Pese a que los diagramas no pueden ser sino complementos de los modelos númericos, uno de los aspectos que se ha destacado en este documento es la utilidad de las aproximaciones en forma de diagrama para plasmar la información acerca de las interacciones y de la red alimentaria del ecosistema. A pesar de la variedad de pautas utilizadas a la hora de elaborar los diagramas de flujo, hay dos aspectos de las relaciones tróficas en el mar que normalmente no son representados, pese a su gran importancia. El primero de estos aspectos representa la distribución espacial de la trasmición de energía - un tema principal en este documento al considerar la red alimentaria como una “estructura de disipación”, en la que las áreas específicas y los habitats juegan un papel clave en la producción y en la diseminación de biomasa (tomemos la Figura 3 como ejemplo de una representación espacial de los vínculos de la red alimentaria, y Laevastu y Larkins (1981) como discusión de la importancia de las distribuciones espaciales y de los factores hidrográficos). Una segunda característica de la mayoría de redes alimentarias marinas que comúnmente no es representada, consiste en que la mayoría de componentes en todos los niveles tróficos inician su vida en rangos de talla similares, y los depredadores apicales, por ejemplo, se mueven de forma inevitable, tróficamente hacia arriba a lo largo de la vida. La conclusión es que las relaciones con la talla (con referencia a las consideraciones espaciones espaciales anteriores), son esenciales para comprender los procesos del ecosistema, pero rara vez aparecen representadas en las redes alimentarias. Todo ello nos incita a sugerir de forma tentativa una aproximación a la representación de redes alimentarias que tome en consideración estos elementos espaciales y relacionados con la talla, junto con el otro elemento informativo importante, la biomasa de las especies.
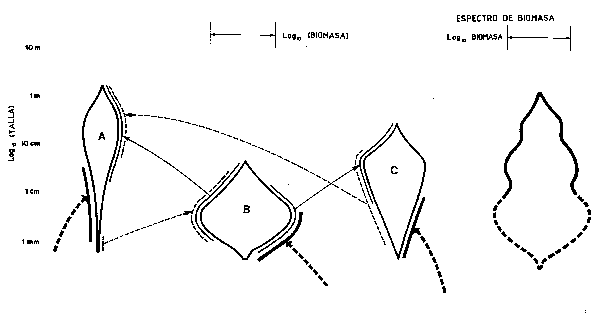
La Figura 68 pretende mostrar de forma gráfica la relación entre un depredador dominante (A), uno subdominante (B) y su presa común (C) evidenciando las interrelaciones con la talla además de la biomasa (ambas en una escala logarítmica). La posibilidad de una depredación “recíproca” de las presas adultas sobre los juveniles de sus depredadores (es el ejemplo del bacalao y el arenque), es fácilmente situable en este planteamiento. También es posible sumar todas las biomasas (no sus logarítmos!) lateralmente para obtener el espectro de biomasa del susbsistema en cuestión. Los diagramas tipo “cometa” de la Figura 68 son, obviamente para cada especie, “réplicas” del espectro de su biomasa. Tales espectros de biomasa (en este caso, obviamente incompleto para los niveles tróficos inferiores), deberían establecerse antes y después de que se hubieran explotado uno o más componentes y son análogos a la pirámide de números mostrada en la Figura 2.
Figura 68 Red alimentaria de tres especies, con el logaritmo de la biomasa representado por la “anchura”,y flechas que indican los intercambios tróficos entre los diferentes rangos de tallas. Se muestra el control recíproco del reclutamiento larvario del “depredador” A por la “presa” B(¿Planctófaga?). También se incluye el espector de biomasa para el ecosistema en su conjunto . (Las flechas sin origen indican elementos indeterminados situados más abajo en la red)
Diferente Vulnerabilidad Frente a la Pesca
La mayor parte de la teoría referida a pesquerías, como hemos venido señalando, aún gira entorno de la asunción de que los componentes individuales pueden ser ordenados con independencia de sus presas, depredadores y competidores, y el mantenimiento de esta escuela tanto en sus facetas teóricas como prácticas a lo largo de medio siglo desde los primeros trabajos de Baronov, demuestra que puede no ser una perspectiva totalmente incorrecta en algunos ecosistemas y en muchos casos. Puede valer la pena especular que esto podría ser cierto a causa de la naturaleza “amortiguada” de muchos de los ecosistemas marinos, en los que los componentes individuales de la red alimentaria tienden a solaparse con otros del mismo sistema generando una cierta elasticidad en la red alimentaria ante la explotación de uno de los componentes, o quizás porqué a niveles de explotación moderados, la producción de las especies sometidas a explotación se ve de hecho incrementada por la pesca.
Habitats Críticos y Refugios
Al tratar de establecer un orden de prioridad, en lo que a nosotros afecta de la ordenación de especies en el contexto del ecosistema, la complejidad de los ecosistemas marinos hance que resulte problemático, excepto en el caso de sistemas muy sencillos. En un futuro próximo será posible dar razón de un impacto dado como consecuencia de una intervención particular en un “nudo” específico de la red alimentaria. Así, aunque por ejemplo, por medio de una pesca selectiva, teóricamente podemos eliminar los componentes “indeseables”, tal operación podría ser de difícil justificación desde un punto de vista económico. Basándonos en algunos ejemplos en que ésto se ha llevado adelante, como es el caso de la eliminación del “dogfish” (un depredador de los salmónidos, de poco valor) en el Pacífico noroeste por medio de un sistema de “subvenciones”, o el control de depredadores como las estrellas de mar y los gasterópodos en los cultivos de moluscos, se puede decir que tales esfuerzos rara vez se ven recompensados por un éxito duradero, incluso en el caso (como ocurre con frecuencia) de que subsecuentemente surja un mercado para estos productos “engorrosos”.
Experiencias a gran escala de manipulación de ecosistemas han sido realmente desarrolladas en todo el mundo, especialmente en pesquerías comerciales de carácter monoespecífico. Aunque estas experiencias no fueron dirigidas a la erradicación de especies, en una proporción relativamente baja de casos encontramos factores diferentes a un descenso en los rendimientos económicos por la baja de las abundancias, habiendo intervenido para proteger las especies objetivo de la extinción. Se han dado casos, por supuesto, en que se ha provocado la extinción o la drástica disminución de especies de valor (en especial de aquellas muy vulnerables y larga vida), pero también ha habido un número sorprendente de casos que han mostrado un significativo grado de elasticidad ante estas intervenciones.
Un examen detallado de ambas categorías parece mostrar que se dan dos tipos de intervenciones que con mayor probabilidad pueden provocar la caída del stock y la situación de sobrepesca:
(a) La fase explotada de una población pasa por un período (generalmente cuando el stock está concentrado en un área dada), en que es muy vulnerable a la pesca, estas áreas pueden ser denominadas “habitats críticos”. Ejemplos comunes de habitats críticos son las áreas de puesta de especies tales como los cefalópodos, el arenque y las tortugas marinas en que la mayor parte de la biomasa adulta se concentra en un área reducida por un corto período de tiempo, siendo vulnerable a altos niveles de explotación en que el éxito de pesca es francamente independiente de la biomasa del stock.
(b) La especie pasa por una fase en la que su desarrollo está asociado a un habitat en particular, por ejemplo el manglar y las lagunas litorales en el casó de algunas especies de gambas y cangrejos, praderas submarinas o asentamientos de macrofitos para algunas especies de langosta, etc. La “salut” de las “especies substrato” tiene en consecuencia una influencia fundamental en los organismos asociados.
En sentido contrario, si las especies resisten una fuerte explotación, puede ser porque la eficiencia en la captura es proporcional a la densidad o porque existe un “refugio” que protege a la especie de la fuerte explotación en una parte de su rango de tallas y de su distribución geográfica.
Un “refugio” no es fácil de definir, pero puede consistir en una zona del fondo de rocas, no rastreable, o en el caso de una pesquería de pequeñas embarcaciones, en una zona de fuertes corrientes y alejada de la costa. También puede darse el caso de que se faene con aparejos tales como nasas o redes de enmalle que tienen una selectividad de tallas particular, de tal forma que la luz de la malla dominante o la dimensión de la boca de entrada del aparejo constituya un “refugio” para aquellos individuos que evitan la captura el tiempo suficiente para alcanzar las tallas mayores, correspondientes a la madurez.
Distribución de la Biomasa por Clases de Talla y Reclutamiento
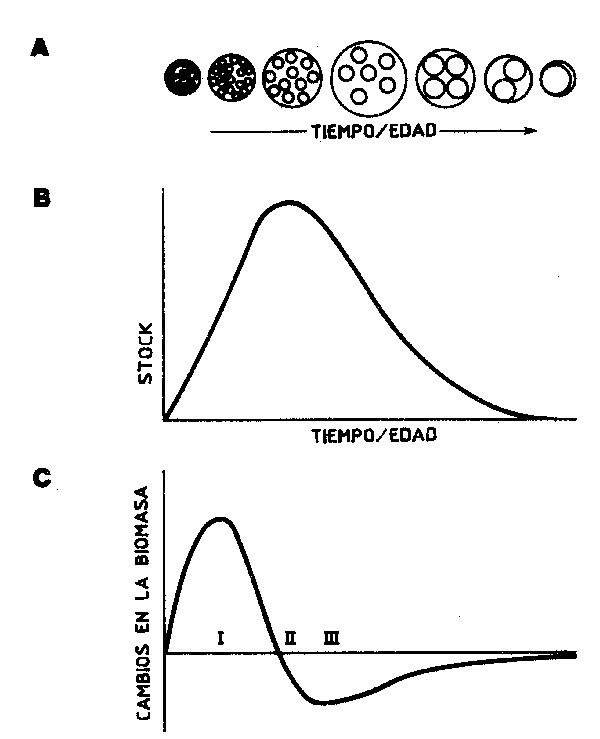
La importancia de la información acerca del espectro de tallas de una especie para el análisis de las tendencias, y la estimación de los parámetros de la población, va siendo en los últimos años más apreciada, y el equivalente en términos de biomasa tiene una importancia análoga de cara a los cálculos tróficos. Las distribución de la biomasa por clase de talla y de edad de una especie muestra un incremento hasta una edad/talla en que as máxima, cuando el crecimiento se ve compensado por la mortalidad debida a todas las causas, después va cayendo al aumentar la talla (Figura 69). En el caso de recursos sometidos a explotación, este punto de biomasa máxima sería la talla ideal para llevar a cabo una explotación instantánea. En la práctica, no obstante (y aquí entran en juego las metodologías de rendimiento por recluta de, por ejemplo, Beverton y Holt, 1957), es corriente para una tasa de mortalidad natural dada (M) que se ha asumido como constante para el rango de tallas sometido a explotación, calcular la tasa de explotación que puede proporcionar un rendimiento por recluta máximo para una talla de primera captura dada, o los efectos de un cambio de malla o de la talla de primera captura. Esta approximación no tiene en cuenta el posible impacto que una fuerte explotación podría causar sobre las poblaciones del depreddor o de la presa de la especie en cuestión.
|
Figura 69 (A) Evoloción de una cohorte con la edad partiendo de numerosos
individuos pequeños hasta llegar a unos pocos de gran talla.
