![]()
![]()
![]()
Los insectos son los únicos invertebrados conocidos que usan los frutos y las semillas de Prosopis como fuente alimenticia (Kingsolver et al, 1977). Su alimentación por parte de los insectos puede dar por resultado la destrucción total de los frutos (por ejemplo, por lepidópteros), pero el resultado más común es que los frutos o las semillas aborten o que las semillas vengan destruídas. El autor llama depredadores a los insectos que se alimentan de la semilla puesto que ellos destruyen totalmente el organismo (por ejemplo, la semilla), pero los insectos podrían también describirse como parasitas cuando su alimentación no interfiere con la capacidad de la semilla de germinar.
Por conveniencia, el autor se ajusta a Kingsolver et al. (1977) dividiendo en dos grupos los insectos que se alimentan con los frutos y semillas de Prosopis, los que lo hacen principalmente desde afuera y los que se alimentan desde adentro. Los que se alimentan desde afuera son adultos y ninfas de hemípteros y larvas de lepidópteros. Los que se alimentan internamente incluyen larvas de lepidópteros y coleópteros de las familias Curculiónides, Cerambícides y Brúcides. Entre los gorgojos, los brugos constituyen a la larga las pestes más importantes. Hasta un cierto punto, la edad del fruto influye sobre el tipo de alimentación por parte de los insectos. Los hemípteros tienen elementos de la boca perforantes y chupadores tan delicados que los frutos inmaduros de Prosopis son rápidamente comidos, mientras que las vainas secas y leñosas ni les atraen ni son fácilmente perforadas. Las larvas de lepidópteros que se alimentan desde afuera utilizan también las vainas delicadas, inmaduras. La edad de la vaina, por otra parte, no es tan importante para la mayoría de los que se alimentan desde adentro, ya que la mayoría come la semilla y ésta es a menudo tierna. Para los que comen desde adentro, el poder entrar en la vaina y en la semilla es el paso más crucial, por lo menos para los que han sido más estudiados, o sea, los gorgojos brúcides. La alimentación interna, en todos los casos conocidos, es a través de las fases larvales. Se cree que los adultos de los que se alimentan internamente se alimentan a su vez con néctar y polen, si bien algunos gorgojos brúcides no se alimentan cuando adultos.
Las verdaderas chinches (hemípteros) son importantes pestes nativas del mezquite. Según Ueckert (1973), la chinche patifoliada, Mozena obtusa Uhler (Coreidae), destruye grandes cantidades de vainas de Prosopis glandulosa en América del Norte. En Texas, estos insectos comienzan su actividad a principios de la primavera y se mantienen activos hasta Agosto. Swenson (1969) estimó que los daños al mezquite en Texas alcanzaban el 50% de la producción potencial de frutos. Se trata del hemíptero más frecuentemente hallado sobre frutos jóvenes de Prosopis velutina cerca de Tucson, Arizona (Werner y Butler, 1958). Ueckert (1973) halló que el aborto de vainas inmaduras de P. glandulosa variaba entre el 33% y el 89% cuando eran comidas por estas chinches. Además, el comer de las chinches redujo el peso seco de las vainas del 23% al 59% y el porcentaje de germinación de la semilla; desde un valor normal del 65% bajó hasta el 4,3%. Puesto que el mezquite se reproduce sólo por semilla, la intervención de este enemigo natural puede ser significativa para limitar la reproducción de la planta. Otra chinche, la conchuela (Chlorochroa ligata (Say), Pentatomidae), destruye las semillas de mezquite en el Texas chupando los jugos de las semillas, dejando en la vaina solamente los tegumentos secos de la semilla (Smith y Ueckert, 1974). Unas vainas en una jaula de control produjeron semillas con un porcentaje de germinación mucho más alto (76%) del de las semillas producidas en jaulas con conchuelas (de 0% a 4% germinables). En un estudio sobre Prosopis flexuosa en Argentina, sólo 40 de los 145 frutos incipientes, y en P. chilensis, sólo 4 de los 70 frutos iniciales, alcanzaron su madurez (Solbrig y Cantino, 1975). Según Kingsolver et al. (1977), es posible que un daño por insecto, similar al que en América del Norte produce la chinche patifoliada, fuese responsable de gran parte de este aborto frutal en Argentina. Es asimismo posible que otros factores, como la tensión del agua y el autoaborto hayan contribuido también a esta mortandad.
