![]()
![]()
![]()
J. J. HAVELJ. J. HAVEL est Directeur de l'Ecole forestière de Bulolo, Nouvelle-Guinée.
ON DIT, non sans raison, que la forêt tropicale humide est le paradis du botaniste et le cauchemar du forestier. La surabondance des espèces pose des problèmes pour l'évaluation des ressources, l'utilisation initiale de la forêt et, par la suite, sa régénération et sa gestion. L'évaluation d'une simple parcelle devient un exercice de haute taxonomie; cependant, l'évaluation volumétrique, sans détermination des espèces, ne permet pas de prévoir quelle pourrait être la production. La récolte et la transformation d'arbres d'espèces aussi variées, et particulièrement leur commercialisation, présentent des difficultés quasi insurmontables, de sorte que, généralement, seule une proportion minime de la forêt est exploitée. La régénération des forêts complètement exploitées pose souvent un problème insoluble qui amène fréquemment les forestiers à effectuer des coupes à blanc pour replanter des espèces exotiques.
L'état souvent peu satisfaisant de la forêt dans les pays tropicaux peut, en partie au moins, être attribué à l'incapacité dans laquelle se trouvent les forestiers de déterminer de façon sûre ne serait-ce qu'une faible partie des espèces qui la composent. La plupart des forestiers de métier formés dans les pays tempérés, ne voient pour la première fois les forêts tropicales humides qu'après leurs études. Pour aggraver les choses, il n'existe généralement pas, dans les pays où ils travaillent d'ouvrager complets sur la flore. L'emploi des noms locaux et le recours aux indigènes pour repérer les arbres ne vont pas sans difficultés. Dans bien des régions, particulièrement en Nouvelle-Guinée, la diversité des dialectes est telle que l'établissement d'une équivalence entre les noms scientifiques et ceux qui sont employés par chaque tribu est aussi difficile que la détermination proprement dite des espèces. De plus, le système de classification scientifique ne correspond pas toujours aux notions des indigènes. Lorsqu'il s'agit d'arbres qui ont pour ceux-ci une importance économique, des variétés à peine distinctes les unes des autres portent des noms différents, alors que des arbres d'espèces et même de genres différents, mais dépourvus de valeur économique, sont groupés sous un seul nom commun, tel que le medang en Malaisie.
Mais c'est la complexité même du problème qui constitue la principale difficulté.
Contrairement aux forêts tempérées ou subtropicales les forêts tropicales humides sont extrêmement riches à la fois en espèces et en familles. Sauf dans certaines conditions climatiques et édaphiques particulières, aucune espèce ne présente d'importance capitale du point de vue écologique ou économique. En Malaisie, Menon (4) a estimé que le nombre d'espèces d'arbres se situait entre 2 000 et 2 500; pour la Nouvelle-Guinée, le chiffre serait encore supérieur. Ces arbres appartiennent à une gamme de familles très étendue, y compris celles qui, dans les pays tempérés, se composent pour une large part de plantes herbacées, par exemple, les Datiscacées.
Il en résulte qu'aucune caractéristique, si ce n'est la structure florale, n'est suffisamment diversifiée pour fournir un moyen de détermination suffisant. Menon, qui a étudié la situation en Malaisie, a conclu que les tissus ligneux sont trop simples pour pousser la détermination jusqu'au stade des espèces et qu'en de nombreux cas il est même impossible de distinguer des genres voisins. Endert (1) a mis au point, pour déterminer les genres d'arbres de l'Indonésie, un moyen fondé sur l'observation des seuls organes végétatifs mais cette méthode nécessite un examen morphologique et anatomique des rameaux si détaillé que seuls les botanistes professionnels peuvent l'utiliser.
Bien que l'examen de l'écorce ait été envisagé récemmant par quelques botanistes (7, 8, 6) pour déterminer les espèces, il ne permet pas d'identifier de façon exacte et sûre toutes les essences.
