![]()
![]()
![]()
by
V.A. Shevchenko
Institute of General Genetics
Academy of Sciences
Moscow, U.S.S.R.
We speak of mutation when a character transmitted by the laws of Mendel appears in some hereditary line of an organism which initially was free from this character. The observed mutation reflects a change in an individual gene. Though the mutant gene differs from the original in its chemical composition or structure, it is capable of reproducing itself with great exactness. There is no way of getting rid of mutation, except in rather rare cases, when a subsequent mutation restores the original gene. Mutation is a change in number, type and arrangement of nucleotides which determine the structure of a gene. According to the process of replication of DNA (deoxyribonucleic acid), such changes should repeat themselves further.
Mutations occur both in germinal and in somatic cells, but in sexually reproducing organisms it is only mutations occurring in germinal cells that are transmitted to the progeny. Mutations appearing in somatic cells may give rise to mutant lines only in asexually reproducing organisms.
Mutations occur regularly in all species of living things, though individual gene mutations are very rare (probably one in 100 000 or 1 000 000 cells). But since the number of genes in the majority of species is extremely large, the total rate of mutations in a given generation may appear to be rather high. However, only a small part of new mutations can be detected immediately in the mutant offspring. The greatest part of mutations turn out to be recessive when they appear. The newly-produced recessive gene is transmitted in the heterozygous form throughout many generations (phenotypically the progeny will be normal) until the crossing of such heterozygotes produces a homozygous (phenotypically changed) progeny. It is only recessive mutations linked with sex that are discovered in the first generation of Xy individuals. Even dominant or partially dominant mutations do not always produce a distinguishable mutant offspring, especially when the mutations affect quantitative characters.
Since every species possesses a limited number of genes, and each gene can give but a limited number of mutant alleles, every mutation of the given gene is repeated at a certain low rate, and it is quite probable that in the case of a species with a long evolutionary process, each possible mutation has occurred repeatedly. In examining millions of cells in microorganisms it may be found that the same mutations occur over and over again. This important fact explains why the majority of mutations are deleterious. In a species, which during its long evolution has adapted itself to a certain way of life, all the useful alleles are already included into the genotype, and the majority of individuals of the species carry them in the homozygous state. Once this stage is reached, only deleterious or less valuable alleles can occur as new mutations (e.g., mutations changing the protective coloration).
The term mutation was first suggested by H. de Vries in his classical work “The mutation theory” (1901). He called the phenomenon of a sudden intermittent change in the hereditary character, a mutation. The following basic principles of his theory have not lost their significance up to now:
Mutations occur suddenly, without any transition periods;
New forms are quite constant;
Unlike non-hereditary changes, mutations do not form continuous sets; neither do they group around the medium type (mode); they are qualitative changes;
Mutations act in different directions and may be either beneficial or detrimental;
Detection of mutations depends on the number of individuals analyzed;
Same mutations may occur repeatedly.
It was only after the particulate inheritance of chromosomes was established that the mutation theory became the basis for research. Though the nature of genes has not been clearly elucidated a series of general regularities of gene mutation has been firmly established.
A gene may be defined by its structure and action; mutations may be classified accordingly by the characteristics and properties affected by the gene, that is, by the phenotype and the kind of changes in chromosomes. A genotype is formed in the process of the natural or artificial selection of mutant genes and chromosome reconstructions. Alleles of genes, typical of wild forms of a species, are called wild-type genes, and changed genes are called mutants. There is no principal difference between them (wild-type genes also were mutants formed by natural selection in the process of species evolution).
Since genes determine a successive chain of processes in the development of an organism, i.e. a morphological, physiological and biochemical differentiation of cells and tissues, the classification of mutations by their effect is valuable when studying the regularities of the mechanism of heredity in ontogenesis. Mutations are often defined by the function which is altered by the mutant gene (mutations of sterility and fertility when fecundity is affected). Morphological mutations are those changing the rate of growth and the pattern of the formation of organs in animals and plants (short legs in domestic animals, absence of eyes or wings in insects, absence of hair in mammals, gigantism or dwarfism. (Fig.1). Mutations can affect not only the morphology of an organ, but also its situation; thus, a leg can develop in an insect in place of the antenna. Physiological mutations increase or decrease the viability of organisms. They involve, in particular, semi-lethal and lethal mutations. Biochemical mutations are those inhibiting or changing the synthesis of certain chemical substances in the organism. The carriers of these mutations will not develop into organisms without the introduction of growth factors or the media necessary for their growth. Such mutants are called auxotrophs in contrast to prototrophs or wild-type organisms, which have no special nutritional requirements for their growth. There are mutant genes influencing the rate of mutations at other loci. In all probability the behaviour of chromosomes themselves is controlled by genes.
