![]()
![]()
![]()
Les insectes sont les seuls invertébrés connus comme se nourrissant des fruits et des graines de Prosopis (Kingsolver et al. 1977). Il peut en résulter une destruction totale des fruits (avec les lépidoptères, par exemple), mais le résultat le plus fréquent est l'avortement des fruits ou des graines, ou la destruction des graines. Nous considérons ici les insectes comme ravageurs en ce sens qu'ils détruisent des organes entiers tels que les semences, mais dans la mesure où les dégâts qu'ils causent n'empêchent pas la semence de germer nous pourrions aussi bien les désigner sous le terme de simples parasites.
Pour des raisons de commodité, nous suivrons Kingsolver et al. 1977) qui divisent les insectes se nourrissant des fruits et des graines de Prosopis en deux groupes, ceux qui les attaquent principalement de l'extérieur, et ceux qui les dévorent par l'intérieur. Les premiers sont des adultes et nymphes d'hémiptères, et des larves de lépidoptères. Les seconds comprennent des larves de lépidoptères, et des coléoptères appartenant aux familles des Curculionidae, Cerambycidae et Bruchidae. Parmi les coléoptères, les bruchidés sont de loin les insectes nuisibles les plus importants.
Dans une certaine mesure, l'âge du fruit influe sur le mode d'alimentation des insectes. Les hémiptères ont des pièces buccales perceuses et suceuses, de sorte qu'ils s'attaquent volontiers aux fruits de Prosopis immatures et tendres, tandis que les gousses sèches et lignifiées ne les attirent pas, et sont délaissées par eux. Les larves de lépidoptères se nourrissant de l'extérieur s'en prennent également aux gousses immatures et tendres. L'âge de la gousse, en revanche, a moins d'importance pour la plupart des parasites internes, parce que la plupart se nourrissent des graines, et que celles-ci sont souvent tendres. Parvenir à pénétrer dans la gousse et dans la graine est pour eux l'étape la plus critique, du moins pour ceux qui ont été le plus étudiés, c'est-à-dire les bruchidés. Tous les parasites internes connus le sont au stade larvaire; on pense que les adultes se nourrissent de nectar et de pollen, quoique certains bruchidés ne s'alimentent pas à l'état adulte.
Les hémiptères sont d'importants parasites indigènes des mesquites. Selon Ueckert (1973) la “punaise à pieds foliacés” Mozena obtusa Uhler (Coreidae) détruit de grandes quantités de gousses de Prosopis glandulosa en Amérique du Nord. Au Texas cet insecte entre en activité au début du printemps, et peut rester actif jusqu'en août. Swenson (1969) a estimé ses dégâts sur les mesquites au Texas à 50 pour cent de la production potentielle de fruits. C'est l'hémiptère que l'on trouve le plus fréquemment sur les jeunes fruits de Prosopis velutina à Tucson en Arizona (Werner et Butler 1958). Ueckert (1973) a constaté que la proportion de gousses immatures de P. glandulosa avortées variait entre 33 et 89 pour cent lorsqu'elles avaient été attaquées par cette punaise. En outre, les attaques de punaises se traduisent par une diminution de poids des gousses sèches de 23 à 59 pour cent, et une baisse du taux de germination des graines, qui n'était plus que de 4,3 pour cent au lieu d'un taux normal de 65 pour cent. Etant donné que les mesquites se reproduisent uniquement par graines, l'intervention de cet ennemi naturel peut réduire de manière importante la propagation de l'espèce. Un autre hémiptère, Chlorochroa ligata (Say), connu sous le nom de “conchuela” (Pentatomidae), détruit les graines de mesquite au Texas en en aspirant le suc, ne laissant à l'intérieur de la gousse que leurs téguments desséchés (Smith et Ueckert 1974). Des gousses placées dans des cages témoins donnèrent des graines avec un pourcentage de germination de 76 pour cent, bien supérieur à celui des graines développées dans des cages renfermant des conchuelas (0 à 4 pour cent de semences viables). Dans une étude effectuée en Argentine, sur 145 jeunes fruits de Prosopis flexuosa seulement 40 atteignirent la maturité, et avec P. chilensis seulement 4 sur 70 (Solbrig et Cantino 1975). Selon Kingsolver et al. (1977), il est possible que des dégâts d'insectes comparables à ceux de Mozena obtusa en Amérique du Nord aient été responsables pour une large part de cet avortement de fruits en Argentine, mais il se peut également que d'autres facteurs, tels qu'un déficit en eau ou un avortement spontané aient contribué à cette mortalité.
