollination work to develop a satisfactory crossability pattern because many of the most easily crossed species do not occur together in nature. There is still much work to be done after the crossability studies, which will show that species A and B can be crossed and perhaps that the hybrids are promising. How can the hybrids be mass-produced, and which particular races or individual trees of the parent species should be used to give the best results? Those are questions the answers to which may need many years of research.
The role of hybridization varies among regions and among genera. There are groups such as Castanea in which the best timber species are deficient in a single character (in this case, disease resistance), which can best be obtained by crossing with another species. The majority of hybrids are intermediate between their parents in morphology and site requirements. Thus hybrids may be easily made but be of little value to forestry when large areas of intermediate habitats are not available. Sometimes disturbance by man has created the intermediate habitats on which intermediate trees are best adapted, showing hybrid vigor, and there are examples of this in Eucalyptus and Aesculus. Again, a species sometimes finds its largest use outside its natural range, in which case hybrids may prove better adapted, an example being the hybrid Larix decidua x leptolepis. Hybridization offers relatively little hope of improving the American Pinus resinosa, which has resisted nearly all attempts to cross it with other species. Id the southeastern part of the United States the Southern pines are unlikely to be improved by interspecific hybridization; they reproduce readily and grow fast, while there are no other species of Pinus which surpass them in wood specific gravity - one of their most important properties. In any case, hybrids must be tested empirically before their potentials can be realized.
Reports of natural hybrids between species belonging to the same genus are frequent. Also, there is one well-authenticated report of hybrids between tree genera, namely the cross Cupressus x Chamaecyparis. Evidence on the presumed Tsuga x Picea hybrid is less convincing.
The probability of obtaining natural hybrids is great when one sows seed collected from isolated specimens growing in an arboretum or park. If a species does not ordinarily self-pollinate under such conditions, most of the seed produced may be from crossing with another species. Also, the nursery practice is usually intensive enough to permit recognition of unusual types. Most Abies and Acer natural hybrids have occurred under such circumstances. So have a large number of those reported in Picea, Pinus Populus and Quercus.
In the majority of cases, natural hybridization leads to the production of one or a few F1 trees only. The group of hybrids is in existence for one generation, then disappears. The next occurrence of the same combination may be in a quite different locality. This is true for most American reports of Quercus hybrids. It is also true of some interesting pine hybrids, such as Pinus sylvestris x nigra and P. palustris x taeda.
In Populus, Pinus and Picea study of the once-occurring natural hybrids provided the initial impetus for the later intensive work on artificial hybridization, but it was the study of the thousands of artificial hybrids which provided most information. On the other hand, study of the once-occurring hybrids of Quercus and Aesculus has contributed most of the knowledge about hybridization in those genera because artificial hybrids would be difficult to produce in quantity.
Natural hybrid swarms are infrequent but instructive. They occur most commonly on "hybrid" habitats, especially on disturbed areas. The oaks of southwestern United States furnish some of the best-studied examples (Muller, 1952). In central and northern Texas Quercus havardii and Q. stellata typically occupy sandy and clay soils respectively. Hybrids are common at the zone of contact between the two soils but not away from the zone of contact. The "pure" form of the species is found on the "pure" habitats typical of that species.
In the same region Q. mohriana and Q. havardii occur on soils derived from limestone and sand respectively. Hybrids are found where the soil is derived from a mixture of limestone and sand but not on the "pure" habitats. In California the hybrid species Quercus x alvordiana is really a group of hybrid swarms between Q. douglasii and Q. turbinella and is found only on intermediate habitats where the two parents come in close contact. Some of these Quercus x alvordiana swarms have persisted for many generations since the close of the Pleistocene Period.
In Australia Eucalyptus dalrympleana was one of the components of the original forest on undisturbed sites. As a result of man's activities, many of those sites have been damaged badly by fire. Now such areas are being invaded by hybrid swarms of E. pauciflora x dives (Pryor, 1957) whereas under natural conditions the two species excluded each other because of competition. Several other examples of hybrid swarms of Eucalyptus have recently been found on disturbed habitats.