La mayoría de las larvas de lepidópteros que se alimentan desde el externo consumen las flores, los frutos inmaduros y las vainas en maduración de los Prosopis (Kingsolver et al, 1977). Una cierta cantidad de lepidópteros en Norteamérica se alimenta dentro de las vainas de Prosopis. Tanto los interiores de las vainas como las semillas son consumidos por algunos representantes de los Lycaenidae (Strymon leda), Olethreutidae (Ofatulena spp.), Pyralidae (Paramyelois spp.), Blastodacnidae (Chaetocampa spp.), Notodontidae (Didigua argentilinea) y Cochyledae (Phalonia leguminana). Ha sido mencionada, por criarse en gran cantidad en vainas de mezquite en almacenaje en Hawai (Bridwell, 1920b), la polilla “indian-meal” (Pyralidae: Plodia interpunctella (Hubner). Los huevos de esta tiña vienen depositados sobre las bolsas en las que se almacenan las vainas, o sobre las vainas y luego los brugos penetran en la vaina, preferentemente en las vainas rotas, donde se alimentan con la pulpa azucarada. Al llegar los brugos a su madurez, dejan las vainas y buscan un área protegida donde puedan pupar. Según Koch y Campos (1978), la Cryptophlebia carpophagoides Clarke (Lepidoptera: Olethreutidae) se alimenta sobre las vainas de Enterolobium contortisiliquum Vell. en Argentina y en Prosopis tamarugo y P. juliflora en Chile. Esta especie se alimenta dentro de la vaina con las semillas y con el tejido tierno y con frecuencia pega sus excrementos sobre el exterior del fruto. Estos autores atribuyen a esta polilla hasta el 30% de las pérdidas de las vainas de P. tamarugo. Habit et al. (1981) describieron el ciclo vital y la estructura de esta falena en Chile y mencionan que los daños alcanzaron el 17,4% de los frutos de P. tamarugo. Koch y Campos (1978) y Habit et al. (1981) informaron que tambien Leptotes trigemmatus Butler (Lepidoptera) se alimentaba con los frutos de P. tamarugo en Chile. Esta especie prefiere comer los frutos en sus primeras fases de desarrollo, lo que eventualmente provoca la caída del fruto.
En Arizona, tres especies de curculiónides (Apion subornatum, A. ventricosum, Microtychius sp.) se alimentan con la semilla de P. velutina (Kingsolver et al., 1977). En Argentina, los gorgojos de Sibinia asulcifera y S. mastuerzo se alimentan probablemente con la semilla de Prosopis (Clark, 1979). Los gorgojos del género Lophopoeum (cerambícide) son conocidos por desarrollarse dentro de las vainas de Prosopis en Argentina y de consumir todas las semillas dentro de la vaina mientras se están desarrollando (Kingsolver et al. 1977).
Los gorgojos de las semillas (Bruchidae) son los insectos a la larga más numerosos (tanto en cantidad de especies como en individuos) mejor conocidos, que comen los frutos de Prosopis en el Nuevo Mundo (Kingsolver et al., 1977). Las larvas de los brúcides se alimentan de las semillas de 32 familias vegetales registradas (Johnson 1981a), pero la mayoría de las especies se alimenta dentro de una familia, las leguminosas. Los brúcides están estrechamente relacionados con las familias Chrysomelidae y Cerambycidae. Si bien las otras dos familias se alimentan con diferentes partes vegetales, los brúcides son conocidos por comer sólo las semillas. En la actualidad, la familia Bruchidae comporta alrededor de 1 300 especies, agrupadas en 56 géneros en las subfamilias Amblycerinae, Brucinae, Eubaptinae, Kyrtorhininae, Pachymerinae y Rhaebinae (Johnson, 1981a). La mayoría de las especies (80%) de los brúcides ha sido en la actualidad asignada a la subfamilia Bruchinae.
