![]()
![]()
![]()
On peut distinguer deux types de données, les statistiques de pêche (prise, effort) d'une part et les données biologiques (sexes, mensurations, âge, état sexuel, etc.) d'autre part.
Le recueil des statistiques permettant de retracer l'évolution simultanée du stock et du système d'exploitation est l'un des aspects essentiels de l'étude des stocks exploités. On distinguera, pour des raisons pratiques, les deux types principaux d'exploitation des crevettes, la pêche chalutière industrielle sur les adultes et la pêche artisanale sur les juvéniles.
4.1.1 L'exploitation industrielle
La pêche à la crevette en mer est un exemple classique de pêche au chalut de fond. Son approche ne présente aucune difficulté particulière et on pourra se référer pour la théorie de base, aux travaux classiques de Beverton et Holt (1956) et de Gulland (1969). Les données essentielles concernent la flottile elle-même et son activité. Elles seront, pour la commodité d'un traitement ultérieur, stratifiées en fonction de zones géographiques préétablies.
4.1.1.1 Les sources de données
(a) Les fichiers des administrations chargées des pêches. Ils permettent, lorsqu'ils existent, de reconstituer l'évolution de la flottille en nombre de vaisseaux et éventuellement en puissance totale et autres caractéristiques globales.
(b) Les registres de mouvements journaliers obtenus auprès des capitaineries des ports ou tenus par les bureaux d'enquêtes, permettent de connaître avec précision les bateaux à quai et d'en déduire ceux qui sont en mer. Cette donnée permet de calculer le temps d'absence du port ou temps de mer.
(c) Les feuilles d'enquête, établies par les enquêteurs auprès des patrons de pêche au retour de chaque marée, permettent de connaître, pour toute ou partie de la flottille, la date de départ, de retour, la zone de pêche, les espèces recherchées, les tonnages capturés, etc. Ces feuilles dont la complexité dépendra surtout de la bonne volonté des patrons et des relations entre la recherche et la profession, permettront de calculer le temps de pêche effectif et de contrôler éventuellement les débarquements déclarés par les usines ou les criées. L'expérience montre en effet que les patrons sont capables d'apprécier, avec une bonne précision, le tonnage capturé (Fig. 22).
(d) Les feuilles de maréyage obtenues à la criée et dans les usines permettent d'obtenir, pour chaque bateau, la prise par espèces (principales et accessoires) et le prix de vente au kilo. La date du débarquement permet de rattacher ces données à une marée précise, pour laquelle d'autres renseignements sont obtenus (voir plus haut).
(e) Les cahiers de pêche. Ils représentent une source inestimable de données pour le biologiste des pêches. Le nombre et la précision des renseignements fournis dépendront des possibilités des patrons et de leur niveau d'instruction. Dans les meilleurs cas, on pourra obtenir des données pour chaque coup de chalut effectué (heure de mise en pêche, de fin de pêche, position, profondeur, espèces capturées, quantités, changement dans les pratiques de rejets, informations sur l'engin utilisé).
Ces données très détaillées nécessitent souvent un traitement automatique dont un exemple est donné par Slack-Smith et Stark (1969). Bien utilisées, elles permettent un suivi remarquable de l'évolution de la pêcherie, des tactiques de pêche, des études détaillées sur les distributions géographique et bathymétrique des différentes espèces, des rythmes d'activité (voir Garcia, 1976, par exemple). Elles ne pourront en général être obtenues que pour un échantillon de la flottille dont la représentativité devra être analysée.
Des exemples d'études de pêcheries de crevettes avec une description des données recueillies peuvent être trouvés, outre les travaux cités plus haut, dans Snow (1969) pour le Golfe du Mexique, Slack-Smith (1969) en Australie, Chabanne et Plante (1970) et Marcille (1978) à Madagascar. Un inventaire des systèmes de collecte des statistiques dans les Caraïbes a été réalisé par Mackett (1972).
Ces différentes sources fournissent les données de base pour évaluer l'effort total déployé et la capture réalisée.
4.1.1.2 L'effort de pêche
Plusieurs mesures de cet effort pourront être obtenues suivant l'efficacité du réseau de collecte. De la plus grossière à la plus précise on peut distinguer:
(a) La taille de la flottille. En l'absence de toute donnée détaillée, le nombre total d'unités en activité, la puissance motrice globale ou toute autre approximation de la puissance nominale pourra être utilisée comme mesure de l'effort. Le temps pendant lequel cette puissance a été appliquée est en général sous-entendu et égal à l'année. Il n'est pas possible de déceler des variations dans l'utilisation réelle de cette puissance (évolution des gréements, des temps de pêche effectifs, etc.).
(b) Le nombre de marées est également une donnée grossière qui sans renseignements complémentaires sur la durée moyenne de ces marées ou la puissance motrice des bateaux concernés peut être à l'origine de biais importants.
(c) Le temps d'absence au port ou temps de mer: obtenu facilement à partir des pointages quotidiens, il ne permet pas de déceler les variations dans les temps morts (temps de route par exemple).
(d) Le temps de pêche représente le temps réellement passé sur les lieux de pêche, les temps de route étant décomptés. Il ne permet pas de tenir compte des temps morts (arrêts pour avarie, pour conditionnement des captures ou rendements trop faibles en attendant une phase plus favorable du rythme nycthéméral de capturabilité).
(e) Le temps de drague: il représente l'effort réellement exercé sur le stock. Les temps morts consacrés à des changements de lieux de pêche, au conditionnement des captures, au tri du contenu du chalut, sont décomptés. Pratiquement cet effort ne peut être évalué qu'à partir des cahiers de pêche ou, à défaut, d'enquêtes très détaillées sur un échantillon suffisant d'unités.
4.1.1.2 La normalisation des données d'effort
Les notions classiques d'effort de pêche, nominal ou standard, d'intensité et de puissance de pêche pourront être retrouvées dans les ouvrages classiques de Beverton et Holt (1956), Ricker (1975), Gulland (1969). Une réunion spéciale sur ce problème s'est tenue en 1970 (Pope, 1975). Rothschild (1972, 1977) a tenté de redéfinir la notion d'effort de façon plus moderne. Un document récent de la FAO (1975) rappelle les principes essentiels de l'utilisation de l'effort pour évaluer l'abondance.