Aesculus sect. Pavia includes four species native to the central and southeastern United States. In the original forest the species remained distinct. If hybrids were formed they occurred singly or in very small groups. Now there are many road cuts and cut-over areas on which the hybrids outgrow the parents. On such intermediate habitats there are occasional hybrid swarms of A. glabra x pavia, A. glabra x octandra, and A. octandra x sylvatica. In the latter two cases hybridization has led to considerable gene flow from A. octandra to the other two species. In yet another combination, A. pavia x sylvatica there is considerable gene flow in both directions from one parental species to the other.
By introgression is meant an extended period of hybridization followed by a considerable flow of genes from one species to another. Frequently it is hard to distinguish between the coming together of species by introgression and the divergence of species or races. In Picea introgression is suspected of being a major factor contributing to the wide adaptability of the northern, circumpolar species although it was proved in only two cases (Wright, 1955); Picea engelmannii and P. glauca cross readily in interior British Columbia and Alberta. Evidence of one species is found in the other far from the zone of contact. The spruce population of northern Scandinavia and adjacent parts of the U.S.S.R. is a mixture of intergrades between P. abies and P. obovata. In other cases widely distributed species with adjacent ranges are known to cross artificially and are presumed to have crossed naturally when climate favored a range overlap. On the other hand, lack of genetic variability and lack of recent opportunity for hybridization are characteristic of the widely scattered southern Picea species.
Following the introduction of Populus deltoides from North America into southern Europe there has been a large amount of introgression from that species into P. nigra. Pure P. nigra is now rare in many localities and planting stock consists mostly of hybrids (Schreiner, 1959).
True introgression is rare in Quercus despite the large number of species which have been reported. Intermediates between the north European species Quercus robur and Q. petraea occur where their ranges overlap and may be hybrids. In southwestern United States Quercus x drummondii seems to be an introgressive derivative of Q. stellata with an infusion of Q. gambelii genes. In most other cases the species have maintained their identities even when growing in mixture with several other oak species (Palmer, 1948).
Three factors help to explain why hybridization sometimes leads to only a few intermediates and sometimes to hybrid swarms or introgression. First, there may be limited opportunity for crossing. A few natural hybrids have been reported from arboreta for the cross between P. strobus from eastern America and P. griffithii from the Himalayas. These hybrids frequently appear in northern Italy where the two species are often grown together in gardens and parks. The cross is easily made artificially and introgression might be expected if the parents had the same range. Differences in flowering time may be all that keep Acer rubrum and A. saccharinum from becoming one species. Natural hybrids have not been reported although the two species commonly grow together and can be crossed with ease.
Secondly, the hybrids may be sterile. As yet there are few instances in which this is known to be the case.
The third factor is the nature and amount of selection pressure. Suppose that genes A and a are allelomorphic to each other, with respective frequencies of p = .9 and q = .1. Under random mating the zygotes will be present with relative frequencies of (p + q)²= p²+ 2pq + q²= .81AA + .18Aa + .01aa. If selection favors the heterozygotes for one generation, the proportion of Aa trees will increase to a maximum (the theoretical maximum being .50). But if environmental conditions then change so that selection favors the homozygotes, the Aa trees will tend to disappear. With complete elimination of heterozygotes the hybrid group in which zygote frequencies were .81AA: .18Aa: .01aa will produce gametes (pollen and egg cells) in which the gene frequency is .988A: .012a. These gametes will produce a new generation of trees in which the zygote frequencies are .9757AA: .0242Aa: .0001aa. Notice that selection against the heterozygotes has reduced the frequency of Aa trees by about 90 percent and reduced the frequency of the less common homozygote (aa) by about 99.9 percent. In other words, very few a genes remain with which to form Aa trees in the following generation. If the direction of selection should again be reversed to favor Aa trees, the build up of a hybrid swarm would have to start afresh.