(B)Evolución del stock de esta misma cohorte con el tiempo o la edad (C) Tasa de cambio en biomasa (dB/dt) con relación a la edad(II indica la edad de biomasa máxima, con las tasas de crecimiento y mortalidad utilizadas, y I Y III indican respectivamente el momento en que la biomasa está creciendo o cayendo más rápidamente |
Otra de las limitaciones de este tipo de cálculos conmayor número de implicaciones prácticas, consiste en que la predicción de efectos de un cambio en la talla de primera captura no siempre tiene en cuenta si la especie va a empezar a ser capturada antes de alcanzar la madurez sexual o no. Esto afectará lógicamente al nivel medio de reclutamiento, si en un caso extremo la explotación es intensa y se inicia antes de la madurez. Este tipo de efecto es tratado por medio de los modelos llamados de stock-reclutamiento (para más detalles ver Ricker, 1975). Estos modelos que normalmente intentan predecir el nivel medio de reclutamiento partiendo solamente del tamaño del stock reproductor, generalmente no tienen en cuenta las variaciones en el medio físico y biótico al que están sujectos el stock reproductor y los prereclutas, y en consecuencia la varianza en la predicción del número de reclutas es elevada. Lo que es importante señalar desde la perspectiva del análisis trófico es, sin embargo, que la “pérdida” de individuos de talla comercial en las evaluaciones de renbdimiento por recluta a causa de la mortalidad natural (la proporción M/Z de todas las muertes) supone un retorno de materia a la red alimentaria. Esto sólo es cierto en el caso de modelos de stock reproductor-reclutamiento, de modo que aunque el número de reclutas producidos por unidad de biomasa de reproductores cae al aumentar la biomasa total de reproductores, las “pérdidas” en forma de gametos y prereclutas que conlleva, se incorporan de nuevo a la red alimentaria, si bien por lo general en una talla de prereclutas menor.
En términos prácticos, no obstante, a pesar de que no deberíamos abandonar los tan útiles métodos monoespecíficos que se han desarrollado durante los últimos 50 años, es necesario poner cierto interés en situar este tipo de análisis en su contexto ecológico correcto, es decir que la evaluaciíon de cada especie debería ir precedida de un estudio cuantitativo acerca de:
(a) El sistema físico, y lo que nosotros conocemos de su estabilidad, estacionalidad, extensión y productividad.
(b) Que conocemos acerca de los principales componentes biológicos del sistema y acerca de sus interacciones (tróficas o de otro tipo) con la especie en cuestión.
(c) La variabilidad de la producción del recurso en cuestión y de otros recursos clave considerados en (b), y alguna idea sobre si por lo general la biomasa presente está en los niveles superior, medio o inferior del rango histórico.
Las evaluaciones monoespecíficas por lo tanto pueden convertirse en una valoración cualitativa que ponga los “condicionantes” y los “peros” a las conclusiones numéricas, de tal forma que el administrador pueda establecer algunos juicios acerca de su posible aplicación y por cuanto tiempo puede aplicarse. Esta valoración debería establecerse en base a una experiencia procedente de fuera de la pesquería en cuestión, y podemos sugerir que no sería prematuro ver retrospectivamente la historia de la pesca industrial a lo largo de las últimas décadas. Un rápido examen (por ejemplo, Caddy y Gulland, 1983) puede sugerirnos que la “teoría de las catástrofes” constituye un concepto bastante más aplicable, en algunos casos, que el de la “población en equilibrio”.
Algunas de las primeras secciones se han referido a los components del ecosistema y sus características, lo cual puede habernos inducido a caer en la errónea impresión de que las características de un ecosistema son simplemente la suma de las propiedades de sus componentes. Esto no es exacto; ya que se dan “características emergentes” del ecosistema que no pueden deducirse de las de los componentes individuales. De tal forma que a un nivel más práctico, el rendimientos multiespecífico del sistema está significativamente por debajo de la suma de los rendimientos máximos de cada especie en el supuesto de que éstas fueran explotadas de forma individual. Mann (1982) señala que los diagramas estáticos de las redes alimentarias, en particular los modelos de compartimentos, están condenados al fracaso como predictores cuantitativos, en gran parte porque las características emergentes de estos sistemas no se evidencian, aunque estos diagramas pueden seguir siendo útiles para describir cómo la energía fluye a través del sistema.
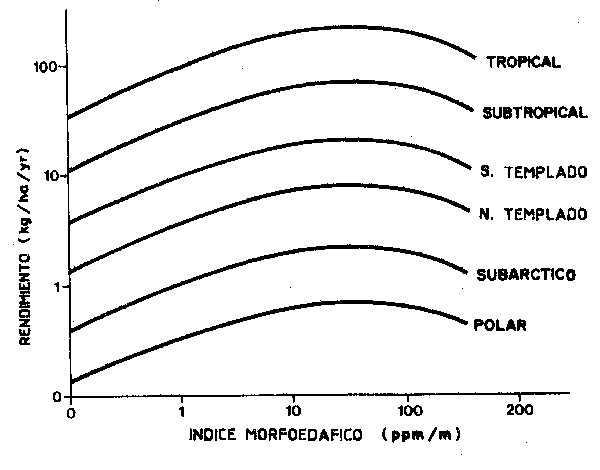
Esto nos lleva a considerar la principal aproximación alternativa, es decir la aplicación de aproximaciones empíricas a la evaluación de los ecosistemas marinos. Podemos fijarnos, por ejemplo en el equivalente marino al índice morfoedáfico (Figura 70), que estima la producción de peces de agua dulce en lagos en función de factores físicos fácilmente medibles. Esto por supuesto encierra dos problemas fundamentals, el primero consistirá en que los sistemas marinos son todos “ilimitados” en diverso grado, el segundo que los cambios en el “estado” del subsistema en que se haya basado el índice trastornan las series históricas de captura y de información medioambiental, obligando a una revisión de las cuestiones estructurales fundamentales.
|
Figura 70 Predicción de curvas de rendimiento en función
del índice morfoedáfico para grupos de lagos en
seis zonas climáticas del mudo (adaptación de
Henderson, Ryder y Kudhongania, 1973) |
Diversos ecólogos se han dedicado a las características emergentes de los ecosistemas, y a pesar de que quizás la ley más fundamental de la ecología sea la más generalizadora y la que conlleve el mayor número de excepciones, las perspectivas que ofrezca demostrarán esperanzadoremente la utilidad de analizar las respuestas a nivel del ecosistema.
Un posible punto de partida sería el planteamiento de Margalef (1978) que sugiere que “todas o la mayoría de las formas con que el hombre puede interferir en el resto de la naturaleza producen efectos coincidentes o paralelos” … “la diversidad se ve reducida y la relación producción/ biomasa se incrementa”. Si esto es cierto, supone realmente una simplificación de muchos problemas aparentemente diversos, y la idea ha sido desarrollada pudiendo con ello abrir un nuevo campo de trabajos, que podría ser la “ecología del stress” (Barrett, Van Dyne y Odum, 1976). Algunos conceptos básicos y la terminología que se ha utilizado (Rapport, Regier y Hutchinson, 1985) provienen de trabajos sobre respuesta de organismos individuales al stress. Este término, impreciso en este contexto, se asocia a la idea de una perturbación impuesta en el sistema partiendo de una situación dada y llevándolo probablemente lejos del estado de equilibrio o de “seudoequilibrio” original, en el sentido en que estos términos son utilizados en la dinámica de poblaciones. Raport, Regier y Hutchinson (1985) destacan los tres estados postulados por Selye (1974) acerca de la respuesta de un organismo, es decir de “una reacción de alarma”, seguida de un estado de resistencia al stress” y finalmente el “agotamiento” o la sumisión pasiva al stress, como si existieran paralelismos desde un punto de vista cibernético a nivel del ecosistema (Figura 71).
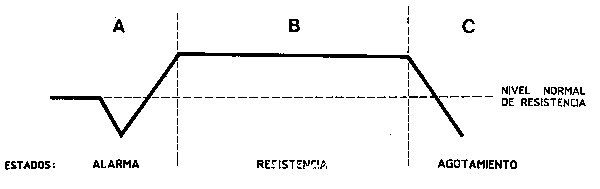
Figura 71 Tres fases del síndrome de adaptación general observados en los organimos.(A) reacción de alarma, (B) estado de resistencia, (C)estado de agotamiento (Rapport, Regier y Hutuchison, 1985)
Los impactos humanos sobre el sistema ecológico podrían incluirse en cinco tipos de stress, los cuales no son exclusivos de la interferencia humana, dado que causas naturales también pueden producir efectos similares:
(1) explotación;
(2) descarga de productos contaminantes (quizás podríamos distinguir los tóxicos producidos por el hombre a los que no se aplica esta generalización de la contaminación orgánica natural);
(3) modificaciones en el terreno, batimetría, grado de protección, etc. (por ejemplo el desarrollo del litoral, demandas de terreno, dragados y extracción de arena, etc.);
(4) introducción de especies exóticas;
(5) acontecimientos devastadores (huracanes, vertidos de petróleo, contaminación costera, etc. ver el apartado (2)).
En el apartado (5) los autores incluyen las guerras también como catástrofes naturales (no obstante de paso debemos señalar aquí que la información más completa acerca de la tasa de recuperación de un sistema pesquero del stress, y que ha servido al desarrollo de la teoría de la dinámica de poblaciones, proviene del cese de la pesca en el Atlántico noreste durante las dos guerras mundiales).
Los cambios por stress del ecosistema, diagnosticados por Rapport, Regier y Hutchinson (1985), incluyen:
(1) un incremento en la tasa de pérdida de nutrientes del sistema (es decir la parcial destrucción de la estructura de disipación o red alimentaria);
(2) cambios en la productividad primaria (como ocurre en las zonas de afloramiento o cuando se produce un incremento en el arrastre de sedimentos o fertilizantes en las zonas litorales debido al desarrollo agrícola);
(3) incremento en la relación producción/biomasa;
(4) disminución de la diversidad de especies;
(5) regresión. Definida como un cambio en la composición de especies debido a la mejor adaptación de algunos organismos a las nuevas y más difíciles condiciones medioambientales, en nuestro contexto, es el caso de los organismos de vida corta r-seleccionados, como se señaló anteriormente. (Si lo vemos desde el punto de vista de la sucesión ecológica y del concepto de clímax en las comunidades biológicas, ésto implicaría un desplazamiento hacia comunidades “menos maduras”);
(6) cambios en la composición de tallas, generalmente hacia tallas medias menores y períodos de vida más cortos. De nuevo tenemos la corta vida de las especies r-seleccionadas y la elevada relación P/B mencionada en (3). (No obstante en sentido contrario un fallo en el reclutamiento puede dar lugar a un incremento de la talla media lo que la convierte en un índice del stress de la población);
(7) otras señales de peligro tales como el “síndrome de peligro biológico” pueden aparecer e incluirían todo lo anterior, más enfermedades, modificación de la tasa de crecimiento y mortalidades masivas.
En relación a la pesca, los cambios en la relación tiempo de búsqueda/tiempo de pesca (por ejemplo, Condrey, 1984), o en el número y localización de áreas de puesta, pueden ser indicadores de situaciones de stress o de cambios en el tamaño de la población, su distribución y abundancia. La desaparición o progresiva escasez de las especies indicadores de sistemas no sometidos a stress, y su sustitución por especies paralelas más resistentes, puede ser otra forma de “reacción de alarma” de las anteriormente mencionadas. Estos cambios también se ponen en evidencia en el mercado local de pescado por medio del aumento del precio relativo de las especies populares (sometidas a stress) y por su sustitución por especies alternativas (consideradas anteriormente de menor calidad). De nuevo los cambios sugerentes del ecosistema, pese a tratarse en este caso de cambios en el mercado o en las pautas de consumo pueden constituirse en agentes causales de una explotación más intensa.