The first attempt to classify mutations was made by H.J.Muller in 1932. He considered the direction and force of action of a mutant allele as compared to a normal allele (wild type), and thus classified mutations into five types:
Hypomorphous mutations act in the same direction as a wild-type gene, but the effect is somewhat weaker; an increased dose of a hypomorphous allele in the genotype results in the restoration of the wild gene character (thus, when a gene determines the amount of pigment, the accumulation of hypomorphous mutant alleles of this type in the genotype will lead to the restoration of the wild type coloration).
Amorphous mutations are not active as regards the typical effect of the original allele (e.g. genes for albinism which fully inhibit the development of skin pigments).
Antimorphous mutations act in the direction opposite to that of the wild type (in maize, for example, there is a wild type gene causing the anthocyanin coloration, and a gene which hinders the development of anthocyanin pigments, accelerating at the same time the synthesis of brown pigment).
Neomorphous mutations with effects quite different from those of wild-type genes.
Hypermorphous mutations act in the same direction as wild-type genes, but produce a somewhat stronger effect.
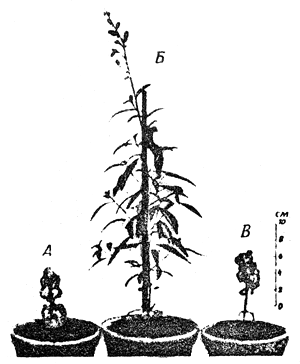 | 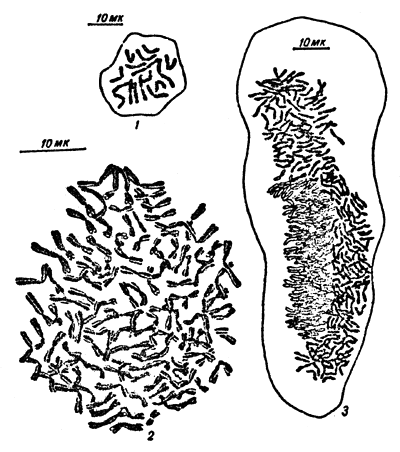 |
Fig.1 An example of gigantism and dwarfism. | Fig.2 Illustration of polyploidy and haploidy, the increase or decrease in the number of complete haploid sets of chromosomes. |
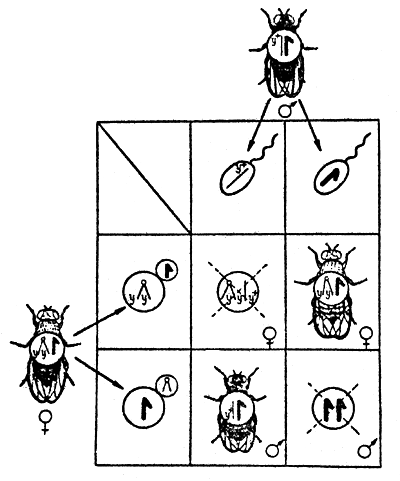 | 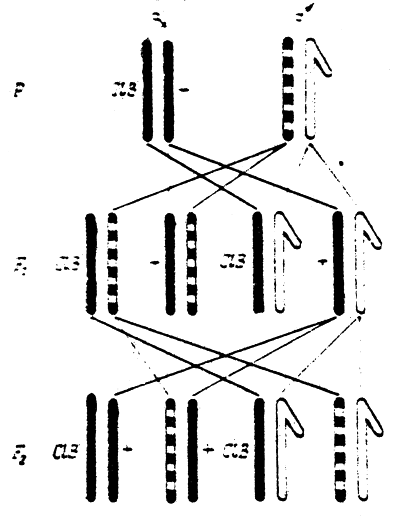 |
Fig.3 Inheritance of characters, linked with sex (body coloration), in Drosophila in case of linkage with the X-chromosome (double-yellow). | Fig.4 The Muller C l B method to count mutations in Drosophila sex chromosomes. |
| Y+ - factor for grey body coloration; y-yellow | |
The study of the action of the mutant gene in ontogenesis is complicated by the fact that its manifestation is determined by the whole system of the genotype - the interaction of genes. One and the same mutation can manifest itself in one genotype and not manifest in another. Moreover, mutations may influence at once a whole set of characteristics, i.e. they may produce a pleiotropic effect.