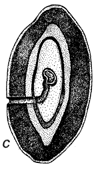
En la Figura 1 se ilustra una típica historia vital de un brúcide, pero los gorgojos de las semillas atacan a éstas en diversas formas (ver más adelante). Su ciclo vital es por lo general que la hembra adulta deposita los huevos sobre una semilla o vaina (Figura 1), la primera etapa de la larva mastica a través de la caparazón del huevo, de la pared de la vaina y/o del tegumento de la semilla y luego entra en la semilla. La primera etapa de la larva (Figura 1) se modifica mucho para penetrar en las semillas y posee muchas espinas, pelos, etc., para ello (Pfaffenberger y Johnson, 1976). Poco después de entrar en la semilla se muda en un gusano sin patas que es muy diferente de la larva de la primera etapa y se modifica para alimentarse dentro de la semilla. La larva por lo general se alimenta dentro de una semilla o en algunos brúcides, de dos a varias semillas, pasando normalmente por tres o más mudas, a medida que continúa alimentándose y aumentando de tamaño. Llega por lo general al estado pupal dentro de una sola semilla, si bien algunas especies dejan la semilla y pupan en un capullo, mientras que otras especies encolan varias semillas juntas formando una cámara pupal. Después del estado pupal, el adulto completa una típica perforación redonda de salida (Figura 1), que había sido casi terminada por la larva y deja la semilla para iniciar un nuevo ciclo de vida. La duración del ciclo vital varía, pero generalmente dura 30 días. Los brúcides adultos probablemente se alimentan con néctar y polen y no se tiene conocimiento de que se alimenten de o sobre las semillas, excepto incidentalmente, como cuando emergen de una semilla o cuando una hembra roe un orificio en la vaina para depositarle luego los huevos. Algunas especies de brúcides, especialmente los de importancia económica, sobreviven el curso de muchas generaciones en los envases de semillas, en el laboratorio o en depósitos, sin que los adultos se alimenten.
 | |
 |  |
 |  |
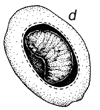 | |
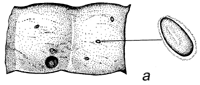
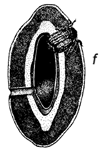
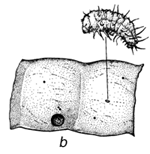
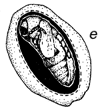
Figura 1 Reconstrucción del ciclo vital de los brúcides en las vainas de Prosopis: a. huevos pegados a la superficie de la vaina o depositados en las grietas de la vaina o en los foros de salida de un brugo adulto (orificio redondo). b. orificio de entrada de la primera etapa de la larva que ha perforado la pared de la vaina, y primera etapa larval, agrandada, para mostrar los pelos, espinas y patas modificadas para entrar en la semilla. c. corte transversal de la vaina y de la semilla para indicar el hueco hecho por la primera fase larval al entrar. d. fase posterior de la larva, dentro de la cavidad roída en la semilla. e. pupa dentro de la cámara de alimentación larval. f. adulto saliendo a través del orificio, preparado por la última fase larval.
Ahora se conocen muchas plantas que sirven de huéspedes para los brúcides (Johnson, 1981a). Esto no significa que se sepa mucho sobre los huéspedes de los brúcides. En efecto, estudios recientes sobre el terreno han inducido al autor del presente manual a creer que los brúcides se alimentan con semillas de muchas más plantas, especialmente de leguminosas neotropicales, de lo que hasta ahora se había observado.
Se conocen cinco géneros de Bruchidae que se alimentan sólo de las semillas de especies de Prosopis (Cuadro 1, pág. 37), pero otros 5 géneros se alimentan de Prosopis y de semillas de plantas que se hallan en estrecha asociación con Prosopis, tales como Acacia, Cercidium y Parkinsonia. En el Nuevo Mundo, alrededor de 32 especies de brúcides se alimentan con los frutos de Prosopis. De estas especies, 28 son obligadamente restringidas a Prosopis, pero especies individuales de brúcides pueden alimentarse con diferentes especies de Prosopis. En el Cuadro 2 (pág. 38) se da una lista de brúcides y de plantas huéspedes, y en el Cuadro 3 (pág. 42) la de las especies de Prosopis con los brúcides que se alimentan con sus semillas.