This mathematical example has two corollaries. First, the existence of a persistent hybrid swarm is strong evidence that selection favors heterozygotes or, in other words, that usable hybrid vigor is present. Second, the absence of a hybrid swarm does not necessarily mean that useful hybrids cannot be produced. Man can take advantage of hybrid vigor exhibited in two generations out of three whereas nature cannot.
If a breeder's interest is in the overdominance1 indicated by the presence of a hybrid swarm, he should attempt to mass-produce by artificial means F1 or possibly F2 populations because it is in those generations that the overdominance will be most pronounced. However, if he is not in a position to undertake such an intensive program he should not attempt an intermediate measure such as a genetic thinning. By another theorem from population genetics, it can be shown that such a hybrid population will come to a natural equilibrium when the
frequency of the a gene = q=
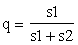
s1 and s2 being the selection pressure against the AA and aa trees respectively. It can also be shown that the population is most productive at this natural equilibrium (if it cannot be maintained in a completely heterozygous state by artificially crossing AA and aa trees). As an example, if AA, Aa, and aa trees have fitness ratings of .60, 1.00, and .85 respectively, the hybrid population will have an average fitness of .89 if at equilibrium. Artificial selection to favor the best trees (which would be the heterozygotes) would increase the number of unproductive AA trees and would lower the average fitness to .862 whereas artificial pollination to ensure that only aa trees were planted would increase the average fitness to 1.00.
1 Overdominance is the condition in which the heterozygote is superior to the best homozygote.
If a breeder's interest is primarily in additive genetic variance, a natural hybrid swarm forms an excellent starting point for a selection program. In southern Michigan there is a large group of hybrids between Pinus thunbergii and P. densiflora. In several characters the additive genetic variance within it is probably two or three times that found in any natural population of the pure species. Heritability and response to selection would also be two or three times as great as in a population of the pure species.
With the exception of Cupressus x Chamaecyparis, mentioned earlier in the chapter, intergeneric hybrids in temperate zone trees can be disregarded and attention can be focused on crosses within genera. The crossability patterns are best known in Populus, Pinus Picea Quercus, Castanea, and Larix. In those groups enough crosses have been attempted to formulate general rules which permit one to forecast which species combinations are most likely to be successful. For Acer, Aesculus, Betula, Fraxinus and Ulmus, information is more fragmentary and permits only the broadest generalizations. For most other genera (including all the tropical ones) it is even more fragmentary, consisting mostly of lists of natural hybrids.
The amount of genetic differentiation within what is considered to be a single genus varies greatly. At one extreme is Populus. There has been so little differentiation that a given species can apparently be crossed with any other species. Castanea may have a similar pattern - all attempted species crosses have been successful. Genetic differentiation is a little more pronounced in Picea. Most species can be crossed with their geographic neighbors but not with those species from a distant locality. There is no sharp break into distinct crossability groups and it is possible (but this has not been tested) that any species could be linked to any other one by crossing with intermediate types. Eucalyptus exhibits a similar pattern, neighboring species in the same section crossing with relative ease. Quercus is divided into 3 subgenera (only 2 are known genetically). A large number of hybrids between species belonging to the white oak subgenus Lepidobalanus have been reported and possibly any two white oaks could be hybridized. The same is true of the red oak subgenus Erythrobalanus. On the other hand, there are no convincing demonstrations of hybridization between the subgenera of Quercus.
The pines are at the other extreme. The genus Pinus is divided into 2 subgenera and a large number of series. With 3 exceptions attempts to cross members of the same series have been successful. The pattern in Acer is similar to that in the pines. Of 60 artificial crosses which have been attempted, 6 succeeded. The successes were all within one of the 13 recognized sections. From the genetic standpoint, series or sections in these 2 genera have approximately the same amount of differentiation as some entire genera. It would make sense biologically (but not practically) to regard these small groups of species as separate genera, as some authors have done in the past.