Almost all changes occurring in chromosomes are referred to as mutations. However, it was established long ago that mutations are variable in their genetics. Practically any change in a chromosome, in which the chromosome and the cell retain the ability to reproduce themselves, determines the hereditary change in the organism's characteristics. Therefore, mutations involve changes in individual genes and chromosome reconstructions, as well as changes in the number of chromosomes. The following genotype classification of mutations is generally accepted:
Gene (or point) mutations - cytologically invisible changes in chromosomes.
Intrachromosome or interchromosome reconstructions:
reconstructions in a chromosome causing changes in the linkage-groups of genes (translocation) or in the sequence of the arrangement of genes within chromosomes (inversion);
fragmentation of chromosomes leading either to the loss of genes or to their doubling (depletion and duplication)
A change in the number of chromosomes:
increase or decrease in the number of complete haploid sets of chromosomes (polyploidy and haploidy) (Fig.2).
change in the number of chromosomes in the diploid set (heteroploidy or aneuploidy).
Mutations arising in sex cells and in embryonic cells are called generative, whereas mutations arising in the cells of other tissues are called somatic. These two types of mutations do not differ in their nature, but as was mentioned above, their evolutionary effect is different and is determined by the type of reproduction of an organism. Generative and somatic mutations manifest themselves differently, and the methods of their detection are also different. Somatic mutations manifest themselves mosaically (the earlier in ontogenesis that they occur, the larger the tissue region which bears them).
Such individuals are called mosaics or chimaeras. Due to the diploid character of the set of chromosomes in the cells of somatic tissue, mutation can manifest itself only when the mutant allele is dominant, or when it is recessive but is in a homozygous state. Somatic mutations do not play any role in the evolution of sexually reproducing organisms, neither are they of any value in selection. But they may be of great significance for asexually reproducing organisms, especially in selection.
Gene mutations in sex cells manifest themselves at the stage of the zygote of the subsequent generations. Dominant mutations are manifest already in the zygote of the first generation, whereas recessive mutations manifest only in the second and subsequent generations when transformed into the homozygous state. Mutation that occurs at the stage of spermatozoa or egg cell formation remains, as a rule single, and the one arising in the cell during the reproduction of spermatogonia or oogonia reproduces itself in proportion to the number of cell divisions that have occurred; as a result a part of the sex cells carry this mutation, and the progeny acquires a bunch of identical mutations. No differences have been found in the frequency of occurrence of somatic and generative mutations.
Gene mutations from the wild type to a new state are called direct, and from the mutant to the wild state is reverse. This reverse process is called gene reversion. Direct mutations are more frequently recessive, whereas reverse mutations are more often dominant. The original gene mutates without any intermediate stages to a new state, and back, correspondingly.
In different genes, direct and reverse mutations may occur with variable frequency:
With equal probability in both directions.
Direct mutations prevail.
Inverse mutations prevail (this is a more rare phenomenon).
Reverse mutations supports the fact that the direct change has resulted not in the loss of the material of inheritance, but only in the change of its state. When the reverse mutation is not detected, it is assumed that the direct change has been caused by the loss of the material of inheritance. The losses can be so minute that it is impossible to detect them in cytological preparations with the help of the light microscope. Sometimes an inverse mutation does not represent a true mutation back to the original gene, but is imitated by the direct mutation of another gene which restores the properties of the original gene (suppressor mutation). That is why any reverse mutation should be analysed genetically before deciding whether it is really inverse. The study of inverse mutations is of principal significance in determining the nature of processes that cause gene mutation.
There are also plasmic mutations - mutational changes in cytoplasmic components of inheritance, which may lead to the appearance of plasmic differences. Plasma mutations have not been studied thoroughly enough.
Theoretically, in the presence of a presumably great number of similar plasmic components, one may expect that the mutation of one of them may become visible only after a selective accumulation of the altered units, the prerequisite for which is the positive selective value of the altered unit - no wonder that plasmic mutations are difficult to detect.