Le recours à la standardisation implique la constitution d'un fichier des bateaux de pêche et de leurs caractéristiques (nom, numéro, taille, puissance, jauge, gréement, maillage utilisé, date de construction, date d'entrée dans la pêcherie, mode de conservation des captures, etc.). Ce fichier doit être tenu à jour régulièrement, ce qui peut être une tâche ardue au début de la période d'exploitation tant que la composition de la flottille n'est pas stabilisée.
Les problèmes posés par la pêche crevettière se rattachent au cas général de la pêche au chalut pour lequel la littérature est plus qu'abondante. Nous rappellerons donc seulement de façon très schématique que la propriété fondamentale d'un effort de pêche correct pour un biologiste des pêches est qu'il est proportionnel à la mortalité par pêche (F) et relié à cette dernière par l'équation
F = q.f
où q est le coefficient de mortalité par pêche par unité d'effort, ou coefficient de capturabilité de Ricker. Pour répondre à cet impératif l'effort de pêche nominal doit être standardisé. L'objectif essentiel de cette opération est de déceler les variations importantes et surtout les tendances de la relation entre f et F. Pour la pêche à la crevette, les principales dérives que l'on peut redouter sont:
l'évolution de la puissance motrice des unités de pêche,
l'évolution du gréement avec passage d'un gréement simple (type Vigneron-Dahl) à un gréement double de type floridien, ou “twin”,
l'évolution des chaluts et une tendance éventuelle à une augmentation de leur taille pour une puissance motrice donnée, ou à un changement de forme affectant leur efficacité,
l'évolution de la tactique de pêche: adoption du chalut d'essai, augmentation du nombre d'heures de drague par jour, chalutage de nuit ou de jour, variation du rapport entre le temps de pêche effectif et la durée d'absence au port avec la modernisation de la flottille et la généralisation de la congélation entrainant un allongement des marées, changements dans la répartition de l'effort par espèce sur un stock multispécifique, passage d'une recherche individuelle des bancs à une recherche en flotille, avec introduction d'un effet de “meute” (Marcille, 1978).
Une des caractéristiques essentielles de la pêche à la crevette est qu'elle peut être effectuée soit de nuit, soit de jour, soit toute la journée. Cela dépend de la saison, de la région, de l'espèce et du type de bateau.
Garcia (1977) a montré, en Côte d'Ivoire, que les congélateurs de 400 ch procédant au conditionnement à bord, sélectionnaient dans une journée les tranches horaires les plus favorables et restaient à l'ancre le reste du temps pour économiser le carburant ou procéder au tri et à l'empaquetage des captures. Il a pu donc distinguer deux tactiques d'exploitation:
La pêche de jour ou “sélective” (11 heures de drague en moyenne),
la pêche de nuit et de jour ou “normale” (15 heures de drague).
Il est évident, dans ce cas, que la sélection des tranches horaires les plus favorables entraine une augmentation du rendement horaire au détriment du rendement journalier. La puissance de pêche relative d'un congélateur utilisant cette tactique dépendra donc de l'unité d'effort utilisée (heure de drague ou jour de pêche) et du nombre d'heures de travail effectif. On voit sur la figure 23 que lorsque l'effort est exprimé en heures de drague, la puissance relative1 passe de 1,25 en pêche “normale” à 1,8 en pêche “sélective”. Inversement, si l'effort est exprimé en jours de pêche, elle passe de 1,25 en pêche “normale” à 1,0 en pêche “sélective”.
Si une flottille est composée d'unités exploitant différemment les tranches horaires et si la composition de la flottille, ou la tactique, change avec la saison, ou au cours des années, on a là une source de biais très importante. La difficulté peut être tournée simplement en adoptant une puissance de pêche relative variable suivant la tactique utilisée. Marcille (1978) a montré des variations saisonnières ainsi qu'une augmentation progressive de 1969 à 1973 du nombre d'heures de drague par jour à Madagascar (Fig. 24A). Il a également mis en évidence une variation saisonnière de la puissance de pêche relative des bateaux (Fig. 24B). Il suggère une liaison avec des variations saisonnières de disponibilité de la crevette affectant différemment les divers types de bateaux. Le passage d'un comportement semi-pélagique (de janvier à juin) à un comportement benthique (de juillet à décembre) des crevettes affecterait différemment les petits et gros crevettiers car dans le premier cas le rapport des rendements est dans le rapport des surfaces d'ouverture des chaluts alors que dans le deuxième cas il est dans le rapport des longueurs de bourrelet.
1 Par rapport à un crevettier type de 250 ch pêchant nuit et jour
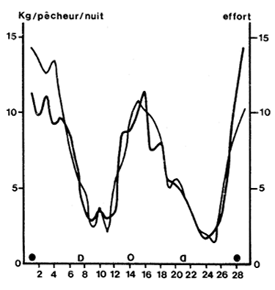 | 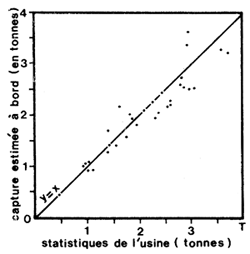 |
Fig. 21: Variations lunaires de l'effort (en nombre de pêcheurs actifs) et du rendement dans la pêcherie artisanale de Côte d'Ivoire (d'après Garcia, 1973) | Fig. 22: Relation entre la capture estimée par le patron de pêche et le tonnage réel, contrôlé en usine (d'après Garcia, 1977) |
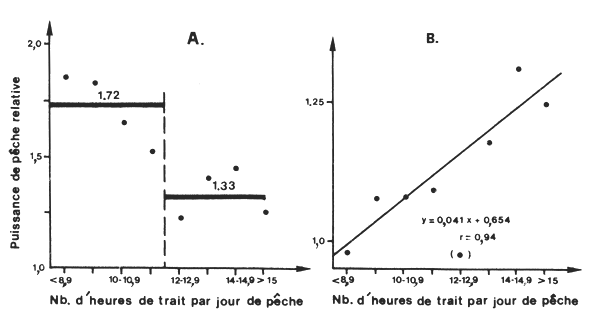
Fig. 23: Variations de la puissance de pêche relative des crevettiers congélateurs de Côte d'Ivoire en fonction du temps de pêche moyen journalier (d'après Garcia, 1977)
A. Quand l'effort est exprimé en heures de trait
B. Quand l'effort est exprimé en jours de pêche
Les principaux procédés de normalisation sont bien connus. Dans la pêche chalutière, les puissances de pêche relatives des bateaux sont en général très liées à leur puissance motrice ou à leur jauge brute. La normalisation s'effectue donc le plus souvent à partir d'une relation entre les captures par unité d'effort nominal et la puissance motrice ou la jauge. Les puissances de pêche relatives sont définies par rapport à un type de bateau de référence. Dans le cas où les zones de pêche relatives sont nombreuses et la distribution des bateaux des différents types est hétérogène, la méthode de Robson (1966) permet de calculer des puissances de pêche relatives avec une bonne précision statistique et de déceler des variations spatio-temporelles éventuelles. Un programme écrit par C. Berude est disponible (Abramson, 1971). Une des caractéristiques des relations entre la puissance de pêche relative et la puissance motrice chez les crevettiers est leur faible pente. Un doublement de la puissance motrice de 200 à 400 CV se traduit par une augmentation de 50 pour cent environ seulement de la puissance de pêche relative, aussi bien en Côte d'Ivoire (Garcia, 1977) qu'à Madagascar (Marcille, 1978). Le tableau suivant rappelle quelques relations proposées:
| Références | Régions | Equations |
| Marcille (1978) | Madagascar | Pr = 0,0029 Pm + 0,530 |
| Garcia (1977) | Côte-d'Ivoire | Pr = 0,0020 Pm + 0,460 |
| Neal (1970, 1975) | U.S.A. | Pr = b.L + a |
0,424 < b < 0,630 | ||
0,022 < a < 0,036 |
Tableau: Relation entre la puissance de pêche relative (Pr) des crevettiers et leurs caractéristiques.