La plupart des larves de lépidoptères parasites externes consomment les fleurs, les fruits immatures et les gousses en cours de maturation des Prosopis (Kingsolver et al. 1977). Un certain nombre d'espèces de lépidoptères se nourrissent à l'intérieur des gousses de Prosopis en Amérique du Nord. Des membres des familles des Lycaenidae (Strymon leda), Olethreutidae (Ofatulena spp.), Pyralidae (Paramyelois spp.), Blastodacnidae (Chaetocampa sp.), Notodontidae (Didigua argentilinea) et Cochylidae (Phalonia leguminana) consomment l'intérieur des gousses aussi bien que des graines. La teigne du blé (Pyralidae: Plodia interpunctella (Hubner)) a été signalée comme se multipliant en grand nombre à Hawaï dans des gousses de mesquite entreposées (Bridwell 1920b). Les oeufs de ce papillon sont pondus sur les sacs dans lesquels les gousses sont entreposées, ou sur les gousses ellesmêmes, et les jeunes chenilles pénètrent ensuite dans les gousses, en choisissant de préférence des gousses cassées dans lesquelles elles mangent la pulpe sucrée. Lorsque les chenilles arrivent à maturité, elles quittent les gousses et cherchent un endroit abrité pour faire leur chrysalide. Selon Koch et Campos (1978), Cryptophlebia carpophagoides Clarke (Lepidoptera: Olethreutidae) se nourrit des gousses d'Enterolobium contortisiliquum Vell. en Argentine, et de Prosopis tamarugo et P. juliflora au Chili. Les chenilles de cette espèce consomment à l'intérieur des gousses les graines et les tissus tendres. Elles laissent fréquemment leurs excréments sur l'extérieur du fruit. Ces auteurs attribuent à ce lépidoptère des pertes atteignant 30 pour cent dans les gousses de P. tamarugo. Habit et al. (1981) décrivent le cycle biologique et la structure de cette “teigne du fruit” au Chili, et mentionnent qu'elle avait endommagé 17,4 pour cent des fruits de P. tamarugo. Koch et Campos (1978) et Habit et al. (1981) indiquent que Leptotes trigemmatus Butler (Lepidoptera) se nourrit également des fruits de P. tamarugo au Chili. Cette espèce s'attaque de préférence aux fruits à leurs premiers stades de développement, ce qui en entraîne finalement la chute.
En Arizona trois espèces de Curculionidae (Apion subornatum, A. ventricosum, Microtychius sp.) se nourrissent des gousses de Prosopis velutina (Kingsolver et al. 1977). En Argentine les charançons Sibinia asulcifera et S. mastuerzo se nourrissent probablement de graines de Prosopis (Clark 1979). On sait que des coléoptères appartenant au genre Lophopoeum (Cerambycidae) se développent à l'intérieur des gousses de Prosopis en Argentine, en en consommant toutes les graines (Kingsolver et al. 1977).
Les Bruchidae sont de loin les insectes les plus nombreux (à la fois par le nombre d'espèces et par le nombre d'individus), et les mieux connus, qui se nourrissent des fruits de Prosopis dans le Nouveau Monde (Kingsolver et al. 1977). Des larves de bruchidés ont été observées dans les graines de plantes appartenant à 32 familles différentes (Johnson 1981a), mais la plupart des espèces s'attaquent à l'une de ces familles, celle des légumineuses. Les Bruchidae sont une famille très voisine de celles des Chrysomelidae, mais alos que les représentants de ces deux dernières se nourrissent de diverses parties de la plante, les bruchidés ne s'attaquent qu'aux graines. Cette famille comprend à l'heure actuelle environ 1300 espèces, groupées en 56 genres et en 6 sous-familles: Amblycerinae, Bruchinae, Eubaptinae, Kytorhininae, Pachymerinae, Rhaebinae (Johnson 1981a). La plupart des espèces de bruchidés (80 pour cent) sont actuellement rangées dans la sous-famille des Bruchinae.
