![]()
![]()
![]()
ÅKE GUSTAFSSON et FRANÇOIS MERGENÅKE GUSTAFSSON est Directeur de l'Institut de génétique forestière, Collège Royal de Sylviculture, Stockholm Suède. François Mergen est professeur de génétique forestière, Université de Yale. Ecole forestière de New Haven Connecticut, Etats-Unis. Les autres membres de l'équipe de rédaction étaient K. Sax (Etats-Unis) et G. Sirén (FAO). KARL SAX (Etats-Unis), Président de la section 1 de la Consultation, a préparé le résumé.
Résumé
La cytogénétique fournit les fondements de la variation héréditaire, et il est indispensable de déterminer quelle est la constitution des chromosomes et la nature de la variation génétique dans les espèces forestières. Quel est le rôle de la polyploïdie et des modifications de structure des chromosomes chez les variétés et chez les espèces? Quelles différences génétiques résultent des modifications dans l'équilibre des chromosomes ou des changements de structure et dans quelle mesure la variation est-elle due à des mutations en un «point»?
Les genres de gymnospermes qui présentent un intérêt en matière forestière ont de grands chromosomes, la polyploïdie est rare et les caryotypes sont semblables à l'intérieur des genres. Dans les Pinacées, les nombres chromosomiques de base sont 10, 11, 12 et 13, ce qui a amené Khoshoo à conclure que le nombre de base originel était 10, et que les nombres de base plus élevés dérivaient des autres par fragmentation des chromosomes. Ceci a été bien démontré dans Pseudolarix et Podocarpus. Le nombre de base de 12 paires de chromosomes est pourtant caractéristique de la plupart des genres des Pinacées et il est possible que les genres qui possèdent un nombre de base inférieur aient pu dériver d'une union de chromosomes faisant suite à une translocation inégale. On a montré que la variation des nombres de base des chromosomes dans certains genres d'angiospermes résultait, d'une part, de la fragmentation des chromosomes et, d'autre part, de l'union des chromosomes.
La polyploïdie est rare chez les Pinacées bien qu'on trouve des polyploïdes dans la nature et qu'on puisse les provoquer. Mergen a trouvé que les polyploïdes spontanés et provoqués ont une croissance retardée et ont un taux de survie faible. Les expériences très étendues de Hyun sur les polyploïdes provoqués chez les conifères fournissent davantage encore de renseignements nécessaires.
La grande taille des chromosomes chez les Pinacées les rend très sensibles aux radiations ionisantes. Gustafsson a montré il y a de nombreuses années que les graines de pins sont détériorées par des doses relativement basses de rayons X. Il faut considérer ces faits en matière d'amélioration par mutation et quand on évalue la valeur relative de la radiation par rapport aux agents mutagènes chimiques.
Au contraire des gymnospermes, les genres d'angiospermes intéressants en matière forestière ont un nombre chromosomique de base qui varie beaucoup, allant de 6 à 41 paires. Les chromosomes sont relativement petits et la polyploïdie fréquente. Dans ces genres, la polyploïdie provoquée peut être très utile chez les arbres forestiers comme elle l'a déjà été pour la production de nombreux types d'arbres et d'arbustes d'ornement.
L'un des aspects les plus prometteurs de la génétique forestière est la production d'hybrides F1 entre espèces de divers milieux. Il se peut que les espèces de conifères qui ont été isolées pendant des milliers de générations se soient croisées et aient produit des hybrides vigoureux et relativement fertiles, comme l'ont démontré des croisements entre le mélèze d'Europe et le mélèze du Japon (Larix decidua et L. leptolepis) et entre des espèces asiatiques et américaines de pins à 5 feuilles. De même, la vigueur hybride a été prouvée dans des croisements entre les espèces américaines et européennes de Populus et Platanus.
L'utilisation de radiations ionisantes comme agent mutagène a été d'un grand intérêt dans l'amélioration des plantes cultivées, même si les mutations intéressantes sont rares. Mais le long cycle biologique des arbres forestiers et l'espace nécessaire pour expérimenter la vigueur et la qualité des mutants recherchés font que l'amélioration par mutation n'est qu'une méthode longue et chère parmi les méthodes d'amélioration forestière. Peut-être la sélection massale au moment de la germination des graines pourrait-elle être employée pour la sélection en vue de la résistance aux maladies comme cela s'est pratiqué avec tant de succès chez les céréales en traitant les graines en cours de germination avec la toxine de l'agent pathogène. La corrélation récemment découverte entre la sensibilité aux rayons X et la vigueur héréditaire des graines de Zea peut être aussi utile pour sélectionner les produits de ségrégation génétique supérieurs dans les populations hybrides d'arbres forestiers.
Chapitre 1
Tous les types d'organismes, même ceux qui n'ont pas atteint le stade chromosomique, sont caractérisés par deux qualités: la répétition, ou constance génétique et la mutation, ou modification génétique. Par suite de phénomènes de mutation, les organismes sont représentés dans la nature par des populations de génotypes. Depuis l'aube même de la vie, il y a une interaction entre l'hérédité et le milieu, et les conditions du milieu dans lequel vivent les organismes sont susceptibles de se modifier. Comme des populations différentes comprennent des individus qui sont plus ou moins bien adaptés à des milieux spécifiques, la composition génétique des populations se trouve modifiée à la suite de la sélection naturelle. Si les contraintes du milieu sont plus sévères que les possibilités génétiques d'une population ou d'une espèce, la population ou l'espèce disparaît et sera remplacée par d'autres.
La composition génétique d'une population d'organismes plus élevés se modifie en raison de la mutation et de la recombinaison. Dans la mutation, les gènes ou molécules de l'hérédité se modifient de diverses façons souvent en ce qui concerne leur ordre normal sur les chromosomes. Les chromosomes forment des groupes de linkage qui limitent les possibilités de recombinaison mais, à cause du crossing-over, les groupes de linkage sont brisés, augmentant ainsi les possibilités de recombinaison au cours de la reproduction sexuelle.
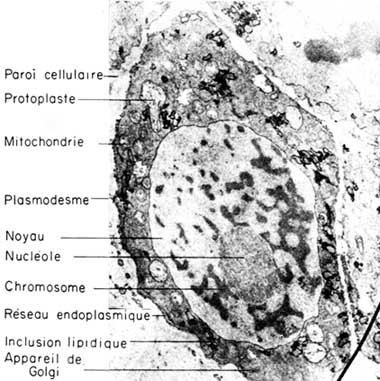
Au cours de la différenciation cellulaire, le matériel héréditaire se concentre sur les chromosomes dans le noyau, tandis que les activités physiologiques et de croissance de la cellule se centrent dans le cytoplasme (figure 1). Cela n'implique pas que le noyau soit inactif du point de vue physiologique, ou qu'il n'y ait pas d'information héréditaire dans le cytoplasme. En fait, les gènes des chromosomes sont l'origine d'instructions biochimiques destinées au cytoplasme et dans ce cytoplasme il y a des plasmagènes (ou plastogènes) qui agissent comme unités héréditaires indépendantes.
LA CYTOLOGIE DES ARBRES FORESTIERS
Ce chapitre traite des arbres forestiers et de leur comportement cytogénétique. D'après l'introduction, il est évident que l'histoire d'une espèce ou d'un genre est d'une importance fondamentale pour sa constitution génétique actuelle, la structure de sa population, son polymorphisme et sa répartition. Il y a aussi des différences fondamentales entre les espèces, les genres, les familles dans leurs caractères cytologiques. Les différences de comportement des chromosomes sont liées:
1. Au nombre chromosomique de base;
2. A la taille des chromosomes et leur structure;
3. Au degré de polyploïdie.
Ces problèmes ont été discutés très en détail par Gustafsson (1960a), qui lui-même se référait au travail de précurseur sur la cytologie des arbres fait par K. et H. J. Sax aux environs de 1930.
Le nombre chromosomique de base
Chez les gymnospermes les nombres de base (x) sont 7 (Ephedra), 8 et 9 (plusieurs genres de Cycadacées), 10 (Sciadopitys), 11, 12 et 13 (dans de nombreux genres), mais ils peuvent aussi atteindre des valeurs plus élevées, comme par exemple 19 et 20 (Podocarpus) et 22 (Pseudolarix). Ces derniers nombres sont sans aucun doute secondaires, c'est-à-dire dérivés, et ils peuvent, dans plusieurs cas, être le résultat d'une sorte de cassure de chromosome ou translocation.
Les genres d'angiospermes ligneux appartiennent à plusieurs familles différentes de la nature. Les différences chromosomiques sont par conséquent complexes et variées. Les nombres de base s'étalent depuis 6 et 7 (Cassia) en passant par 8 (Carpinus), 9 (Hevea), 10 (Cornus), 11 (Corylus et Eucalyptus), 12 (Quercus), 13 (Acer et Ficus), 14 (Alnus et Betula) jusqu'à 19 (Populus), 21 (Platanus), 23 (Fraxinus) et 41 (Tilia).
Les genres de gymnospermes et d'angiospermes diffèrent aussi en ce qui concerne la taille moyenne des chromosomes. Naturellement, cela ne veut pas dire que les angiospermes à petits chromosomes contiennent moins de gènes que les gymnospermes à grands chromosomes; ce fait signifie seulement que le matériel extragénique des chromosomes de gymnospermes a pris un développement extraordinaire. Il semble que des facteurs sélectifs peuvent augmenter ou diminuer la taille des chromosomes suivant le type de milieu, ou comme l'a dit le botaniste danois Raunkiaer, suivant la forme de vie de l'espèce. Voir, par exemple, Babcock (1947), Stebbins (1950) et Gustafsson (1951, 1960a).
Stebbins a souligné que de nombreuses angiospermes ligneuses possèdent des chromosomes plus petits que les genres et les espèces herbacées qui leur sont parents. Cette différence peut dépendre du fait que le bois des angiospermes contient des fibres provenant de cellules initiales du cambium de petite taille ou, suivant Stebbins, de la supposition vraie ou fausse que les végétaux ligneux ont besoin en général d'un système génétique présentant le maximum de recombinaison des gènes, ce système étant favorisé par des chromosomes nombreux et petits. Les principales gymnospermes, les conifères, n'ont pas de fibres de bois et possèdent des cellules initiales du cambium qui sont toutes à peu près de taille égale. Comme on l'a dit plus haut, elles ont un nombre chromosomique de base relativement élevé par comparaison avec les angiospermes, mais que l'on peut considérer comme encore bas, et des chromosomes de grande taille. D'après Stebbins, on peut supposer que la réduction de taille des chromosomes est apparue très tôt au cours de l'évolution des angiospermes ligneuses.