Pr = rendement horaire du bateau considéré/rendement du bateau type
L = longueur du bateau
Pm = puissance motrice
Griffin, Nichols et Lacewell (1973) ont donné une relation plus complexe:
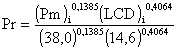
dans laquelle (Pm)i et (LCD)i indiquent la puissance motrice et la longueur de corde de dos du bateau de classe i, et 38,0 et 14,6 sont respectivement la puissance motrice et la longueur de corde de dos du bateau type.
Une autre caractéristique importante de la relation entre la puissance de pêche relative et les caractéristiques du bateau est la grande dispersion des points autour de la droite. Cette relation traduit d'une part la diversité des types de chaluts utilisés (flat, balloon, semi-balloon, etc.) et des stratégies appliquées (pêche de jour, de nuit, de jour et de nuit, pêche individuelle ou en meute, mais également, et d'une manière importante semble-t-il, le facteur personnel lié à l'habileté du patron et à son expérience.
Dans le cas de pêcheries de crevettes multispécifiques, le problème le plus complexe est celui de la détermination de l'effort dirigé sur chacune des espèces. Elle peut être très difficile si les espèces ne sont pas nettement séparées géographiquement. Faute de mieux, on considère souvent dans ce cas que l'effort de pêche est réparti au hasard sur les différentes espèces capturées et que les captures par unité d'effort (cpue) sont proportionnelles à la densité réelle des diverses espèces. Ceci n'est vraisemblablement pas exact dans le cas d'une pêche aussi dirigée que la pêche à la crevette et les risques de biais sont importants lorsque les indices de concentration de ces flottilles, sur une espèce donnée, évoluent dans le temps (ce qui équivaut à un changement de q). C'est la situation la plus courante dans les pêcheries multispécifiques (Louisiane, Madagascar) où l'espèce principale est d'abord exploitée seule, surtout si elle est abondante le jour (Penaeus setiferus et Penaeus indicus) et ce n'est que lorsque les rendements baissent sur l'espèce-cible que les espèces secondaires sont recherchées. Boerema (1974) indique, pour la pêcherie de Panama, des modifications progressives de ce type entre Penaeus et Xyphopenaeus. Dans le cas où la composition des prises dépend de l'heure de chalutage comme à Magadascar (Chabanne et Plante, 1971), un simple changement des tranches horaires exploitées suffit à modifier les coefficients de capturabilité des différentes espèces. Ces changements tendent en général à exagérer les variations des indices d'abondance des différentes espèces.
4.1.1.4 La capture
Il est nécessaire de connaître la capture totale réalisée pour chacune des espèces exploitées lorsqu'il y en a plusieurs.
4.1.1.4.1 Les espèces-cibles
La capture est parfois facile à obtenir au niveau des usines car la haute valeur commerciale de la crevette entraîne un conditionnement soigneux et les espèces sont souvent distinguées. Lorsque la pêcherie porte sur plusieurs espèces de crevettes, les statistiques d'usine sont parfois difficiles à utiliser en raison des mélanges. A Madagascar, Marcille (1978) note les catégories commerciales suivantes:
HL white: Penaeus indicus calibré et étêté,
HL pink: Metapenaeus monoceros calibré et étêté,
HL tiger: Penaeus semisulcatus calibré et étêté, avec en outre quelques P. japonicus,
HO: crevettes entières comprenant une majorité de M. monoceros avec aussi des petites P. indicus et P. semisulcatus
Tiger HO: comportent de grosses P. monodon non étêtées, P. monodon étêtées et quelques P. semisulcatus
Mixed: catégorie pouvant comprendre différentes espèces étêtées et non calibrées.
HO est utilisé pour “head on” (crevette entière) et HL pour “headless” (crevette étêtée).
Dans le cas de tels mélanges, seul un échantillonnage régulier permettra de définir les proportions. Lorsque deux espèces de taille différente sont mélangées dans une catégorie, on a parfois une distribution biomodale des calibres permettant une séparation approximative (Marcille, 1978).
Dans chaque catégorie commerciale, les crevettes sont souvent calibrées suivant parfois une norme locale, mais le plus souvent suivant la norme américaine (en nombre par livre). La collecte de cette information très importante permet de reconstituer les structures de taille des captures (paragraphe 4.2). Compte tenu des divers conditionnements utilisés, il peut être nécessaire d'effectuer des transformations des données (crevettes entières en étêtées ou vice versa). On peut alors utiliser un coefficient empirique global, obtenu en pesant une quantité importante de crevettes avant étêtage et après. Aux Etats-Unis, le rapport utilisé est de 1,680 (entière/étêtée) ou 0,595 (étêtée/entière). D'autres rapports, très proches, sont utilisés ailleurs. Une liste en sera trouvée dans FAO (1972) et Squires (1974).