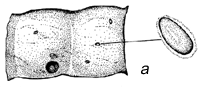
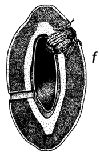
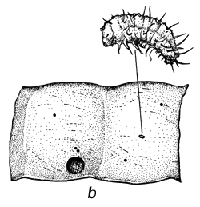
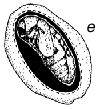
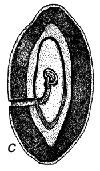
Fig. 1 - Schéma du cycle biologique des bruchidés sur les gousses de Prosopis. a) Oeufs collés à la surface de la gousse ou déposés dans des fissures de celles-ci ou dans les trous de sortie (circulaires) d'insectes adultes. b) Trou d'entrée de la larve au premier stade de développement, percé à travers la paroi de la gousse, et image agrandie de cette larve montrant les poils, les piquants et les pattes qui sont une adaptation lui permettant de pénétrer dans la graine. c) Coupe transversale d'une gousse et d'une graine montrant la galerie creusée par la jeune larve lors de sa pénétration. d) Larve à un stade de développement plus avancé, à l'intérieur de la cavité creusée dans la graine. e) Nymphe à l'intérieur de la chambre larvaire. f) Adulte sortant par le trou préparé par la larve au dernier stade.
 | |
 |  |
 |  |
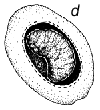 | |
La Figure 1 illustre un cycle biologique typique de bruchidé, mais ces insectes attaquent les graines de manières diverses (voir plus bas). Généralement, la femelle adulte pond ses oeufs sur une graine ou sur une gousse (Figure 1), et la larve à son premier stade de développement perce avec ses mandibules la coque de l'oeuf, la paroi de la gousse (s'il y a lieu), le tégument de la graine, et pénètre à l'intérieur de celle-ci. Elle est fortement modifiée à ce stade (Figure 1) pour pouvoir pénétrer dans la graine, et possède à cet effet de nombreux piquants, poils, etc. (Pfaffenberger et Johnson 1976). Peu après être entrée, elle se mue en un ver apode qui est très différent du premier stade larvaire, et qui est adapté pour se nourrir à l'intérieur de la graine. La larve s'attaque habituellement à une seule graine, mais chez certains bruchidés à deux ou plusieurs graines, et elle se mue en général encore trois fois au cours de sa croissance. Elle fait ensuite sa nymphe en général dans une graine unique, mais certaines espèces quittent la graine et font leur nymphe dans un cocon, tandis que les larves d'autres espèces agglutinent plusieurs graines ensemble en une chambre nymphale. Après la nymphose, l'insecte adulte termine l'ouverture d'un trou de sortie caractéristique (Figure 1), qui avait déjà été fait presque entièrement par la larve, et quitte la graine pour recommencer un nouveau cycle. La durée de ce cycle biologique est variable, mais est généralement d'une trentaine de jours. Les bruchidés adultes se nourrissent probablement de nectar et de pollen, et on ne les voit jamais s'attaquer aux graines, si ce n'est incidemment, par exemple lorsqu'ils sortent d'une graine ou lorsqu'une femelle perce un trou avec ses mandibules dans une gousse pour y déposer ses oeufs. Certaines espèces de bruchidés, notamment celles qui sont économiquement importantes, survivent pendant de nombreuses générations dans des récipients emplis de semences en laboratoire ou en magasin, sans que les adultes se nourrissent.
On connaît à l'heure actuelle de nombreuses plantes pouvant héberger ces insectes (Johnson 1981a), ce qui ne veut pas dire qu'on sache beaucoup de choses sur les hôtes des bruchidés. En fait, de récentes études de terrain amènent l'auteur du présent manuel à penser que ces insectes se nourrissent des graines d'un bien plus grand nombre de plantes, notamment de légumineuses néotropicales, qu'on ne l'a signalé jusqu'à présent.
On connaît 5 genres de bruchidés se nourrissant uniquement des graines de Prosopis spp. (Tableau 1 page 40), mais 5 autres genres s'attaquent également aux graines de plantes se rencontrant en association étroite avec les Prosopis, telles qu'Acacia, Cercicium et Parkinsonia. Dans le Nouveau Monde, environ 32 espèces de bruchidés se nourrissent des fruits de Prosopis. Sur ce nombre, 28 sont obligatoirement limités aux Prosopis, mais certains bruchidés peuvent s'attaquer à plusieurs espèces de Prosopis. Le Tableau 2(page 42) donne une liste des bruchidés et de leurs plantes hôtes, et le Tableau 3 (page 46) présente les différentes espèces de Prosopis avec les bruchidés qui s'attaquent à leurs graines.