II y a une relation intéressante entre une grande taille des chromosomes et la sensibilité aux radiations, qui a été montrée au cours de ces dernières années, principalement par les travaux de Sparrow et de ses collaborateurs. Sparrow et Miksche (1961) ont montré qu'il existe une bonne corrélation positive entre l'inhibition de la croissance par les radiations ionisantes et la dimension des noyaux de l'interphase dans les méristèmes. L'effet de la taille des chromosomes sur la sensibilité aux rayons chez les espèces forestières a été vérifié par Gustafsson et Simak (1958) et Wettstein et al. (1959) qui ont comparé les effets de la radiation sur les espèces: Populus, Pinus, et Picea. Les membres de la famille des Pinacées ont de gros noyaux comparativement aux angiospermes et le volume nucléaire des pins est approximativement 5 fois celui des chênes (Quercus, spp.) mais les deux genres ont un nombre chromosomique de 2n = 24; cela pourrait expliquer la plus grande radiosensibilité chez les Pinacées.
Depuis que Gustafsson et Simak (1958) ont indiqué la haute sensibilité du pin et de l'épicéa aux radiations, de très nombreux renseignements se sont accumulés. Pedigo (1960, 1962) et Platt (1963) en ont décrit les effets sur Pinus taeda, Sparrow et al. (1963) sur Pinus strobus, Mergen et Stairs (1962) sur Pinus rigida, et Brandenburg et al. (1962) sur Pinus monophylla.
Il est très intéressant d'étudier les effets des radiations de longue durée et de basse intensité sur les pins. Des sujets de Pinus rigida ont été tués après une exposition de 5 ans aux rayons gamma à un taux inférieur à 8 r par jour pendant 8 mois par an. Les individus de Quercus exposés dans les mêmes conditions ont survécu à ces rayons gamma chroniques sans doute à cause de leur volume nucléaire inférieur. Toutefois les descendants R1 des deux genres étaient beaucoup plus variables que ces témoins, ce qui reflétait les effets génétiques accumulés dans les arbres irradiés depuis longtemps. Ceci suggère qu'un nombre appréciable de modifications génétiques et physiologiques peuvent se produire chez les arbres à la suite d'irradiations cumulatives de longue durée et de basse intensité.
Polyploïdie
Peu de genres de gymnospermes contiennent des espèces polyploïdes. Les exemples en sont: Ephedra, Gnetum, et Welwitschia. Juniperus chinensis est tétraploïde et Sequoia sempervirens est une espèce hexaploïde naturelle. Bien que la polyploïdie ne joue pas un rôle significatif dans les modalités de variation des Pinacées, on trouve de temps en temps en pépinière et dans la nature des plants polyploïdes. On a trouvé l'aneuploïdie et la mixoploïdie dans des semis nains de Picea abies (Kiellander, 1950; Illies, 1959) et un semis jumeau d'Abies firma a été décrit comme tétraploïde (Kanezawa, 1949). On cite des cas isolés de polyploïdie naturelle chez Larix. Christiansen (1950) a repéré un Larix decidua adulte tétraploïde, et Chiba et Watanabe ont trouvé chez des plants repiqués de 2 ans de Larix leptolepis 8 semis polyploïdes - 2 avaient des racines diploïdes, tandis que les 6 autres étaient tous tétraploïdes. Un seul individu qui provenait d'un croisement Larix decidua et Larix occidentalis était tétraploïde (Larsen et Westergaad, 1938). On a indiqué des cas de polyploïdie ou de mixoploïdie spontanées dans 4 espèces de pins: Zinnai (1953) a répéré 5 semis tétraploïdes de Pinus densiflora; chez Pinus elliottii, Mergen (1958) a décrit des semis mixoploïdes comprenant des compléments chromosomiques de 2 n, 3 n, 4 n; Johnsson (1959) cite la présence de polyploïdes en Suède chez Pinus sylvestris; et Nishimura (1960) a décrit un semis tétraploïde de Pinus thunbergii qui provenait d'une graine di-embryonique.
A l'aide d'un traitement de colchicine, on a rendu polyploïdes des espèces de gymnospermes (Mergen, 1959). On a provoqué avec succès la polyploïdie chez toutes les espèces sur lesquelles on l'a tentée, et les changements étaient, dans l'ensemble, semblables chez les semis et chez les arbres. En général, les aiguilles se raccourcissaient et s'épaississaient, le nombre de cellules était réduit, les branches étaient plus grosses; la floraison était supprimée, et on observait couramment une panification. Chez la plupart des espèces, les tétraploïdes ne sont pas souhaitables, mais ils sont employés comme intermédiaires dans la production triploïde.
On a essayé d'éviter le passage par le stade sporophytique tétraploïde par un traitement des strobiles à microsporanges chez Larix leptolepis (Illies, 1956) et Pinus nigra et Pinus mugo (Mergen, 1959) au cours de la microsporogénèse. On a produit des grains de pollen diploïdes, mais on ne peut pas obtenir de résultat à partir de la descendance de ce type de pollen. Cette méthode est intéressante pour l'avenir et fera l'objet, sans aucun doute, d'un intérêt accru. D'autres renseignements sont donnés sur la polyploïdie chez les gymnospermes par Gustafsson (1960a), Mehra (1960) et Mergen (1963).
La polyploïdie naturelle apparaît souvent à la suite de l'hybridation entre différentes espèces ou populations se comportant comme des espèces et du doublement du nombre chromosomique qui en résulte, dû à la formation de gamètes n'ayant pas subi la réduction chromosomique (phénomène connu sous le nom d'amphiploïdie ou allopolyploïdie). Un tel type de polyploïdie se rencontre couramment chez les angiospermes cultivées (Nicotiana, Gossypium, Triticum, Brassica), mais aussi chez les espèces sauvages (Galeopsis, Robus, Poa), et on peut produire de nouvelles combinaisons artificiellement comme par exemple Triticale, genre nouveau réunissant Triticum et Secale. Dans de nombreux cas, la polyploïdie est intraspécifique à l'origine (on emploie alors le terme d'autopolyploïdie). Les limites entre l'allo- et l'autopolyploïdie sont peu nettes (Muntzing, 1936). Le genre Dactylis est intéressant à ce point de vue car les tétraploïdes qui se produisent naturellement sont souvent considérés comme des polyploïdes inter-spécifiques, bien qu'il n'y ait aucun doute que les diploïdes correspondants soient proches parents (Muntzing, 1956).
Un cas intéressant a été cité par Wright (1959a) chez le frêne blanc, Fraxinus americana qui se divise en trois «écotypes», l'un du nord (2n = 46), l'autre intermédiaire (présentant la polyploïdie) et le troisième du sud (avec 2n = 46, 92, 138). Le frêne citrouille, Fraxinus tomentosa, est une des rares espèces hexaploïdes (2n = 138), qui dérive probablement d'un croisement entre un frêne vert diploïde et un frêne blanc tétraploïde (Wright, 1959b). Chez Fraxinus, d'après ces études, on rencontre à la fois l'autopolyploïdie et l'amphiploïdie.
Le caractère le plus marquant des races polyploïdes interspécifiques consiste dans les modifications du rythme de développement et du comportement écologique comme l'a d'abord montré Müntzing (1936), et comme l'ont ensuite étudié plus en détail Stebbins (1950, 1956) et Müntzing (1956, 1959). L'aspect «géant», l'accroissement de la vigueur végétative, l'altération des réactions d'incompatibilité et les modifications des besoins écologiques font que les autopolyploïdes sont bien adaptés pour une extension de la zone de culture des diploïdes. Ces polyploïdes sont aussi utiles en matière d'amélioration des arbres forestiers, idée émise par Nilsson-Ehle dans ses premières études sur les réactions et les potentialités du tremble géant autotriploïde, Populus tremula. Chez les gymnospermes ainsi que chez de nombreux feuillus, les allopolyploïdes peuvent finalement convenir encore mieux que les autopolyploïdes pour une utilisation pratique immédiate.
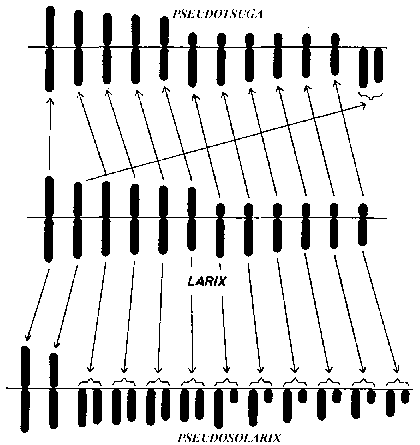
FIGURE 2. - Evolution du caryotype: mutations chromosomiques au cours de l'évolution illustrées par les caryotypes (idiogrammes) de Pseudotsuga, Larix et Pseudolarix.
La disploïdie et la polyploïdie secondaire
Le terme disploïdie (Chiarugi, 1932) signifie qu'on rencontre des nombres de base différents dans un genre, ces nombres ayant une relation phylogénétique. Ce terme, bien que n'étant pas d'emploi courant dans la littérature, est très utile. Dans les genres d'arbres, la disploïdie n'est pas rare non plus. Dans les genres Cycas et Microcycas par exemple, on rencontre les nombres de base 11, 12 et 13 qui dérivent probablement les uns des autres. Chez Podocarpus, avec des nombres de base de 11, 12, 19 et 20 on rencontre probablement une sorte de cassure chromosomique aboutissant à la disploïdie. D'après Barlow (1959), le genre Casuarina d'Australie présente X = 8, 9, 10, 11, 12, 13, etc. Un genre qui présente encore plus d'intérêt pour l'évolutionniste est le genre Pseudolarix, parent de Larix où il semble possible de définir d'une façon plausible ce qui s'est passé phylogénétiquement au moyen de l'analyse chromosomique (fig. 2). Pseudolarix amabilis avec X = 22 a deux paires de chromosomes avec des centromères submédians ou médians et 20 paires avec des centromères presque terminaux. Chez Larix, tous les chromosomes ont des centromères médians, submédians ou subterminaux. Il est fort possible que Pseudolarix dérive de Larix par une cassure de chromosomes dans la région des centromères pour 10 chromosomes du jeu haploïde tandis que deux chromosomes sont restés inchangés (Mergen, 1961).