Il est évident qu'un forestier, pour lequel la nomenclature botanique n'est qu'un des nombreux aspects de son travail, ne peut espérer connaître toutes les espèces de la forêt humide à laquelle il se consacre; paradoxalement, il semble cependant que les forestiers préfèrent se reférer à des espèces bien définies. Il y a à cela de multiples raisons. Nombre de caractéristiques dont ils se servent pour identifier les espèces, telles que l'écorce extérieure ou la forme d'un arbre, sont souvent propres à des espèces particulières. Il est souvent plus facile de reconnaître une espèce particulière que d'avoir une idée du genre tout entier. Deux espèces du même genre présentent souvent de fortes différences du point de vue économique.
En revanche, et pour des raisons tout aussi valables, les botanistes préfèrent étudier les familles. Pour beaucoup de familles communes, comme les Lauracées et les Méliacées, la classification et la nomenclature sont jusqu'à présent loin d'être complètes et il est difficile, sinon impossible, d'en déterminer de façon sûre les espèces.
Il semble également que forestiers et botanistes ne soient pas d'accord quant aux moyens de détermination. Les premiers évitent généralement de tenir compte des organes reproducteurs, arguant que les fleurs et les fruits des grands arbres forestiers sont pratiquement inaccessibles et, dans bien des cas, saisonniers. Ils préfèrent s'en tenir à l'aspect général des arbres et à la structure de leur écorce et de leur bois, faciles à observer du sol, en y ajoutant l'examen des caractéristiques des feuilles tombées. Les botanistes, eux, se refèrent aux caractéristiques qui peuvent être observées sur des échantillons d'herbiers portant les organes reproducteurs - feuilles, fleurs et fruits. Il leur arrive souvent de ne pouvoir reconnaître sur le terrain une espèce qu'ils ont, à maintes reprises, manipulée dans l'herbier. Il ne m'appartient pas de décider qui a tort: ni les uns ni les autres n'ont raison. Fleurs et fruits tombés sont des éléments d'observation beaucoup plus utiles, fréquents et sûrs que ne l'admettent les forestiers. Mais d'un autre côté, une fois qu'a été établie une corrélation sûre entre d'une part les organes reproducteurs et d'autre part l'aspect de l'écorce et du bois, les objections qu'opposent les botanistes à l'utilisation des éléments végétatifs perdent de leur valeur.
Il n'en demeure pas moins que la classification et la nomenclature scientifiques se fondent sur l'observation des organes reproducteurs et qu'une première détermination valable n'est possible que sur cette base. En d'autres termes, c'est le botaniste qui montre la voie au forestier. Après une détermination initiale positive, les identifications ultérieures fondées sur la corrélation entre organes reproducteurs et caractéristiques végétatives sont non seulement possibles mais beaucoup plus efficaces dans la pratique.
L'une des raisons qui font que l'on n'a pas plus souvent recours à ces corrélations est qu'elles ne sont pas toujours simples à établir. Il peut arriver que des genres très voisins du point de vue de la structure florale aient en revanche une structure ligneuse très différente, ou au contraire que l'on observe les mêmes caractéristiques anatomiques sur des genres d'arbres autrement très éloignés (4). En fait, ce problème n'est pas étranger aux taxonomistes: la détermination et la classification ne sont pas fondées sur une seule caractéristique et ne sauraient l'être. Elles reposent au contraire sur un ensemble de caractéristiques de complexité variée.
Le nombre de caractéristiques à observer est d'autant plus réduit qu'elles sont moins communes et plus frappantes. Lorsque l'on passe de l'espèce au genre et du genre à la famille, la combinaison des caractéristiques devient de plus en plus importante, mais également plus difficile à saisir, surtout du fait que l'on va vers des concepts plus larges, plus généraux et par conséquent moins clairement définis.
Nous arrivons ainsi au nœud du problème: les espèces qui composent la forêt tropicale sont trop nombreuses pour qu'on les étudie chacune séparément. Il faut donc généraliser en passant par la notion de genre et de famille. En fournissant un cadre dans lequel les données d'observation peuvent clairement s'insérer, la généralisation aide à retenir les connaissances acquises. Elle permet également de réduire le nombre de données à retenir, car bien des caractéristiques sont communes à tous les membres d'une même famille. Enfin, elle ouvre la voie à un élargissement futur des connaissances, en ce sens que des arbres jusqu'alors inconnus peuvent être rangés dans une famille, puis rattachés, au moyen d'une clé propre à celle-ci, à tel genre ou à telle espèce. Avant d'en arriver à ce stade, il faut toutefois définir le concept de famille, difficile et souvent insaisissable.