Des représentants des genres Algarobius, Neltumius et Mimosestes sont les principaux bruchidés parasitant les graines de Prosopis depuis les Etats-Unis jusqu'au Venezuela (Tableau 1). Toutes les espèces de ces 3 genres qui s'attaquent aux Prosopis sont obligatoirement limitées à ce genre botanique, à l'exception de Mimosestes amicus et M. insularis. Les autres hôtes de ces deux dernières espèces sont indiqués au Tableau 2 (page 42).
Les études les plus poussées effectuées sur le cycle biologique et les dégâts pouvant être causés aux semences de Prosopis concernent Algarobius prosopis, Mimosestes amicus, M. protractus et Neltumius arizonensis (Swier 1974, Kingsolver et al. 1977, Conway 1980), c'est pourquoi nous passerons en revue ci-dessous les informations disponibles sur ces 4 espèces.
Cette espèce est de loin la plus abondante dans les graines de Prosopis nord-américains, et elle est donc susceptible de causer d'importants dégâts aux semences de mesquite. Selon Swier (1974), elle hiverne probablement à l'état immature ou adulte dans les graines de Prosopis velutina en Arizona. Les femelles adultes sortent au printemps et pondent leurs oeufs sur ce qui reste de la fructification de l'année précédente. Lorsque de jeunes gousses apparaissent sur les arbres vers la fin du printemps (mai-juin), les femelles y déposent leurs oeufs, et elles continuent à pondre sur les gousses en cours de maturation ou arrivées à maturité jusqu'à la fin de l'automne. La femelle d'A. prosopis n'agglutine pas ses oeufs aux gousses ou aux graines comme le font de nombreux bruchidés, mais les introduit dans des fentes et crevasses de la gousse (Bridwell 1920a, Swier 1974). Les trous de sortie des adultes sont souvent utilisés par cette espèce comme emplacement de ponte. Les larves d'A. prosopis sont très mobiles, possédant des pattes et des soies sensorielles bien développées (Pfaffenberger et Johnson 1976), ce qui est en corrélation avec une forte activité locomotrice. A l'intérieur des gousses très jeunes, elles se nourrissent probablement du sirop qu'elles renferment, jusqu'à ce que les cotylédons se développent (Bridwell 1920b). Sur des gousses à un stade plus avancé de développement, les larves pénètrent dans la première graine qu'elles rencontrent, ou bien cheminent à travers la pulpe de la gousse pour pénétrer dans une autre graine. En magasin, les larves peuvent percer un trou dans le tégument de la graine depuis la surface de la gousse. Après s'être nourries et avoir mué environ 3 fois à l'intérieur d'une même graine, les larves tapissent les parois de la chambre qu'elles ont creusée avec des déchets, et ensuite se transforment en nymphe.
Dans les conditions naturelles, Algarobius prosopis a 3 générations ou plus par an. Selon Bridwell (1920b), cette espèce ne se reproduit pas dans les gousses sèches de mesquite entreposées, mais continue à se multiplier si les gousses sont encore fraîches. Swier (1974) a observé des larves de cette espèce pénétrant dans des gousses sèches. Il faut encore d'autres recherches, avec des gousses sèches et des gousses fraîches, pour déterminer dans quelle mesure A. prosopis peut détruire les semences entreposées.
Dans la nature, pour une fructification donnée sur un arbre, cet insecte peut détruire entre 8 et 75 pour cent des semences (Swier 1974, Glendening et Paulsen 1955, Kingsolver et al. 1977). Swier a constaté que cette espèce était responsable de 93 pour cent des dégâts de bruchidés sur les semences de Prosopis velutina en Arizona.
Aucune étude écologique n'a été faite sur Algarobius bottimeri Kingsolver, mais on suppose que cette espèce a des caractéristiques écologiques très semblables à celles d'A. prosopis. Selon Kingsolver et al. (1977), A. bottimeri a été introduit aux îles Hawai, où il s'attaque à Prosopis pallida. On peut supposer que les études écologiques sur “Bruchus prosopis” effectuées par Bridwell (1918, 1920a, b) concernaient en réalité A. bottimeri.
Les applications d'insecticides pour lutter contre A. prosopis devraient sans doute débuter lorsque les gousses sont immatures et que la présence d'adultes de cette espèce peut être vérifiée.