D'après Barner et Christiansen (1962), il y a chez Pseudotsuga taxifolia 13 paires de chromosomes, les deux plus petits étant télocentriques, c'est-à-dire ayant des centromères terminaux ou presque terminaux. Si l'on considère ces deux chromosomes comme dérivant d'un seul chromosome long à centromère médian ou submédian, le schéma est tout à fait semblable à celui de Larix. Il y a, en outre, d'autres particularités génériques qui font penser à des relations très étroites entre ces deux genres.
Quand les schémas des espèces et des genres de gymnospermes auront été étudiés plus en détail, on pourra tirer une série de conclusions concernant les relations entre les altérations des chromosomes et la phylogénie.
Le noyau et le cytoplasme
Dans l'introduction, on a souligné qu'il y a dans le cytoplasme des éléments cellulaires que l'on peut considérer comme se reproduisant eux-mêmes et en un certain sens indépendants des gènes du noyau. Ce fait est en rapport avec l'opposition qui existe entre les termes de génome (la somme des gènes du noyau, Winkler, 1920) et plasmone (la somme des facteurs héréditaires du cytoplasme, Wettstein, 1926), ainsi que le terme moins clair de plastome (les facteurs héréditaires des plastes, Renner, 1934, Michaelis, 1957-58 et Gustafsson et Wettstein, 1957-58). Du point de vue général, il est intéressant de savoir que la stérilité mâle des espèces hermaphrodites est d'origine cytoplasmique (maïs, canne à sucre, betterave à sucre, oignon et Dactylis), ce qui fait de ce facteur cytoplasmique un outil important pour la production de vigueur hybride ou hétérosis. Il en est de même du problème de la différenciation des sexes pour lequel les facteurs cytoplasmiques jouent un rôle sur la formation des organes sexuels femelles ou mâles (chez Aquilegia, Godetia, Bryonia, Satureja, Cirsium et Streptocarpus). Les études classiques de Correns (1908, 1916) sont fondamentales à ce point de vue. Les conifères sont généralement monoïques, mais chez de nombreuses espèces comme Pinus sylvestris, on trouve des individus qui sont mâles ou femelles de façon prédominante. Une sélection systématique pour la dioïcité peut être très utile pour la production d'hétérosis, ainsi que l'utilisation d'individus mâles stériles dans les croisements. Au contraire, pour produire des lignées autofécondées, le passage de l'état monoïque ou dioïque (par exemple chez Salix ou Populus) à l'hermaphrodisme peut dans certains cas être avantageux. Ces modifications ont été réalisées chez le chanvre (Sisov, 1937; Sengbusch, 1952), en passant de l'état dioïque à l'état monoïque ou à l'hermaphrodisme.
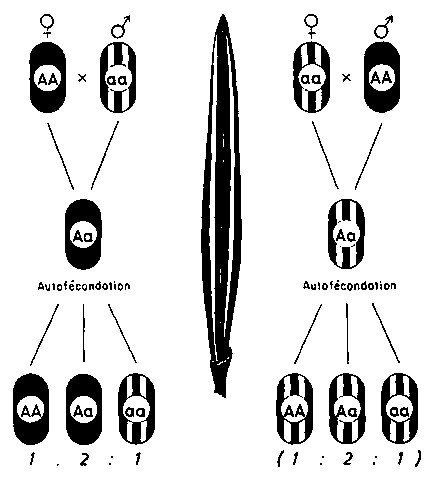
Au cours des dernières années, on a découvert des indices qui montrent que l'hérédité cytoplasmique peut être déclenchée dans une espèce au moyen de mutations induites dans les gènes nucléaires (fig. 3). En ce qui concerne certains gènes léthaux-chlorophylliens, par exemple, des mutations nucléaires provoquent des modifications irréversibles dans les plastes ou plastogènes. Si l'on enlève ensuite les gènes nucléaires, l'absence de plastes persiste et se transmet cytoplasmiquement (par l'intermédiaire de la plante mère) à la descendance (Wettstein, 1961 et études non publiées, Gustafsson, 1960b). Il serait très souhaitable de provoquer aussi ou d'isoler chez les espèces forestières une stérilité mâle ou une dioïcité commandée par le cytoplasme et par les gènes.
En ce qui concerne la résistance aux maladies, quelques données découvertes par Langner (1952) indiquent une origine cytoplasmique dans la sensibilité à la maladie des aiguilles dans les croisements entre Larix decidua et T, leptolepis.
FIGURE 3. - Mutations cytoplasmiques (dans ce cas, dérèglement des plastes) induites par des mutations de gènes.
LA GÉNÉCOLOGIE DES ESPÈCES FORESTIÈRES
La structure de l'espèce et de la population
Dans un certain sens, l'espèce est l'unité fondamentale de l'évolution. Cependant, la définition de ce qui constitue une espèce est différente suivant les points de vue de l'observateur et de l'expérimentateur. Turesson a essayé, en 1922, de faire une division tranchée fondée non pas sur des caractères morphologiques mais sur des caractères biologiques entre et à l'intérieur des espèces ou des groupes d'espèces. Il distingue:
1. Les écotypes qui sont des sous-unités interfertiles;2. Les écoespèces, qui peuvent former des hybrides, mais de fertilité diminuée et de viabilité réduite;
3. Les coenoespèces, qui consistent en une ou plusieurs écoespèces qui peuvent échanger leurs gènes entre elles mais ne peuvent pas se recombiner avec d'autres groupes d'espèces.
Les conceptions de Turesson ont été soutenues plus tard particulièrement par Clausen, Keck et Hiesey dans leurs remarquables études sur la flore de Californie (Clausen, 1951). Cependant, ces notions ont fait l'objet de beaucoup de critiques. Il suffit de savoir qu'en général une espèce est hétérogène et polymorphe, qu'elle se divise en populations adaptées ou en cours d'adaptation à des climats, à des stations ou à des niches variés. Que les populations à l'intérieur d'une espèce soient différentes ou non du point de vue morphologique n'a qu'une importance secondaire; de même, le fait que les populations forment des écotypes plus ou moins nettement délimités ou qu'elles constituent des clines avec des modifications progressives des caractères physiologiques et morphologiques (Huxley, 1938; Langlet, 1959 a, b).
Le degré d'hétérogéneité et l'intensité de la recombinaison des gènes dépendent dans une large mesure du type de fécondation, autofécondation ou fécondation croisée, ou même de la disparition de la fécondation dans les espèces parthénogénétiques et à reproduction végétative. La présence d'autofécondation naturelle a été établie avec certitude pour une série d'espèces forestières, même chez des conifères. C'est le cas de l'espèce très uniforme Picea omorika, d'après Langner (1959). Cependant, même dans ces cas-là, des croisements se produisent avec très souvent une ségrégation très prononcée dans les générations ultérieures. Dans les petites populations, l'autofécondation peut jouer, ayant pour conséquence, de même que dans la nature, une tendance à l'état homozygote et une diminution générale de la viabilité des variante homozygotes due aux effets de la consanguinité. Chez les espèces forestières hermaphrodites ou monoïques, le degré d'autofécondation naturelle peut varier suivant les années, car il dépend de la floraison et de la dispersion du pollen dans le peuplement (pour une étude détaillée de la distance de vol du pollen, voir Andersson, 1955). Il faut prendre sérieusement en considération ce fait quand on utilise la régénération naturelle en sylviculture. On a souligné que chez les espèces à fécondation croisée normale comme Picea abies, Pinus sylvestris et Pinus monticola, l'autofécondation peut se produire très facilement et que l'on rencontre d'ordinaire certains variante pleinement autocompatibles (Sylvén, 1910; Plym Forshell, 1953; Barnes et al., 1962; Eklundh Ehrenberg, 1963). Des biotypes autofertiles surgissent soit spontanément, soit par l'expérimentation sous l'action d'agents mutagènes par mutation-perte ou par destruction des gènes et des allèles d'incompatibilité.
Cependant, chez de nombreuses espèces végétales, y compris les espèces forestières de conifères et d'angiospermes, il existe une série de transitions entre une pleine auto-incompatibilité et une aptitude à l'autofécondation prononcée. Néanmoins, ces espèces, pour des raisons écologiques, historiques et de migration, sont partagées en populations qui se croisent dans la nature et sont plus ou moins adaptées à leurs habitats particuliers, de même qu'elles constituent des éléments des communautés naturelles végétales, avec une compétition et une coopération des biotypes individuels à l'intérieur des espèces et entre espèces.
Une étude des plus intéressantes est celle des effets de l'autofécondation comparée à la fécondation croisée dans quatre sujets de Pinus monticola (Barnes et al., 1962) dont deux étaient autofertiles, et deux en partie autostériles. On a mélangé le pollen des arbres et les mélanges ont été employés pour la fécondation. La proportion de graines résultant de fécondation croisée ou d'autofécondation était variable, et elle dépendait du génotype de l'arbre mère ainsi que du génotype du père utilisé. Dans quelques cas, le pollen de l'arbre lui-même était aussi efficace que le pollen d'un autre arbre; dans la plupart des cas, cependant, il était moins efficace.
Chez les espèces apomictiques, des hybrides fortement hétérozygotes peuvent vraiment se reproduire et se répandre largement dans la nature en raison de leur aptitude à la formation asexuée des graines, ou en raison de leur mode de reproduction végétative. Ces espèces sont plutôt rares parmi les arbres forestiers ayant un intérêt économique, bien qu'on ait rencontré l'apomixie chez Alnus, Euonymus, Sorbus et d'autres genres, et que de nombreuses espèces, comme les trembles, les peupliers et les saules se reproduisent végétativement.