On peut également utiliser des nomogrammes pour des transformations plus précises des calibres en taille moyenne ou poids moyen individuel, ou pour passer d'un conditionnement à l'autre. Ces nomogrammes sont établis à partir des relations biométriques classiques (Fig. 25). Kutkuhn (1962) en donne quelques exemples et la manière de les construire. Cependant, compte tenu du fait que la droite doit être utilisée dans les deux sens, il serait préférable de calculer une relation fonctionnelle telle que l'axe majeur réduit (ou droite de Tessier ou “GM regression” d'après Ricker, 1973).
Compte tenu de la précision nécessaire, il est également préférable que chaque espèce fasse l'objet d'une relation propre à calculer avec soin. On verra au paragraphe 4.2 la possibilité d'utiliser avec une meilleure précision des clés de transformation pour obtenir, à partir des débarquements par calibre, des distributions de tailles.
4.1.1.4.2 Les espèces accessoires
Pratiquement toujours négligés, les poissons qui constituent un complément de la capture sont débarqués en tonnages très faibles car la capacité de stockage des bateaux est souvent limitée. Aussi, seules les espèces de belle qualité sont-elles conservées. Elles sont cependant un élément non négligeable du compte d'exploitation et doivent être prises en considération pour toute étude économique. Enfin, toute politique globale d'aménagement des ressources d'une région doit tenir compte de toutes les espèces en raison des interactions possibles sur une espèce donnée de deux types de pêcheries (par exemple la pêche chalutière et la pêche crevettière sur les sciaenidés).
Nous avons déjà vu qu'en général le patron de pêche est capable d'estimer sa capture avec une bonne précision (Fig. 22) et qu'en l'absence de données précises à terre (en cas de transbordement en mer par exemple), les données de prises obtenues par enquête sont utilisables.
4.1.1.4.3 Les rejets (Fig. 26)
La combinaison des données d'effort et de prise conduit à calculer les captures par unités d'effort (cpue). Si l'effort est correctement représenté, la capture est proportionnelle à la densité du stock
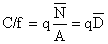 | d'après Gulland (1969) |
où N et D représentent respectivement l'abondance numérique moyenne et la densité moyenne pendant une période observée, et A l'aire de la zone de pêche. Il en résulte que la donnée de base doit être la capture réelle et non la quantité débarquée dans la mesure où une partie des captures peut être rejetée. Le plus souvent, seuls les débarquements sont connus et ceci peut entraîner un biais grave dans la mesure où les pratiques de rejet changent souvent avec l'évolution de la pêcherie et les prix de vente par catégorie de taille.
Notons que les rejets ont été étudiés par Berry et Benton (1969), Baxter (1973) et Juhl et Drummond (1977) dans le golfe du Mexique et la région Caraïbes. Ils ont une importance considérable, non seulement pour l'espèce-cible elle-même, mais pour toutes les espèces d'accompagnement.
Dans un premier temps, les ressources des fonds à crevettes ont le plus souvent été considérées comme mono ou paucispécifiques et en tous cas constituées uniquement de crevettes, en raison de la valeur marchande nettement supérieure de ces dernières et du faible intérêt des crevettiers pour le poisson. Il n'en reste pas moins qu'un plan d'aménagement global des ressources d'une région implique la connaissance de l'effort appliqué par la pêche à la crevette sur les espèces associées. La grande majorité de ces espèces est actuellement rejetée et Juhl et Drummond (1977) évaluent à 350 000 t/an les quantités rejetées dans le Golfe du Mexique et les Caraïbes. Klima (1976) les évalue à 900 000 tonnes maximum, à raison de 1,4 à 9,0 kg de rejets par kilo de crevettes capturées.
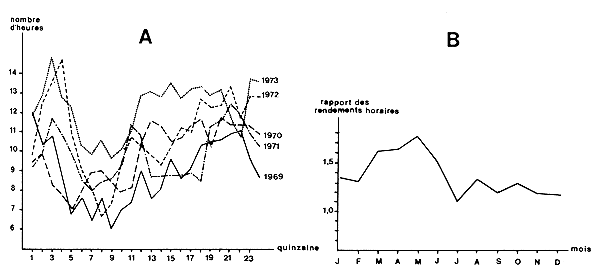
Fig. 24: Variations saisonnières des caractéristiques de l'effort de pêche
A. Evolution du nombre d'heures de drague par jour entre 1969 et 1973 à Madagascar
B. Variations saisonnières de la puissance de pêche relative
(d'après Marcille, 1978)
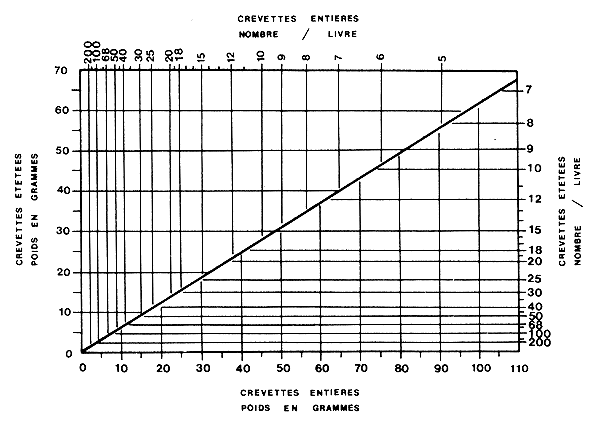
Fig. 25: Nomogramme de conversion des poids et nombre de crevettes entières et étêtées (d'après Kutkuhn, 1962)
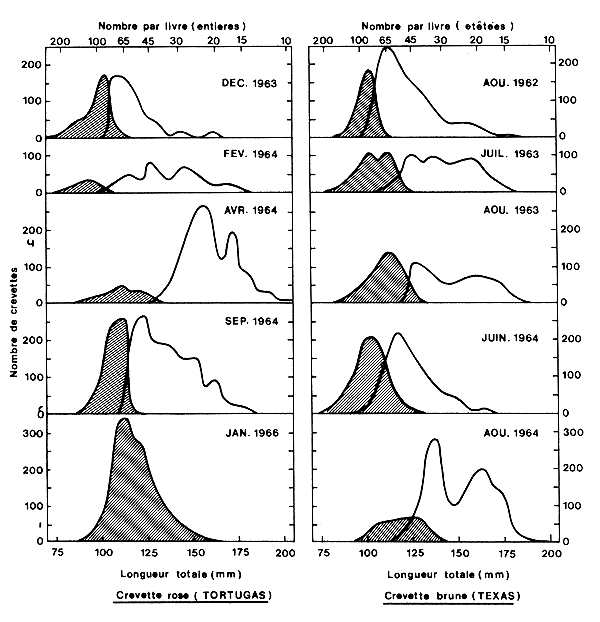
Fig. 26: Distribution de fréquence des crevettes débarquées (en blanc) et rejetées (hachures) en Floride et au Texas (d'après Berry et Benton, 1969)
Il existe plusieurs méthodes d'évaluation des rejets: elles sont de deux types:
(a) Méthodes directes. On peut effectuer des enquêtes auprès des patrons de pêche, à la fin de chaque marée, pour obtenir leur estimation des tonnages rejetés et une idée des principales espèces affectées. Si l'on peut vaincre la réticence générale des pêcheurs à parler de ce phénomène, on arrive de cette manière assez rapidement à une évaluation grossière des rejets. La méthode peut être affinée si les patrons acceptent de remplir les cahiers de pêche mis à leur disposition. Si l'on désire avoir une idée plus précise des rejets, la meilleure méthode est l'observation directe par un scientifique embarqué, effectuant des échantillonnages avant le tri de la capture.