Dans une fructification donnée, dans des conditions exceptionnelles, cette espèce peut attaquer un nombre important de semences de Prosopis. Selon Swier (1974) et Conway (1980), toutefois, elle cause beaucoup moins de dégâts qu'A. prosopis. Elle a un cycle biologique analogue à ce dernier, mis a part le fait qu'elle ne pond ses oeufs à la fin du printemps (juin) que sur les gousses immatures dont les graines ont des cotylédons bien développés et sur les gousses mûres, qu'elle colle ses oeufs au hasard sur la gousse, les plaçant souvent les uns sur les autres, et que les larves pénètrent directement dans la gousse à travers la base de l'oeuf. Swier (1974) a constaté que ce bruchidé pouvait détruire jusqu'à 3 pour cent des semences de Prosopis velutina en Arizona. Il se nourrit aussi de graines autres que celles de Prosopis (Tableaux 2 et 3, pages 42 et 46).
Comme M. amicus, cette espèce peut être localement abondante et détruire une quantité importante de semences de Prosopis. Son cycle biologique est analogue à ceui d'A. prosopis, mis à part le fait qu'elle hiverne probablement à l'état adulte (Swier 1974), qu'elle ne pond que sur les gousses immatures, et que c'est l'espèce de bruchidé qui est la première à les quitter à la fin de l'été. M. protractus colle de préférence ses oeufs au voisinage de la suture de la gousse; ses larves sont moins mobiles, et penètrent dans la gousse directement à travers la base de l'oeuf. Du fait qu'elle n'utilise que les gousses immatures, cette espèce ne produit qu'une génération par an sur P. velutina en Arizona (Swier 1974). Bien qu'étant assez rare, Conway (1980) a constaté que M. protractus était responsable de 61 pour cent des dégâts de bruchidés sur les semences de P. velutina près de Black Canyon City en Arizona, tandis que M. amicus et A. prosopis intervenaient pour 22 et 14 pour cent respectivement. Par conséquent, dans certaines conditions écologiques, des bruchidés peu communs peuvent se multiplior en abondance.
Cette espèce peu commune n'a pas été signalée comme détruisant des quantités importantes de semences de Prosopis, même dans des conditions locales particulières. Son cycle biologique est analogue à celui d'A. prosopis, si ce n'est qu'elle ne pond pas ses oeufs sur les gousses très immatures mais sur celles dont les graines ont des cotylédons développés, et qu'elle colle ses oeufs au hasard sur la gousse, les larves perforant la base de l'oeuf pour pénétrer directement dans la gousse. N. arizonensis se reproduit continuellement dans les graines, et a probablement 3 générations ou plus par an (Swier 1974). Sur un arbre donné il peut détruire jusqu'à 3 pour cent de la fructification, mais habituellement il n'attaque que moins de l pour cent de la fructification de P. velutina en Arizona (Swier 1974). Conway (1980) a constaté que ses effectifs étaient insignifiants en comparaison des 3 autres espèces de bruchidés des mesquites qu'il a étudiés en Arizona.
Des espèces appartenant aux genres Acanthoscelides, Pectinibruchus, Rhipibruchus, Rhipibruchus et Scutobruchus sont les principaux bruchidés s'attaquant aux semences de Prosopis en Argentine, au Chili et au Pérou, les trois derniers genres étant strictement liés pour leur alimentation aux graines des Prosopis (Tableaux 1, 2 et 3, p. 40, 42, 46).
Leur cycle biologique et leur nocivité sont loin d'avoir été étudiés aussi en détail que ceux des bruchidés d'Amérique du Nord. Selon Kingsolver et al. (1977), on constate des différences de comportement entre les espèces de ces quatre genres. Les Scutobruchus pondent leurs oeufs isolément sur les gousses immatures et mûres, aussi bien sur l'arbre que tombées à terre. Une espèce de Rhipibruchus pond ses oeufs en amas se recouvrant partiellement, de la même façon que Mimosestes amicus en Arizona (Swier 1974). Les adultes de Rhipibruchus sortent des gousses de Prosopis chilensis avant ceux de Scutobruchus, et les femelles sortent avant les mâles. Les représentants des deux genres hivernent à l'état adulte, se cachant dans des endroits abrités tels que vieilles gousses ou fissures de l'écorce. A la différence de certains bruchidés d'Amérique du Nord, les adultes de ces deux genres ne visitent pas les fleurs.