C'est peut-être déjà trop que de vouloir atteindre un stade aussi avancé. Cependant, si, pour définir ce concept, l'on se fonde sur l'observation d'essences importantes du point de vue écologique ou économique, on y gagne de pouvoir les reconnaître, même s'il s'avère difficile d'aller plus avant dans la généralisation.
La formation des concepts de genre et de famille est une tâche difficile qui demande des observations nombreuses et approfondies. Il conviendrait pour cela de suivre deux principes: premièrement, considérer les arbres dans leur ensemble, et pas simplement comme des morceaux de bois couverts d'écorce ou des rameaux collés sur une page d'herbier. Wyatt-Smith (8) a donné un excellent exemple de l'emploi d'une gamme étendue de caractéristiques pour déterminer les arbres. Deuxièmement, l'enseignement ou l'acquisition des concepts devrait se faire en tenant compte des découvertes de la psychopédagogie dans ce domaine.
Il y a deux formules pour arriver au concept de famille: la plus généralement utilisée, surtout dans les manuels est déductive et formelle. Elle consiste à énumérer les caractéristiques de la famille, en la subdivisant au moyen d'une clé dichotomique et en décrivant les genres et les espèces qui la composent. Cette méthode paraît simple et commode. Sa principale faiblesse consiste en ce que peu de chercheurs peuvent concevoir la famille à partir de la définition, parce que la distinction entre les caractéristiques dont on se sert pour établir celle-ci est rarement nette. Par exemple, il n'est pas rare que des représentants de plusieurs genres, appartenant à une même famille, aient des feuilles alternes, opposées ou verticillées ou encore quatre, cinq ou six pétales. Ce processus de classification incomplet peut convenir à un spécialiste, mais il est tout à fait inutilisable à un niveau inférieur.
La deuxième méthode est inductive. Elle consiste à étudier un groupe d'arbres dont on sait qu'ils appartiennent à une famille donnée, pour arriver par l'observation et le raisonnement au concept de famille. C'est la méthode naturelle, applicable même lorsqu'il est difficile de décrire de façon précise les caractéristiques. Elle peut être pratiquée, et elle l'est, par des personnes manquant de formation; par exemple, la plupart des indigènes de Nouvelle-Guinée savent rattacher Viticipremnanovae pomeraniae à Vitex cofassus, bien que le premier ait des feuilles composées, palmatilobées, et l'autre des feuilles simples. Beaucoup d'entre eux reconnaissent Tectona, qui a été introduit dans le pays, comme étant un «frère» du Gemlina indigène; bien qu'ils n'aient jamais entendu parler de la famille des Verbenaceae, certains vont même jusqu'à classer les quatre genres mentionnés ci-dessus dans un «clan».
On peut distinguer cinq phases dans le processus inductif:
a) Observation des divers sujets;b) Formation d'une hypothèse fondée sur leurs éléments communs;
c) Mise à l'épreuve de l'hypothèse par l'étude de cas d'espèce;
d) Choix d'une catégorie générale dans laquelle peuvent s'insérer tous les sujets;
e) Dénomination de la catégorie.
Le même processus doit être suivi pour l'étude de la taxonomie. L'élève atteindra plus rapidement le concept de famille avec l'aide d'un professeur, mais, avec une bonne formation de base et un manuel de référence approprié, il pourra y parvenir seul. Voici maintenant quelques commentaires sur l'application de la méthode ci-dessus.
OBSERVATION DES ARBRES DONT ON SAIT QU'ILS APPARTIENNENT A TELLE OU TELLE FAMILLE
Il n'est certainement pas superflu, malgré les apparences, de donner des instructions sur la manière d'observer des arbres. Il faut apprendre à l'étudiant à observer des caractéristiques significatives; par exemple, de nombreuses légumineuses ont des fleurs qui attirent immédiatement le regard par leurs brillantes couleurs, alors que ce détail est sans importance; ce qui compte c'est la disposition des pièces florales, le type de fruit et la nature du bois. On peut répartir en trois catégories principales les nombreuses caractéristiques observables sur chaque arbre:
a) Caractères principaux utiles pour classer l'arbre considéré dans une famille ou un genre donné;b) Traits marquants utiles pour déterminer l'espèce particulière;
c) Traits secondaires utiles seulement pour confirmer la première détermination.