(b) Méthodes indirectes. L'évaluation des quantités rejetées, de leur composition spécifique et des tailles concernées peut être obtenue par un navire de recherche effectuant des pêches avec des engins comparables à ceux des professionnels. Le profil spécifique des captures expérimentales est ensuite comparé à celui des captures débarquées. On pourra en déduire, pour chaque espèce, une courbe de rejet analogue à une courbe de sélectivité.
Une méthode mixte consiste à embarquer un professionnel à bord du navire de recherche et à lui demander d'effectuer le tri de la capture comme il l'aurait fait à bord de son navire. On a alors une approche directe et expérimentale des pratiques de rejets. Ces pratiques sont complexes dans la mesure où elles dépendent de plusieurs facteurs tels que:
l'importance des crevettes de petite taille dans les captures,
leur valeur au débarquement,
la méthode de tri (Berry et Benton, 1969),
les règlementations éventuelles sur une taille minimale commercialisable.
Les rejets d'espèces associées dépendent, eux:
des rendements obtenus sur l'espèce-cible,
de la taille des espèces associées rencontrées et de leur valeur marchande,
du mode de conservation utilisé par le bateau.
Les rejets sont donc susceptibles de variations saisonnières importantes, et l'analyse de ces variations peut conduire à la constitution d'un modèle simple. C'est ainsi que Berry et Benton (1969) ont établi une régression multiple de la forme
Y = a + b X1 + c X2 + d X3
ou Y est le tonnage de crevettes rejeté par mois, X1 le tonnage total débarqué, X2 le tonnage de petites crevettes débarqué par mois et X3 le prix de vente de l'unité de poids des petites crevettes.
Ils ont également utilisé l'équation
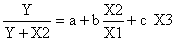
Ces deux modèles ne permettent cependant d'expliquer que 42 et 55 pour cent de la variance totale des rejets.
4.1.2 L'exploitation artisanale
4.1.2.1 Les sources de données
Elles dépendent du niveau d'organisation du système. S'il est bien organisé, le problème se ramène à celui du chapitre précédent, compliqué par le fait que les points de débarquement sont nombreux et dispersés. C'est le cas des pêcheries artisanales qui se sont développées dans les lagunes et estuaires des Etats-Unis. Des formulaires classiques peuvent alors être mis en place.
Dans le cas d'un système artisanal à collecte organisée, comme ceux que l'on rencontre en Afrique de l'Ouest, de bonnes statistiques de débarquement peuvent être obtenues au niveau des usines de conditionnement. Les rejets devront alors être évalués comme précédemment.
Dans le cas d'une pêcherie totalement inorganisée, plusieurs sources de données doivent être recherchées.
(a) Les fiches de pêche. Il est souvent difficile, mais parfois possible, d'obtenir la collaboration d'un habitant lettré qui accepte de remplir des fiches de pêche portant l'heure de mise en pêche et de fin de pêche, la quantité capturée, le nombre de filets posés. Ce système a été utilisé avec succès par l'un des auteurs en Côte d'Ivoire (Garcia, 1978). Il est certain qu'il peut difficilement être généralisé à toute une région, et ne peut concerner qu'un échantillon de la pêcherie.
(b) Les données de maréyage. Lorsque les circuits de commercialisation ne sont pas trop diffus, ou lorsqu'il existe des points de passage obligatoires (bacs, ports secondaires), une statistique des quantités transportées peut être obtenue à l'aide d'un pointage des véhicules (camionnettes ou pinasses) et d'une évaluation de leur contenu. La partie des captures autoconsommées échappe à l'enquêteur. Elle est en général négligeable si la commercialisation est bien organisée car le prix de vente des crevettes est si élevé que le pêcheur préfère vendre toute sa capture. Elle peut être importante si une règlementation interdit la vente des petites tailles qui sont alors transformées ou consommées sur place.
(c) Les enquêtes classiques par sondage sur les lieux de pêche. Il est évident que, bien plus que pour l'exploitation industrielle, une bonne connaissance de la pêche artisanale requiert un bon réseau d'enquête et un excellent contact des chercheurs avec le milieu de la pêche et les professionnels du maréyage.
4.1.2.2 La notion d'effort de pêche
Elle est très complexe dans le cas de la pêche artisanale au filet fixe. Garcia (1977), utilisant des feuilles de pêche remplies tous les soirs dans quelques campements en lagune Ebrié (Côte d'Ivoire), a mis en évidence de nombreuses sources de variation. Nous les résumons ci-après:
(a) L'effort improductif. Lorsque l'on utilise les statistiques d'usines ou celles des coopératives, les sorties produisant des prises nulles ne sont pas enregistrées. Il y a donc sous-estimation de l'effort et surestimation de la prise par unité d'effort. Le biais est maximum en morte saison.
L'unité d'effort utilisée est la nuit de pêche d'un pêcheur ou la pose d'un filet pendant la durée du jusant. Si le nombre de filets par pêcheur est stable, ces deux unités sont équivalentes.