Morphological or physiological similarity and geographic distribution provide excellent clues as to the crossability pattern in a genus. The most easily made combinations are between similar species with nearly overlapping ranges. Often distance was a sufficient barrier to render unnecessary another type of isolating mechanism. If this barrier is removed by interplanting or by controlled pollination, hybrids result. On the other hand, species which occur in mixture in the same stands usually do not cross. Sometimes differences in flowering time or in microsite requirements provide an isolating barrier which can easily be overcome by artificial crossing. Sometimes the isolating barrier arises as the result of genic or chromosomal differentiation. Irradiation of pollen and the use of pollen mixtures offer some promise of overcoming such a barrier. It is also difficult to cross morphologically dissimilar species or species with widely separated distribution areas. In such cases, long separation has usually been accompanied by great genie or chromosomal differentiation.
Crosses which fail in one environment may succeed at another place or time. Knowledge of the nature of the barrier often helps to break it down.
Examples of the relationship between crossability, similarity, and geographic distribution are numerous, and for illustration the genus Pinus can be cited. The series Strobi comprises several similar species. All except one have separate (but neighboring, if considered in the geological sense) distribution areas and can be crossed with each other rather easily. The single exception (Pinus lambertiana) is not typical of the series and does not cross with the other members. Several attempts have been made to cross species of ser. Strobi with members of other series but only one such attempt succeeded.
Pinus ser. Sylvestres comprises about a dozen hard pines. The Asiatic members are morphologically similar and can be crossed with each other in nearly all possible combinations. They are less similar to the European representatives of the series. Only a few European x Asiatic crosses successful and these gave low seed sets. The single United States representative in series Sylvestres (P. resinosa) has several distinctive morphological and chromosomal characters. Over 20 years of effort by many people resulted in only 4 hybrid trees between P. resinosa and another species.
Pinus ser. Australes comprises groups of southeastern and southwestern United States species. Members of each group are more similar to each other than to members of the other group. All possible southeastern x southeastern species combinations and a great many southwestern x southwestern combinations have resulted in hybrids. However, no southwestern x southeastern combination has been successful (Duffield, 1952).
The spruces (Picea) also show a relation between crossability, similarity, and distribution. In northern United States, Canada, and northeast Asia is a group of 6 species, each with a nearly separate distribution area and each with certain similarities to its neighbors. All attempted hybridizations within the group were successful but several attempts to cross those spruces with others outside it were not. Across northern Europe, Siberia, and northern China is another "chain" of similar species, within which crossing occurs very easily. The 6 species which inhabit the same mountain in Japan do not cross, but one of them can be crossed with mainland spruces. One of the notable exceptions to the general crossability pattern is the cross P. omorika x sitchensis, which has been successful in Europe.
The genus Larix contains about 10 species, each with its own separate geographic range. Apparently differentiation has been entirely geographic and the species cross easily.
Of 7 attempted artificial hybridizations in Fraxinus only one succeeded. That was between an eastern United States and a western United States representative of the same section. The 2 species can be considered neighbors in the geological sense and are so similar that one could mistake aberrant specimens of one for the other. Hybrid swarms are reported between members of a similar species-pair in southwestern Europe.
Several cases are known in which crosses between species with different chromosome numbers yielded good seed. The general relationship between chromosome number and crossability has not yet been worked out.
These generalizations can be summarized by pointing out that a good monograph and a detailed set of range maps are of great help in planning a species crossing program. If it is assumed that there are 90 species in the genus Pinus, there would be a total of (90 x 89)/2 = 4,005 different species x species combinations. Work to date indicates that 10 percent or 400 of these might be successful. A breeder who crossed at random would have only a 10 percent probability of success. But one who studied the taxonomy and distribution could increase this probability to 80 percent.
Most emphasis in species hybridization has been on the production of vigorous F1 hybrids. Enough work has been done to show that the approach is extremely promising. The majority of the hybrids are useless but enough possess hybrid vigor to make the work worthwhile. Often hybrid vigor is evident in one part but not in another part of the range of a species.