Las especies de los géneros Algarobius, Neltumius y Mimosestes son los principales brúcides que infestan la semilla de Prosopis, desde Estados Unidos de Norteamérica hasta Venezuela (Cuadro 1). Todas las especies en estos tres géneros que se alimentan sobre Prosopis se restringen obligadamente a Prosopis, haciendo excepción las Mimosestes amicus y M. insularis. En el Cuadro 2 se indican las otras plantas huéspedes para estas dos últimas especies.
La historia vital y el daño potencial a las semillas de Prosopis han sido estudiados más a fondo para Algarobius prosopis, Mimosestes amicus, M. protractus y Neltumius arizonensis (Swier 1974, Kingsolver et al. 1977, Conway 1980), de manera que se presenta a continuación la información disponible.
En las semillas de Prosopis de Norteamérica esta especie es enormemente más abundante y por ello es potencialmente grande el daño que puede producir a las semillas de mezquite. Según Swier (1974), esta especie probablemente pasa el invierno, como madura o adulta, en las semillas de Prosopis velutina en Arizona. Las hembras adultas emergen en la primavera y depositan los huevos sobre los restos de las cosechas de semilla de los años precedentes. Cuando las hembras son pequeñas, las vainas inmaduras comienzan a formarse sobre los árboles a fines de la primavera (mayo, junio); ellas depositan huevos sobre estas vainas continuamente a medida que maduran, hasta su madurez a fines del otoño. El A. prosopis no encola sus huevos sobre las vainas o las semillas, como muchos brúcides hacen, sino que las hembras los colocan dentro de rajaduras y grietas de la vaina (Bridwell, 1920a; Swier, 1974). A menudo, esta especie utiliza como sitio para depositar los huevos las perforaciones de donde han salido los adultos. Las larvas de A. prosopis son muy móviles, con patas bien desarrolladas, así como el tabique sensorial (Pfaffenberger y Johnson, 1976), lo que se correlaciona bien con un fuerte comportamiento locomotor. Dentro de las vainas muy inmaduras las larvas probablemente se alimentan con el jugo dulce de las vainas hasta que se desarrollan los cotiledones (Bridwell, 1920b). Sobre vainas más maduras, o penetran dentro de la primera semilla que encuentran o se arrastran a través de la pulpa de la vaina para entrar en otra semilla. En los depósitos, las larvas pueden perforar el tegumento de la semilla desde la superficie de la vaina. Después de alimentarse y haber mudado alrededor de tres veces dentro de una determinada semilla, las larvas recubren la cámara que han abierto con desperdicios y luego se mudan en pupas.
En condiciones naturales, Algarobius prosopis desarrolla tres o más generaciones por año. Según Bridwell (1920b), esta especie no se cría sobre vainas secas, almacenadas de mezquite pero continúa a criarse, si las vianas son húmedas. Swier (1974) ha observado larvas de esta especie penetrar vainas secas. Se debe investigar más, empleando tanto vainas secas como húmedas, para determinar el peligro potencial que representa A. prosopis en la destrucción de semillas almacenadas de Prosopis.
Para una determinada cosecha de un árbol, esta especie puede destruir, en condiciones normales, del 8% al 75% de las semillas (Swier, 1974; Glendening y Paulsen, 1955; Kingsolver et al., 1977). Swier halló que esta especie era responsable del 93% de la depredación brúcida de las semillas de Prosopis velutina en Arizona.
No se han hecho estudios ecológicos de Algarobius bottimeri Kingsolver, pero se cree que tiene características ecológicas muy similares a las de A. prosopis. Según Kingsolver et al. (1977) A. bottimeri ha sido introducida en las islas de Hawai, donde se alimenta sobre Prosopis pallida. Es concebible que los estudios ecológicos hechos por Bridwell (1918, 1920a, b) sobre “Bruchus prosopis” fueron en realidad sobre A. bottimeri.
Las aplicaciones de insecticidas para el control de A. prosopis deberían probablemente empezar cuando las vainas son inmaduras y cuando puede verificarse la presencia de adultos de esta especie.