(b) Le cycle lunaire. On voit sur la figure 21 que la prise par unité d'effort et l'effort nominal enregistrés suivent la même oscillation lunaire. Ceci est une constatation quasi générale en pêche artisanale en Afrique de l'Ouest où l'effort est souvent lié à l'abondance. Tout le monde met en pêche quand l'abondance est élevée et une grande partie des pêcheurs s'abstiennent en morte période pour s'adonner à d'autres travaux, dans les champs en particulier.
Un examen plus approfondi montre qu'à ce rythme, qui affecte le nombre de pêcheurs au travail, se superpose un autre rythme: celui de la durée du temps de pêche. Ce dernier est délimité par la conjonction entre le crépuscule et le jusant et il est maximum quelques jours après la pleine et la nouvelle lune (Fig. 27).
(c) Le cycle saisonnier. Quelle que soit la phase de lune concernée, le temps de pêche varie dans l'année. En lagune Ebrié, il augmente de février à août, où il est maximum, et passe par un second maximum en décembre. En octobre–novembre il accuse une nette diminution qui paraît liée aux crues (Fig. 28A). Le schéma saisonnier peut étre différent si l'on examine deux années successives en un même point ou trois points de pêche successifs la même année (Fig. 28B). Ces phénomènes qui sont vraisemblablement sous la dépendance des processus hydrauliques (cycle des marées, des pèches, des crues, phénomènes de résonnance, opposition marées/crues) n'ont pas été expliqués.
(d) Variations géographiques du temps de pêche. L'onde de marée se propageant et se déformant au fur et à mesure qu'elle avance dans l'estuaire, l'heure de mise en pêche et la durée de la pêche (de même que la force du courant utilisable et donc le volume filtré) varient de l'embouchure de l'estuaire vers l'amont de manière complexe (Fig. 29).
(e) Le choix de l'unité d'effort.
Les filets fixes: Le mètre cube d'eau filtrée serait la meilleure unité d'effort ainsi que l'a démontré expérimentalement Yokel (1970 - cf. paragraphe 2.3.4.2), dans la mesure où la prise par mètre cube serait un bon indice de la densité dans la masse d'eau. Cette donnée nous échappe cependant le plus souvent. D'autre part, si l'objectif de l'étude est de définir un indice de recrutement, il nous faut connaître, non la densité des crevettes en migration, mais le nombre total qui est sorti en mer ou un indice correspondant. Dans ce cas, on utilisera de préférence une donnée intégrative: la nuit de pêche d'un filet. La prise par filet, en un lieu donné, par nuit, dépendra de la densité des crevettes, de la durée de la pose et de la vitesse du courant. Nous verrons au chapitre concernant les mortalités qu'elle dépend aussi de l'endroit où est placé le filet sur l'axe amont-aval et de la configuration de la lagune.
Les chaluts: S'ils sont tirés par un bateau, la durée de la pêche (quelques heures) sera la meilleure unité. S'ils sont tirés à pied par deux hommes, la sortie sera vraisemblablement la seule donnée utilisable avec les risques que cela comporte. Si la pêche est bien organisée, la situation se rapproche beaucoup de celle de la pêche industrielle et les mêmes méthodes sont utilisables.
Les sennes: Le temps de pêche inclut la recherche de la crevette et la manoeuvre de la senne. L'unité d'effort pourra être la sortie. Cependant, la modification du rapport temps de pêche/temps de recherche au cours de l'existence de la pêcherie peut être une source de biais lorsque l'espèce se raréfie. Il s'agit cependant d'un engin relativement peu utilisé.
Lorsque plusieurs engins sont utilisés dans une même pêcherie, il est pratiquement impossible de standardiser l'effort de pêche si les structures démographiques exploitées diffèrent.
Si les différents engins capturent des structures démographiques identiques, une estimation de l'effort total pourra être obtenue en divisant simplement la prise totale par la cpue d'un engin type dont l'effort est facile à cerner.
L'échantillonnage des débarquements représente un problème de statistique classique que l'on résoudra par les techniques habituelles. L'objectif essentiel de cet échantillonnage est d'estimer la composition, en tailles; des débarquements des différentes espèces (puisque l'âge ne peut être déterminé directement), avec la plus petite variance possible, compte tenu des moyens disponibles. Les propriétés biologiques des deux sexes (croissance en poids et en valeur) étant très différentes, il convient également d'obtenir des données séparées par sexes, pour une modélisation aussi fine que possible des conséquences de la règlementation, quand bien même il est évident que l'on ne pourra pas gérer séparément les deux sexes. De bonnes analyses du problème général et des méthodologies sont proposées par Gulland (1955), Pope (1956), CIEM (1956), et Gulland (1966).
4.2.1 Les structures de taille
On mesure en général la taille de la carapace (du creux post orbitaire au bord médiodorsal postérieur du céphalothorax) ou la longueur totale (de l'extrémité du rostre à l'extrémité des uropodes ou du telson). La première mesure est plus précise et non sensible aux variations d'élasticité de la crevette en fonction de son état de fraîcheur et de son mode de conservation.