Les populations d'arbres et leur adaptation
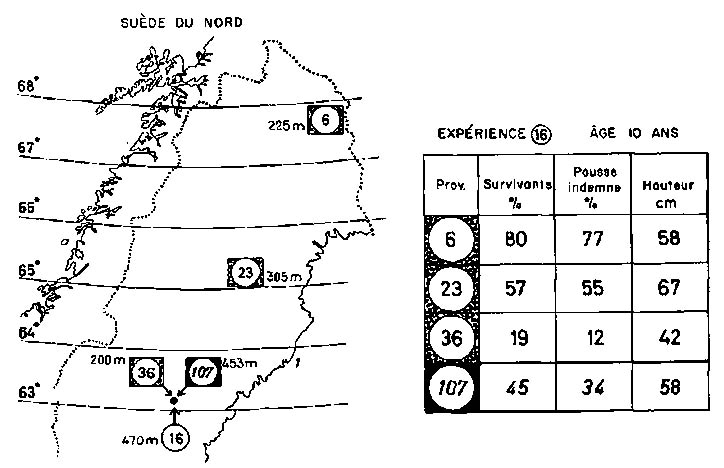
On considère souvent que les populations naturelles sont en général bien adaptées au milieu extérieur. Dans un récent article de Duffield (1962), par exemple on trouve la phrase suivante: «Par conséquent, en tant que procédé d'amélioration, l'induction de mutations est un gros risque à courir, car chez des organismes aussi bien adaptés que le sont la plupart des arbres forestiers, toute modification est virtuellement susceptible d'amener le pire» (passage souligné par les auteurs). Au contraire, une population d'arbres échoue souvent dans la recherche d'une adaptation complète à son habitat. Cela est vrai des aptitudes à la floraison, par exemple chez Picea abies, qui fleurit de façon irrégulière sur une grande portion de son aire de répartition, ou pour la production de graines, qui soit ne s'opère pas du tout, soit est empêchée de diverses façons comme dans le cas de Pinus sylvestris et Picea abies aux latitudes et aux altitudes élevées (Simak et Gustafsson, 1954). Cependant, en ce qui concerne également la survie et la croissance du système végétatif, on observe souvent une résistance imparfaite aux gelées et aux basses températures des populations locales quand elles sont soumises à des conditions sévères. Pour des populations de Scandinavie de Pinus sylvestris (fig. 4), ce fait a été bien démontré par Wibeck (1933) et ensuite développé par Eiche (1962 et documents non publiés; voir aussi Gustafsson, 1962). Il ne faut pas oublier qu'un milieu naturel est en perpétuelle modification et que des populations locales ont à subir des processus d'adaptation successifs aboutissant aussi à des modifications génétiques de la variabilité potentielle. En outre, des raisons historiques comme l'hétérozygotie originale, le mode de migration depuis des conditions extérieures plus ou moins sévères, des attaques irrégulières ou des catastrophes dues à des insectes, à des champignons, des rongeurs, des incendies, etc., peuvent influer beaucoup sur la constitution de la population locale.
Si l'on calcule en unités de temps et peut-être aussi par génération, les arbres forestiers présentent une adaptation lente à des modifications du milieu. Les délais sont rendus encore plus longs par les floraisons médiocres ou l'absence de production de graines. (Dans certains districts de la Suède, par exemple, on n'observe une bonne année de semence pour Pinus sylvestris que tous les 30 ou 40 ans.) En général, dans les populations nordiques de pins et d'épicéas, le stade juvénile jusqu'à 15 ou 20 ans est très sensible à un climat accentué. Quand ils ont dépassé ce stade, le peuplement et les biotypes individuels peuvent être considérés comme adaptés végétativement. Par conséquent, suivant les conditions climatiques au cours des années qui suivent la plantation ou la germination en forêt, les populations locales peuvent être sélectionnées plus ou moins sévèrement pour une localité donnée, impliquant des conséquences à long terme pour les générations à venir.
Les populations d'arbres sont en général mal adaptées à de nombreuses interventions de l'homme telles que les différentes méthodes d'éclaircie et, comme les espèces sauvages, elles ne sont pas suffisamment sélectionnées pour répondre aux exigences de l'homme quant à la qualité du bois, la production de pâte à papier, la composition chimique. Cet état de choses rend l'amélioration des arbres forestiers d'autant plus urgente, au besoin grâce à une domestication poussée des espèces forestières.
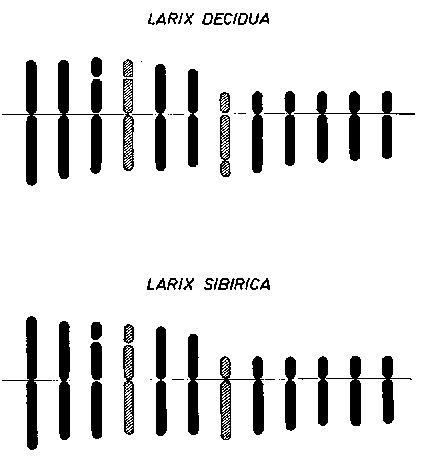
FIGURE 5. - Idiogrammes d'espèces de mélèzes: les espèces apparentées ont souvent des caryotypes (idiogrammes) différents, comme on le voit ici pour Larix decidua et L. sibirica.
La composition génotypique des populations à fécondation croisée
II est de la plus haute importance pour l'amélioration des plantes de connaître la composition génotypique des populations, comment sont répartis les gènes sur les chromosomes, leurs effets sur l'état hétérozygote et homozygote, leurs possibilités de recombinaison libre ou limitée. Les études sur les espèces de Drosophila de Mather (1943, 1960), Dobzhansky (l951) et de leurs collaborateurs ont donné une assez bonne description du comportement et de la constitution des populations, dans des conditions soit naturelles soit artificielles et sous la pression d'une sélection sévère ou faible. L'extrême hétérozygotie des populations naturelles, mettant aussi en jeu la cassure et les dispositions nouvelles des segments de chromosomes, est un fait bien établi chez de nombreuses espèces végétales; de même l'abondance de mutations à l'état hétérozygote, qui, à l'état homozygote, diminuent la viabilité et agissent ensuite comme des gènes léthaux ou semi-léthaux, est bien prouvée. De nombreuses mutations de ce type augmentent la viabilité au-dessus de la normale quand elles sont hétérozygotes, par exemple les gènes léthaux pour la chlorophylle (Gustafsson, 1954). Eiche (1955) a montré que ces gènes léthaux pour la chlorophylle sont communs dans les populations naturelles d'arbres forestiers. De toute façon, il existe un système complexe d'équilibre entre tous les types de gènes et de mutations, qu'ils soient hétérozygotes ou homozygotes. En raison des phénomènes de linkage, la variabilité potentielle est réduite au premier stade de la sélection mais elle s'élargit lorsque se sont produits des crossing-over et des recombinaisons de gènes dans des segments chromosomiques voisins.
Malheureusement, on sait peu de choses sur la présence d'inversions spontanées ou de translocations dans les espèces forestières. Sugihara (1940) a montré que des translocations se produisent dans des populations locales de Cephalotaxus drupacea puisque l'on cite l'existence de quadrivalents et hexavalents au moment de la méïose des biotypes de cette espèce. De petites inversions peuvent se produire dans toutes les espèces sans qu'on les ait remarquées dans les quelques études qui ont été effectuées jusqu'ici sur la méïose. Une étude attentive des schémas des espèces parentes, par exemple pour Larix (fig. 5) ou Pinus chez les conifères, révélerait si la différenciation des espèces apparaît non seulement grâce à la mutation génique, mais aussi grâce à des réarrangements chromosomiques faibles ou importants. Il est évident, d'après les travaux d'hybridation, que l'on peut faire des croisements de grande portée dans de nombreuses espèces et de nombreux genres de conifères. On peut utiliser ce fait pour la production de vigueur hybride (Righter, 1946, 1960; Hyun, 1960). Dans la nature, l'hybridation entre espèces est souvent liée à une bonne part d'introgression spécifique (Andersson, 1949; Stebbins, 1950); c'est-à-dire que les gènes sont transférés d'une espèce à une autre par hybridation et ensuite par croisement de retour entre l'hybride et l'une ou l'autre des espèces parentes. Des cas d'introgression semblable ont été cités pour les espèces d'Alnus, Quercus et Pinus.
Le développement harmonieux de nombreuses populations d'arbres est intimement associé avec des phénomènes de symbiose, par exemple avec la présence de mycorrhizes. Cela aboutit à une interaction mutuelle et à une adaptation continuelle entre groupes d'organismes entièrement différents. En outre, comme l'ont montré de nombreux chercheurs, un peuplement forestier forme souvent, ou même d'une façon générale, une énorme «collectivité vivante» qui est le résultat d'une symbiose des racines de grande envergure, avec des transports d'éléments nutritifs, d'hormones, d'exsudations, d'eau et d'autres matériaux d'un individu à l'autre. Les conséquences biologiques des influences biotiques mutuelles de cette sorte n'ont pas été jusqu'ici considérées sous l'angle de la génétique ou de la biométrie.
LA STRUCTURE GÉNÉTIQUE ESPÈCES FORESTIÈRES, FONDEMENT DE L'AMÉLIORATION
L'hérédité quantitative et ses bases génétiques
Dans le paragraphe précédent, on a mis l'accent sur la complexité des espèces végétales à fécondation croisée. Les populations réagissent à la sélection naturelle et artificielle comme des sortes de systèmes polygéniques, c'est-à-dire que de nombreux gènes, dont la plupart ont des effets individuels légers, au moins à l'état hétérozygote, constituent la base génétique des caractères quantitatifs. La théorie de l'hérédité quantitative vient d'être analysée récemment par Mather dans de nombreux articles (Mather, 1960). Dès 1915, Nilsson-Ehle soulignait l'existence de ces systèmes héréditaires complexes et il écrivait: «Il est évident, d'après les études d'ensemble qui ont été faites jusqu'ici dans différents pays, que les caractères ayant une importance pratique, pourvu qu'ils soient quantitatifs, doivent être considérés en général comme étant constitués par plusieurs facteurs génétiques, et de temps en temps par de nombreux facteurs, qui obéissent aux lois de Mendel... Toutes les propriétés héréditaires, non seulement celles qui sont de nature morphologique externe - non indispensables pour un programme d'amélioration - mais aussi celles de nature physiologique ou biologique, comme la résistance au froid, la résistance génétique aux maladies, la précocité, la résistance à la verse, la capacité de germination, etc., produisent une ségrégation nette à la suite des croisements et forment de nouvelles combinaisons. En général, les propriétés à hérédité quantitative réagissent de cette manière, par exemple, les dimensions et la teneur de protéines dans le blé. L'auteur de cet article n'a pratiquement trouvé dans aucune recherche des caractéristiques importantes ayant un autre comportement... La ségrégation peut être plus ou moins complexe - ce qui suivant notre théorie, dépend du nombre et de l'action des différents facteurs génétiques - mais il est indubitable que les différentes propriétés agissent fondamentalement de la même façon. La composition des caractères ayant une importance pratique, du fait qu'ils dépendent d'un grand nombre de facteurs génétiques, est de la plus grande importance pour les principes et les méthodes de l'amélioration des plantes.» (Op. cit., p. 57 de la traduction.)