En una determinada cosecha y en condiciones muy especiales, esta especie se alimenta con una notable cantidad de semillas de Prosopis. Según Swier (1974) y Conway (1980), esta especie, sin embargo, provoca menos daños que A. prosopis. Esta especie se parece en su ciclo vital a A. prosopis, con la excepción de que M. amicus deposita los huevos a fines de primavera (junio) sólo sobre vainas inmaduras con cotiledones bien desarrollados y sobre vainas maduras, adhiere los huevos al azar sobre la vaina, a veces superponiendo los huevos uno arriba del otro, y las larvas entran en la vaina directamente por la parte inferior del huevo. Swier (1974) halló que esta especie destruye hasta el 3% de la semilla de Prosopis velutina en Arizona. Esta especie se alimenta de semillas diferentes de las de Prosopis (Cuadros 2, 3).
Como en el caso de M. amicus, esta especie puede ser localmente abundante y destruir significativas cantidades de semillas de Prosopis. El ciclo vital de M. protractus se parece al de A. prosopis, excepto que probablemente hiberna en estado de adulto (Swier, 1974), deposita huevos sólo sobre vainas inmaduras y es la primera especie de brúcides que utiliza las vainas a fines de la primavera, y la primera que abandona las vainas a fines de verano. M. protractus cementa preferentemente los huevos cerca de la sutura de la vaina, las larvas son menos movedizas y penetran la vaina de la semilla, directamente a través de la parte inferior del huevo y, dado que utiliza solamente las vainas inmaduras, esta especie produce una sola generación cada año, sobre P. velutina en Arizona (Swier, 1974). Si bien M. protractus es un brúcide bastante raro, Conway (1980) encontró que era responsable del 61% de los daños por brúcides sobre semillas de P. velutina cerca de Black Canyon City, Arizona, mientras que M. amicus y A. prosopis eran respectivamente responsables del 22% y del 14% del dano. Por lo tanto, bajo ciertas condiciones ecológicas, los gorgojos de brúcides poco comunes pueden multiplicarse abundantemente.
Aún bajo condiciones locales, esta especie poco común no destruye grandes cantidades de semillas de Prosopis, según las referencias existentes. Su ciclo vital es parecido al de A. prosopis, con excepción de que no deposita huevos sobre vainas demasiado inmaduras, sino que sobre las que han desarrollado los cotiledones de las semillas y cementa lo huevos al azar sobre la vaina y las larvas perforan a través de la parte inferior del huevo directamente en la vaina. N. arizonensis se cría en las semillas en forma continua y desarrolla probablemente tres o más generaciones por año (Swier, 1974). En un determinado árbol, esta especie puede destruir hasta el 3% de una cosecha de semillas, pero, por lo general, se alimenta sobre menos del 1% de la semilla cosechada de P. velutina en Arizona (Swier, 1974). Conway (1980), halló que la cantidad de N. arizonensis era insignificante comparada con las otras tres especies de brúcides de mezquite que había estudiado en Arizona.
Las especies en los géneros Acanthoscelides, Pectinibruchus, Rhipibruchus y Scutobruchus son los principales brúcides que infestan la semilla de Prosopis en Argentina, Chile y Perú. Las especies de los tres últimos géneros se limitan a comer exclusivamente semillas de Prosopis (Cuadros 1,2,3).
Los ciclos vitales y los daños potenciales no han sido estudiados en forma tan extensa como en Norteamérica. Según Kingsolver et al. (1977) hay diferencias de comportamiento entre las especies de estos géneros. Las especies de Scutobruchus depositan huevos en forma individual sobre vainas inmaduras y maduras, y sobre vainas -sea en el árbol que sobre el suelo. Una especie de Rhipibruchus deposita sus huevos parcialmente sobrepuestos en pilas como en el caso de Mimosestes amicus en Arizona (Swier, 1974). Los adultos de Rhipibruchus emergen de las vainas de Prosopis chilensis antes de los adultos de Scutobruchus, y las hembras salen antes de los machos. Especies de ambos géneros pasan el invierno como adultos, que se esconden en lugares protegidos como viejas vainas o grietas en las cortezas. Contrariamente a las especies de brúcides de Norteamérica, los adultos de estos dos géneros no visitan las flores.