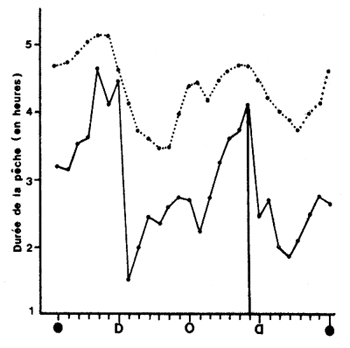
Fig. 27: Durée de la pêche en fonction de la phase lunaire en lagune Ebrié (Côte d'Ivoire) à Ngadykro (—) et Adoukro (…) (d'après Garcia, 1977)
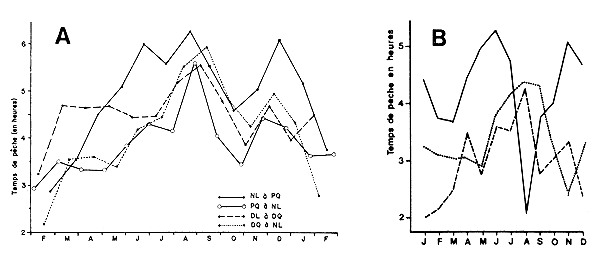
Fig. 28: Variations saisonnières du temps de pêche/nuit en lagune Ebrié (Côte d'Ivoire)
A. En 1974–75 à Adoukro, et pour chaque phase lunaire
B. En 1975 à Adoukro (—), Ngadykro (---) et Ndiemi (…)
(d'après Garcia, 1977)
Si tout ou partie de la flottille débarque des crevettes entières conservées dans la glace pour traitement à terre, l'échantillonnage peut être fait avant passage à la trieuse par les méthodes classiques. Le seul problème est que les glaciers débarquent une cargaison peu importante et vite triée; l'opération d'échantillonnage doit donc être rapide. Il est préférable, la plupart du temps, de tirer parti du fait que les crevettes sont pratiquement toujours conditionnées par calibre. La variance sera donc très sensiblement diminuée si l'on collecte l'ensemble des statistiques de capture par calibre et qu'on les transforme ensuite en structures de tailles classiques. La transformation se fait à l'aide d'une table (cf Tableau I) donnant les distributions de fréquence des tailles dans chaque calibre commercial, distributions obtenues par simple échantillonnage des crevettes classées dans ces calibres au moment du tri. Ces tables permettent de convertir les captures enregistrées dans chaque calibre en captures par classes de taille (centimétriques par exemple). La distribution de fréquence totale des tailles des captures pour la période considérée est obtenue simplement en additionnant les distributions élémentaires correspondant à chaque calibre. Au cas où l'information ainsi obtenue ne couvrirait pas la totalité des captures débarquées une simple élévation de la distribution partielle obtenue à la totalité des captures devra être effectuée.
On peut ainsi obtenir une bonne information à peu de frais. Si il existe un risque que les réglages des trieuses soient modifiés les distributions de fréquence correspondant à chaque calibre devront être vérifiées périodiquement.
Si une partie de la flotille débarque des crevettes étêtées et déjà triées et empaquetées et l'autre des crevettes entières en vrac, on choisira d'échantillonner la fraction permettant l'extrapolation dans les meilleures conditions ou alternativement, les distributions de fréquence seront établies pour les deux types de capture et combinées ensuite. Il faut noter que les crevettes sont souvent triées étêtées. Les échantillons de distribution de fréquence de longueurs obtenus pour chaque calibre concernent donc les longueurs (ou poids) des queues. En déterminant les relations entre la taille (ou poids) de la queue et la longueur totale ou céphalothoracique, il est possible de transformer les distributions de fréquence des queues en ces autres unités. La table de transformation ainsi obtenue est alors utilisée comme précédemment. La mesure de la longueur de la queue n'est cependant pas très précise et cette imprécision introduit un flou dans la distribution reconstituée. Squires (1974) propose de mesurer le poids de la queue, ce qui est certainement plus précis.
Caillouet, Patella et Jackson (1980) proposent un mode de représentation très synthétique de la structure de taille des captures. Si la composition par calibres internationaux est exprimée en pourcentage, la relation entre le loge du pourcentage cumulé et le calibre est une droite. En effet, ils admettent
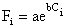
où Fi est le pourcentage cumulé en poids (crevettes étêtées) de la catégorie i,
Ci est la limite inférieure du calibre, par exemple, 21 pour le calibre 21–25 crevettes/livre.
i = rang du calibre,
a et b étant des constantes, la transformation loge permet de linéariser la relation.
L'étude de l'évolution à long terme de la pente (b) de la relation permet de déceler les tendances de la composition en taille et donc également les tendances de la mortalité totale.
4.2.2 Les structures par sexes
Lorsque l'on utilise les statistiques de production des usines, on ne dispose évidemment pas des sexes. Les mâles étant plus petits que les femelles, ils sont plus nombreux dans les petits calibres que dans les gros. A l'intérieur du calibre, le sex-ratio n'est pas uniforme car il varie avec la taille. La relation taille-âge étant différente pour les deux sexes, les structures ne sont pas utilisables telles quelles pour l'étude de la croissance ou de la mortalité.
Tableau I: Clé de conversion des catégories commerciales en longueurs totales pour Penaeus schmitti (d'après Cadima et al., 1972)
| Longueur totale (cm) | FREQUENCE, EN POURCENTAGE, DES LONGUEURS TOTALES DANS CHAQUE CATEGORIE | ||||||||
| 61/70 | 51/60 | 41/50 | 36/40 | 31/35 | 26/30 | 21/25 | 16/20 | 11/15 | |
| 8 | - | - | - | - | - | - | - | - | - |
| 9 | - | - | - | - | - | - | - | - | - |
| 10 | - | - | - | - | - | - | - | - | - |
| 11 | 15 | 12 | 1 | - | - | - | - | - | - |
| 12 | 72 | 43 | 26 | 1 | 4 | 1 | - | - | - |
| 13 | 13 | 38 | 56 | 8 | 9 | 1 | - | - | - |
| 14 | - | 3 | 15 | 24 | 14 | 2 | - | - | - |
| 15 | - | 3 | 1 | 52 | 25 | 21 | 4 | - | - |
| 16 | - | - | - | 14 | 33 | 44 | 33 | 5 | - |
| 17 | - | - | - | 1 | 12 | 26 | 29 | 22 | 2 |
| 18 | - | - | - | - | 2 | 5 | 20 | 43 | 20 |
| 19 | - | - | - | - | 1 | 1 | 4 | 23 | 33 |
| 20 | - | - | - | - | - | - | 1 | 6 | 35 |
| 21 | - | - | - | - | - | - | - | 1 | 9 |
| 22 | - | - | - | - | - | - | - | - | - |
| 23 | - | - | - | - | - | - | - | - | - |
| 24 | - | - | - | - | - | - | - | - | - |
Garcia et Albaret (1977), à la suite des travaux de Wenner (1972), ont proposé de considérer, avec la prudence qui s'impose, la possibilité d'utiliser la relation entre le sex-ratio et la taille pour décomposer à postériori des distributions de fréquences établies sans distinction de sexes, soit que le sexe soit indiscernable de l'extérieur, soit que les données disponibles ne le précisent pas. La relation moyenne obtenue en Côte d'Ivoire (Fig. 30) par des campagnes expérimentales en 1969–70, a été appliquée à la structure en taille des débarquements de Penaeus notialis en avril 1974, à titre d'exemple. La structure globale (mâles + femelles) a été décomposée et ses éléments mâles et femelles, connus auparavant, ont été reconstitués avec une excellente précision (Fig. 31). Il est évident qu'il est nécessaire de vérifier que ce procédé peut être généralisé. Il est vraisemblable que la relation peut être utilisée dans les grands nombres (débarquements mensuels), encore faut-il s'assurer qu'elle ne subit pas de modifications saisonnières par suite de migrations différentielles ou de changements de capturabilité de l'un des sexes. Boerema (comm.pers.) a signalé que lorsque les variations saisonnières du recrutement sont importantes, la relation peut varier d'un mois à l'autre. Marcille (1978) a utilisé la méthode et montré la similitude des courbes obtenues pour P. indicus à plusieurs années d'écart et la différence des courbes obtenues pour des espèces différentes. Lhomme (MS) travaillant sur P. notialis au Sénégal a montré l'existence d'une courbe identique à celle obtenue par Garcia et Albaret, mais nettement décalée, ce qui indique que cette courbe est caractéristique d'un stock et ne doit pas être extrapolée à toute l'espèce.