Cette observation fragmentée doit toutefois être complétée par l'observation de l'arbre dans son ensemble, car le tout est plus que la somme des parties. Il est possible, en effet, de reconnaître d'un coup d'œil et sans vérifier mentalement les caractéristiques de la famille, les représentants de familles nettement définies, telles que les Myristicacées. Or, il est difficile de dire sur quelles caractéristiques se fonde cette détermination: inclinaison des branches, disposition des feuilles, nature de l'écorce. C'est pourquoi seule l'expérience permet de déterminer un arbre au premier coup d'œil, le nombre d'observations nécessaires diminuant progressivement et l'observateur s'attachant moins aux caractères détaillés et bien définis qu'à l'impression d'ensemble.
Un bon herbier, une collection de bois et une facilité d'accès à la forêt sont bien entendu indispensables à cette étude. L'accès à la forêt présente moins de difficultés pour des établissements de formation au niveau inférieur que pour des universités.
Il faut que les étudiants puissent voir presque en même temps les éléments de l'herbier, les échantillons de bois et les arbres. Si l'on consacre un après-midi à l'étude d'une famille donnée, il faut exclure les représentants d'autres familles, même s'il s'en trouve à proximité. La clarté des résultats acquis de cette manière vaut que l'on fasse cet effort.
Une détermination rapide n'étant possible qu'après des observations répétées, il peut être utile aussi d'aménager un itinéraire botanique, en marquant les arbres d'une étiquette avec leur nom.
FORMATION DE L'HYPOTHÈSE
Dans ce cas, plus encore que dans la première phase, l'instructeur peut être d'un grand secours en attirant l'attention des élèves sur les caractères communs à une famille. Avant cela, il importe de donner aux élèves des notions de taxonomie. La terminologie employée doit s'étendre à l'arbre tout entier et non seulement à ses éléments reproducteurs, mais une extrême précision ne s'impose pas. Mieux vaut omettre les caractères invisibles à l'œil nu ou à la loupe, car ils sont de peu d'intérêt pour la détermination sur le terrain. Il est à noter que bien des termes utilisés dans la taxonomie et l'anatomie des arbres évoquent en eux-mêmes des notions déjà suffisamment complexes, puisqu'ils traduisent la corrélation existant entre les éléments composants: par exemple l'épigyne chez les Meurs et le parenchyme confluent dans le bois. A ce titre, ils sont utiles pour définir le concept de famille.
Lorsqu'on se sert d'un manuel, le parallélisme des descriptions et des illustrations aide à mettre en lumière les similitudes. La définition d'un concept impliquant une généralisation à partir de nombreuses observations particulières, l'utilisation d'auxiliaires tels que formules florales ou diagrammes hâtent le processus.
Durant l'étude d'une famille et au cours des observations ultérieures de sujets qui en font partie, il importe de reviser constamment le concept puisque celui-ci résulte d'une fusion des perceptions antérieures et des nouvelles observations. Le nouveau concept ainsi formé sera soit plus large soit plus restreint que le précèdent, selon les sujets qui auront été observés en premier lieu. S'inspirant de sa vaste expérience, l'instructeur peut à juste titre écarter des exemples qui conduiraient à un concept inutilement large. Pour la plupart des familles, l'étude des arbres exploités pour leur bois dans un pays donné aboutira à un concept plus étroit et plus précis que celle des plantes et des arbres du monde entier.
Le concept de famille´ est plus facile à définir dans certains cas que dans d'autres. Les représentants de familles étroites et denses comme les Myristicacées sont beaucoup plus faciles à grouper que ceux de familles plus larges, aux nombreuses variétés telles que les Euphorbiacées.