Quizás el menos optimista, aunque bastante realista, punto de vista del uso potencial de las aproximaciones corrientes al modelado de las interrelaciones tróficas, es el planteado por Gulland y Garcia (1984), que en relación a la cuestión de la interacción y estabilidad en pesquerías multiespecíficas destaca que a pesar de que estas cuestiones merezcan un incremento práctico y atención teórica, los estudios teóricos no han ido muy lejos excepto en cuanto a “abrir la puerta de la complejidad de los problemas interrelacionados”. Una visión similar fue expresada por Sainsbury (1982) quien después de revisar los modelos matemáticos utilizados, incluido el seminario realizado por el ICLARM/CSIRO en 1981, concluye que “un detenido examen de la teoría ecológica y de pesquerías indica que en el momento presente no existen modelos ecológicos adecuados a la dinámica de la comunidad, y que de entre las aproximaciones revisadas no destaca un cuerpo coherente de reglas y métodos aplicables a la ordenación de las pesquerías multiespecíficas tropicales. Como se ha señalado, con cierta frecuencia la complejidad del problema proviene de la aproximación científica adoptada por el mundo académico, que da una notable prioridad a la innovación frente a los trabajos de síntesis, dado que como señaló Kuhn y demuestra perfectamente la teoría ecológica, la ciencia consiste en una serie de disciplinas separadas, compitiendo cada una de ellas en la publicación de trabajos y disputándose los presupuestos en el “mercado de la ciencia”.
A pesar de la visión pesimista que se acaba de plantear acerca de la posibilidades de producir un modelo multiespecífico generalizado en breve (lo que constituye un proyecto ambicioso dada la amplia diversidad de pesquerías multiespecíficas; ver, por ejemplo, Pauly y Murphy, en Gulland y Garcia, 1984) ellos no implica que una aproximación dirigida a una pesquería en particular no pueda resultar en cierto grado exitosa. Lo que se ha intentado en esta revisión es ilustrar algunas de las ideas y conceptos que deben ser tenidos en cuenta en tales aproximaciones. No obstante resulta bastante evidente que los sistemas multiespecíficos no pueden ser abordados, con ciertas posibilidades de éxito, a menos que se establezca un importante compromiso, preferentemente con unas bases de cooperación amplias.
Así aunque no existen fórmulas sencillas apropiadas, parece que hay ciertas directrices generales para proceder al análisis de los sistemas pesqueros multiespecíficos de una forma objectiva que pueden demostrar ser útiles en una situación dada, pero un plan de acción establecido aplicable a todas las circunstancias no es factible.
(1) La primera sugerencia que hacemos va dirigida a establecer categorías entre las principales asociaciones o comunidades presentes (separando las especies transeúntes de las residentes), y cartografiarlas sobre un mapa de caladeros. Incluso en el caso de que lo hagamos de forma preliminar, mostrará las relaciones espaciales de estas asociaciones con referencias a los principales puertos pesqueros, puiendo constituir la base de un sistema estadístico pesquero que en la medida de las posibilidades pueda determinar las capturas efectuadas en cada caladero separadamente. Las concepciones derivadas pueden facilitar la caracterización de las zonas litorales, de las masas de agua y de las áreas locales de alta producción.
(2) Si es posible, las series históricas de datos de la pesquería, incluidas las informaciones de carácter anecdótico proporcionadas por los pescadores, así como la información proveniente de la revisión de la bibliografía disponible, nos debe ayudar a establecer una base informativa referida a las especies clave, sus hábitos, interrelaciones tróficas y ciclos vitales. La existencia de cambios importantes en la abundancia y en la dominancia de especies en el pasado (incluso de carácter cualitativo) debe quedar documentada, junto con la información específica acerca del esfuerzo pesquero ejercido en cada área, tamaño de la flota, caladeros y serie histórica de capturas, así como todos los datos históricos referidos a la tasa de captura y a la composición de tallas que se puedan encontrar.
(3) Siempre que sea posible, sin que ello suponga mayores dificultades, también se pueden recoger informaciones de contenidos estomacales de las especies comerciales clave referidos a tallas, estaciones y áreas determinadas a fin de configurar una idea de las interacciones de las especies en el sistema.
(4) Un aspecto clave de la aplicación de los planteamientos ecológicos a los problemas pesqueros consiste en la identificación de las fuentes de producción biológica principales y de las áreas en que se sitúan estos centros con relación a las actividades desarrolladas por el hombre.
(5) Una posible sugerencia que deriva de estas consideraciones es la de establecer un mapa de habitats críticos y de los centros de las “estructuras de disipación” del ecosistema, tal como se ha representado en la portada de este trabajo, con el fin de que se de una gran prioridad a su conservación, incluso en el caso de que los detalles de las interacciones de la red alimentaria todavía no se hayan resuelto. Este tipo de actuaciones pueden calificarse como de conservación de “opciones de futuro”, una vez que los estudios ecológicos de la pesquería hayan sido realizados.
Estrategias de Explotación de Ecosistemas Multiespecíficos
Las estimaciones de producción pesquera total basadas en las estadísticas pesqueras, están posiblemente subestimadas con referencia al potencial que podría obtenerse si la pesquería fuese explotada con diversos artes de pesca de forma equilibrada con el fin de alcanzar el máximo rendimiento (Marten y Polovina, 1982). ¿Cual es la estrategia de explotación que se ajusta a esto?.
Teóricamente, una red alimentaria puede mantenerse “en equilibrio” pescando cada uno de sus componentes en la proporción a la tasa de depredación natural a que está sometido.
Esta estrategia de explotación “utópica” de una red alimentaria puede considerarse equivalente a una situación en que:
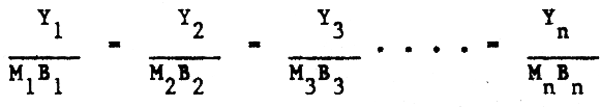
es decir que una proporción constante d ela producción total pi = MiBi para las especies i = 1,2,3 …. n sea retirada (capturada) como rendimiento (Yi), donde Mi es la tasa de mortalidad natural, y Bi la biomasa. Esto es equivalente a situar la relación Fi/Mi a un nivel constante para cada especie, lo que supone una forma de control del esfuerzo pesquero para cada especie.
Con referencia a cualquier tipo de esquema simplificador se pueden hacer toda una serie de objeciones lo mismo que con respecto a su aplicación práctica:
¿De qué manera se podrían medir todos los parámetros con precisión, sin modificar la tasa de explotación de cada especie?
¿Cuáles serían los niveles “de seguridad” de Fi/Mi a utilizar?
Caddy y Csirke (1983) demuestran que los valores de FRMS/M encontrados en la biobliografía varían considerablemente y que un buen número de valores se sitúan por debajo de 1 e incluso valores del orden de 0,33 podrían resultar apropiados a algunos grupos utilizando el escaso número de estimaciones de los parámetros de que se dispone.
Obviamente como hemos visto resulta peligroso asumir que la abundancia o incluso la naturaleza de los componentes de una red alimentaria sea constante. Con una relación F/M demasiado elevada la sustitución de especies podría ser otra posibilidad, especialmente si la talla media de los componentes de la red alimentaria cambia, con lo que cambia la posición efectiva de la especie en la red, generando un excedente de organismos disponible como alimento lo que permite la colonización del ecosistema por “nuevas” especies o grupos de depredadores.
La idea de que se puede incrementar la producción total del sistema retirando los depredadores superiors y procediendo después a explotar las especies “presa” situadas por debajo, en la red alimentaria, intensivamente con el fin de obtener grandes rendimientos, parece que, dado que muchas de las especies de peces marinos de mayor talla recorren secuencialmente los diferentes niveles tróficos a lo largo de su vida, no funciona en la mayoría de los casos.
Al parecer (Regier y Henderson, 1973; Ursin, 1982) al incrementarse la actividad pesquera ejercida sobre los depredadores dominantes o apicales, el papel de agente controlador es asumido, por lo menos en parte, por el depredador inferior más próximo, los individuos más viejos que pueden situarse en una categoría trófica superior a la ocupada en un principio. En realidad, las experiencias llevadas a cabo en grandes lagos (Regier y Loftus, 1972) demuestran que eliminando los grandes depredadores el rendimiento se incrementa en un porcentaje inferior al 100 %, lo que no se ajusta a las predicciones de 5 a 10 veces obtenidas por medio de experiencias de laboratorio en las que se asumía que la composición de la dieta de los peces era fija.
Una estrategia alternativa consistente en la pesca indiscriminada de todas las especies con un arte de pesca de amplio espectro del tipo de los artes de arrastre, favorecería a las especies oportunistas poco exigentes en relación al nicho ecológico y con una elevada fecundidad. No obstante la experiencia demuestra que las respuestas del sistema a la pesca no son fácilmente predecibles, puesto que el éxito de la actividad pesquera al ser una actividad de carácter económico depende de a cual de los dos tipos de depredadores de amplio espectro se esté favoreciendo: los de escaso valor como los peces espinosos o los de elevado valor como son los peces tipo bacalao.
Pauly (1979) resumió algunas de estas “opciones de ordenación de las redes alimentarias” de forma semihumorística, y tomamos algunas de estas formas que frecuentemente se dan en el mundo real y que por lo menos de forma implícita han sido tratadas anteriormente. Haciendo uso de los títulos descriptivos utilizados por Pauly, seleccionamos los siguientes:
(1) La estrategia “inexistente” o “nula”:
Es el caso en que el recurso es dejado en su estado virgen.
(2) La estrategia de “los túnidos”:
Básicamente consiste en tomar el RMS de los depredadores apicales pescando los mayores ejemplares utilizando para ello métodos selectivos. Como ya ha sido indicado por May et al., esta estrategia, si bien no proporciona el máximo rendimiento del ecosistema, es sostenible.
(3) La estrategia “del mar del Norte”:
Pauly (1979) caracteriza esta estrategia por la sistemática sobreexplotación de los depredadores apicales hasta que la depredación ejercida sobre las presas se hace insignificante. Ello conduciría a un máximo en la biomasa de las presas (ver sin embargo la hipótesis de Ursin!) pero en ese momento este aumento de biomasa, es contrarrestado por la pesca ejercida sobre las presas, con lo que su producción es trasferida a la captura. Esta puede ser la estrategia más productiva en latitudes situadas más al norte donde encontramos un buen número de comedores generalizados, pudiendo incluso resultar sostenible en estas áreas.
(4) La estrategia del golfo de Tailandia:
Esta consiste en pescar tanto depredadores como presas con artes de malla muy pequeña y con niveles de esfuerzo muy altos. El resultado, es el colapso tanto del stock del depredador como del de la presa, incrementándose el plancton y la biomasa bentónica y una, en cierta forma generalizada, rápida reproducción de especies r-seleccionadas, tales como calamares y pequeños peces planos.
También se pueden considerar otras variantes, pero estas tres estrategias configuran de alguna forma todo el espectro posible, incluyendo la “estrategia nula” la cual curiosamente es practicada conjuntamente con una de las otras, es decir, preservando parte de los ecosistemas vírgenes intactos calificándolos como reservas o parques marinos.
En un apartado anterior se postuló una estrategia diferente que se denominó estrategia “Utópica”, en la que se pretende equilibrar la explotación en su conjunto, por medio de una pesca selectiva de todos los componentes que asegure el equilibrio multiespecífico. Quizás la pesquería litoral japonesa constituya el caso más próximo a la realidad de esta estrategia. Esto puede poner en evidencia la utilidad de este concepto que se debe tener presente a la hora de discutir alternativas multiespecíficas de ordenación.