4.2.3 Données diverses
Parmi les données biologiques qu'il est intéressant de recueillir, on peut noter également la fréquence des crevettes molles et l'état de maturité sexuelle des femelles. La première donnée, facile à obtenir, permet de mettre en évidence les rythmes de mues, phénomènes de première importance dans l'étude de la croissance. La deuxième permet de déterminer le cycle saisonnier de reproduction et sera discutée plus en détail au chapitre suivant.
La disponibilité de statistiques fiables est l'une des conditions essentielles à la mise en place des modalités de gestion rationnelle des ressources. Ces statistiques pourront être obtenues:
si on dispose d'un réseau permanent dans les points de débarquement, à la conception duquel les chercheurs, utilisateurs des données, doivent être associés étroitement,
si on respecte scrupuleusement la règle du secret professionnel et s'il est convenu que les statistiques deviennent anonymes et ne seront pas utilisées à des fins de rétorsions fiscales.
si un minimum d'information en retour est apporté. Ce retour peut se faire de façon informelle par des contacts personnels, ou formelle sous forme de bulletins d'information destinés aux usines, aux commerçants ou aux armateurs (bulletins statistiques, informations sur les marchés),
D'autre part, la complexité et la quantité des données recueillies supposent un traitement automatique de l'information. Ceci implique de disposer d'un équipement de base et de personnel ayant une formation adéquate. Le principe d'organisation de la collecte des données, de leur codage et de leur traitement reste celui qui a été décrit plus haut, mais chaque pêcherie représentant un problème particulier dans un contexte différent, il est difficile de proposer un schéma général. On pourra se référer à quelques travaux déjà cités (Berry, 1967; Snow, 1969; Slack-Smith et Stark, 1969; Garcia, 1978 et Marcille, 1978).
En plus des données concernant les crevettes il est important de prendre en considération les captures accessoires de poisson. Jusqu'à présent dans les pêcheries de crevettes côtières les informations concernant les poissons capturés, conservés ou rejetés ont été le plus souvent négligées. Ceci resulte du prix démesurément plus élevé de la crevette et de la haute spécialisation des unités de pêche. Cet état de fait n'est pas satisfaisant et une approche multispécifique de l'aménagement devant être envisagée, la conséquence directe au niveau de la collecte des statistiques sera la nécessité de mettre l'accent sur des données jusqu'alors négligées telles que les captures et les rejets d'espèces accessoires.
De même, une estimation plus rigoureuse de l'effort exercé sur chaque espèce de crevette - lorsque l'exploitation en concerne plusieurs à la fois - est nécessaire.
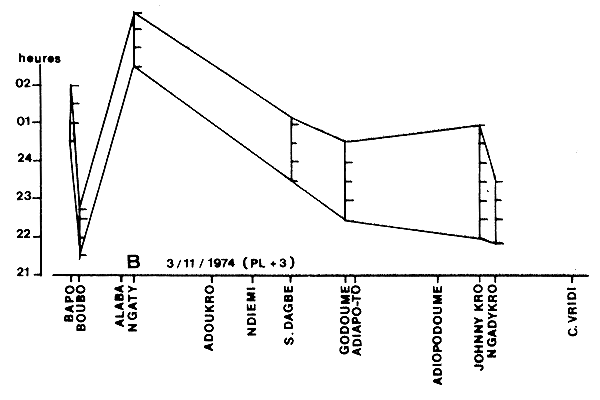
Fig. 29: Variations de l'heure et de la durée de la pêche (en heures) le long de l'axe amont-aval (de Bapo au Canal de Vridi) en lagune Ebrié (d'après Garcia, 1977)
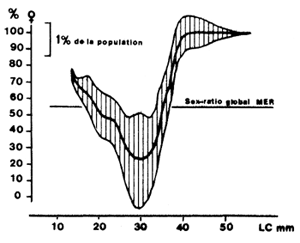
Fig. 30: Relation entre la taille et le sex-ratio chez Penaeus notialis (les lignes verticales sont proportionnelles à l'abondance de chaque classe, en %, dans la population) (d'après Garcia, 1974)
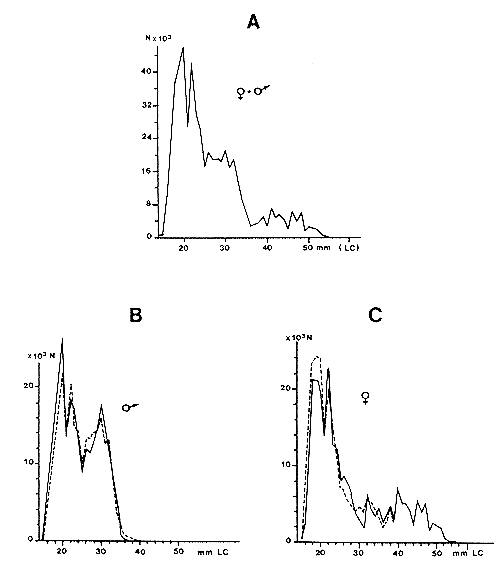
Fig. 31: Comparaison des courbes de fréquence des tailles par sexes, observées et déduites par l'utilisation de la relation sex-ratio/taille chez Penaeus notialis (d'après Garcia et Albaret, 1977)
![]()
![]()
![]()