The introduction of a successful F1 hybrid combination into commercial practice is carried out in four stages. First comes a crossability study to determine what combinations can be made. As already indicated, this work is well advanced in some genera and scarcely started in others. Next comes the production of a few hybrids of each promising combination and their testing in comparison with the nonhybrids they might replace. A good start has been made in this direction, but in scarcely any case have the tests been extensive enough to indicate a hybrid's true potentialities. Most of the Pinus hybrids produced in California and Pennsylvania are scarcely known outside a few test localities; and the potentialities of Larix decidua x leptolepis, which is very useful in Europe, are scarcely known in America. The third stage is the determination of mass production possibilities. This is no problem in those species of Populus which can be propagated readily by cuttings, however, it is a very real problem in other genera. The Institute of Forest Genetics at Placerville, California, did enough work on several combinations of Pinus to show that mass production is possible. In recent years, the Korean Institute of Forest Genetics has done a large amount of work, and obtained extremely promising results, on a few Pinus combinations. The fourth step in introducing a successful F1 hybrid into commercial practice is determining which particular trees would make the best parents. This has barely started for any species combination. Take the cross Pinus sylvestris x densiflora for example. It was produced in the United States, originally in Connecticut and later in Pennsylvania. In both cases the first available trees, of unknown origin, were used as parents. The hybrids grew extremely fast but cannot be used because of poor form and low seed set.
The Pacific southwest and the northeastern forest experiment stations of the United States Forest Service have produced a large number of species combinations in Pinus but only a few can be mentioned here. Pinus monticola x strobus is one of the most promising. Seed sets are high enough to indicate good mass production possibilities; growth in California and in many other parts of the United States equals or exceeds that of the faster-growing parent; there is intensive work under way on breeding disease-resistant cultivars of both parent species. When the work is completed, truly remarkable F1 hybrids will be possible. The hybrid P. attenuata x radiata also shows promise. It could be mass-produced. Although it does not outgrow the faster-growing parent (P. radiata) on warm sites, it promises to be one of the most productive pines on sites too cold for that species.
Seed sets are very high for the hybrid P. densiflora x thunbergii. This combination shows considerable hybrid vigor in Pennsylvania, but not in Japan where the parents are native. The potential usefulness of a fourth combination, P. nigra x resinosa, is limited because of mass production difficulties. Only four hybrids were produced after many years' effort. Thus, these hybrids have good growth characters but remain botanical curiosities.
The potentialities of F. pine hybrids are being exploited to the full in Korea. There thousands of bags are used to make the cross P. rigida x taeda each year and the program is proving to be economic. The hybrids can produce the most income even though they are more expensive to produce than nonhybrids.
The cross Larix decidua x leptolepis is the subject of more than 27 different publications from as many parts of northern Europe, almost all reporting greater growth than for either parent species (for the references see Wright, 1962). Seed sets are high but Larix does not lend itself as well to mass-controlled pollinations as does Pinus Syrach Larsen of Denmark has therefore designed a seed orchard containing a self-incompatible clone of one species and seedlings or clones of the other. The self-incompatible clone serves as the female parent and produces hybrid seed.
A small amount of American and a much greater amount of Russian work has resulted in the production of several artificial hybrid combinations in Quercus. Some of these are intermediate between their parents but others exhibit useful hybrid vigor (Piatnitsky, 1960). However, at present even the latter appear to be of very limited usefulness. Controlled pollination is extremely tedious and likely to be too expensive to be done on a commercial scale. Vegetative propagation is impractical. Seed orchards where natural crossing can take place may be the answer, but efforts to find trees which set large percentages of hybrid seed after natural gross pollination have been unrewarding.
The silviculture of Populus is now based much more on hybrids than on pure species. Cultivars derived from crosses involving P. deltoides, P. nigra P. trichocarpa and other species are commonly planted in Europe. Most of these have not been compared with their parents but hybrid vigor is usually assumed. As the result of Schreiner's pioneer work in the northeastern United States, many promising hybrids have been tested in America and can soon be planted on a commercial scale.
Species of Populus other than the aspens can be propagated readily by cuttings so that mass production is no problem. Ease of rooting has also permitted breeders to conduct a large number of tests for clonal differences within F1 full-sib families. As expected, significant clonal differences were rarely found except when the parents were themselves hybrids.