Cependant, même si Nilsson-Ehle, East, Fisher, Mather et d'autres auteurs insistent sur l'importance des gènes à effet réduit, il ne faut pas oublier que les plantes cultivées diffèrent de leurs ancêtres sauvages pour une série de qualités fortement et brusquement modifiées (Schwanitz, 1957). Les mutations à effets importants ont contribué de façon continuelle au processus de la domestication. A la suite de ces modifications importantes, la recombinaison suivie de la mutation se met au travail, ajustant entre elles des modifications grandes ou petites pour en faire un tout équilibré, dans lequel de nombreux gènes agissent ensemble sur le même caractère quantitatif. Dans les espèces à fécondation croisée, les gènes nuisibles et les mutations exercent aussi des effets légers de modifications à l'état hétérozygote. La raison pour laquelle il est si difficile d'étudier en hérédité quantitative les modifications dues à un seul gène, chez les arbres forestiers, est assez simple; les dispositifs expérimentaux et les méthodes de mesure sont souvent imprécis et inadéquats et les influences du milieu cachent des effets génétiques légers.
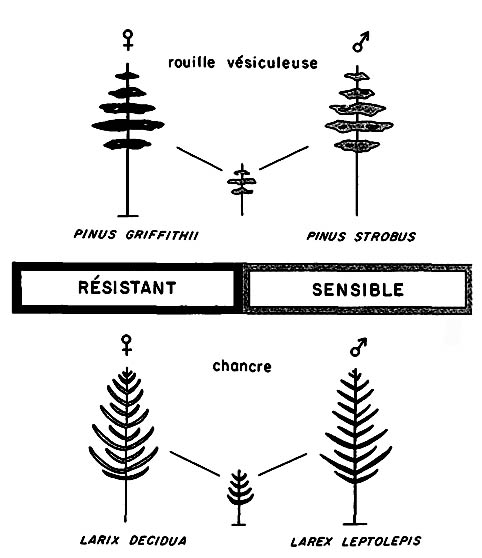
FIGURE 6. - La résistance aux maladies chez les hybrides spécifiques dans le cas des conifères, peut être récessive (en haut) ou dominante (en bas)
La résistance aux maladies et les effets des gènes majeurs
Ce n'est que récemment que la génétique de la résistance des arbres forestiers aux maladies a pris de l'importance. Dans une étude publiée en 1962, Heimburger formulait ce problème de la façon suivante: «Les bases génétiques de la résistance chez l'hôte peuvent être polygéniques ou bien commandées par un petit nombre de gènes majeurs, bien que l'on ait trouvé que la résistance aux maladies chez les végétaux était basée dans la majorité des cas sur une combinaison de gènes majeurs et de polygènes. Les gènes majeurs qui commandent la résistance aux maladies constituent le point de départ de la plupart des progrès spectaculaires effectués dans l'amélioration pour la résistance en agriculture et en horticulture.» D'autre part, Heimburger écrit plus loin: «On doit bien garder à l'esprit le fait que la plupart des plantes cultivées ont été produites par sélection et amélioration, principalement en utilisant les polygènes et que de nombreux caractères nouveaux et utiles, comme la résistance aux maladies, peuvent être ainsi sensiblement amplifiés si l'on ne dispose pas d'autres plants proches parents à résistance supérieure.» (Op. cit., p. 358.) Les données sur la résistance aux maladies chez les arbres forestiers sont vagues et contradictoires. Cela dépend dans une très grande mesure du fait que l'hérédité de la résistance a été étudiée surtout dans les croisements d'espèces, pour lesquels la condition d'hybride complique l'action physiologique des gènes pour la résistance provenant de sources tout à fait différentes. On veut s'attendre à ce que les hybrides d'espèces entre biotypes résistants deviennent souvent sensibles. Heimburger cite lui-même le cas d'hybrides entre Pinus griffithii et Pinus strobus dans lesquels «le mécanisme d'inhibition n'existait plus, l'infection s'étendait rapidement depuis les aiguilles jusqu'au tronc et le résultat était une forte mortalité des jeunes plants.» (Op. cit., p. 360.) D'autre part, on connaît des cas où une espèce rend l'hybride presque totalement résistant, par exemple dans des croisements entre Larix leptolepis, résistant au chancre, et Larix decidua (fig. 6).
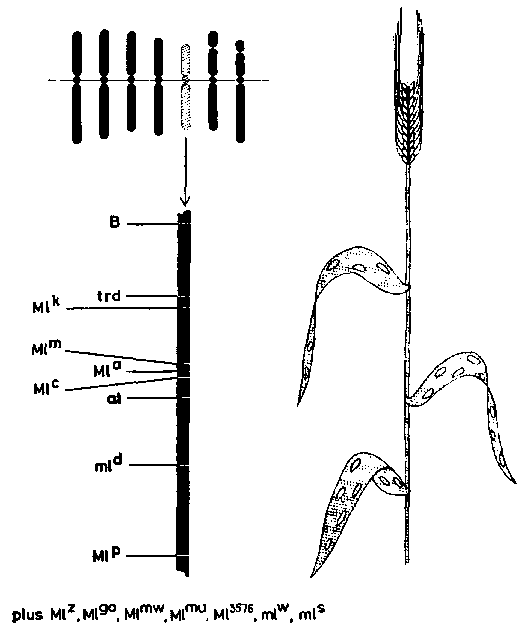
Il existe, provenant d'études sur les plantes agricoles et horticoles, des données abondantes prouvant que des gènes majeurs jouent pour la résistance. Knight (1946) présente une liste de 33 plantes cultivées chez lesquelles on a démontré l'existence d'un gène majeur de la résistance à 84 parasites ou maladies cryptogamiques. En outre, des travaux récents de Briggs, Flor, Favret et d'autres auteurs ont montré qu'il existe dans une espèce de nombreux gènes majeurs conférant la résistance, et qu'ils ne sont pas répartis au hasard dans le génome de la plante-hôte, mais qu'ils ont tendance à être groupés sur des segments génétiques, concentrés sur quelques chromosomes. Briggs fut le premier à suggérer l'hypothèse d'un phénomène de cet ordre dans la maladie de la carie du blé. La génétique de la résistance au mildiou chez l'orge (fig. 7) est probablement l'exemple le plus frappant (Favret, 1960a, b). Dix-huit facteurs différents de la résistance au mildiou sont répartis sur 14 loci; 17 facteurs se trouvent sur le chromosome 5 et un sur le chromosome 4. Treize loci forment un grand «segment isophénique» de 45 à 50 unités de crossing-over. On peut diviser ce segment isophénique en quatre sections suivant la nature des allèles de la résistance. Certains loci ne contiennent qu'un allèle de résistance, d'autres constituent des séries de 2 à 5 allèles. Trois loci sont étroitement linkés, et forment un complexe de gènes sur un segment court d'une longueur d'environ une unité de crossing-over. La plupart des gènes de la résistance sont dominants ou semi-dominants, mais on peut rencontrer aussi des facteurs récessifs.
Il est à retenir que ce système complexe de résistance a été trouvé dans plusieurs espèces étudiées avec soin. Cela peut jouer en faveur de l'hypothèse d'une situation identique chez les résineux et les feuillus. Bien que de nombreux gènes entrent en jeu, chacun d'eux a en général un effet «majeur». Par conséquent, l'hérédité n'est pas «polygénique» comme le dit Heimburger, à la suite de Mather, c'est-à-dire qu'elle présenterait de petits effets additifs, mais elle est «multifactorielle» comme le dit Nilson-Ehle, basée peut-être sur une série de petits doublements, comme l'a suggéré Favret dans ses travaux sur les mutations, de nombreux facteurs de la résistance étant dominants ou semi-dominants. En outre, il faut souligner ici que la résistance aux maladies et la tolérance à une maladie en forêt peuvent mettre en jeu des phénomènes différents.
FIGURE 7. - La résistance à une maladie cryptogamique (ici, le mildiou de l'orge) est souvent déterminée par différents gènes, dominants ou récessifs, qui sont localisés dans des segments particuliers des chromosomes appelés segments isophéniques.
La notion d'héritabilité
En expérimentation forestière, dès qu'il s'agit de caractères quantitatifs, comme la production en volume, l'accroissement en hauteur ou en diamètre, la forme du tronc et les qualités du bois, les expériences devraient être faites de manière à distinguer les influences génétiques de celles dues au milieu et à estimer dans quelle mesure un phénotype donné est déterminé par l'hérédité et dans quelle mesure il l'est par le milieu. En matière de sélection animale ces problèmes sont aussi importants qu'en matière de sélection forestière, en raison de la longueur des générations et du coût élevé des tests de descendance. La notion d'héritabilité (h2) (Lush, 1948) rend compte de la partie de la variance phénotypique totale (VP) due à des facteurs génétiques à effets additifs (VA). A l'origine, le terme «héritabilité» s'appliquait seulement à la corrélation entre parents et descendance. En matière d'amélioration des arbres, on emploie souvent le mot héritabilité suivant deux significations, selon les conceptions de Lush (1948). L'héritabilité au sens large s'applique au fonctionnement du génotype entier pris comme unité et s'emploie par opposition à la variance due au milieu. La définition de l'héritabilité au sens strict ne comprend que les effets moyens des gènes, transmis des parents à leur descendance par l'intermédiaire de la méiose (séparation des chromosomes) et de la fécondation (recombinaison des chromosomes) qui la suit. «Cette signification étroite de l'héritabilité s'emploie quand on fait ressortir quelle est la fraction des différences phénotypiques entre parents et descendance qu'on peut espérer raisonnablement retrouver dans la descendance.» (Lush, Op. cit.) Voir également Toda, 1957; Zobel, 1961; et Eklundh Ehrenberg, 1963.