Reyes et Hermosilla (1974) ont étudié le cycle biologique de Scutobruchus gastoi Kingsolver en laboratoire, sur des fruits de Prosopis tamarugo. Ils décrivent l'oeuf, la larve et la nymphe, et constatent qu'en laboratoire la durée du cycle varie entre 80 jours et 4 mois. Habit et al. (1981) indiquent que dans la nature cette espèce hiverne dans des gousses sur le sol, et en sort lorsque la floraison de P. tamarugo commence de nouveau.
Selon Kingsolver et al. (1977), les interactions entre espèces observées dans les semences de P. velutina se retrouvent pour une large part chez les bruchidés parasitant les algarrobos en Argentine. Des échantillonnages détaillés effectués en Argentine sur 9 pieds de P. flexuosa ont montré que le total de semences détruites croissait avec le temps en fonction logarithmique depuis la maturation de la première gousse jusqu'environ 75 jours plus tard. Des comptages de fruits attaqués en Amérique du Nord ont montré un palier dans les attaques avec le temps; cette diminution relative était due au fait que les fruits deviennent plus difficiles à localiser au fur et à mesure que la saison avance. Dans la zone étudiée en Argentine, il ne se produisait pas de palier dans les attaques de bruchidés parce que les moutons et les chèvres enlevaient toutes les gousses qui tombaient au sol; en l'absence de troupeaux, on aurait pu s'attendre à un tel palier comme en Amérique du Nord.
En Argentine Scutobruchus ceratioborus a détruit 26 pour cent des graines de Prosopis flexuosa, et 90 pour cent de celles de P. chilensis (Kingsolver et al. 1977), et il semble être l'équivalent écologique sud-américain d'Algarobius prosopis en Amérique du Nord.
L'importance des dégâts de bruchidés varie considérablement d'un arbre à l'autre aussi bien en Arizona qu'en Argentine (Kingsolver et al. 1977). Sur 9 pieds de P. flexuosa, en Argentine, les dégâts causés aux semences variaient entre 0,7 pour cent dans les gousses tombées au sol pour un arbre, et 53 pour cent pour un autre. Swier (1974) rapporte une variation analogue d'un arbre à l'autre pour P. velutina en Arizona. Il constate que la différence dans la proportion de semences détruites peut être attribuée en partie à des différences individuelles entre les arbres. D'autres facteurs qui contribuent à cette variation dans les dégâts sont la station sur laquelle se trouve la population de mesquite, la quantité de fruits et de graines produites par un arbre donné, la direction d'où proviennent les bruchidés à partir d'autres, arbres, et la hauteur relative des gousses dans les cimes. Toutefois, l'influence de chacune de ces variables diffère selon les espèces de bruchidés. C'est ainsi par exemple que Swier (1974) constate une tendance significative pour la plupart des espèces à l'exception de Neltumius arizonensis à préférer les fruits situés dans la partie haute des cimes de Prosopis.
Caryedon serratus attaque les semences de Prosopis à Hawaï et en Israël (Bridwell 1920b, Belinsky et Kugler 1978). Cette espèce de l'Ancien Monde se nourrit principalement des graines du tamarinier (Tamarindus indica), et a par suite été introduite en même temps que cet arbre dans la plupart des pays tropicaux; dans le Nouveau Monde elle est largement répandue sur les tamariniers au Mexique. C. serratus est un sérieux ennemi de l'arachide (Arachis hypogaea) en Afrique; étant donné que l'arachide a été introduite du Nouveau Monde en Afrique, ce bruchidé a donc la faculté de changer d'hôte, et peut constituer un danger économique sérieux. Il pourrait donc s'attaquer également aux Prosopis du Nouveau Monde. Belinsky et Kugler (1978) ont étudié son cycle biologique sur Prosopis en Israël, de même que Bridwell (1918, 1920b) à Hawaï. Les premiers constatèrent que la sous-espèce qu'ils étudiaient, Caryedon serratus palaestinicus, préférait en conditions de laboratoire son hôte naturel, Prosopis farcta, à l'arachide; ils en concluent que cette sous-espèce n'est pas une menace pour l'arachide en Israël.