MISE A L'ÉPREUVE DE L'HYPOTHÈSE
Cette phase est importante non seulement parce qu'elle permet d'éprouver la qualité des observations antérieures, mais parce qu'elle: suscite la participation active des élèves. Deux attitudes sont possibles: on peut, soit exclure une espèce importante du concept initialement formé, soit choisir une espèce secondaire relativement peu importante. Dans l'un et l'autre cas, il faut à l'improviste montrer un arbre à l'élève pour qu'il s'efforce de le déterminer tout seul. L'élimination de caractéristiques qui ne présentent qu'une valeur spécifique aide à éclaircir le concept de famille.
DÉFINITION D'UNE ET DÉNOMINATION
Les élèves peuvent s'essayer à définir une famille mais généralement ils auront besoin de l'aide de l'instructeur. A ce stade, le nom de la famille peut être lié au concept. Cela peut paraître illogique mais sans formation préalable du concept, le nom est vide de toute signification.
Nous avons dit plus haut que l'emploi du concept de famille facilitait la mémorisation, en ce sens que les faits observés sont plus significatifs. Il serait intéressant de savoir si le même principe est valable pour les divisions plus vastes de la classification, telles que l'ordre. Dans la plupart des pays, la constitution des herbiers se fonde sur le système d'Engler et Prantl, dont l'intérêt réside dans le fait qu'il groupe les familles d'après leurs similitudes. Ainsi, les familles de dicotylédones se classent de la façon suivante: plantes apérianthées, ensuite celles qui sont dotées d'un périanthe à un verticille, puis d'un périanthe à deux verticilles disposés en spirale, puis d'un périanthe à deux verticilles cycliques, les pièces du verticille intérieur étant distinctes (dialypétales) ou, enfin, soudées (gamopétales). Cette méthode offre un cadre plus large pour ranger les données d'observations dans l'ordre qui convient. Il existe également des séries parallèles mais moins constantes, passant de l'apocarpie à la syncarpie, et de l'hypogynie à l'épigynie en passant par la périgynie. Etant donné le volume considérable de données que doit assimiler la mémoire, toute formule qui facilite la mémorisation vaut la peine d'être essayée.
PHASES FINALES
Au cours de l'élaboration du concept de famille, il conviendrait de faciliter le passage à la seconde phase de l'acquisition des connaissances, lorsque l'élève travaillera seul. La terminologie devrait donc, dès le début, suivre dans toute la mesure du possible les ouvrages de référence dont il se servira plus tard. Pour la Malaisie, ce sont incontestablement la Flora Malesiana et le fichier du Commonwealth scientific and industrial research organization (CSIRO) sur les arbres exploités dans la région du Pacifique du Sud-Ouest. La principale difficulté à cet égard réside dans la multiplicité des termes techniques utilisés par les botanistes professionnels.
Enfin, il faudrait enseigner aux élèves une méthode appropriée. pour déterminer les espèces qu'ils n'ont pas encore rencontrées. L'une de ces méthodes, utilisée par l'auteur du présent article, est la suivante: si, à première vue, l'espèce inconnue peut être rangée dans un genre ou une famille, il convient de vérifier immédiatement cette impression en comparant les caractéristiques de
l'espèce inconnue avec les caractéristiques connues des représentants de ce genre ou de cette famille. Si l'observateur n'a pas d'intuition immédiate, il note les caractéristiques de l'espèce inconnue et les compare avec les caractéristiques connues des différentes familles. La simple énumération des caractéristiques met souvent sur la bonne voie. Si l'on parvient à déterminer approximativement l'espèce, il faut, là encore, effectuer une vérification complète. Lorsqu'on dispose d'un herbier, le meilleur moyen d'opérer cette vérification est de comparer directement le représentant de l'espèce inconnue avec les spécimens; faute d'herbier, les manuels de référence doivent suffire. L'emploi de clés taxonomiques est indispensable dans la seconde phase. Comme la plupart des manuels traitent suffisamment de cette question, il est inutile de s'y attarder ici.