L'héritabilité au sens large n'est rien d'autre que l'opposition soulignée par Johannsen entre génotype et phénotype. Cette notion est éclairée magnifiquement par les expériences sur la couleur des fleurs et la température chez Primula sinensis décrites par Erwin Baur (1919) dans son manuel classique de génétique (fig. 8). Chez ceux des arbres forestiers où la multiplication végétative représente le processus normal, les études sur les clones mettent facilement en relief quelle est la proportion de la variation due au génotype. C'est là un principe important de la sélection des arbres plus, chez lesquels de nombreuses influences fortuites telles les propriétés du sol, le bilan hydrique, la densité du peuplement, les éclaircies irrégulières, etc., peuvent faire apparaître comme «plus» des arbres forestiers inférieurs du point génétique ou comme «moins» des arbres supérieurs du point de vue génétique. Cependant, la forme du fût, le port des branches et particulièrement leur angle d'insertion, sont souvent gouvernés en grande partie par des facteurs génétiques. Les propriétés de la graine, par exemple celle de Pinus sylvestris, montrent bien l'influence relative de l'hérédité et du milieu (Simak et Gustafsson, 1954). Des caractères tels que la couleur et la taille de la graine se manifestent selon le degré de maturité de la graine et sont facilement influencés par des modifications du milieu. Au contraire le milieu modifie moins la forme de la graine, c'est-à-dire le rapport longueur/largeur, la structure de la surface et la forme de l'aile. Les détails morphologiques de la graine qui sont particuliers à des biotypes donnés, par exemple l'extrémité du micropyle recourbée ou renflée, ou le hile asymétrique et très marqué sont très fixes et indépendants des modifications du climat et du sol. A l'aide des caractères très fixes, il est possible de contrôler avec précision les pins qu'on utilise dans les greffes et les vergers à graines.
En ce qui concerne les qualités du bois, on s'y est beaucoup intéressé depuis quelques années (Zobel, 1961; Ericson, 1960, 1961; Wellwood et Smith, 1962). Chez certaines espèces, la corrélation existant pour le poids spécifique, entre les arbres plus sélectionnés et leurs clones en plantations comparatives, est assez prononcée. Plus la densité théorique est élevée pour un clone de Pinus sylvestris, plus le rendement en pâte et la résistance à la déchirure de la pâte seront élevés. Comme l'ont montré Wellwood et Smith (Op. cit.) pour Pseudotsuga taxifolia et Tsuga heterophylla, il n'y a pas de relation entre les caractères externes d'un arbre plus et les qualités de son bois. Dans un programme d'amélioration visant à augmenter par la génétique le rendement en pâte, il faut étudier à part les qualités du bois.
L'héritabilité au sens strict est une notion utile, avec quelques restrictions. Dans une bonne expérimentation, l'analyse statistique indiquera quelle est l'importance de la composante génétique de la variance, au moins chez les descendances issues de croisements à l'intérieur d'une population. Le problème reste de supprimer les influences non spécifiques du milieu. Si les arbres plus employés dans l'opération d'amélioration sont choisis soigneusement, on empêchera automatiquement les biotypes ayant des qualités héréditaires moins de prendre part à la production de la graine. Ce procédé est négatif, mais n'en reste pas moins important. Cependant, on renforcera l'aspect positif de la sélection si non seulement on sélectionne avec soin dans la nature les arbres plus que l'on utilise, mais si on les éprouve dans des tests de clones et si on les croise ensuite avec des sujets appropriés. Il est intéressant de noter que des plantations comparatives de provenances qui utilisaient un mélange de graines ont révélé un net parallélisme entre le phénotype des peuplements qui participaient au mélange et sa descendance; Petrini (1959) a décrit un parallélisme du même ordre dans un essai de provenance de Pinus sylvestris âgés de 50 ans. Dans ses études sur l'hérédité comparée des arbres plus et des arbres moins, Eklund Ehrenberg (1963) a montré que la descendance d'arbres plus bien sélectionnés présente une production et un développement supérieurs a ceux de la descendance des arbres moins.
FIGURE 8. - Influence du milieu sur le génotype.
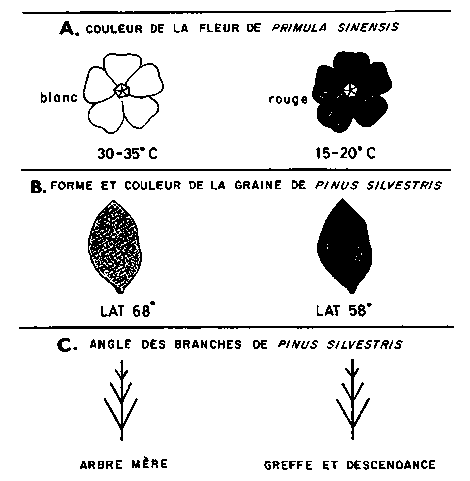
A. Réactions phénotypiques d'un qénotype de Primula à une légère variation de température. B. La forme des graines d'un génotype de Pinus est très indépendante des conditions climatiques, tandis que la couleur des graines est fortement influencée par le milieu. C. Chez Pinus sylvestris l'angle d'insertion des branches a une forte «héritabilité» aussi bien au sens large qu'au sens strict.
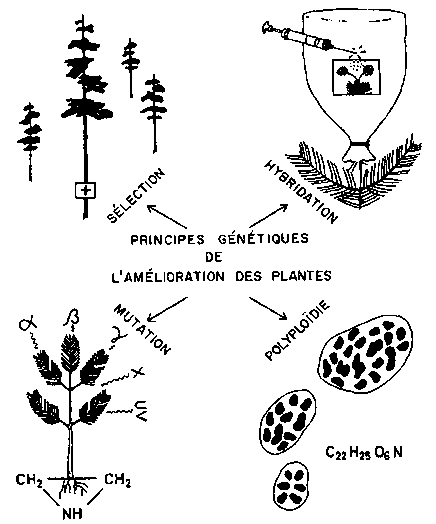
FIGURE 9. - Principes génétiques de l'amélioration des plantes.
Cependant, l'amélioration des arbres ne vise pas à obtenir seulement les effets généraux de la combinaison, mais aussi ses effets spécifiques. La mise en place en vergers à graines de 30, 40 clones ou plus d'arbres plus dans une région donnée est une précaution nécessaire, particulièrement recommandée pour les régions forestières à conditions très difficiles et à changements brutaux. D'autre part, dans les régions de production à conditions climatiques favorables, le nombre de clones pourrait être réduit et, à mesure que l'on acquiert de l'expérience, on pourrait insister davantage sur les aptitudes spécifiques à la combinaison. Dans les vergers à graines qui utilisent la vigueur hybride au moyen de croisements entre espèces et entre provenances, le nombre des clones pourrait être ramené à quelques unités, à la suite de tests de descendance soignés. C'est particulièrement le cas quand on n'a pas recours à la régénération naturelle et que les futurs peuplements se feront par plantation. Les calculs mathématiques sur les relations parents-descendance (l'héritabilité au sens strict) ne seront souvent d'aucun recours si l'on ne tient pas compte des effets spécifiques de la combinaison.
Principes génétiques de l'amélioration des plantes
II est nécessaire de résumer quels sont les facteurs génétiques qui entrent en jeu dans l'amélioration des plantes, tous ces facteurs régissant aussi les phénomènes naturels (fig. 9). Ce sont:
1. La sélection;2. L'hybridation,
a) l'hétérosis en F1, vigueur hybride des croisements d'espèces, de populations et de biotypes,b) la recombinaison et la transgression de F2 en Fn,
c) les croisements de retour des individus de F1 à Fn avec l'un des parents;
3. Les mutations spontanées ou induites par instabilité des molécules, radiations ionisantes ou substances mutagènes chimiques;
4. La polyploïdie naturelle ou artificielle qui se traduit par l'allo- et l'autopolyploïdie.
De nombreux procédés peuvent faciliter les opérations d'amélioration à long terme, tels que la multiplication végétative, l'utilisation de la stérilité mâle, la production de lignées consanguines d'homozygotes que l'on multiplie ensuite en grande quantité chez les arbres forestiers par greffage. L'étude aux rayons X permet de décrire le développement de l'embryon et de l'endosperme. On peut effectuer des séries d'inoculations en serre pour révéler la résistance génétique aux maladies. En soumettant les plantes à des conditions climatiques variées, on facilitera l'étude des relations génotype-phénotype et des conditions physiologiques et écologiques provoquant une floraison précoce et abondante. Les corrélations existant entre le comportement des parents et celui de leur descendance même à un stade précoce simplifient le problème d'une bonne sélection des pieds mères en vue des croisements. Il faut insister sur la nécessité des analyses mathématiques. Dans un proche avenir, de nombreux autres moyens de caractère scientifique ou technique, dont on ne dispose pas encore, accéléreront les recherches et procureront plus rapidement des résultats pratiques.
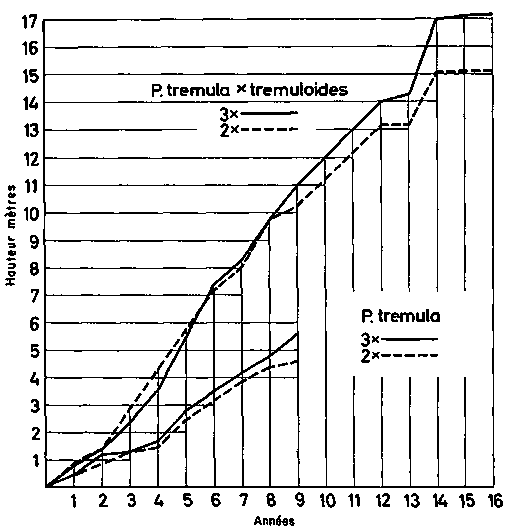
FIGURE 10. - Accroissement en hauteur des hybrides spécifiques et des triploïdes chez Populus, montrant la combinaison des deux méthodes d'amélioration.
LA DOMESTICATION DES ESPÈCES FORESTIÈRES
Un des aspects les plus attrayants de la biologie moderne est la portée générale et l'applicabilité des principes de la génétique dans de nombreuses espèces et de nombreux genres. Cela permet de transposer les résultats acquis sur les bactéries, les algues et les mouches aux populations d'arbres forestiers et à l'homme. L'application de ces résultats peut être limitée par la taille de l'organisme ou la longueur du cycle de reproduction.
II y a quelque chose de fataliste dans la manière qu'a le généticien d'envisager la vie: le matériel héréditaire qui est constitué d'unités fixes ayant un degré de permanence élevé, se transmet inchangé dans sa totalité de génération en génération, mais avec une possibilité de mutation. Cependant le matériel héréditaire n'est pas placé dans le vide et, par le moyen de mutations successives et de recombinaisons, des génotypes ont été sélectionnés et modelés depuis des milliers ou des millions d'années dans les milieux les plus variés. Autrement dit, on ne peut donc concevoir un génotype sans le milieu ambiant.