En la realidad, el mejor planteamiento puede ser una combinación de estrategias. Por ejemplo, una combinación de (1) y (3) o de (1) y (4) son posibilidades a considerar, en cuyo caso algunos habitats críticos permanecerían completamente cerrados a la pesca, o por lo menos lo estarían en períodos estacionales.
Con el fin de comparar la posibilidad de manipulación de un ecosistema (o de la vulnerabilidad de los ecosistemas a ser modificados), existe una marcada tendencia a clasificar los diversos tipos de intervenciones propias de ecosistemas de mar abierto y que hemos discutido, en el siguiente orden de posibilidad e impacto:
| Posibilidad | Tipo de manipulación | Vulnerabilidad |
|---|---|---|
| Alta | - Impactos que afectan a los habitats críticos | Alta |
| Alta | - Impactos en las especies “sustrato” | Alta |
| Media | - Impactos que afectan a las principales fuentes de producción (estructuras de disipación) | Media |
| Baja-Alta | - Introducción de vectores de enfermedades y especies exóticas | (Depende de la situación) |
| Media | - Control de la depredación | Media |
| Baja | - Control de los competidores | Baja |
| Baja | - Control de las presas | Baja |
El anterior orden de posibilidad encierra, con excepciones obvias, un alto grado de subjetividad, pero está basado en la conclusión de que las manipulaciones de especies, con la excepción de las exóticas, cuya introducción en algunos ecosistemas marinos han tenido consecuencias dramáticas (y por lo general efectos adversos), son de una efectividad más incierta que la manipulación de habitats críticos y de las especies substrato (como arrecifes de coral, manglares y praderas submarinas). Igualmente se considera que la protección o mejora del habitat probablemente resultará más efectiva en la ordenación de recursos, que la intervención directa sobre las redes alimentarias controlando las especies de forma individual tal como se indica en los últimos casos de los tipos de manipulación referidos anteriormente. La protección de los habitats críticos posee la ventaja de que no precisa muchos conocimientos detallados de las interacciones entre especies, y no impide que en el futuro cuando sean factibles se puedan realizar intervenciones más sofisticadas sobre la red alimentaria.
ACMRR/IABO 1976 Working Party on Ecological Indices of Stress to Fishery Resources, Indices for 1976 measuring responses of aquatic ecology to various human influences. A report of the ACMRR/IABO Working Party on ecological indices of stress to fishery resources. FAO Fish.Tech.Pap., (151):66 p.
Allen, K.R., 1971 Relation between production and biomass. J.Fish.Res.Board Can., 28(10):1573–81
Allen, P.M. y J.M. McGlade, 1986 Dynamics of discovery and exploitation: the case of the Scotian Shelf groundfish fisheries. Can.J.Fish.Aquat.Sci., 43(6):1187–200
Alvarino, A., 1980 The relation between the distribution of zooplankton predators and anchovy larvae. Rep.CCOFI, (21):150–60
Alverson, D.L. y M.J. Carney, 1975 A graphic review of the growth and decay of population cohorts. J. Cons.CIEM, 36(2):133–43
Amaratunga, T., 1983 The role of cephalopods in the marine ecosystem. In Advances in assessment of world cephalopod resources, edited by J.F. Caddy. FAO Fish.Tech.Pap., (231):452 p.
Anderson, J.M., 1981 Ecology for environmental sciences: biosphere, ecosystems and man. Nueva York, Halstead Press, Resource and environmental science series, 195 p.
Anderson, K.P. y E. Ursin, 1977 A multispecies extension to the Beverton and Holt theory of fishing, with accounts of phosphorus circulation and primary production. Medd.Danm.Fisk. Havunders., 7:319–435
Armstrong, M.J., 1982 The predator-prey relationships of Irish Sea poor-cod (Trisopterus minutus L.), pouting (Trisopterus luscus L.) and cod (Gadus morhua L.). J.Cons.CIEM, 40(2):135–52
Arnold, G.P., 1979 Squid: a review of their biology and fisheries. Lab.Leafl.Dir.Fish.Res.G.B., (48):39 p.
Bacon, P.R., 1984 Overview of mollusc resources and exploitation in the Caribbean region. Documento presentado a 37th Meeting of the Gulf and Caribbean Fisheries Institute, y a Ninth Mexus-Gulf Cooperative Research Conference, Cancun, México, noviembre 1984 (mimeo)
Bagenal, T. (ed.), 1978 Methods for assessment of fish production in fresh waters. IBP Handb., (3): 365 p. 3rd ed.
Baisre, J.A., 1985 Los complejos ecológicos de pesca: definición e importancia en la administración de las pesquerías cubanas. FAO Fish.Rep./FAO Rapp.Pêches/FAO Inf.Pesca, (278) Suppl.:251–72
Bajkov, A.D., 1935 How to estimate the daily food consumption of fish under natural conditions. Trans. Am.Fish.Soc., 45:288–9
Bakun, A. y R.H. Parrish, 1980 Environmental inputs to fishery population models for eastern boundary current regions. IOC Workshop Rep., (28):67–104
Bakun, A., et al., 1980 Ocean sciences in relation to living resources. Can.J.Fish.Aquat.Sci., 39(7): 1059–70
Baronov, F.I., 1918 On the question of the biological basis of fisheries. Izv.Nauchno.-Issled.Ikhtiol. Inst., 1:81–128 (en ruso)
Barrett, G.W., G.M. Van Dyne y E.P. Odum, 1976 Stress ecology. BioScience, 26(3):192–4
Bas, G., R. Margalef y P. Rubies (eds), 1985 Simposio internacional sobre las áreas de afloramiento más importantes del oeste africano (cabo Blanco y Benguela). Barcelona, Instituto de Investigaciones Pesqueras, 2 vols:1114 p.
Beverton, R.J.H., 1966 Manual of methods for fish stock assessment. Part 2. Tables of yield functions for fishery assessment. Manuel sur les méthodes d'évaluation des stocks ichtyologiques. Partie 2. Tables des fonctions de rendement. Manual de métodos para la evaluación de los stocks de peces. Parte 2. Tablas de functiones de rendimiento. FAO Fish.Tech.Pap./FAO Doc.Tech.Pêches/FAO Doc.Téc.Pesca, (38):Rev.1:67 p.
Beverton, R.J.H. y S.J. Holt, 1957 On the dynamics of exploited fish populations. Fish.Invest. Minist.Agric.Fish.Food G.B. (2 Sea Fish.), 19:533 p.
Beverton, R.J.H. y S.J. Holt, 1959 A review of the life spans and mortality rate of fish in nature, and their relation to growth and other physiological characteristics. In CIBA Foundation Colloquium on ageing, editado por G.E.W. Wolstenholme and M. O'Connor. Londres, Churchill, Vol.5:142–81
Beyer, J.E., 1981 Aquatic ecosystems: an operational research approach. Seattle, University of Washington Press, 315 p.
Beyer, J.E. y G.C. Laurence, 1981. Aspects of stochasticity in modelling growth and mortality of clupeoid fish larvae. Rapp.P.-V.Réun CIEM, 178:17–23
Blackburn, M., 1968 Micronekton of the eastern tropical Pacific Ocean: family composition, distribution, abundance, and relations to tuna. Fish.Bull.U.S.Fish.Wildl.Serv., 67(1):71–115
Blauenweiss, L., et al., 1978 Relationship between body size and some life history parameters. Oecologia, 37:257–72
Borgmann, U., 1983 Effect of somatic growth and reproduction on biomass transfer up pelagic food webs, as calculated from particle-size-conversion efficiency. Can.J.Fish.Aquat.Sci., 40(11): 2010–8
Bradbury, R.H., R.E. Reichelt y D.G. Green, 1984 Fractals in ecology: methods and interpretation. Mar.Ecol.(Prog.Ser.), 14(2–3):295–6
Breen, P.A. y K.H. Mann, 1976 Changing lobster abundance and the destruction of kelp beds by sea urchins. Mar.Biol., 34:137–42
Brett, J.R., 1979 Environmental factors y growth. In Fish physiology. Vol.8. Bioenergetics and growth, editado por W.S. Hoar, D.J. Randall and J.R. Brett. Nueva York, Academic Press, p. 599–675
Brett, J.R. y T.D.D. Groves, 1979 Physiological energetics. In Fish physiology. Vol.8. Bio energetics and growth, editado por W.S. Hoar, D.J. Randall and J.R. Brett. Nueva York, Academic Press, p. 280–352
Butler, M.J.A., et al., 1986 Marine resource mapping: an introductory manual. FAO Fish.Tech.Pap., (274):256 p.
Caddy, J.F., 1975 Spatial model for an exploited shellfish population and its application to the Georges Bank scallop fishery. J.Fish.Res.Board Can., 32(8):1305–28
Caddy, J.F., 1980 Surplus production models. In Selected lectures from the CIDA/FAO/CECAF Seminar on fishery resource evaluation. Casablanca, Morocco, 6–24 March 1978. Rome, FAO, Canada Funds-in-Trust, FAO/TF/INT 180(c) (CAN) Suppl., pp. 29–55. Issued also in French
Caddy, J.F., 1983 The cephalopods: factors relevant to their population dynamics and to the assessment and management of stocks. FAO Fish.Tech.Pap., (231):419–52
Caddy, J.F., 1983a Algunas consideraciones a propósito de la definición de stocks compartidos y su distribución entre zonas económicas adyacentes. FAO Circ.Pesca, (749):52 p. Publicado también en inglés
Caddy, J.F.,1984 An alternative to equilibrium theory for management of fisheries. FAO Fish.Rep., (289)Suppl. 2:173–214
Caddy, J.F., 1985 Species interactions and stock assessment - some ideas and approaches. In Simposio Internacional sobre las áreas de afloramiento más importantes del Oeste Africano (Cabo Blanco y Benguela), editado por C. Bas, R. Margalef y S.P. Rubies. Barcelona, Instituto de Investigaciones Pesqueras
Caddy, J.F., 1986 Modelling stock-recruitment processes in Crustacea: some practical and theoretical perspectives. Can.J.Fish.Aquat.Sci., 43(11):2330–44
Caddy, J.F. y G.P. Bazigos, 1985 Practical guidelines for statistical monitoring of fisheries in manpower-limited situations. FAO Fish.Tech.Pap., (257):86 p.