Reyes y Hermosilla (1974) han estudiado el ciclo vital de Scutobruchus gastoi Kingsolver, en el laboratorio sobre frutos de Prosopis tamarugo. Describieron el huevo, la larva y la pupa y hallaron que, en el laboratorio, el ciclo vital varía desde 80 días a 4 meses. Habit et al. (1981) citan que en el campo esta especie pasa el invierno en vainas sobre el suelo y emerge de las vainas cuando la floración comienza de nuevo en P. tamarugo.
Según Kingsolver et al. (1977) muchos de los mismos tipos de interacciones observados entre las especies, sobre las semillas de P. velutina, tienen también lugar en los brúcides que infestan los algarrobos en Argentina. Muestreos detallados de 9 árboles de P. flexuosa, en Argentina, mostraron que la destrucción total de la semilla subía logarítmicamente con el tiempo, desde la maduración de la primera vaina hasta alrededor de 75 días después. Los recuentos de depredación de frutos en Norteamérica indicaron que, con el tiempo, la depredación se allanaba. La reducción proporcional en la depredación se debía a que resultaba cada vez más difícil localizar los frutos a medida que la estación adelantaba. En la zona estudiada en Argentina, una nivelación de la depredación no tuvo lugar puesto que ovejas y cabras domésticas recogieron todas las vainas que caían al suelo. Si no hubiese sido por las ovejas y las cabras, la nivelación de la depredación por parte de los brúcides podía esperarse como en el caso de Norteamérica.
Scutobruchus ceratioborus destruyó el 26% de las semillas de Prosopis flexuosa y el 90% de las de P. chilensis en Argentina (Kingsolver et al., 1977) y parece que es el equivalente ecológico sudamericano de Algarobius prosopis de Norteamérica.
La magnitud del daño causado por brúcides varía notablemente de un árbol a otro, tanto en Arizona como en Argentina (Kingsolver et al., 1977). En 9 árboles de P. flexuosa en Argentina, el daño a la semilla varió del 0,7% en las vainas caídas de un árbol, hasta el 53% en las de otro. Variaciones similares entre daños sobre semillas por parte de brúcides entre un árbol y otro fueron citadas por Swier (1974) sobre P. velutina en Arizona. Swier (1974) halló que las diferencias entre las cantidades de semilla dañada pueden atribuirse en parte a las diferencias entre los árboles individuales. Otros factores que contribuyeron en la variabilidad de las cantidades de semillas destruidas eran, el sitio de la población de mezquite, la magnitud de la producción de semillas y frutos de un determinado árbol, y la altura relativa de las vainas en el dosel. Sin embargo, el peso relativo de la participación de cada variable es diverso para cada especie de brúcide. Por ejemplo, Swier (1974) halló que había una significativa tendencia en la mayoría de las especies, exceptuando Neltumius arizonensis, de preferir los frutos ubicados en las partes más altas de las plantas de Prosopis.
Caryedon serratus come las semillas de Prosopis en Hawai e Israel (Bridwell, 1920b; Belinsky y Kugler, 1978). Esta especie del Viejo Mundo se alimenta principalmente de semillas de tamarindo (Tamarindus indica) y ha sido por lo tanto introducida en la mayoría de las áreas tropicales junto al tamarindo. Este brúcide ha sido introducido en el Nuevo Mundo y se halla ampliamente disperso sobre tamarindo en México. C. serratum es una grave peste del maní (Arachis hypogaea) en Africa. Puesto que el cacahuete ha sido introducido en Africa desde el Nuevo Mundo, esta especie tiene la característica potencial de cambiar de huésped y transformarse así en una seria peste económica. Potencialmente, por lo tanto, podría convertirse en un problema para Prosopis del Nuevo Mundo. Belinsky y Kugler (1978) estudiaron su ciclo vital sobre Prosopis en Israel, como lo hizo Bridwell (1918, 1920b) en Hawai. Belinsky y Kugler (1978) hallaron que la subespecie que ellos estudiaron, Caryedon serratus palaestinicus Southgate, prefería su huésped natural, Prosopis farcta, al maní en condiciones de laboratorio, y llegaron a la conclusión de que esta subespecie no representaba una amenaza para el cacahuete en Israel.