En sylviculture pratique, on peut voir un exemple de cette interaction du génotype et du milieu. Grâce aux éclaircies, aux élagages, et aux autres opérations culturales, la valeur économique d'un peuplement peut être accrue, mais aucun forestier ne peut produire un peuplement plus à partir d'un peuplement qui végète à cause de son origine mal choisie. Il est impossible d'appliquer les méthodes de l'amélioration des plantes et de la génétique aux arbres forestiers si l'on ne tient pas compte des autres disciplines du métier forestier. A cet égard, il y a une différence entre la sylviculture et l'agriculture.
Des modifications nettes se sont produites chez les plantes agricoles et horticoles au cours de leur développement à partir de l'état sauvage originel. Ces différences font entrer en jeu des processus extérieurs morphologiques et physiologiques qui aboutissent à l'augmentation de la productivité et à des variations dans le caryotype sous-jacent. Les modifications du caryotype font entrer en jeu le nombre et la structure des chromosomes et la disposition des gènes dans les chromosomes. Un exemple typique est fourni par la comparaison des espèces diploïdes sauvages de Triticum et d'Aegilops avec l'espèce de blé commerciale hexaploïde Triticum aestivum. Même à l'intérieur du groupe des plantes agricoles diploïdes, la modernisation est remarquable; il n'est que de comparer le maïs hybride de la ceinture du maïs des Etats-Unis avec les spécimens primitifs de maïs qu'on a découvert dans des cavernes du Nouveau-Mexique ou d'Amérique du Sud (Mangelsdorf, 1958). Il y a, en outre, des espèces qui ne sont devenues d'usage agricole que tout récemment. C'est le cas des lupins, qui grâce aux habiles méthodes de Sengbusch ont été transformés en cultivars sans alcaloïdes à la fin de la période 1920-30 et par la suite ont été améliorés à l'aide de mutations et de recombinaisons.
Chez quelques arbres forestiers, une ère semblable de domestication a commencé et a débuté par la découverte des trembles triploïdes à croissance rapide par Nilsson-Ehle (1936). En croisant des tétraploïdes avec des triploïdes, il est possible d'obtenir des triploïdes en grande quantité. Une amélioration supplémentaire a été obtenue par Johnsson (1953 et documents non publiés) qui a pu montrer (cf. Gustafsson, 1960 b) que les hybrides entre Populus tremula et Populus tremuloides ont en Suède une meilleure production que le Populus tremula indigène (fig. 10). En introduisant un Populus tremula tétraploïde dans les croisements, Johnsson a pu en augmenter encore le rendement et en même temps transmettre la résistance ou au moins la tolérance à Valsa nivea du tétraploïde à l'hybride. D'autre part, de nombreux cultivars de Populus sont des clones hautement sélectionnés au niveau diploïde et mettent aussi en jeu des croisements entre espèces variées. Cependant, on a l'impression que jusqu'ici l'hybridation chez les peupliers a été menée dans une large mesure au hasard, et qu'un programme soigneusement établi de croisements aboutira à des résultats encore meilleurs.
Chez les conifères, comme chez les feuillus et les plantes agricoles, la domestication causera une modification de grande envergure de la structure des populations. Chez les genres Pinus et Larix, il est facile de produire des hybrides. Beaucoup d'entre eux allient la vigueur hybride à la résistance ou à la tolérance aux maladies. On ne sait rien sur les hybrides triploïdes chez Pinus, mais il est possible qu'un certain nombre d'entre eux prennent de la valeur dans l'avenir. Au niveau intraspécifique, les croisements entre populations auront probablement plus rapidement une importance pratique. Par exemple, les provenances de Picea abies venant d'Europe centrale, de Pologne ou de l'ouest de la Russie transportées en Scandinavie, sont souvent plus productives que les populations scandinaves indigènes. On a importé en grande quantité des graines et des plants de ces provenances dans les pays scandinaves et des arbres plus sélectionnés parmi des origines indigènes ou étrangères font déjà partie des vergers à graines mis en place en Suède (Andersson et Andersson, 1962). On a réalisé de larges fécondations croisées dans des vergers à graines de clones expérimentaux qui modifieront radicalement la structure génétique des populations cultivées que l'on utilisera en F 1 sous forme de graine. Les peuplements peuvent être très hétérogènes mais seront à productivité élevée. Dans ces peuplements, on peut continuer la sélection de nouveaux arbres plus.
Le degré de domestication chez les arbres forestiers sera naturellement plus faible que chez les cultures annuelles ou bisannuelles et variera suivant les espèces et suivant les caractères soumis à la sélection. Le produit final doit varier avec les besoins d'utilisation et avec les conditions écologiques de la station où il pousse. Les populations mises à la disposition des praticiens par les sélectionneurs de plantes doivent être parfaitement adaptées aux climats et aux stations dans lesquels on projette de les utiliser. Cela implique une parfaite conformité entre la somme des génotypes de la population et le milieu, principe fondamental que l'on retrouve au commencement et à la fin de ce chapitre.
Bibliographie
ANDERSSON, E. 1949. Introgressive hybridization. New York. 109 p.
ANDERSSON, E. 1955. Pollenspridning och avståndsisolering av skogsfröplantager. Norrlands Skogsv. Förb. Tidskr., p. 35-100.
ANDERSSON, E. 1955. Pollenverbreitung und Abstandsisolierung von Forstsamenplantagen. Z. Forstgenet., 4: 150-152.
ANDERSSON, E. & ANDERSSON, S. 1962. Förteckning över Domänverkets och Skogevårdsstyrelsernas skogsfröplantager. Stockholm, Samarbetsnämnden för skoglig växtförädling och genetik. Distribution restreinte. 158 p.
BABCOCK, E. B. 1947. The genus Crepis. Part One. The taxonomy, phylogeny, distribution and evolution of Crepis. Berkeley, University of California Press. 197 p.
BARLOW, B. A. 1959. Chromosome numbers in the Casuarinaceae. Aust. J. Bot., 7 (3): 230-237.
BARNER, H. & CHRISTIANSEN, H. 1962. The formation of pollen, the pollination mechanism, and the determination of the most favourable time for controlled pollination in Pesudotsuga menziesii. Silvae Genet., 11: 89-102.
BARNES, B. V., BINGHAM, R. T., & SQUILLACE, A. E. 1962. Selective fertilization in Pinus monticola Dougl. II. Results of additional tests. Silvae Genet., 11: 103-111.
BAUR, E. 1919. Einführung in die experimentelle Vererbungslehre. Berlin. 410 p.
BRANDENBURG, M. K., MILLS, H. L., RICKARD, W. K. & SHIELDS, L. M. 1962. Effects of acute gamma radiation on growth and morphology in Pinus monophylla Torr. and Frem. (Pinion pine). Radiation Botany, 2; 251-264.
CHIARUGI, A. 1962. La cariologia nelle sue applicazioni a problemi di botanica. Atti della Società Italiana Progresso delle Science, XXI Riunione Roma, 1932, III: 1-38.
CHIBA, S. & WATANABE, M. 1952. Tetraploids of Larix kaempferi appeared in the nurseries. J. Jap. For. Soc., 34: 276-279.
CHRISTIANSEN, H. 1950. A tetraploid Larix decidua Miller. Det Kgl. Danske Videnskap. Selsk., Biol. Meddel., 18 (9). 9 p.
CLAUSEN, J. 1951. Stages in the evolution of plant species. New York. 206 p.
CORRENS, C. 1908. Die Rolle der männlichen Keimzellen bei der Geschlechtsbestimmung der gynodioecischen Pflanzen. Ber. dtsch. bot. Ges., 26: 686-701.
CORRENS, C. 1916. Untersuchungen über Geschlechtsbestimmung bei Distelarten. Sitz. Ber. Kön. Preuss. Akad. Wiss., 20: 448-477.
DOBZHANSKI, TH. 1951. Genetics and the origin of species. 3rd ed. New York, Columbia University Press. 364 p.
DUFFIELD, J. W. 1962. Forest tree improvement: old techniques and the new science of genetics. H. R. MacMillan Lectureship, University of British Columbia, Columbia, 12. 16 p.
EICHE, V. 1955. Spontaneous chlorophyll mutations in Scots pine (Pinus silvestris L.). Medd. Statens skogsforskn. Inst. Stockh., 45: 1-69.
EICHE, V. 1962. Nya aspekter på plantavgång och bristande vinterhärdighet i norrländska tallkulturer. Skogen: 423-426, 432.
EKLUNDH EHRENBERG, C. 1963. Genetic variation in progeny tests of Scots pine (Pinus silvestris L.). Studia Forestalia Suecica, 10: 1.-135.
ERICSON, B. 1960. Studies of the genetical wood density variation in Scots pine and Norway spruce. Statens skogsforskn. Inst. Stockh., Avdel. skogsproduktion, Rappt. 4, 1-52.
ERICSON, B. 1961. Skogsträdsförädling med sikte på ökat massautbyte. Tek. Vetenskaplig Forskning, 32: 194-203. (Statens skogsforskn. Inst. Ser. Uppsatser 81).
FAVRET, E. A. 1960a. Spontaneous and induced mutations of barley for the reaction to mildew. Hereditas, 46: 20-28.
FAVRET, E. A. 1960b. Induced mutations for resistance to diseases. Genet. Agr., 13: 1-26.
GUSTAFSSON, Å. 1951. Some aspects on variation and evolution in plants. Evolution, 5: 181-184.
GUSTAFSSON, Å. 1954. Mutations, viability and population structure. Acta Agr. Scand., 4: 601-632.
GUSTAFSSON, Å. 1960a. Polyploidy and mutagenesis in forest-tree breeding. Proc. 5th World For. Congr., Seattle, U.S.A., p. 793-805.
GUSTAFSSON, Å. 1960b. Chemical mitagenesis in higher plants. Chemische Mutagenese Erwin-Baur-Gedächtnisvorlesungen I (1959). Abhandl. Deut. Akad. Wiss. Berlin, Kl. Med 1, 14-29.
GUSTAFSSON, Å. 1962. Genetick och växtförädling i skogsbrukets tjänst. Svenska Skogsv. Fören Tidskr., 111-150.
GUSTAFSSON, Å. & SIMAK, M. 1958. Effect of X- and g -rays on conifer seed. Medd. Statens. skogsforskn. Inst. Stockh., 48 (5): 120.
GUSTAFSSON, Å. & WETTSTEIN, D. von. 1957-58. Mutationen und Mutationszüchtung. Handb. Pflanzenzücht., 1: 612-699.