Caddy, J.F. y J. Csirke, 1983 Approximations to sustainable yield for exploited and unexploited stocks. Oceanogr.Trop., 18(1):3–15
Caddy J.F. y S. Garcia, 1983 Production modelling without long data series. FAO Fish.Rep./FAO Rapp.Pêches/FAO Inf.Pesca, (268) Suppl.:309–13
Caddy J.F. y S. Garcia, Fisheries thematic mapping: a prerequisite for intelligent management and de velopment of fisheries. (en preparación)
Caddy, J.F. y J.A. Gulland, 1983 Historical patterns of fish stocks. Mar.Policy, 7(4):267–78
Caddy, J.F. y T.D. Iles, 1973 Underwater observations on herring spawning grounds on Georges Bank. ICNAF Res.Bull., (10):131–9
Caddy, J.F. y G.D. Sharp, 1983 Some useful ecological concepts in fisheries assessment. Documento presentado a ACMRR Working Party on the management of living resources in near-shore tropical waters. Roma, 28 febrero - 4 marzo, 1983. Roma, FAO, Working paper (8) (mimeo)
Carey, F.G. y R.J. Olson, 1982 Sonic tracking experiments with tunas. Collect.Vol.Sci.Pap.ICCAT/ Rech.Doc.Sci.CICTA/Colecc.Doc.Sci.CICAA, 17:458–66
Carlenton, R.G., 1985 Man and the sea: the ecological challenge. Am.Zool., 25:451–68
Chapman, A.R.O., 1981 Stability of sea urchin dominated barren grounds following destructive grazing of kelp in St Margarets Bay, eastern Canada. Mar.Biol., 61:307–11
Christy, F.T., Jr., 1983 Derechos de uso territorial en las pesquerías marítimas: definiciones y condiciones. FAO Doc.Téc.Pesca (227):11 p. Publicado también en inglés y francés
Christensen, B., 1978 Primary production of mangrove forests. In Proceedings of International Workshop for Mangrove and estuarine area development for the Indo-Pacific region. 14–19 November 1977, Manila. Los Baños, Laguna, Philippine Council for Agriculture and Resources Research, pp. 131–6
Coelho, M.L. y A.A. Rosenberg, 1984 Causal analysis of some biological data for Illex illecebrosus from the Scotian Shelf. NAFO Sci.Counc.Stud., (7):61–6
Condrey, R.E., 1984 Density-dependent searching time: implications in surplus production models. Fish Bull.NOAA/NMFS, 82(3):537–42
Conover, R.J., 1978 Transformation of organic matter. In Marine ecology. Vol.4. Dynamics, editado por O. Kinne. Chichester, U.K., John Wiley and Sons, pp. 221–456
Conrad, M., 1972 Statistical and hierarchical aspects of biological organization. In Towards a theoretical biology, editado por C.H. Waddington. Edinburgo, Edinburgh University Press, Vol.4:189–221
Conrad, M., 1976 Biological adaptability: the statistical state model. BioScience, 26(5):319–24
Coulter, G.W., 1970 Population changes within a group of fish species in Lake Tanganyika following their exploitation. J.Fish Biol., 2:329–53
Cousins, S., 1985 Ecologists build pyramids again. New Sci., 106(1463):50–4
Csirke, J., 1980 Recruitment in the Peruvian anchovy (Engraulis ringens) and its dependence on the adult population. Rapp.P. -V.Réun.CIEM, 177:307–13
Cushing, D.H., 1982 Climate and fisheries. Londres, Academic Press, 373 p.
Cushing, D.H. y R.R. Dickson, 1976 The biological response in the sea to climatic changes. Adv.Mar. Biol., 14:1–122
Davies, R.G., 1971 Computer programming in quantitative biology. Nueva York, Academic Press, 492 p.
Day, D.S. y W.G. Pearcy, 1968 Species associations of benthic fishes on the continental shelf and slope of Oregon. J.Fish.Res.Board Can., 25(12):2665–75
De Sylva, D.P., 1975 Nektonic food webs in estuaries. In Estuarine research, editado por L.E. Cronin. Nueva York, Academic press, pp. 420–47
Devries, T.J. y W.G. Pearcy, 1982 Fish debris in sediments of the upwelling zone off central Peru: a late Quaternary record. Deep-Sea Res. (A Oceanogr.Res.Pap.), 28(1):87–109
Diana, J.S., 1979 The feeding pattern and daily ration of a top carnivore, the northern pike (Esox lucius). Can.J.Zool., 57(11):2121–7
Dickie, L.M., 1972 Food chains and fish production. ICNAF Spec.Publ., (8):201–21
Domain, F., 1972 Poissons démersaux du plateau continental Sénégambien: application de l'analyse aux composantes principales à une série de chalutages. Cah.ORSTOM (Sér.Océanogr.), 1(2): 111–23
Dotson, R.C., 1978 Fat deposition and utilization in albacore. In The physiological ecology of tunas, editado por G.D. Sharp y A.E. Dizon. Nueva York, Academic Press, pp. 343–55
Doubleday, W.G., 1976 Environmental fluctuations and fisheries management. Sel.Pap.ICNAF, (1)1:141–50
Eggers, D.M., 1977 Factors in interpreting data obtained by diel sampling of fish stomachs. J.Fish. Res.Board Can., 34(2):290–4
Elliott, J.M., 1971 Some methods for the statistical analysis of samples of benthic invertebrates. Sci.Publ.Freshwat.Biol.Assoc.Ambleside, (25):144 p.
Elliott, J.M. y L. Persson, 1978 The estimation of daily rates of food consumption for fish. J.Anim. Ecol., 47:977–91
FAO, 1984 Yearbook of fishery statistics. Annuaire statistique des pêches. Anuario estadístico de pesca, 1983. Catches and landings. Captures et quantités débarquées. Capturas y desembarques. Yearb.Fish.Stat./Annu.Stat.Pêches/Anu.Estad.Pesca, (56):72
Fänge, R. y D. Grove, 1979 Digestion. In Fish physiology. Vol.8. Bioenergetics and growth, editado por W.S. Hoar, D.J. Randall y J.R. Brett. Nueva York, Academic Press, pp. 161–260
Fasham, M.J.R., 1978 The statistical and mathematical analysis of plankton patchiness. Oceanogr.Mar. Biol., 16:43–79
Fasham, M.J.R., (ed.), 1984 Flows of energy and materials in marine ecosystems: theory and practice. Nueva York, Plenum Press, NATO Conference series, IV, Marine sciences, 13:733 p.
Fisher, R.A., 1960 The design of experiments. Edinburgo, Oliver and Boyd, 7th ed.
Fortunatova, K.R. y O.A. Popova, 1973 Feeding and food relationships of predatory fish in Volga delta. Moscow, Nauka Press
Fry, F.E.J., 1971 The effects of environmental factors on the physiology of fish. In Fish physiology, editado por W.S. Hoar y D.J. Randall. Nueva York, Academic Press, Vol.6:1–98
Fuller, K.H., 1979 Shoreline as a controlling factors in commercial shrimp production. NASA Tech.Memo., (72–73):227 p.
Gabriel, W.L. y A.V. Tyler, 1980 Preliminary analysis of Pacific coast demersal fish assemblages. Mar.Fish.Rev., 42(3–4):83–8
Gallucci, V.F., 1973 On the principles of thermodynamics in ecology. Annu.Rev.Ecol.Syst., 4:329–57
Garcia, S., 1982 Distribution, migration and spawning of the main fish resources in the northern CECAF area. CECAF/ECAF Ser., (82/25):9 p.
Garcia, S., 1983 The stock-recruitment relationship in shrimps: reality or artefacts and misinter pretations? Oceanogr.Trop., 18(1):25–48
Garcia, S. y L. Le Reste, 1986 Ciclos vitales, dinámica, explotación y ordenación de las poblaciones de camarones peneidos costeros. FAO Doc.Téc.Pesca, (203):180 p. Publicado también en inglés y francés
Gilliland, R.L., 1982 Solar, volcanic and CO2 forcing of recent climatic changes. Climat.Change, 4:111–31
Goeden, G.B., 1982 Intensive fishing and a “keystone” predator species: ingredients for community instability. Biol.Conserv., 22:273–81
Golley, F.B., 1972 Energy flux in ecosystems. In Ecosystem structure and function, editado por J.A. Wiens. Proceedings of the Thirty-First Annual Biology Colioquium. Corvallis, Oregon State University Press, pp. 69–89
Grady, J.R., 1971 The distribution of sediment properties and shrimp catch on two shrimping grounds on the continental shelf of Mexico. Proc.Gulf Caribb.Fish.Inst., 23:39–48
Graham, M., (ed.), 1956 Sea fisheries: their investigation in the United Kingdom. Londres, Edward Arnold Publishers, 487 p.
Grainger, R.J.R., 1980 Irish west coast herring fluctuations and their relation to oceanographic con ditions. Rapp.P.-V.Réun.CIEM, 177:444 (Abstr.)
Greig-Smith, P., 1964 Quantitative plant ecology. Londres, Butterworths, 256 p. 2nd ed.
Gulland, J.A., 1970 Food chain studies and some problems in world fisheries. In Marine food chains, editado por J.H. Steele. Edinburgo, Oliver and Boyd, pp. 296–315
Gulland, J.A., (comp.), 1971 The fish resources of the ocean. West Byfleet, Surrey, Fishing News (Books) Ltd., por FAO, 225 p. Ed.rev. de FAO Fish.Tech.Pap., (97):425 p.(1970)
Gulland, J.A. y L.K. Boerema, 1973 Scientific advice on catch levels. Fish.Bull.NOAA/NMFS, 71(2): 32–5
Gulland, J.A. y S. Garcia, 1984 Observed patterns in multispecies fisheries. In Exploitation of marine communities edited by R.M. May. Report of the Dahlem Workshop on exploitation of marine communities. Berlin, 1–6 April 1984. Berlin, Springer-Verlag, Life sciences research report, 32:155–90
Gunderson, D.R., 1980 Using r-K selection theory to predict natural mortality. Can.J.Fish.Aquat.Sci., 37(12):2266–71
Hackney, P.A. y C.K. Minns, 1974 A computer model of biomass dynamics and food competition with implications for its use in fishery management. Trans.Am.Fish.Soc., 103(2):215–25
Hamblyn, E.L., 1966 The food and feeding habits of Nile perch Lates niloticus (Linne) (Pisces: Centroponidae). Rev.Zool.Bot.Afr., 74(1–2):28 p.
Hancock, D.A. y A.E. Urquhart, 1965 The determination of natural mortality and its causes in an exploited population of cockles (Cardium edule L.). Fish.Invest.Minist.Agric.Fish.Food G.B.(2 Sea Fish.), 24(2):40 p.
Hardy, A.C., 1924 On food and feeding habits of the herring. Fish.Invest.Minist.Agric.Fish.Food G.B. (2 Sea Fish.), 7(3)
Henderson, H.F., R.A. Ryder y A.W. Kudhongania, 1973 Assessing fishery potentials of lakes and reservoirs. J.Fish.Res.Board Can., 30(12) Part 2:2000–9
Hennemuth, R.C. y S.M. Autges, 1982 Effects of variability of recruitment on management service. ICES Pelagic Fish.Comm., CM 1982/H:22:4 p. (mimeo)
Hennemuth, R.C., J.E. Palmer y B.E. Brown, 1980 A statistical description of recruitment in eighteen selected fish stocks. J.Northwest Atl.Fish.Sci., 1:101–11
Heron, A.C., 1972 Population ecology of a colonizing species, the pelagic tunicate Thalia democratica. Part 1. Individual growth rate and generation time. Oecologia, 10(4):269:93
Heron, A.C., 1973 Population ecology of a colonizing species, the pelagic tunicate Thalia democratica. Part 2. Population growth rate. Oecologia, 10(4):294–312
Hewitt, R.P., 1981 The value of pattern in the distribution of young fish. Rapp.P.-V.Réun.CIEM, 178: 229–36
Hewitt, R.P., 1982 Spatial pattern and survival of anchovy larvae: implications of adult reproductive strategy. Ph.D. Dissertation. University of California at San Diego, 187 p.
Hoar, W.S. y D.J. Randall (eds), 1978 Fish physiology. Vol.7. Locomotion. Nueva York, Academic Press, 576 p.
Hoar, W.S., D.J. Randall y J.R. Brett (eds), 1979 Fish physiology. Vol.8. Bio-energetics and growth. Nueva York, Academic Press, 786 p.
Hobson, E.S., 1968 Predatory behavior of some shore fishes in the Gulf of California. Res.Rep.U.S.Fish Wildl.Serv., (73):92 p.