En conclusion, on peut se demander si la méthode proposée est applicable. L'auteur a constaté que certains élèves intelligents et curieux, ayant un niveau d'instruction moyen, pouvaient, après avoir observé pendant une année 150 des espèces les plus importantes en Nouvelle-Guinée, ranger correctement dans une famille ou un genre, beaucoup d'espèces jusqu'alors inconnues d'eux. D'autres élèves ne pouvaient reconnaître que les espèces qu'on leur avait effectivement fait observer, mais étaient incapables de saisir le concept de famille. Même lorsque la détermination est faite, elle n'est pas toujours correcte, et il est conseillé d'envoyer un échantillon de l'espèce pour confirmer le résultat au moyen d'un herbier. L'essentiel est qu'une base ait été établie et qu'un cadre ait été défini, ce qui permet de faire de nouveaux progrès. Avec des élèves d'un niveau d'éducation supérieur, on peut s'attendre à des résultats meilleurs encore.
Remerciements:
Je tiens à remercier M. W. Finlayson pour l'aide qu'il m'a apportée en lisant et en corrigeant mon manuscrit.
ENDERT, F.H. 1956. Key to the tree genera of Indonesia. Traducción al inglés por R.D. Hoogland, CSIRO, Canberra.
KRECH, D. y CRUTCHFIELD, R. S. 1959. Elements of psychology. Knopf, New York
LAWRENCE, G.H.M. 1951. Taxonomy of vascular plants. Macmillan, New York.
MENON, P.K.B. 1961. Taxonomic value of wood anatomy seen through Malayan woods. Malayan Forester, 24: 290-301.
MERRILL, E.D. 1954. Plant life of the Pacific. New York, Macmillan.
WHITMORE, T.C. 1963. Bart, morphology as an aid to forest recognition and taxonomy of Dipterocarpaceae. Flora Malesiana Bulletin, 18: 1017-1019.
WOOD, G.H.S. 1952. Bark as means of tree identification. Journal of the Oxford University Forest Society. Ser. 3, 6: 15-27.
WYATT SMITH, J. 1954. Suggested definitions of field characteristics. Malayan Forester, 17: 170-183.
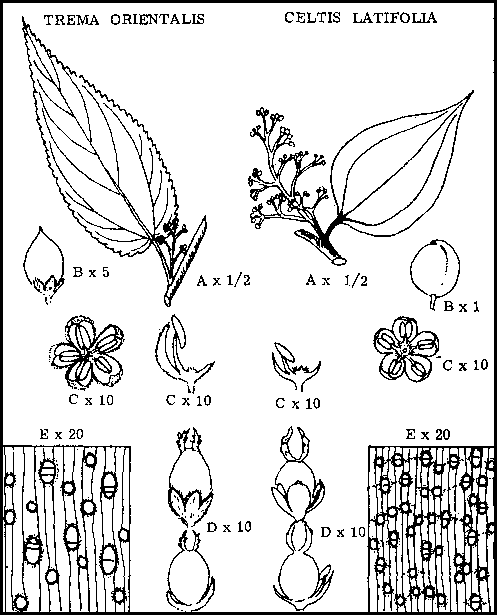
Il existe en Nouvelle-Guinée deux autres genres peu importants d'arbres qui ressemblent à Trema. Comme Celtis, ils appartiennent à la famille des Ulmacées.
Les caractéristiques de cette famille, observées chez Celtis et Trema sont les suivantes:
Les feuilles sont simples, alternes, à bord lisse ou denté.
Arbres monoïques, fleurs surtout unisexuées, composées d'un périanthe à 4 ou 6 lobes, avec un nombre égal d'étamines chez les fleurs mâles et un ovaire unique à une seule loge portant deux styles chez les fleurs femelles. Le fruit est la plupart du temps une petite drupe, faiblement charnue a l'extérieur, avec un endocarpe dur.
Bois avec rayons médullaires étroits, pores de taille petite à moyenne en chaînes courtes, et parenchyme entourant les pores.
Le trait discontinu indique les caractéristiques propres a la famille; le trait continu souligne les caractéristiques spécifiques importantes.