Le genre Acanthoscelides renferme un plus grand nombre d'espèces (environ 300) que n'importe quel autre genre de bruchidés du Nouveau Monde (Johnson 1981b). Etant donné qu'on y trouve un important parasite des haricots, Acanthoscelides obtectus (Say), de nombreuses informations ont été accumulées sur l'écologie d'A. obtectus et d'autres espèces du même genre n'ayant pas d'importance économique (Johnson 1970, 1981b). En raison des nombreuses espèces qu'il renferme, et de la diversité de leurs plantes hôtes (9 familles botaniques), on ne peut guère prédire le comportement et la nocivité possible des Acanthoscelides qui se nourrissent des graines de Prosopis.
Le genre Amblycerus est semblable au genre Acanthoscelides en ce qu'il renferme un grand nombre d'espèces, et que celles-ci se nourrissent aux dépens de plantes de familles diverses. Etant donné que 3 espèces d'Amblycerus ont été signalées sur des Prosopis, elles représentent un danger potentiel pour d'autres espèces de ce genre.
On a émis l'hypothèse que la spécificité et la grande compétitivité des bruchidés vis-à-vis des semences hôtes sont dues essentiellement à des relations chimiques précises (Kingsolver et al. 1977). Cela ne semble pas être le cas pour les bruchidés qui s'attaquent aux Prosopis. Des interactions concurrentielles semblent limiter les effectifs des bruchidés sur une espèce donnée de Prosopis. On peut soutenir qu'il y a absence de spécificité chimique, parce que les graines de Prosopis ne sont pas connues pour être toxiques, qu'il y a de nombreux exemples de changements d'hôte de bruchidés lorsqu'il n'y a pas de représentants des Prosopis, et qu'il y a plusieurs espèces de bruchidés du genre Stator qui se reproduisent librement dans les graines de Prosopis velutina en conditions expérimentales de laboratoire (Johnson 1981d). Au siècle dernier Prosopis pallida, espèce indigène du Pérou, a été introduit aux îles Hawaï comme source d'aliments pour le bétail (Fosberg 1966). Plusieurs espèces de bruchidés ont également été involontairement introduites; elles comprennent Mimosestes nubigens (= M. sallaei), que l'on trouve normalement dans les graines de 6 espèces d'Acacia en Amérique du Nord (Kingsolver et al. 1977). Aux îles Hawaï où ces bruchidés ont été introduits, tous utilisent les gousses de P. pallida. Toutes ces observations montrent que les fruits des Prosopis sont vulnérables aux attaques de bruchidés, et que jusqu'à 4 espèces peuvent coexister dans la même localité. Cette coexistence peut s'expliquer en partie par le fait qu'elles attaquent leur hôte à des moments différents de la saison (Figure 2), ou qu'elles pondent leurs oeufs sur des fruits situés à des hauteurs différentes sur l'arbre, ou encore par des différences dans l'aptitude à utiliser d'autres hôtes.
Diverses espèces de bruchidés sont connues comme ayant un cycle biologique particulier, adaptépour leur permettre de se nourrir des graines d'hôtes spécifiques. Par exemple, les adultes de bruchidés qui attaquent les graines de Prosopis sont modifiés pour pondre leurs oeufs à la surface ou dans des fissures de gousses ligneuses indéhiscentes; leurs larves sont spécialisées pour pénétrer dans les gousses et les graines. Johnson (1981c) a constaté récemment que des gousses de légumineuses indéhiscentes, déhiscentes et partiellement déhiscentes sont toutes attaquées par différents groupes écologiques de bruchidés. Il constata que dans un même groupe écologique les genres de bruchidés qui déposent leurs oeufs sur les gousses sont différents de ceux qui pondent sur les graines. Il constata également que les graines de certaines espèces de légumineuses sont attaquées par un autre groupe écologique de bruchidés seulement après être tombées sur le sol. Ce dernier groupe écologique, dit “des graines dispersées”, se compose de plusieurs espèces du genre Stator qui sont très spécifiques vis-à-vis des graines de leurs hôtes respectifs. Ce qui est important dans le contexte de ce manuel sur les insectes des Prosopis est que dans des conditions expérimentales de laboratoire Johnson (1981d) a constaté que plusieurs espèces du groupe écologique des graines dispersées se développent facilement sur des graines autres que celles de leur hôte. Une de ces espèces, Stator sordidus (Horn), se montrait capable d'accomplir son développement dans les graines de 20 plantes non hôtes différentes. Le plus intéressant est que 4 espèces de ce groupe écologique se développaient dans des graines de Prosopis velutina, plante non hôte. Il convient de noter pour les futures recherches sur les bruchidés des Prosopis que les représentants de ce groupe écologique de bruchidés sont effectivement susceptibles de s'attaquer aux graines de Prosopis dans la nature et en magasin.