Hobson, E.S., 1973 Diel feeding migrations in tropical reef fishes. Helgol.Wiss.Meeresunters., 24: 361–70
Hobson, E.S., 1974 Feeding relationships of teleostean fishes on coral reefs in Kona, Hawaii. Fish. Bull.NOAA/NMFS, 72(4):915–1031
Hobson, E.S., 1975 Feeding patterns among tropical reef fishes. Am.Sci., 63:382–92
Hobson, E.S. y J.R. Chess, 1976 Trophic interactions among fishes and zooplankters near shore at Santa Catalina Island, California. Fish.Bull.NOAA/NMFS, 74(3):567–98
Hobson, E.S. y J.R. Chess, 1978 Trophic relationships among fishes and plankton in the lagoon at Enewetak Atoll, Marshall Islands. Fish.Bull.NOAA/NMFS, 76(1):133–53
Holden, M.J., 1978 Long-term changes in landing of fish from the North Sea. Rapp.P.-V.Réun.CIEM, 172: 11–26
Holling, C.S., 1973 Resilience and stability of ecological systems. Annu.Rev.Ecol.Syst., 4:1–23
Horne, E.P.W. y T. Platt, 1984 The dominant space and time scales of variablity in the physical and biological fields on continental shelves. Rapp.P.-V.Réun.CIEM, 138:8–19
Hughes, R.N. y M.L.H. Thomas, 1971 The classification and ordination of shallow-water benthic samples from Prince Edward Island. Can.J.Exp.Mar.Biol.Ecol., 7:1–39
Hunter, J.R. y R. Leong, 1981 The spawning energetics of female northern anchovy, Engraulis mordax. Fish.Bull.NOAA/NMFS, 79(2):215–30
Hurley, A.C., 1976 Feeding behaviour, food consumption, growth and respiration of the squid Loligo opalescens raised in the laboratory, Fish.Bull.NOAA/NMFS, 74(1):176–82
Huston, M., 1979 A general hypothesis of species diversity. Am.Nat., 13(1):81–101
Hutchinson, B., 1965 The ecological theatre and the evolutionary play. New Haven, Yale University Press, 139 p.
Hutchinson, C.E., 1957 Concluding remarks. Cold Spring Harbour. Symp.Quant.Biol., 22:415–27
Iles, T.D. y M. Sinclair, 1982 Atlantic herring: stock discreteness and abundance. Science, Wash., 215(4533):627–33
Isaacs, J.D., 1972 Unstructured marine food webs and “pollutant analogues”. Fish.Bull.NOAA/NMFS, 70(3):1053–9
Isaacs, J.D., 1973 Potential trophic biomasses and trace substances concentrations in unstructured marine food webs. Mar.Biol., 22:97–104
Isaacs, J.D., 1976 Reproductive products in marine food webs. Bull.South.Calif.Acad.Sci., (75):220–3
Ivlev, V., 1961 Experimental ecology of the feeding of fishes. Trad. del ruso. New Haven, Connecticut, Yale University Press, 302 p.
Johannes, R.E., 1972 The metabolism of some coral reef communities a team study of nutrient and energy flux at Eniwetok. BioScience, 22(9):541–3
Johnson, L., 1981 The thermodynamic origin of ecosystems. Can.J.Fish.Aquat.Sci., 38(5):571–90
Jones, R., 1978 Estimates of the food consumption of haddock (Melanogrammus aeglefinus) and cod (Gadus morhua). J.Cons.CIEM, 38:18–27
Jones, R., 1982 Species interactions in the North Sea. Can.Spec.Publ.Fish.Aquat.Sci., (59):48–63
Jones, R., 1982a El uso de datos de composición por tallas en la evaluación de poblaciones de peces (con notas sobre VPA y análisis de cohortes). FAO Circ.Pesca, (734):61 p. Publicado también en inglés y francés
Jones, R., 1982a Ecosystems, food chains and fish yields. ICLARM Conf.Proc., (9):195–239
Jones, R., 1984 Assessing the effects of changes in exploitation pattern using length composition data (with notes on VPA and cohort analysis). FAO Fish.Tech.Pap., (256):118 p.
Jones, R. y E.W. Henderson, 1980 Further observations on energy flow through the marine food chain. ICES Biological Oceanography Cmtee. CM 1980/L:26 (mimeo)
Kapetsky, J.M., 1981 Some considerations for the management of coastal lagoon and estuarine fisheries. FAO Fish.Tech.Pap., (218):47 p. Issued also in French and Spanish
Kapetsky, J.M., 1982 Some potential environmental effects of coastal aquaculture with implications for site selection and aquaculture engineering. Manila, South China Sea Fisheries Development and Coordinating Programme, SCS/82/GEN/42:76–82
Kapetsky, J.M., 1985 Mangroves, fisheries and aquaculture. FAO Fish.Rep./FAO Rapp.Pêches/FAO Inf. Pesca, (338):Suppl.:17–36
Kawasaki, T., 1980 Fundamental relations among the selections of life history in the marine Teleost. Bull.Jap.Soc.Sci.Fish., 46(3):289–93
Kenchington, R.A. y B.E.T. Hudson (eds), 1984 Coral reef management handbook. Jakarta, Indonesia, Unesco Regional Office for Science and Technology for Southeast Asia, 281 p.
Kerr, S.R., 1974 Structural analysis of aquatic communities. In Proceedings of the First International Congress on ecology, The Hague. Wageningen, Netherlands Centre for Agricultural Publishing and Documentation
Kerr, S.R., 1977 Structure and transformation of fish production systems. J.Fish.Res.Board Can., 34(10):1989–93
Kerr, S.R., 1980 Niche theory in fisheries ecology. Trans.Am.Fish.Soc., 109:254–60
Kerr, S.R., 1982 Estimating the energy budgets of actively predatory fish. Can.J.Fish.Aquat.Sci., 39(3):371–9
Kerr, S.R. y R.A. Ryder, 1977 Niche theory and percid community structure. J.Fish.Res.Board Can., 34(10):1952–58
Kinne, O., (ed.), 1970–84 Marine ecology. A comprehensive, integrated treatise on life in oceans and coastal waters. Chichester, U.K., John Wiley and Sons, Vol.1, Pts 1–3; Vol.2. Pts 1–2; Vol.4; Vol.5; Pts 1–4
Knight, W. y A.V. Tyler, 1973 A method for compression of species association data by using habitat preferences, including an analysis of fish assemblages on the southwest Scotian Shelf. Tech.Rep.Fish.Res.Board.Can., (402):15 p.
Kondo, K., 1980 The recovery of the Japanese sardine - the biological basis of stock-size fluctuations. Rapp.P.-V.Réun.CIEM, 177:332–54
Laevastu, T. y F. Favorite, 1980 Holistic simulation models of shelf-seas ecosystems. In Analysis of marine ecosystems, editado por A.R. Longhurst. Londres, Academic Press, pp. 701–27
Laevastu, T. y H.A. Larkins, 1981 Marine fisheries ecosystem: its quantitative evaluation and management. Farnham, Surrey, Fishing News Books Ltd., 162 p.
Lamson, C. y A.J. Hanson (eds), 1984 Atlantic fisheries and coastal communities: fisheries decisionmaking case studies. Halifax, Nueva Escocia, Dalhousie University, Institute for Resource and Environmental Studies, 262 p.
Lane, E.D., 1975 Quantitative aspects of the life history of the diamond turbot, Hypsopsetta gluttulata (Girard), in Anaheim Bay. Fish.Bull.Calif.Dep.Fish Game, (165):153–73
Lane, E.D., M.C.S. Kingsley y D.E. Thornton, 1979 Daily feeding and food conversion efficiency of the diamond turbot: an analysis based on field data. Trans.Am.Fish.Soc., 108:530–5
Larkin, P.A., 1963 Interspecific competition and exploitation. J.Fish.Res.Board Can., 20(3):647–78
Larkin, P.A. y S.W. Gazey, 1982 Applications of ecological simulation models to management of tropical multispecies fisheries. ICLARM Conf.Proc., (9):123–40
Lasker, R., 1966 Feeding, growth, respiration and carbon utilization of a euphausid crustacean. J.Fish.Res.Board Can., 23(9):1291–317
Lasker, R., 1978 Ocean variability and its biological effects-regional review northeast Pacific. Rapp.P.-V.Réun.CIEM, 173:168–81
Lessios, H.A., P.W. Glynn y J.D. Cubit, 1984 Spread of Diadema mass mortality through the Caribbean. Science, Wash., 226:335–7
Li, H.W. y P.B. Moyle, 1981 Ecological analysis of species introductions into aquatic systems. Trans.Am.Fish.Soc., 110:772–82
Lie, U. y J.C. Kelley, 1970 Benthic infauna communities off the coast of Washington and in Puget Sound: identification and distribution of the communities. J.Fish.Res.Board Can., 27(1):621–51
Lindemann, R.L., 1942 The trophic-dynamic aspect of ecology. Ecology, 23:399–418
Livingston, R.J., 1982 Trophic organization of fishes in a coastal seagrass system. Mar.Ecol.(Prog. Ser.), 7:1–12
Lleonart, J., J. Salet y E. MacPherson, 1983 Effect of the cannibalism in hake population. ICES Demersal Fish Committee CM 1983/G:54:22 p. (mimeo)
Lleonart, J., J. Salet y E. MacPherson, 1983a Efecto del canibalismo en la población de Merluccius capensis en la División 1.5. Collect.Sci.Pap.ICSEAF/Recl.Doc.Sci.CIPASE/Colecc.Doc.Cient.CIPASO, 10(1):111–28
Longhurst, A.R., 1981 Analysis of marine ecosystems. Nueva York, Academic Press, 741 p.
Louins, A.B. y L.H. Louins, 1981 Energy: what is the problem? Ecologist, 11(6):302–13
MacArthur, R., 1955 Fluctuations of animal populations and a measure of community stability. Ecology, 36(3):533–6
MacArthur, R.H. y E.O. Wilson, 1967 The theory of island biogeography. Princeton, N.J., Princeton University Press, 25 p.
MacDonald, J.S., et al., 1984 Fishes, fish assemblages and their seasonal movements in the lower Bay of Fundy and Passamaquoddy Bay, Canada. Fish.Bull.NOAA/NMFS, 82(1):121–39
MacDonald, N., 1983 Trees and networks in biological models. Nueva York, John Wiley, 216 p.
Mackinnon, J.C., 1973 Analysis of energy flow and production in an unexploited marine flatfish popula tion. J.Fish.Res.Board Can., 30(11):717–28
Mackinnon, J.C., 1973a Metabolism and its relationship with growth rate of American plaice, Hippoglossoides platessoides Fabr. J.Exp.Mar.Biol.Ecol., 11:297–310
MacPherson, E., 1981 Resource partitioning in a Mediterranean demersal fish community. Mar.Ecol.(Prog. Ser.), 4(2):183–94
Magnuson, J.J., 1969 Digestion y food consumption by skipjack tuna (Katsuwonis pelamis). Trans.Am. Fish.Soc., 98:379–92
Magnuson, J.J. y J.G. Heitz, 1971 Gill raker apparatus and food selectivity among mackerels, tunas and dolphins. Fish.Bull.NOAA/NMFS, 69(2):361–70
Mahon, R., H. Oxenford y W. Hunte (eds), 1986 Development strategies for flying fish fisheries of the eastern Caribbean. Proceedings of an IDRC-sponsored Workshop at the University of the West Indies, Cave Hill, Barbados, 22–23 October 1985. Manuscr.Rep.Int.Dev.Res.Cent., Ottawa, (IDRC-MRI 28e):147 p.
Mahon, R., et al., 1984 Spatial y temporal patterns of groundfish distribution on the Scotian Shelf and in the Bay of Fundy, 1970–1981. Can.Tech.Rep.Fish.Aquat.Sci., (1300):164 p.
Mandelbrot, B.B., 1982 The fractal geometry of nature. Nueva York, W.H. Freeman, 460 p.