In short-term tests Acer platanoides x mayrii and A. platanoides x cappadocicum exhibited some hybrid vigor. The crosses were easily made on an experimental basis at the rate of a few hundred per day. However, that rate could not be increased greatly because of the flower structure, and mass-controlled pollination does not seem feasible. Neither do seed orchards because of differences in flowering time. Polyploidy is possibly the answer. Perhaps the few hybrids now available could be made truebreeding by having their chromosome number doubled. But would the tetraploids be as desirable as the diploids? That must be determined.
Theoretically one can expect a certain amount of hybrid vigor to persist into the F2 generation. This raises the possibility of utilizing the F2 generation if the F1 cannot be produced cheaply. At least in some of the pines the F1's are sufficiently fertile for F2 seed to be produced in quantity. The Institute of Forest Genetics in California has been pursuing this possibility for several Pinus, combinations. In the United Kingdom there are plans to raise large F2 progenies of European x Japanese larch (Larix decidua x leptolepis) in the expectation that some hybrid vigor will persist.
In most of the illustrations quoted here, the hybrids are still young, since there are no adequate older tests of many of the most interesting combinations. Also, many of the preliminary growth tests have been conducted on only one or two sites, and it is well known that hybrid vigor may be evident at one place and not at another. There is, therefore, considerable testing still to be done.
Practically all the Pinus, Picea and Populus hybrids to date have been made with the first available parents. This leaves considerable work for future tree breeders. The first Pinus, densiflora x thunbergii hybrids produced in Philadelphia seem quite useful but it is inconceivable that they can be the best. It is much more likely that even better hybrids can be produced by crossing selected races of the two parent species or perhaps selected individuals within those selected races.
Almost every tree hybridizer hopes he will produce an F1 or F2 generation which possesses enough hybrid vigor to be worth planting on a large scale. That is not always possible. Sometimes segregates arising in a later generation will be the valuable product. This will be true under three conditions:
1. if controlled pollination is so difficult that F1's and F2's cannot be mass-produced, (as in Juglans, Castanea, Acer, Quercus and possibly Abies);2. if hybrid vigor is present but is not sufficient to make a plantable cultivar (Castanea dentata x mollissima);
3. if the genetic variance is mostly additive.
If it is desired to incorporate one character from species A with many characters from species B, back-crossing is indicated. The F1 hybrid between A and B should be produced artificially, then backcrossed to species B and the process repeated again if necessary. In crop plants four or five generations of backcrossing are frequently necessary. If it is desired to incorporate approximately equal amounts of species A and B, an F1 generation is produced; the F1's are crossed with each other to produce an F2, those are crossed with each other to make an F3, etc.
The necessity for this approach has been appreciated for many years but investigators have been loath to follow it because of the long periods of time involved. Such delays may be necessary but need not be interminable. Numerous, important tree species produce flowers at ages from one to five years. If a tree breeder had known their desirability at the start, within 20 years he could have had F4 generations in production.
At present most many-generation breeding projects are unplanned developments of F1 hybridization studies. This situation must be changed if the most rapid progress is to be made. The field tests designed to produce a later generation segregate should be different from those designed to test the usefulness of the F1 or F2 generation per se.
The aim in a many-generation project should be to produce and test an F3 or second generation backcross as rapidly as possible. Why? The F1 generation is a uniform one, offering almost no opportunity for selection. ² Suppose that there are 25 F1 seedlings and 10 of them fruit at a very early age. The genetic component of the variation in the F1 population is almost nil, so that it makes little difference whether those 10 are used as parents of an F2 population or there is a wait of several years in order to see whether the other 15 might have superior phenotypes. The F2 population is a genetically variable one and offers opportunity for genetic selection. But a large proportion of the F2 genetic variance is nonadditive and therefore cannot be fixed by crossing the best trees. This means that a breeder is justified in getting only a moderately good evaluation of the F2 because no matter how long he looks at the trees he cannot tell their breeding potential as well as if he were to wait until the F3 or F4.³ It is better to look at the F2 for relatively few years and to practice a mild selection. The 10 or 20 percent best trees might be chosen and used as parents of a replicated F3 test in which the identity of all families is maintained.4
² It is true that wild trees are variable. The F1 hybrid between race I of species A and race VI of species B will not be absolutely isogenic but will be relatively isogenic if considered in comparison with the mixture between the two. Selection within such an F1 offers so little chance of improvement (in comparison with that offered by selection in a later generation) that it can safely be neglected.