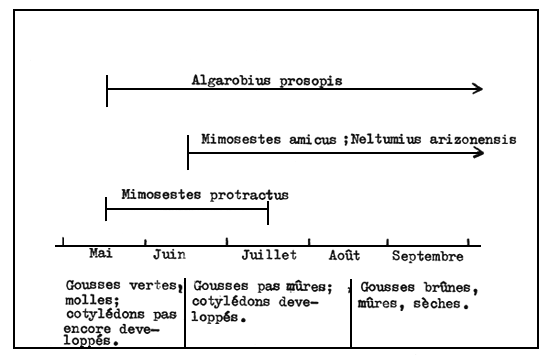
Fig. 2 - Périodes de ponte et stades de développement des gousses de Prosopis velutina sur lesquelles pondent 4 espèces de bruchidés coexistant sur les fruits de ce Prosopis en Arizona
Un certain nombre d'études récentes ont décrit l'écologie et les interactions évolutives entre les bruchidés et leurs plantes hôtes (Janzen 1969, 1971a, b, c, 1980; Forister 1970; Center et Johnson 1974; Johnson et Slobodchikoff 1979; Johnson 1981c). Selon Janzen (1969), la plupart des légumineuses semblent suivre l'une des deux stratégies suivantes pour éviter les déprédations des bruchidés sur leurs semences. Certaines produisent de grosses graines toxiques qui écartent tous les ravageurs ou presque. D'autres ont de petites graines qui sont mangées par les bruchidés, mais qui sont produites en si grand nombre que certaines échappent aux ravageurs. Center et Johnson (1974) et Johnson (1981c) ont apporté la preuve que les bruchidés ont développé des mécanismes pour contrecarrer les dispositifs protecteurs des plantes. Par exemple, certains bruchidés sont capables de se nourrir de graines très toxiques, tandis que d'autres s'attaquent aux petites graines après qu'elles se sont dispersées. Janzen (1969) discute de 31 dispositifs protecteurs développés par des plantes pour faire obstacle aux déprédations des bruchidés. Certains Prosopis ont semble-t-il développé la stratégie qui consiste à produire une fructification très abondante qui assure qu'au moins un certain nombre de semences échapperont aux bruchidés.
Des mécanismes plus spécifiques auxquels les légumineuses recourent pour réduire au minimum les dégâts d'insectes parasites des graines sont les suivants: 1) les arbres poussant dans un milieu relativement constant et uniforme ont la faculté d'échelonner dans le temps la fructification de certains individus de sorte que la multiplication des populations d'insectes se trouve freinée; 2) formation de graines charnues qui se dessèchent trop rapidement pour que les ravageurs puissent s'en nourrir au stade larvaire; 3) production de graines si petites qu'elles ne contiennent pas assez de nourriture pour assurer le développement complet d'un insecte. Selon Kingsolver et al. (1977), aucune de ces possibilités ne s'offre aux Prosopis poussant dans des milieux désertiques, parce que la fructification de tous les individus est synchronisée de façon à se produire au moment de l'année le plus favorable pour la germination des semences, et que les graines doivent être capables à la fois de résister à la dessication et de fournir au jeune semis une quantité d'énergie initiale adéquate. De nouvelles recherches sont encore nécessaires avant que les hypothèses de Kingsolver et al. (1977) puissent être acceptées ou rejetées.
La plupart des espèces de bruchidés sont extrêmement spécifiques vis-à-vis de l'hôte, et la plupart des plantes hôtes n'entretiennent qu'un petit nombre de bruchidés dans leurs semences. Johnson et Slobodchikoff (1979) ont constaté que 82,5 pour cent des bruchidés s'attaquant aux Cassia avaient 3 hôtes ou moins, et que 90,5 pour cent des espèces de Cassia avaient dans leurs semences 3 espèces de bruchidés ou moins. Johnson (1981b) indique qu'environ 70 pour cent des espèces de bruchidés du genre Acanthoscelides ont 1 ou 2 hôtes, et que 74 pour cent de leurs hôtes ont 1 ou 2 espèces de bruchidés se nourrissant de leurs semences. Des résultats identiques sont présentés par Janzen (1980) dans sa discussion sur les causes de la spécificité vis-à-vis de l'hôte de coléoptères parasites des graines au Costa Rica.
![]()
![]()
![]()