Mann, K.H., 1972 Ecological energetics of the seaweed zone in a marine bay on the Atlantic coast of Canada. 2. Productivity of the seaweeds. Mar.Biol., 14:199–209
Mann, K.H., 1977 Destruction of kelp-beds by sea urchins: a cyclic phenomenon or irreversible degradation? Helgol.Wiss.Meeresunters., 30:455–67
Mann, K.H., 1978 Estimating the food consumption of fish in nature. In Ecology of freshwater fish production, editado por S.D. Gerking. Edinburgo, Oxford Blackwell Scientific Publications, pp. 250–73
Mann, K.H., 1982 Ecology of coastal waters: a systems approach. Berkeley, California. University of California Press, 322 p.
Margalef, R., 1968 Perspectives in ecological theory. Chicago, University of Chicago Press, 111 p.
Margalef, R., 1978 General concepts of population dynamics and food links. In Marine ecology. Vol.4. Dynamics, editado por O. Kinne. Chichester, R.U., Wiley-Interscience, pp. 617–704
Mark, D.M., 1984 Fractal dimension of a coral reef at ecological scales: a discussion. Mar.Ecol. (Prog.Ser.), 14:293–4
Marten, G.C. y J.J. Polovina, 1982 A comparative study of fish yields from various tropical ecosystems. ICLARM Conf.Proc., 9:255–85
Martosubroto, P. y N. Naamin, 1977 Relationship between tidal forests (mangroves) and commercial shrimp production in Indonesia. Mar.Res.Indones., 18:81–6
May, R.M., 1974 Stability y complexity in model ecosystems. Princeton, N.J., Princeton University Press. Princeton N.J.
May, R.M., 1984 (ed.) Exploitation of marine communities. Report of the Dahlem Workshop on exploitation of marine communities. Berlin 1984, April 1–6. Berlin, Springer-Verlag, Dahlem Workshop reports, Life sciences research report, 32:366 p.
May, R.M., et al., 1979 Management of multispecies fisheries. Science, Wash., 205(4403):267–77
McNeill, S. y J.H. Lawton, 1970 Annual production and respiration in animal populations. Nature, Lond., 225(5231):472–4
Mearns, A.J., et al., 1981 Trophic structure and the caesium-potassium ratio in pelagic ecosystems. Rep.CCOFI, (22):99–110
Miller, R.B., 1985 A succession in sea urchin and seaweed abundance in Nova Scotia, Canada. Mar.Biol., 84:275–86
Miller, R.B., 1985a Sea urchin pathogen: a possible tool for biological control. Mar.Ecol.(Prog.Ser.), 21:169–74
Miller, R.J., K.H. Mann y D.J. Scarratt, 1971 Production potential of a seaweed lobster community in eastern Canada. J.Fish.Res.Board Can., 28(11):1733–8
Mills, E.L., K. Pittman y F.C. Tan, 1982 Food web structure on the Scotian Shelf, eastern Canada: a study using 13C as a food-chain tracer. Documento presentado a Symposium on Biological productivity of continental shelves in the temperate zone of the north atlantic. Kiel, F.R.G., 2–5 March 1982, Doc. No. 29 (mimeo)
Morse, D.R., et al., 1985 Fractal dimension of vegetation and the distribution of arthropod body lengths. Nature, Lond., 314:731–3
Munro, J.L., 1967 The food of a community of East African fish. J.Zool.Lond., 151:389–415
Munro, J.L., 1977 Actual and potential fish production from the coralline shelves of the Caribbean Sea. FAO Fish.Rep., (200):301–21
Munro, J.L., 1979 Stock assessment models: applicability and utility in tropical small-scale fisheries. In Stock assessments for tropical small-scale fisheries. Proceedings of the International Workshop, University of Rhode Island, 19–21 September 1979, editado International Center for marine Resource Development, pp. 35–47
Munro, J.L., 1982 Estimation of biological and fishery parameters in coral reef fisheries. ICLARM Conf.Proc., (9):71–82
Munro, J.L., (ed.), 1983 Caribbean coral reef fishery resources. ICLARM Stud.Rev., (7):276 p.
Munro, J.L., 1984 Yields from coral reef fisheries. ICLARM Fishbyte, 2(3):13–5
Murdoch, W.W., 1969 Switching in general predators: experiments on predator specificity and stability of prey populations. Ecol.Monogr., 39(4):335–54
Newell, R.C., 1984 The biological role of detritus in the marine environment. In Flows of energy and materials in marine ecosystems: theory and practice editado por M.J.R. Fasham. Nueva York, Plenum Press, NATO Conference Series, IV, Marine sciences, Vol. 13, pp. 317–43
Nicolis, G. y I. Prigogine, 1977 Self-organization in non-equilibrium: to dissipative structures to order through fluctuations. New York, Wiley-Interscience, 491 p.
Nixon, S.W., 1982 Nutrient dynamics, primary production and fisheries yields of lagoons. Oceanol.Acta, 5(4) Suppl. pp. 357–71
Norman, J.R., 1963 A history of fishes. Londres, Ernest Benn Ltd., 398 p. 2nd ed. Editado por P.H. Greenwood
Odum, E.P., 1968 Energy flow in ecosystems: a historical review. Am.Zool., 8:217–24
Odum, E.P., 1969 The strategy of ecosystems development. Science,Wash., 164:262–70
Odum, H.T., 1967 Biological circuits and the marine systems of Texas. In Pollution and marine ecology, editado por T.A. Olson y F.J. Burgess. Nueva York, Wiley Interscience, pp. 99–157
Odum, H.T., 1972 An energy circuit language for ecological and social systems: its physical basis. In System analysis and simulations in ecology, editado por B.C. Patten. Nueva York, Academic Press, pp. 140–211
Olson, R.J., 1982 Feeding and energetics studies of yellowfin tuna; food for ecological thought. Collect.Vol.Sci.Pap.ICCAT/Rech.Doc.Sci.CICTA/Colecc.Doc.Cient.CICAA, 17:444–57
Overholtz, W.J. y A.V. Tyler, 1985 Long-term responses of the demersal fish assemblages of Georges Bank. Fish.Bull.NOAA/NMFS, 83(4):507–20
Owen, R.W., 1980 Patterning of flow and organisms in the larval anchovy environment. IOC Workshop Rep., (28):1167–200
Paine, R.T., 1969 The Pisaster-Tegula interaction: prey patches, predator food preference and inter tidal community structure. Ecology, 50:930–8
Paine, R.T., 1969a A note on trophic complexity and community stability. Am.Nat., 103:91–3
Paloheimo, J.E., 1961 Studies on estimation of mortalities. 1. Comparison of a method described by Beverton and Holt and a new linear formula. J.Fish.Res.Board Can., 18(5):645–62
Paloheimo, J.E. y L.M. Dickie, 1970 Production and food supply. In Marine food chains, editado por J.H. Steele. Edinburgo, Oliver and Boyd, pp. 499–527
Paloheimo, J.E. y H.A. Regier, 1982 Ecological approaches to stressed multispecies fisheries resources. Can.Spec.Publ.Fish.Aquat.Sci., (59):127–32
Parrish, J.D. y R.J. Zimmerman, 1977 Utilization by fishes of space and food resources on an offshore Puerto Rican coral reef and its surrounding. In Proceedings of the third International Coral Reef Symposium. Miami, Florida, University of Miami, Rosentiel School of Marine and Atmospheric Science, Vol. 1:297–303
Passet, R., 1979 L'économique et le vivant. Paris, Payot, Petite Bibliothèque, 286 p.
Parrish, R.H. y A.D. MacCall, 1978 Climatic variation and exploitation in the Pacific mackerel fishery. Fish Bull.Cal.Dep.Fish Game, (167):110 p.
Parrish, R.H., C.S. Nelson y A. Bakun, 1981 Transport mechanisms and reproductive success of fishes in the California Current. Biol.Oceanogr., 1 (2):175–203
Pauly, D., 1979 Theory and management of tropical multispecies stocks: a review, with emphasis on the Southeast Asian demersal fisheries. ICLARM Stud.Rev., (1):35 p.
Pauly, D., 1979a Gill size and temperature as governing factors in fish growth: a generalization of the von Bertalanffy growth formula. Ber.Inst.Meereskd.Christian-Albrechts Univ.Kiel, (63)
Pauly, D., 1980 A selection of simple methods for the assessment of tropical fish stocks. FAO Fish. Circ., (729):54 p. Publicado también en francés
Pauly, D., 1980a On the interrelationships between natural mortality, growth parameters and mean en vironmental temperature in 175 fish stocks. J.Cons.CIEM, 39(3):195–212
Pauly, D., 1981 Fish stock assessment in coral reefs - notes on the state of the art. ICLARM Newsl., 4(3):19
Pauly, D., 1983 Algunos métodos simples para la evaluación de recursos pesqueros tropicales. FAO Doc.Téc.Pesca, (234):49 p. Publicado también en inglés y francés. Revisión de FAO Fish. Circ., (729):54 p.
Pauly, D., 1984 Fish population dynamics in tropical waters: a manual with application programs for programmable calculators. ICLARM Stud.Rev., (8):325 p.
Pauly, D. y A.N. Mines, (eds), 1982 Small-scale fisheries of San Miguel Bay, Philippines: biology and stock assessment. ICLARM Tech.Rep., (7):124 p.
Pauly, D. y G.I. Murphy (eds), 1982 Theory and management of tropical fisheries. Proceedings of the ICLARM/CSIRO Workshop on the theory and management of tropical multispecies stocks. 12–21 January 1981, Cronulla, Australia. ICLARM Conf.Proc., (9):360 p.
Peterson, I. y J.S. Wroblewski, 1984 Mortality rate of fishes in the pelagic ecosystem. Can.J.Fish. Aquat. Sci., 41(7):1117–20
Pianka, E.R., 1970 On r-selection and K-selection. Am.Nat., 104:592–7
Pianka, E.R., 1972 r and K - selection or b and d selection? Am.Nat., 106:581–8
Pielou, E.C., 1969 An introduction to mathematical ecology. New York, Wiley Interscience, 286 p.
Pimm, S.L., 1980 Properties of food webs. Ecology, 61(2):219–25
Pinkas, L., M.S. Oliphant y I.L.K. Iverson, 1971 Food habits of albacore, bluefin tuna and bonito in California waters. Fish Bull.Calif.Dep.Fish Game, (152):1–105
Pitt, T.K., 1972 Estimates of annual mortality coefficients of American plaice. ICNAF Res.Doc., (72/15) Serial No. (2699):12 p.
Pitt, T.K., 1978 American plaice from ICNAF subarea 2 and divisions 3K, a stock assessment update. CAFSAC Res.Doc./CSPA Doc.Rech., Dartmouth, N.S., (78/11):6 p. (mimeo)
Platt, T., 1971 The annual production by phytoplankton in St. Margaret's Bay, Nueva Escocia. J.Cons. CIEM, 33(3):324–33
Platt, T. y K. Denman, 1975 A general equation for the mesoscale distribution of phytoplankton in the sea. Mem.Soc.R.Sci.Liege(60 Ser.), 7:31–42
Platt, T. y K. Denman, 1977 Organization in the pelagic ecosystem. Helgol.Wiss.Meersunters., 30:575–81
Platt, T. y K. Denman, 1978 The structure of pelagic marine ecosystems. Rapp.P.-V.Réun.CIEM, 173:60–5
Platt, T., K.H. Mann y R.E. Ulanowicz (eds), 1981 Mathematical models in biological oceanography. Monogr.Oceanogr.Methodol., (7):156 p.