CELTIS LATIFOLIA Planch.
|
Nom commercial |
Celtis |
|
Port |
Arbre de grande taille pouvant atteindre 42 m de hauteur et 1 m de diamètre. Fût à contreforts, long, mince, allant en décroissant, cime de largeur moyenne. |
|
Ecorce |
Epaisseur 1,2 cm. Assise externe grise lisse, sauf quelques pustules et quelques arêtes horizontales saillantes. Assise intermédiaire verte; assise intérieure de couleur ivoire à jaune paille, structure en relief rouge foncé «poivre et sel», dure, non fibreuse. Les coupes foncent rapidement. |
|
Bois |
Couleur ivoire à jaune paille, dureté et densité moyennes, rayons médullaires étroits, pores de taille moyenne, assez nombreux et disposés en courts rayons, entourés de parenchyme. Etalé en forme d'aile et confluent. |
|
Feuilles |
Simples, alternes, ovales, souvent asymétriques, 3,8 sur 7,62 cm, sommet acuminé, base ronde, bord lisse, multinervées, ternées, d'épaisseur moyenne, coriaces, de couleur vert foncé luisant sur la face supérieure; face inférieure vert clair glabre. |
|
Inflorescences |
Sous forme de panicules axillaires ou terminales, petites, jaunes, vertes, faiblement odorantes. Les fleurs sont soit unisexuées et monoïques, soit d'apparence bisexuées. Les fleurs mâles ont un périanthe composé de 5 éléments et 5 étamines qui leur sont opposées Les fleurs femelles ont aussi un périanthe composé de 5 éléments avec un gros ovaire ovoïde muni de deux styles et des étamines faiblement développées. |
|
Fruit |
Drupe globuleuse pointue peu charnue d'un bleu noir; brillant lorsqu'elle est arrivée à maturité. |
|
Habitat |
Forêts humides de plaine dans tout le territoire, associées avec différentes espèces d'arbres feuillus. |
|
Autres espèces |
Il existe sur le territoire six espèces de Celtis qui diffèrent surtout par la taille et la forme des feuilles. Chez deux d'entre elles, le réseau de nervures n'est pas nettement terné. |
Pour identifier les espèces sur le terrain, on se fonde sur la structure «poivre et sel» de l'écorce, sur la structure du bois et dans la plupart des cas, sur le réseau de nervures ternées.
TREMA ORIENTALIS (Linn.) Bl.
|
Port |
Arbre de taille petite à moyenne, pouvant atteindre 21 m pour 38 cm de diamètre. Fût droit, se couvrant facilement de branches gourmandes. Cime moyenne à large, feuillage peu dense. |
|
Ecorce |
Epaisseur de 63 à 84 mm, assise extérieure grise, lisse; assise intérieure verdâtre, très fibreuse, utilisée pour la fabrication de cordages. |
|
Bois |
Tendre et clair, couleur paille, à grain grossier, pores de taille moyenne disposés en chaînes courtes, rayons médullaires étroits et sinueux, pores entourés de parenchyme. |
|
Feuilles |
Simples, alternes, distiques, ovales, acuminées au sommet à base cordiforme et bord denté, uninervées mais avec deux nervures basilaires fortement développées, d'épaisseur moyenne, vertes, légèrement pubescentes sur la face supérieure; argentées, fortement pubescentes sur la face inférieure. |
|
Inflorescences |
Sous forme de petites panicules axillaires, fleurs petites, vert jaunâtre unisexuées sur le même arbre. Les fleurs mâles se composent d'un périanthe à 5 lobes; 5 étamines leur sont opposées. Les fleurs femelles se composent d'un périanthe à 5 lobes avec un gros ovaire portant deux styles. |
|
Fruits |
Petits, 3,7 mm de diamètre, ovoïdes, drupes noires - peu charnues; endocarpe dur, lobes du périanthe persistants. |
|
Autres espèces |
Il existe sur le territoire deux autres espèces de Trema qui diffèrent surtout par le degré de pilosité des feuilles. |
|
Habitat |
Les trois espèces de Trema se rencontrent sur tout le territoire de la Nouvelle Guinée, dans les basses terres et les régions montagneuses jusqu'à environ 1500 m d'altitude; elles sont très communes dans les parcs. |
TREMA ORIENTALIS - CELTIS LATIFOLIA
![]()
![]()
![]()