Plastid mutations are spontaneous alterations in the system of plastids, causing changes in plastid characteristics, independent of nuclear genes. Plastids are relatively large cell organellae, and plastid mutations occur not in mature plastids but in embryonic proplastids, changing the genetic centre in them. Typical plastid mutations are subject to the regularities of plastid inheritance.
Registration of mutations, as they arise, is essential for understanding the nature of a gene, its changes, the mechanism of the influence of the surroundings and the physiological condition of the organism on the mutation process.
The difficulty of registration lies in the fact that it is often impossible to distinguish between mutation and recombination. It is practically impossible to objectively register the total number of mutations at present; however, the count of mutations at separate loci and the registration of mutations of a certain type is quite feasible.
Visible dominant mutations that can manifest themselves in the homozygous state in the first generation are most easily detected. Special genetic analysis in a series of generations is required to detect recessive mutations. Recessive mutations should be transferred to the homozygous state.
To make a further analysis, the mutant line is crossed with the analyzer-line, having one or several marked linkage-groups.
For objects with well-known genetics, there are special methods of registering the rate of mutations occurring in individual chromosomes. Thus to detect visible mutations in the Drosophila sex chromosome, a method of X-linked chromosomes - yy (double yellow) - is used. By this scheme (Fig.3) it is possible to detect separate, visible sex-linked recessive mutations which occur in the X-chromosome of the sex-cells in the paternal organism and manifest themselves in males of the first generation.
Recessive lethal mutations, which in the homozygous state lead to the death of individuals bearing them, can be most objectively registered. H.J.Muller has developed the ClB method (Fig.4) to count such mutations in the Drosophila sex chromosome. Another more simple and effective method, called Muller-5, is also used now (Fig.5).
To determine the frequency of lethal mutations in autosomes, a method of balanced lethals has been developed, by which the count of recessive lethal mutations is done in the third generation. It should be noted that lethal mutations by their genetic character are blended types of mutations, embracing various types of chromosome reconstructions as well as changes in individual genes.
A mutation process is characterized by the frequency of mutation occurrence and the direction of gene mutation. The frequency of mutation occurrence is one of the determining factors for every species of animals, plants and micro-organisms: some species are characterized by a higher mutation variability than others. These variations are caused by many factors of general and private significance: a species genotypic structure, a degree of its adaptation to the environment, the place of its distribution, the force of action of natural factors, etc. No matter how well protected from the environmental effects an organism may be, chemical processes relating to its metabolism may cause spontaneous mutation variability.
The causes of spontaneous gene mutations have not been elucidated. A number of factors, which are usually divided into external (in relation to the organism) and internal, underlie the phenomenon of spontaneous (natural) mutations. External factors include the natural radioactive background, the influence of temperature, which may be direct (on the genetic material), or indirect (through metabolic processes). Internal factors include genotypically controlled physiological factors, and chemical factors arising in metabolic processes. Many researchers hold that the main cause of the appearance of spontaneous mutations is cell metabolism. In all probability, mutations may also be caused, in part, by the physical processes in the genetic material (e.g. thermal movements of atoms may lead to the formation of isomeric molecules), or because of changes in the normal process of duplication of DNA.
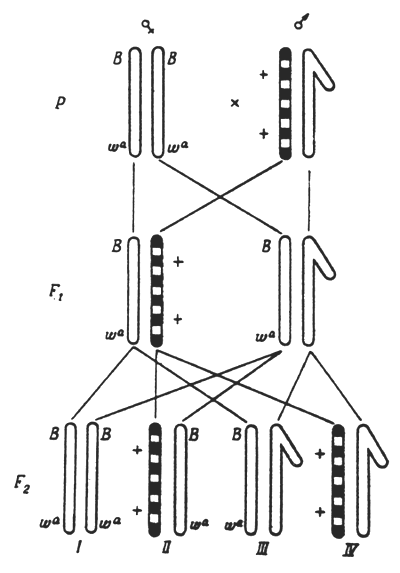 | 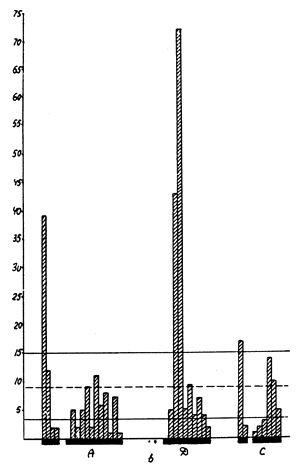 |
Fig.5 The Muller - 5, a simpler method to count mutations in Drosophila sex chromosomes. | |
 | |
Fig.6 Varieties of grain heads produced by irradiation. | Fig.7 Arrangement of chromosome breaks under the influence of ethylenimine on sprouts of Crepis capillaris. |
As has been mentioned the frequency of mutation occurrence within one generation is not quite known. This is accounted for by the fact that mutations are exceedingly diverse, in their phenotypical manifestation, and in their genetic causes, whereas the methods of registering them are imperfect. And only as regards the mutability of individual loci can a more or less accurate estimate be made.