El género Acanthoscelides abarca más especies (alrededor de 300) que cualquier otro género de brúcides del Nuevo Mundo (Johnson, 1981b). Puesto que la importante peste de los frijoles, Acanthoscelides obtectus pertenece a este género, una notable cantidad de datos se han acumulado sobre la ecología de A. obtectus y otras especies no económicas de Acanthoscelides (Johnson 1970, 1981b). Debido a sus muchas especies y variadas plantas huépedes (9 familias vegetales), puede predecirse muy poco sobre el comportamiento y la capacidad como peste potencial de los Acanthoscelides que se alimentan con las semillas de Prosopis.
El género Amblycerus es similar al Acanthoscelides en cuanto es grande y sus especies se alimentan en diversas familias vegetales. Puesto que tres especies de Amblycerus han sido citadas en Prosopis, estas especies representan una amenaza para las semillas de otras especies de Prosopis.
Se ha sugerido que la mayor parte de la especificidad y de las ventajas competitivas de los brúcides sobre las semillas que los hospedan se debe a precisas relaciones químicas (Kingsolver et al., 1977). No parece ser el caso de los brúcides que se alimentan en Prosopis. Parece que las interacciones competitivas limitan la cantidad de brúcides sobre una determinada especie de Prosopis. Puede argumentarse que hay una falta de especificidad química porque las semillas no se sabe que sean tóxicas, que existen numerosos ejemplos de brúcides que se desplazan a nuevos huéspedes cuando falta la flora de Prosopis, y que hay varias especies de brúcides del género Stator que se crían libremente en las semillas de Prosopis velutina en condiciones experimentales de laboratorio (Johnson, 1981b). En los últimos 100 años, Prosopis pallida, nativa del Perú, ha sido introducida en las islas de Hawai, como fuente de alimento para el ganado (Fosberg, 1966). Varias especies de brúcides se han introducido también por descuido, e incluyen Mimosestes nubigens (=M. sallaei), especie que se halla normalmente en semillas de 6 especies de Acacia en Norte América (Kingsolver y Johnson, 1978), Caryedon serratus, y Algarobius bottimeri que se alimenta sólo con P. glandulosa en Texas (Kingsolver et al., 1977). En las islas de Hawai, donde fueron introducidos estos brúcides, todos utilizan las vainas de P. pallida. Todas estas observaciones indican que los frutos de Prosopis son vulnerables al consumo por parte de brúcides y que hasta 4 especies pueden coexistir en la misma localidad. Esta coexistencia puede deberse, en parte, a que ellos atacan un huésped en diferentes momentos durante la estación (Figura 2) o por depositar los huevos sobre frutos a diferentes alturas del árbol, o por la característica que los diferencia por usar otros huéspedes.
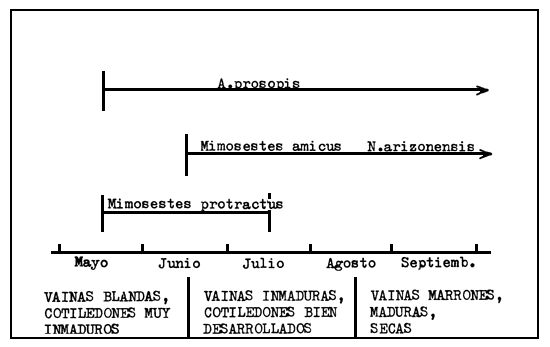
Figura 2 Períodos de oviposición y las fases de las vainas de Prosopis sobre las cuales vienen depositados los huevos de cuatro especies de gorgojos que coexisten en vainas de P. velutina en Arizona.