The ineffectiveness of F1 selection applies to selection within progenies. Suppose race I of species A is crossed with race VI of species B and race IV of species A is crossed with race II of species B. The two F1 progenies can be expected to differ and should form the foundation for two selection projects which might later be united.
³ The question of how long to assess the F2 before proceeding to the F3 cannot be answered categorically. There is probably enough additive genetic variance in an F2 population to make it a mistake to use the first 1 percent of the trees to flower if they were noticeably deficient in an important growth character. Pronounced hybrid vigor in the F1 and F3 is an indication of the presence of nonadditive variance. Hence, the greater the departure of the F1 and F2 from the midparent means, the shorter the F2 test period need be. If growth tests show the F1 to equal the midparent means in every character, selection could as well start in the F2 as in the F3.
4 The term family is used to designate the progeny of a single open-pollinated parent or the progeny of a single cross between two individuals.
It has already been mentioned that selection is more productive in the F3 than in the F2 generation because of the smaller amount of nonadditive genetic variance. Another reason for preferring the F3 is that it offers an opportunity to practice family selection. This means selecting breeding stock on the basis of a tree's phenotype as well as on the performance of other trees having the same parent(s). This is much more effective than selection on the basis of phenotype alone. (For a good discussion of the relative gains rom mass and family selection, see Lerner, 1958).
Should one use control-pollinated (both parents known) or open-pollinated (only the seed parent known) progenies when conducting a selective breeding program such as outlined here? The F1 and F2 populations can be relatively small, consisting of a few dozen or few hundred trees. They could well be control-pollinated. As long as a tree breeder is working with multiple factor inheritance (which is most likely), the F3 population should be very large, containing scores or hundreds of progenies and thousands of individual seedlings. The likelihood of success with small F3 tests is too small to warrant the trouble involved. In Populus Picea, and many Pinus species controlled pollination can be easily done on the required scale. On the other hand, workers with genera in which controlled pollination is laborious, as in Abies in which the female flowers are rather inaccessible, or Castanea Juglans or Acer in which each pollination yields few seeds, would be well advised to work with open-pollinated progenies in order to have tests of adequate size.
Vegetative propagation is commercially possible with many species of Populus. Hence, in that genus it may be desirable to establish clonal tests in the F2 or F3 generations in order to select clones with desirable properties. In genera such as Pinus and Quercus, clonal tests are expensive and probably not needed because seedling families will be planted commercially.
FIGURE 14. - Isolated female flowers on Pinus sylvestris.
Family selection in segregating generations will play a large role in the Castanea breeding program. The American C. dentata is a fast-growing timber tree with desirable growth habit and wood properties but without resistance to chestnut blight (Endothia parasitica). That must be furnished by the Asiatic C. mollissima or C. crenata, neither of which becomes a large timber tree. A moderate number of F1 hybrids have been produced which show more or less intermediacy between the parents. Also, moderate numbers of F2's and backcrosses have been produced. Some of the F2 segregates have good vigor and much more resistance than the C. dentata parent. That is not enough, however, and it appears that large F3 and F4 generations must be raised to yield a cultivar which is commercially acceptable for timber production.
A great deal of the Populus breeding work in southern Europe is probably aimed toward the production of desirable later-generation segregates. However, no formal many-generation program has been started, selection being among trees of unknown generation.