The established general regularities of the frequency of spontaneous mutations can be summed up as follows:
Different genes in the same genotype mutate with different frequency;
Identical genes in different genotype mutate with different frequency.
Thus there are mutable and stable genes. For example, a certain gene in maize mutates in about 50 cases out of 10 000, and sh-gene (shrunken endosperm) only in 0.1 whereas in various maize lines the same gene can mutate with the frequency of 6.2 (Columbian line) and 18.2 (Cornwell line) per 10 000 gametes. By selection, lines with different spontaneous mutability can be created.
The fact that the spontaneous mutation process is caused by the physiological condition and biochemical changes in cells is confirmed, for example, by instances of more frequent lethal mutations in Drosophila when the sperm is kept in the seminal receptacle, and by cases, of more frequent gene mutations and chromosome reconstructions in seeds when stored for a number of years.
At present it may be stated with good reason that in related species of common origin there occurs similar mutations. Moreover, we come across parallelism even in individuals of different classes and types of animals: homologous sets of mutations by morphological, physiological and biochemical characters and properties (the law of homologous sets by N.I.Vavilov).
Today the investigation of mutations is carried out almost exclusively on experimentally induced mutations. A whole number of factors can cause hereditary change - temperature, ultraviolet rays, various types of ionizing radiation and a great many chemical substances.
The first mutations induced by X-rays were made by H.J.Muller in Drosophila and by L.Stadler in maize and barley. Later it was shown that X-rays cause mutations in a number of plants and animals as well as fungi, bacteria and viruses. Already at the early stages of these investigations, it was evident that the frequency of mutations is proportional to the irradiation doze measured in röntgens. A definite number of röntgens may be passed on to the object within a short period of time (at a high irradiation intensity) or within a longer period (at a low irradiation intensity). It was found that the same number of röntgens produced, at an average, the same number of mutations, irrespective of the irradiation intensity. The probability that the given gene mutates, when affected by one röntgen, turned out to be equal to approximately 10-8, and was almost the same for such different organisms as Drosophila, mice, bacteria and viruses. Hence, it follows that there is a linear dependance between the irradiation dose and the frequency of mutations. It should be noted that sometimes this linear dependance is disturbed, as for example, when irradiating seeds. Because genetic alterations occur only in a part of the seed cells, and the mutant tissues are often weakened as compared to the normal ones, the frequency of detected mutations decreases.
Early research showed also that, along with the induction of gene mutations, irradiation caused many structural alterations of chromosomes - translocations, inversions, loses and duplications. All these structural changes are associated with chromosome breaks under the influence of irradiation, as a result of ionization produced by irradiation; electrons are liberated from some atoms and join others, forming positive or negative ions. The formation of ions is the beginning in the complex chain of physico-chemical processes resulting, in the long run, in the reconstruction of molecules and changes in the various biochemical processes, which may lead to gene mutations and breaks of chromosomes.
Different types of ionizing irradiation: gamma-rays, X-rays, protons, neutrons of various energies and α -particles are widely used at present. They all cause ionization in the tissue. However, in spite of the fact that the final result is similar, there are essential differences in the character of ionization caused by various primary particles. These differences involve the unequal density of ionization (the number of ion pairs formed per unit of volume), and an unequal distribution of ionization throughout the thickness of the irradiated substance. The effect of light particles (electrons, protons) creates ionization of low density in the object, dispersed more or less equally throughout the irradiated substance. Heavy loaded particles (protons, α - particles) produce, on the contrary, ionization with a very high local density.