HEIMBURGER, C. 1962. Breeding for disease resistance in forest trees. For. Chron., 38: 356-362.
HUXLEY, J. 1938. Clines: an auxiliary taxonomic principle. Nature, Lond., 142: 219-220.
HYUN, S. K. 1960. Mass production of control-pollinated seed of conifers. Proc. 5th World For. Congr., Seattle, U.S.A., p. 787-792.
ILLIES, Z. M. 1952. Colchicineversuche on Larix decidua Miller und Picea abies (L). Karst. Z. Forstgenet., 1: 36-39.
ILLIES, Z. M. 1956. Veränderungen der Pollengrösse bei Lärche nach Blütenbehandlung mit Colchicin. E. Forstgenet., 5: 112-115
ILLIES, Z. M. 1959. Polysomatie im Meristem von Einzelbaumabsaaten bei Picea abies. Silvae Genet., 7: 94-97.
JOHNSSON, H. 1953. Hybridaspens ungdomsutveckling och ett fõrsök till framtidsprognos. Svenska Skogsv. Fören. Tidskr., p. 73-96.
JOHNSSON, H. 1959. Föreningen för Växtförädling av Skogsträd 1936-1958. Norrtelje Tidn. Boktr. AB. 68 p.
KANEZAWA, R. 1949. Polyploids appeared in twin seedlings of forest trees. J. Jap. For. Soc., 31: 22-24.
KIELLANDER, C. L. 1950. Polyploidy in Picea abies. Hereditas, 36: 513-516.
KNIGHT, R. L. 1946. The role of major genes in the evolution of economic characters. J. Genet., 48: 370-387.
LANGLET, O. 1959a. Norrlandstallens praktiska och systematiska avgränsning. Svenska Skogsv. Fören. Tidskr., 1959: 425-436.
LANGLET, O. 1959b. A cline or not a cline - a question of Scots pine. Silvae Genet., 8: 13-22.
LANGNER, W. 1952. Kreuzungsversuche mit Larix europaea D.C. und Larix leptolepis Gord. Z. Forstgenet., 1: 40-56.
LANGNER, W. 1959. Selbstfertilität und Insucht bei Picea omorika (Pancic) Purkyne. Silvae Genet., 8: 84-93.
LARSEN, C. S. & WESTERGAARD, M. 1938. Contributions to the cytogenetics of forest trees. I. A triploid hybrid between Larix decidua Miller and Larix occidentalis. Nutt. J. Genet., 36: 523-530.
LUSH, J. L. 1948. Heritability of quantitative characters in farm animals. Proc. 8th Intern. Congr. Genet., p. 356-375.
MANGELSDORF, P. C. 1958. Reconstructing the ancestor of corn. Proc. Am. Phil. Soc., 102: 454-463.
MATHER, K. 1943. Polygenic inheritance and natural selection. Biol. Rev., 18: 32-64.
MATHER, K. 1960. Evolution in polygenic system. Accad. Naz. dei Lincei. Evoluzione e Genetica 1960 (Colloquio Internazionale, Roma, 1959), p. 131-152.
MEHRA, P. N. 1960. Cytology and breeding of conifers. Proc. 5th World For. Congr., Seattle, U.S.A., II: 728-733.
MERGEN, F. 1958. Natural polyploidy in slash pine. For. Sci., 4: 283-295.
MERGEN, F. 1959. Colchicine-induced polyploidy in pines. J. For., 57: 180-190.
MERGEN, F. 1961. The chromosomes of Pseudolarix amabilis. Cytologia, 23: 213-216.
MERGEN, F. 1963. Evaluation of spontaneous, chemical, and radiation-induced mutations in the Pinaceae. FAO/FORGEN 63-1/1.
MERGEN, F. & STAIRS, G. R. 1962. Low level chronic gamma irradiation of Pitch-Pine-Oak forest - its physiological and genetical effects on sexual reproduction. Radiation Botany, 2: 206-216.
MERICLE, L. W., MERICLE, R. P. & SPARROW, A. H. 1962. Cumulative radiation-damage in oak trees. Radiation Botany, 2: 265-272.
MICHAELIS, P. 1957-58. Plasma-Vererbung. Hand. Pflanzensücht., I: 140-175.
MÜNTZING, A. 1936. The evolutionary significance of autopolyploidy. Hereditas, 21: 263-378.
MÜNTZING, A. 1956. Chromosomes in relation to species differentiation and plant breeding. Wageningen, Conference on Chromosomes, p. 1 -37.
MÜNTZING, A. 1959. Darwin's views on variation under domestication in the light of present-day knowledge. Proc. Am. Phil. Soc., 103 (2): 190-220.
NILSSON-EHLE, H. 1915. Den modärna ärftlighetsläran och dess betydelse för växtodlingen. Stockholm, Nordiska Bokhandeln. 83 p.
NILSSON-EHLE, H. 1936. Über eine in der Natur gefundene Gigantsform van Populus tremula. Hereditas, 21: 379-382.
NISHIMURA, S. 1960. Chromosome numbers of poly-embryony seedlings of Pinus thunbergii Parl. J. Jap. For. Soc., 42: 263-264.
PEDIGO, R. A. 1960. The effects of ionizing radiation on the ecology of Pinus taeda. Bull. Ecol. Soc. Am., 41: 94.
PEDIGO, R. A. 1962. The effects of ionizing radiation on the ecology of Pinus taeda L. Thèse, Emory Univ.
PETRINI, S. 1959. De två äldsta svenska tallproveniensförsöken. Medd. Statens skogsforsk. Inst. Stock., 48 (11): 1-49.
PLATT, R. B. 1963. Ecological effects of ionizing radiation on organisms, communities, and ecosystems. Proc. lst Nat. Symp. Radioecol. AIBS and Reinhold Publ. (Sous presse.)
PLYM FORSHELL, C. 1953. Kottens och fröets utbilding efter själv- och krosbefruktning hos tall (Pinus silvestris). Medd. Statens skogsforskn. Inst. Stockh., 43 (10): 1-42.
RENNER, O. 1934. Die pflanzlichen Plastiden als selbstständige Elements der genetischen Konstitution. Ber. Math. - Phys. Kl. Sächs. Akad. Wiss. (Leipzig), 86: 241-266.
RIGHTER, F. I. 1946. New perspectives in forest tree breeding. Science, 104: 1-3.
RIGHTER, F. I. 1960. Forest tree improvement through inbreeding and interspecific hybridization. Proc. 5th World For. Congr., Seattle, U.S.A., p. 783-787.
SCHWANITZ, F. 1957. Die Entstehung der Kulturpflanzen. Berlin. 151 p.
SENGBUSCH, R. VON. 1952. Ein weiterer Beitrag zur Vererbung des Geschlechts bei Hanf als Grundlage für die Züchtung eines monözischen Hanfes. Z. Pflanzenzücht, 31: 319-338.
SIMAK, M. & GUSTAFSSON, Å. 1954. Fröbeskaffenheten hos moderträd och ympar av tall. Medd. Statens skogsforskn. Inst. Stockh., 44: 2, 1-73.
SISOV, L. A. 1937. Breeding of hemp. Dans Vavilov, N. I. Handbook of plant breeding. Leningrad, 1937. (Cité dans R. von Sengbusch, 1952.)
SPARROW, A. H. & MIKSCHE, J. P. 1961. Correlation of nuclear volume and DNA content with higher plant tolerance to chronic radiation. Science, 134: 282-283.
SPARROW, A. H. et al. 1963. The radiosensitivity of gymnosperms. I. The effect of dormancy on the response of Pinus strobus seedlings to acute gamma irradiation. Radiation Botany. (Sous presse.)
STEBBINS, G. L. 1950. Variation and evolution in plants. New York, Columbia University Press. 643 p.
STEBBINS, G. L. 1956. Artificial polyploidy as a tool in plant breeding. Dans Genetics in plant breeding, Brookhaven Symp. Biol. N° 9, p. 37-52.
SUGIHARA, Y. 1940. On the multipartite chromosome-ring in Cephalotaxus drupacea Sieb. et Zucc. Sci. Rep. of Tohoku Imp. Univ. Sendai, 4: 13-18.
SYLVÉN, N. 1910. Om pollineringsförsök med tall och gran. Medd. Statens skagsförsöksanstalt, 7: 219-228.
TODA, R. 1957. Variation and heritability of some quantitative characters in Cryptomeria. Silvae Genet., 7: 87 -93.
TURESSON, G. 1922. The genotypical response of the plant species to the habitat. Hereditas, 3: 211-350.
WELLWOOD, R. W. & SMITH, J. G. H. 1962. Variation in some important qualities of wood from young Douglas fir and hemlock trees. Res. Publ. Faculty of Forestry, Univ. Brit. Columbia, N° 50, p. 1-15.
WETTSTEIN, D. VON. 1961. Nuclear and cytoplasmic factors in development of chloroplast structure and function. Can. J. Bot., 39: 1537-1545.
WETTSTEIN, D. VON, GUSTAFSSON, Å. & EHRENBERG, L. 1959. Mutationsforschung und Züchtung. Arbeitsgemeinschaft für Forschung Landes Nordrhein-Westfalen, 73: 7-48, 59-60.
WETTSTEIN, F. VON. 1926. Über plasmatische Vererbung, sowie Plasma- und Genwirkung. Nachr. Ges. Wiss. Göttingen. Math.-Phys. Kl., p. 250-281.
WIBECK, E. 1933. Till skogens rasfrågå på lokala kallorter. Norrlands Skogvs. Förb. Tidskr., p. 227-250.
WINKLER, H. 1920. Vererbung und Ursache der Parthenogenese im Pflanzen- und Tierreich. Jena. 166 p.
WRIGHT, J. W. 1959a. Silvical characteristics of White Ash (Fraxinus americana). Northeast. For. Expt. Sta., Upper Darby, Pa. Sta. Paper N° 123, p. 1-19.
WRIGHT, J. W. 1959b. Silvical characteristics of Green Ash (Fraxinus pennsylvanica). Northeast For. Expt. Sta., Upper Darby, Pa. Sta. Paper N° 126, p. 1-18.
ZINNAI, I. 1953. The morphological characters and the fertillity of the pollen of a tetraploid Japanese red pine induced by the colchicine method. J. Jap. For. Soc., 35: 245-248.
ZOBEL, B. 1961. Inheritance of wood properties in conifers. Silvae Genet., 10: 65-70.
![]()
![]()
![]()
{kind=link}
{kind=link}