Se conocen varias especies de brugos que tienen singulares ciclos de vida, modificados para permitirles comer las semillas de huéspedes específicos. Por ejemplo, los brúcides adultos que atacan las semillas de Prosopis se han modificado para depositar huevos sobre el exterior de vainas leñosas indehiscentes, o sobre grietas en las mismas. Las larvas se han especializado para penetrar la vaina y las semillas. Johnson (1981c) halló recientemente que las vainas leguminosas que son indehiscentes, dehiscentes y parcialmente dehiscentes, todas alimentan diferentes agrupaciones de brúcides; halló que los géneros de brúcides en la agrupación que deposita huevos sobre legumbres son diversos de los géneros que los depositan sobre semillas; halló también que las semillas de algunas especies de leguminosas eran atacadas por otra agrupación de brúcides solamente después que han caído al suelo. Esta última agrupación, el “grupo sobre semilla suelta”, está representado por varias especies de Stator que son muy específicas con respecto a las semillas de sus respectivos huéspedes. Lo que es importante en el contexto de este manual sobre insectos de Prosopis es que bajo condiciones experimentales de laboratorio, Johnson (1981d) halló que varias especies del “grupo sobre semillas sueltas” se desarrollan rápidamente en semillas no huéspedes. Una especie, Stator sordidus (Horn), fue capaz de completar su desarrollo sobre semillas de 20 diferentes especies no huéspedes. Lo más interesante fue que 4 especies de este grupo se desarrollaron sobre semillas de Prosopis velutina, que no es su huésped. Para las investigaciones futuras sobre los brúcides de Prosopis resulta significativo que brúcides de este agrupamiento puedan efectivamente alimentarse con semillas de Prosopis en el ambiente natural y en depósitos.
Una cantidad de estudios recientes han descrito la ecología y las interacciones evolucionarias entre los brúcides y las plantas que los hospedan (Janzen, 1969, 1971 a, b, c, 1980; Forister 1970; Center y Johnson 1974; Johnson y Slobodchikoff 1979; Johnson 1981c). Según Janzen (1969), la mayoría de las legumbres parece seguir una o dos estrategias para evitar la depredación de semillas por parte de los brúcides. Algunas plantas producen semillas grandes, tóxicas, lo que excluye todos o casi todos los depredadores de semillas. Otras plantas producen semillas muy pequeñas que son comidas por los brugos, pero son tantas las semillas producidas que algunas escapan al pillaje. Center y Johnson (1974) y Johnson (1981c) ofrecieron evidencias que prueban que los brúcides han desarrollado mecanismos que se contraponen a las formas protectivas de las plantas. Por ejemplo, algunos brúcides pueden alimentarse con semillas muy tóxicas, mientras que otros comen semillas muy pequeñas, una vez que se han dispersado. Janzen (1969) analizó alrededor de 31 mecanismos protectivos que las plantas han generado para defenderse de la depredación de los brugos. Aparentemente, las especies de Prosopis han desarrollado la estrategia de producir una muy grande cosecha de semillas, que asegure que al menos algunas de ellas evitarán la depredación de los brúcides.
Los mecanismos más específicos que las legumbres usan para minimizar los efectos del pillaje por parte de los insectos que comen las semillas son que los árboles que crecen en ambientes relativamente constantes, uniformes, tienen la posibilidad de espaciar los momentos de fructificación de los individuos de manera que las poblaciones de insectos no pueden acumularse, la formación de semillas carnosas que se secan demasiado rápidamente como para que un depredador pueda usarlo como alimento larval, o la producción de semillas que son tan pequeñas con muy poco alimento para permitir que se complete el desarrollo del insecto. Según Kingsolver et al., 1977, ninguna de estas posibilidades se ofrece a Prosopis en ambientes desérticos, ya que la fructificación de todos los individuos está sincronizada para que tenga lugar en el momento más ventajoso del año para la germinación de la semilla, y la semilla debe estar en condiciones de soportar la desecación como también de proporcionar una nueva fuente inicial de energía a las nuevas plántulas. Debe investigarse más antes de aceptar o rechazar la hipótesis de Kingsolver et al. (1977).
La mayoría de las especies de brúcides son sumamente específicas con respecto a su huésped y la mayor parte de los huéspedes mantienen pocos brúcides en sus semillas. Johnson y Slobodchikoff (1979) encontraron que el 82,5% de los brúcides que se alimentan sobre Cassia tienen 3 o menos huéspedes y el 90,5% de las especies de Cassia tienen 3 o menos especies de brúcides en sus semillas. Johnson (1981b) mencionó que alrededor del 70% de los brúcides del género Acanthoscelides tiene 1–2 huéspedes y el 74% de sus huéspedes tiene 1–2 especies de brúcides que se alimentan con sus semillas. Resultados similares fueron presentados por Janzen (1980) cuando pasó a analizar la especificidad del huésped por parte de los gorgojos de Costa Rica, que se alimentan con semillas.
![]()
![]()
![]()