The irradiation effect depends not only on its own quality, but also on the state of the chromosomes during irradiation. It has been established that during mitosis, chromosomes at the stages of prophase and metaphase are damaged more readily than chromosomes at the stage of interphase. If chromosomes are irradiated in the interphase, the type of chromosome reconstructions obtained in mitosis depends on the period (before the reduplication of chromosomes or after it) when the irradiation produces its effect. In the pre-synthetic stage of the interphase (g1) the reconstructions that occur are of the chromosome type, and in the synthetic stage (S) and post-synthetic one (g2), of a chromatid type.
The g2 stage being much more sensitive to irradiation than g1 (the stage of synthesis usually occupies an intermediate position as regards sensitivity). Meiosis stages, as a rule, are more sensitive to irradiation than mitosis stages.
Rather recently it was found that the relation between irradiation and the mutation process, in all probability, is not as direct as it was previously thought to be. The irradiation energy is evidently absorbed not only within chromosomes, but by their environment as well. This can cause chemical changes, which in their turn, lead to gene mutations or the breakage of chromosomes. A great role in the indirect action of ionizing irradiation is played by hydrogen peroxide and free radicals (e.g. water can form OH and HO2 radicals which are highly reactive, and therefore can split nucleic acids and other organic substances).
The efficiency of ionizing irradiation in causing mutations and chromosomes reconstructions is to a certain extent controlled by the genotype. It has been established that the radio resistance and the ability to mutate under the influence of ionization are different in organisms of different genotypes, in different forms, species and genera. The genotype plays an important role in controlling the primary and secondary radiation effects.
The length of a single gene and its chemical composition can well reveal the frequency of occurrence of mutant alleles in it. It has been shown that different genes mutate at different frequencies under a uniform radiation dose. Even the movement of the gene from one area of the chromosome to another is accompanied by the alteration of its mutability. Moreover, genes, in all probability, determine the relatively different chemical composition of the cell nucleus, the amount of water and oxygen in the cell, the colloid state of the karyoplasm, etc., that is, the genotype determines the nature of the secondary processes during irradiation.
The achievements of radiation genetics are widely used nowadays in the radiation selection of plants, animals and micro-organisms. Radiation selection began to develop especially rapidly after the erroneous views - that radiation only upsets and damages heredity and that it may lead only to malformations had been discarded. Hundreds and thousands of new forms have been produced by radiation selection. For instance, new radiation varieties of a series of agricultural plants have been on sale in Sweden for a long time (Fig.6).
Ultraviolet rays (u v rays) do not cause ionization; they can only change the cell electrons from a lower orbit to a higher level. Though u v rays have a lower penetrating power than ionizing radiation, they are still capable of penetrating small objects (bacteria, spores of fungi, plant pollen) causing rather frequent mutations in them. Especially powerful are u v rays with the wave-length of about 2600A, which are absorbed by DNA. U v rays produce a milder effect on chromosomes than X-rays; and it was believed previously that u v rays cause only true gene mutations, not associated with the structural changes in chromosomes. However, to-day it is established that u v rays can also induce structural changes, so that the difference between X-rays and u v rays refers rather to the degree than to the essence of the mutation effect. U v rays are widely applied in the radiation selection of micro-organisms (producers of antibiotics), contributing not only to a radical qualitative rise in the activity of strains, but also to some qualitative changes. Thus, the production of oxytetracycline was complicated by some unfavourable properties of the producer strain. With the help of u v rays, mutants were produced that increased the productivity and eliminated the defects in the strain.
By now, several hundreds of various chemical substances have been discovered that can induce changes in the hereditary material. To be an active mutagen, a substance should easily penetrate the cells of an organism (which retains its viability) reach the nucleus of the cell and influence the chemical processes in it.
Chemical mutagens may be divided into four big groups. The first group includes highly active chemical substances, which can transfer alkyl groups to other molecules. These substances are called alkylizing agents. The most active mutagens (yperite, ethylenimine, ethylmethanesulphonate) belong to this group. The second group includes peroxides. It is free radicals (OH, H, HO2) that are active in this group. That is why all the factors contributing to the formation of free radicals (oxygen, water, u v rays and others) intensify the mutagenic effect of peroxides. The mechanism of action of the substances, that belong to the third group of metabolite-analogues, consists in their substituting normal metabolites during metabolic processes in the cell. To this group belong, for example, various derivatives of purine and pyrimidine bases (bromine and fluorinouracil), aminopurine, aminopterine and others. The fourth group includes substances whose mechanism of action is not yet clear: these are different mineral salts, some dyes and alkaloids.