Several F1 Pinus hybrids have reached fruiting size at the Institute of Forest Genetics in California. Most have been fertile. One of the most promising crosses is P. jeffreyi x coulteri. Some F2's and backcrosses exhibit the desirable growth characters of the former and resistance to the pine reproduction weevil (Cylindrocopturus eatoni) of the latter. The degree of recombination is sufficient to indicate that a group of F3 families might comprise a commercially acceptable cultivar.
The crossing of different races within a species offers the same possibilities as does the crossing of different cultivars of a crop plant or different breeds of animals. The F1 hybrids may exhibit hybrid vigor as do crosses between inbred lines of maize. Or they may exhibit a combination of desirable characters and furnish the foundation for a several-generation selection program.
The pioneer German tree breeder, Dengler, made a start on interracial hybridization in 1926 when he crossed German, French, and Scottish origins of Pinus sylvestris He thought some combinations showed hybrid vigor at 10 years old, but Scamoni later reported, when the trees were 20 years old, that the hybrids were intermediate between their parents.
In 1948 Johnsson and his co-workers at Ekebo, Sweden, made a systematic series of crosses among European aspens (Populus tremula) originating at three different latitudes in Sweden. At 10 years all the hybrids were considerably superior to the midparent mean and one combination (southern x central) was taller than the faster-growing parent.
Other Swedish tree breeders, especially Nilsson and Langlet, have had promising results from interracial hybridization in Pinus sylvestris and Picea abies. These were seen during the study tours preceding the world consultation. Following their lead, workers in other regions will undoubtedly show greatly increased interest in this subject.
Good collections of breeding material are the prime requisite for productive hybridization work. The breeder interested in determining a crossability pattern needs several flowering specimens of several races or species. The breeder interested in mass-producing a particularly promising hybrid needs many selected specimens of each parent (Figures 11 and 12). One or two specimens of each parental type are not enough because they may not fruit nor give enough nonhybrid seed to provide controls.
In temperate climates a period in the spring should be reserved for at least five years when embarking on a serious hybridization program. Time is needed for bagging, pollinating, removal of bags and observation of flower development (Figures 13 and 14). Seed sets are often so variable that the same cross needs to be repeated a few times to establish a reliable average pattern.
A hybridizer needs a small but good research nursery and several test sites. Usually few seeds are available of combinations which are made for the first time. This means that the testing must be done with greater precision than is usual in planting research. Both nursery and experimental forests must, therefore, be equipped to give the tests better than average care.
If the breeder is interested in developing both F1 and F3 cultivars, duplicate test plantings may be necessary. The design used to determine comparative growth rates of the F1's and their parents may not be the one which would insure earliest fruit production. Also, a good design to test hybrid vigor would not necessarily be the one used to obtain F2 and F3 hybrids by natural open pollination.
DUFFIELD, J. W. 1952. Relationships and species hybridization in the genus Pinus. Z. Forstgenet., 1: 93-100.
LERNER. I. M. 1958. The genetic basis of selection. New York, Wiley. 298 p.
MULLER, C. H. 1952. Ecological control of hybridization in Quercus: a factor in the mechanism of evolution. Evolution, 6: 147-161.
PALMER, E. J. 1948. Hybrid oaks of North America. J. Arnold Arbor., 29: 1-48.
PIATNISKY, S. S. 1960. Evolving new form of oak by hybridization. In Questions of forestry and forest management. Proc. 5th World For. Congr., Seattle, U.S.A., p. 231-244.
PRYOR, L. D. 1957. Selecting and breeding for cold resistance in Eucalyptus. Silvae Genet., 6: 98-109.
SCHREINER, E. J. 1969. Production of poplar timber in Europe and its significance and application in the United States. U.S. Dept. Agric. Handbook 150. 124 p.
WRIGHT, J. M. 1955. Species crossability in spruce in relation to distribution and taxonomy. For. Sci., 1: 319-349.
WRIGHT, J. W. 1962. Genetics of forest tree improvement. Rome. FAO Forestry and Forest Products Studies No. 16. 399 p.
![]()
![]()
![]()
{kind=link}
{kind=link}
{kind=link}
{kind=link}