The substances belonging to the first group are often called radiomimetics. In general, the effect they produce is the same as that of X-rays, but there are two essential differences. The first is that radiomimetics are capable of producing a distant effect. As a result of this effect, for several generations after the mutagen has acted, individuals appear which represent mosaics from normal and changed cells. There are different explanations of this effect and its mechanism is being studied currently. The second difference lies in the fact that usually, under the influence of radiation, structural changes in chromosomes are more numerous than gene mutations, whereas the opposite is the case when chemical substances are used.
Chemical mutagens are capable of exerting not only a direct, but also an indirect influence on the hereditary material. There is evidence, testifying to the presence of anti-mutagens in organisms, which prevent, or, at least, counteract the mutagenic effect of a number of substances normally taking part in metabolic processes. The catalase enzyme serves as a natural anti-mutagen. The catalase immediately stops the mutagenic action of hydrogen peroxide; and it is probable that the failure in attempts to produce mutations in some organisms with the help of hydrogen peroxide is due to a high content of this enzyme in the cells of these organisms.
One of the basic problems of modern genetics and selection is the problem of producing directed mutations. To solve this problem it is necessary to study the specificity of the action of various mutagenic factors. The problem is being studied at different levels (molecular, gene and chromosome). The means of direct chemical influence on the molecular structure of DNA leading to qualitatively definite alterations have already been found. As is known, the structure of all genes is somewhat monotonous, since they are formed only of four nucleotides arranged in a different sequence. Due to this, their reaction to mutagenic factors must also be rather monotonous; i.e. the action of mutagens, as it seems, should not always be directed. However, every chemical mutagen possesses definite chemical properties, typical only of it, so that it may be assumed that it can induce not just any change, but definite changes in pairs of bases. With this chemical characteristic should be associated the mutagenic characteristic - the capability of inducing not any, but definite mutations.
The reasons for the selective effect of mutagens on definite loci may also depend on the gene material itself: e.g. the given gene may contain a relatively greater amount of the nucleotide which is preferably influenced by the mutagen used; and, consequently, this gene will mutate more frequently. The nucleotide which is mainly affected by the mutagen, may, within one gene, be part of a triplet, determining the synthesis of the amino-acid found in the active centre of the protein, and within another gene the synthesis of a not very essential amino-acid in the absence of which the protein will be active all the same.
Using alkylizing compounds, oxidizers and reducing agents, purine and pyrimidine derivatives, we cannot, as yet, induce directed mutations in individual genes; we can only speak of the group specificity, i.e. of specific changes in the spectrum of mutations.
The analysis of the spontaneous mutation in a number of organisms has shown some specific pattern in the distribution of the frequency of mutations, both between different genes and within genes. Thus, a clearly outlined intra-genic specificity has been found within the locus r,II of the phage T-4, and in the thymine locus Entamoeba coli, K-12. In the X-chromosome of Drosophila were found points, where many spontaneous, visible and lethal mutations were being accumulated, and there were also many empty places, where no mutations were observed.
The gene specificity drew the attention of researchers already at the early stage of investigations on radiation mutagenesis. It was shown that different genes of Drosophila react differently to X-rays. The mutation frequency of some increases considerably, of others, only slightly or not at all, as compared to the mutation frequency in spontaneous mutagenesis. Many papers were published in 1950's – 60's which demonstrated clearly the presence of specificity in the mutation of different loci and in the spectrum of mutations in barley when influenced by various kinds of radiation.
In general, it can be said that the neutron radiation produced more mutations and a wider spectrum of them than the rarely-ionizing radiation. The gene and intra-genic specificity was also found in some micro organisms (E. coli, Streptomyces) as a reaction to the effect of various kinds of radiation. It was proved that the spectrum of mutations can change not only due to the kind of radiation, but also due to the radiation conditions and the dose of radiation applied.
The different reaction of the hereditary material when influenced by chemical mutagens is often expressed by the ratio: gene mutations/chromosome reconstructions. The ratio changes sharply depending on the application of various mutagens and determines the specific character of their effect. The change in the ratio, gene mutations/chromosome reconstructions, essentially contributes to change in the spectrum of mutations, as definite types of mutation (e.g., erectoids in barley and wheat) are often associated with chromosome reconstruction.
It is of interest to note that out of chemical mutagens it was only oxygen that produced a mutagenic effect under high pressure, which by its spectrum did not differ from the effect produced by radiation.
It should be mentioned that the study of the specificity in the reaction of definite loci to various mutagens is hindered by a series of modifying factors. For example, in plants the specificity of the effect of chemical mutagens can depend on the kind of plant, the mutagen dose, the length of its action and the conditions of cultivation of the first and subsequent generations.
In recent years the study of specificity of chemical mutagens on micro-organisms (bacteria, phages, yeast, actinomycetes, neurospore and others) has been especially intensive. S.Benzer and E.Freese made a detailed map of mutations, induced in the phage T-4 in the locus r II by the analogues of purines and pyrimidines, nitrous acid and ethylmethanesulphonate. Each of the tested mutagens gave its own typical distribution of hot spots on the gene map. Freese believes that since the occurrence of almost all induced mutants in the locus r II is associated with the substitution of the single pair of nucleotides in DNA, the heightened specificity of the mutagen effect may be also accounted for (except for the essential differences between both pairs of bases in relation to some mutagens) by the influence of some structural factors: e.g. bases adjacent to the given base may intensify its trend for mutations; or a high specificity will characterize mutagens interacting simultaneously with one or more bases.
Specific action of various mutagens analyzed at the chromosome level may manifest itself as follows:
in the ability of mutagens to cause various correlations in the frequency of certain types of chromosome reconstructions, as well as in the characteristic distribution of various types of reconstructions in individual cells of the population treated with the mutagen;
in the distribution of chromosome breaks among different cells of the tissue irradiated or treated with chemical mutagens (at random distribution the number of cells with 1,2,3 n breaks, should correspond to the values estimated by the Poisson formula);
in the distribution of breaks among different chromosomes or clearly-distinguished groups of chromosomes within one cell (at random distribution, the relation of the frequency of breaks in separate chromosomes or groups of chromosomes will be proportional to their lengths);
in the distribution of breaks along the length of separate chromosomes (at random distribution, the breaks should be positioned evenly along the length of the chromosome arms).
At present, numerous data are available on the distribution of chromosome breaks under various types of radiation, and, especially, when influenced by different chemical mutagens. The following factors can affect the localization of breaks in chromosomes.
Spatial arrangement of chromosomes in the nucleus;
Size and position of the nucleolus;
Centromere and telomere;
Pairing of chromosomes (in meiosis);
Presence and arrangement of heterochromatic regions.
An especially strongly pronounced specificity of action, at the chromosome level, characterized a number of alkylizing compounds, e.g. in Crepis capillaris, under the influence of ethylenimine, more than one third of all the breaks observed were concentrated at a small site of D-chromosome (during radiation the frequency of breaks at this site was casual (Fig.7). Unfortunately, there are no data, as yet, explaining why certain sites of chromosomes are so highly sensitive to some mutagens. It is probable that the reason lies in the unequal distribution of pairs of bases along the length of chromosomes, or in the protein component, which is different at different sites of chromsomes, or in the fine structure of chromosomes themselves. It is quite possible that methods will appear which will enable the scientists to answer these questions. Further research into the specificity of the effect of various mutagens, at the levels of molecules, genes and chromosomes, and the elucidation of reasons underlying this specificity will bring the scientists nearer to the problem of producing directed mutations.
Benzer, S. and E. Freese, 1958 Induction of specific mutation with 5-bronouracil. Proc.nat.Acad. Sci., Wash., 44:112–9
de Vries, H., 1901 Die Mutationstheorie. Veit, Leipzig
Freese, E., 1959 The difference between spontaneous and base-analogue induced mutations of phage T4. Proc.nat.Acad.Sci., Wash., 45:622–33
Muller, H.J., 1932 Further studies on the nature and causes of gene mutations. Proc.6th int. Congr. Genet., 1:213–55
Stadler, L.J., 1928a Genetic effects of x-rays in maize. Proc.nat.Acad.Sci., Wash., 14:69
Stadler, L.J., 1928b Mutations in barley induced by x-rays and radium. Science, 68:186–7
Vavilov, N.I., 1922 The law of homologous series in variation. J.Genet., 12:47–89
![]()
![]()
![]()