Pollard, D.A., 1984 A review of ecological studies on seagrass - fish communities, with particular reference to recent studies in Australia. Aquat.Bot., 18:3–42
Polovina, J.J. y M.D. Ow, 1985 An approach to estimating an ecosystem box model. Fish.Bull.NOAA/NMFS, 83(3):457–60
Pomeroy, L.R., 1974 The oceans' food web, a changing paradigm. Bioscience, 24(9):499–504
Pope, J.G., 1979 A modified cohort analysis in which constant natural mortality is replaced by estimates of predation level. ICES CM. 1981/G:14 (mimeo)
Pope, J.G., 1980 Phalanx analysis: an extension of Jones; length cohort analysis to multispecies cohort analysis. ICES Demersal Fish Committee. CM. 1980/G:19:5 p. (mimeo)
Pope, J.G., 1980a Assessment of multispecies resources. In Selected lectures from the CIDA/FAO/CECAF Seminar on fishery resource evaluation. Casablanca, Morocco, 6–24 March 1978. Roma, FAO, Canada Funds-in-trust, FAO/TF/INT 180(c) (CAN) Suppl. Publicado también en francés
Pope, J.G. y L. Woolner, 1981 A simple investigation into the effects of predation on recruitment to some North Sea fish stocks. ICES CM. 1981/G:14:4 p. (mimeo)
Popova, O.A., 1978 The role of predacious fish in ecosystems. In Ecology of freshwater fish produc tion, editado por S.D. Gerking. Oxford, Blackwell Scientific Publications, pp. 215–49
Powles, H. y C.A. Barans, 1980 Groundfish monitoring in sponge-coral areas off the Southeastern United States. Mar.Fish.Rev., 42(5):21–35
Prigogine, I., 1978 Time structure and fluctuations. Science, Wash., 201(4358):777–85
Pringle, J.D., G.J. Sharp y J.F. Caddy, 1982 Interactions in kelp bed ecosystems in the northwest Atlantic: review of a Workshop. Can.Spec.Publ.Fish.Aquat.Sci., (59):108–15
Ran, G.H., et al., 1983 Animal 13C/12C correlates with trophic level in pelagic food webs. Ecology, 64(5):1314–8
Randall, J.E., 1967 Food habits of reef fishes of the West Indies. Stud.Trop.Oceanogr., 5:665–847
Rapport, D.J., H.A. Regier y T.C. Hutchinson, 1985 Ecosystem behaviour under stress. Am.Nat., 125 (5):617–38
Rasmusson, E.M., 1984 El Niño: the ocean/atmosphere connection. Oceanus, 27(2):5–12
Regier, H.A., 1973 Sequence of exploitation of stocks in multispecies fisheries in the Laurentian Great Lakes. J.Fish.Res.Board Can., 30(12)pt.2:1992–9
Regier, H.A. y H.F. Henderson, 1973 Towards a broad ecological model of fish communities and fisheries. Trans.Am.Fish.Soc., 102(1):56–72
Regier, H.A. y K.H. Loftus, 1972 The ecological role of fisheries exploitation in salmonid communities. J.Fish.Res.Board Can., 29(7):959–68
Reintjes, J.W. y J.E. King., 1953 Food of yellowfin tuna in the Central Pacific. Fish.Bull U.S. Fish Wildl. Serv., 54:90–110
Rheinheimer, G., 1980 Aquatic microbiology. Chichester, U.K., John Wiley and Sons, 235 p. 2nd ed. Trad. del alemán
Ricker, W.E., 1958 Handbook of computations for biological statistics of fish populations. Bull.Fish. Res. Board Can., (119):300 p.
Ricker, W.E., 1973 Linear regression in fishery research. J.Fish.Res.Board Can., 30(3):409–34
Ricker, W.E., 1975 Computations and interpretation of biological statistics of fish populations. Bull. Fish. Res. Board Can., (191):382 p.
Ricker, W.E., 1979 Growth rates and models. In Fish physiology. Vol. 8. Bioenergetics and growth, editado por H.S. Hoar, D.J. Randall y J.R. Brett. Nueva York, Academic Press, pp. 677–743
Rigler, F.H., 1982 The relation between fishery management and limnology. Trans.Am.Fish.Soc., 111(2): 121–32
Rikhter, V.A. y V.N. Efanov, 1976 On one of the approaches to estimates of natural mortality of fish populations. ICNAF Res.Doc., 76(VI) 8:12 p.
Robson, D.S., 1963 Maximum likelihood estimation of a sequence of annual survival rates from a capture/ recapture series. Spec.Publ.ICNAF, (4):330–5
Roger, C. y R. Grandperrin, 1976 Pelagic food webs in the tropical Pacific. Limnol.Oceanogr., 21: 731–5
Rollet, B., 1984 La ecología de los manglares con referencia especial a la base biológica para la ordenación sostenida, forestal y pesca. Documento presentado al Seminario regional de la FAO sobre la ordenación integrada de los manglares. Cuba-Venezuela-Colombia, 5–23 de noviembre de 1984. 33 p. (mimeo)
Romesburg, H.C., 1984 Cluster analysis for researchers. Belmont, California, Lifetime Learning Publications
Rowe, G.T., 1971 Benthic biomass and surface productivity. In Fertility of the sea, editado por J.D. Costlow. Nueva York, Gordon and Breach, vol. 2:441–54
Ryther, J.H., 1969 Photosynthesis and fish production in the sea. Science, Wash., 166:72–6
Saenger, P., E.J. Hegerl y J.D.S. Davie (eds), 1983 Global status of mangrove ecosystems. IUCN Comm. Ecol. Pap., (3):88 p.
Saila, S.B. y T.A. Gaucher, 1966 Estimation of the sampling distribution and numerical abundance of some mollusks in a Rhode Island Salt pond. Proc.Natl.Shellfish.Assoc., 56:73–80
Saila, S.B. y J.D. Parrish, 1972 Exploitation effects upon interspecific relationships in marine ecosystems. Fish.Bull.NOAA/NMFS, 70(2):383–93
Sainsbury, K.J., 1982 The ecological basis of tropical fisheries management. ICLARM Conf.Proc., 9:167–94
Sanders, M.J., 1977 Estimation of mortality coefficients for exploited fish populations in which the sexes exhibit divergent growth rates. J.Cons.CIEM., 37(2):192–3
Santander, H., 1981 Patrones de distribución y fluctuaciones de desoves de anchoveta y sardina. In Volumen extraordinario: Investigación cooperativa de la anchoveta y su ecosistema - ICANE-entre Perú y Canadá. Callao, Instituto del Mar del Perú
Saunders, P.T., 1980 An introduction to catastrophe theory. Cambridge, R.U., Cambridge University Press, 144 p.
Schaefer, M.B., G.C. Broadhead y C.J. Orange, 1963 Synopsis on the biology of yellowfin tuna, Thunnus (Neothunnus) albacares (Bonnaterre) 1788 (Pacific Ocean). FAO Fish.Rep., (6) vol. 2: 538–61
Schlesinger, D.A. y H.A. Regier, 1982 Climatic and morphoedaphic indices of fish yields from natural lakes. Trans.Am.Fish.Soc., 111:141–50
Seaburg, K.G. y J.B. Moyle, 1964 Feeding habits, digestion rates, and growth of some mennesota warmwater fishes. Trans.Am.Fish.Soc., 93(3):269–85
Selye, H., 1974 Stress without distress. Filadelfia, Pennsilvania, J.B. Lippincott Co., 171 p.
Sharp, G.D., 1978 Behavioral and physiological properties of tuna and their effects on vulnerability to fishing gear. In The physiological ecology of tunas, editado por G.D. Sharp y A.E. Dizon. Nueva York, Academic Press, pp. 397–450
Sharp, G.D., 1980 (Rapporteur), Report of the Workshop on the effects of environmental variation on survival of larval pelagic fishes. IOC Workshop Rep., (28):1–47
Sharp, G.D., 1980a Colonization: modes of opportunism in the ocean. IOC Workshop Rep., (28):125–48
Sharp, G.D., (ed.), 1980b Workshop on the effects of environmental variation on the arrival of larval pelagic fishes. IOC Workshop Rep., (28):323 p.
Sharp, G.D., 1983 Tuna fisheries, elusive stock boundaries and the illusory stock concepts. Collect. Vol.Sci.Pap.ICCAT/Rech.Doc.Sci.CICTA/Colecc.Doc.Cient.CICAA, 18(3):812–29
Sharp, G.D., 1983a Ecological efficiency and activity metabolism. In Flows of energy and materials in marine ecosystems: theory and practice, editado por M. Fasham. New York, Plenum Press, NATO Conference series, IV, Marine sciences, vol. 13
Sharp, G.D. y J. Csirke (eds), 1983 Proceedings of the Expert Consultation to examine changes in abundance and species of neritic fish resources. San José, Costa Rica, 18–29 April 1983. Actas de la Consulta de Expertos para examinar los cambios en la abundancia y composición por especies de recursos de peces neríticos. San José, Costa Rica, 18–29 abril 1983. A preparatory meeting for the FAO World Conference on fisheries management and development. Una reunión preparatoria para la Conferencia Mundial de la FAO sobre ordenación y desarrollo pesqueros. FAO Fish.Rep./FAO Inf. Pesca, (291)Vol.2:553 p. FAO Fish.Rep./FAO Inf.Pesca, (291):Vol.3:557–1224
Sharp, G.D., J. Csirke y S. Garcia, 1983 Modelling fisheries: what was the question? FAO Fish.Rep./ FAO Inf. Pesca, (291) Vol.3:1177–224
Sharp, G.D. y R.C. Dotson, 1977 Energy for migration in albacore, Thunnus alalunga. Fish.Bull.NOAA/ NMFS, 75(2):447–50
Sharp, G.D. y R.C. Francis, 1976 An energetics model of the exploited yellowfin tuna, Thunnus albacares, population in the eastern Pacific Ocean. Fish.Bull.NOAA/NMFS, 74(1):36–50
Sheldon, R.W. y T.R. Parsons, 1967 A continuous size spectrum for particulate matter in the sea. J.Fish.Res.Board Can., 24(5):909–15
Sheldon, R.W., A. Prakash y W.H. Sutcliffe, 1972 The size distribution of particles in the ocean. Limnol.Oceanogr., 17(3):327–40
Sheridan, P.F., J.A. Browder y J.E. Powers, 1984 Ecological interactions between penaeid shrimp and bottomfish assemblages. In Penaeid shrimps - their biology and management, editado por J.A. Gulland y B.J. Rothschild. Farnham, Surrey, Fishing News Books, Ltd., pp. 235–53
Shirakihara, K. y S. Tanaka, 1981 A predator-prey model for two-species populations with nonlinear interactions and implications for fisheries management. Bull.Jap.Soc.Sci.Fish., 47(4): 487–93
Silliman, R.P., 1943 Studies on the Pacific pilchard or sardine (Sardinops caerulea). 5. A method of computing mortalities and replacements. Spec.Sci.Rep.U.S.Fish Wildl.Serv., (24):10 p.
Simpson, J.H., 1979 et al., The Islay front: physical structure and phytoplankton distribution, estuarine and coastal. Mar.Sci., 9:713–26
Sinclair,M., Marine populations : an essay on population regulation and speciation in the oceans. Seattle University of Washington/US Sea Grant Publication (in press)
Sissenwine, M.P., 1984 Why do fish populations vary? In Exploitation of marine communities, editado por R.M. May. Berlin, Springer Verlag, Dahlem Workshop reports, Life sciences research reports, 32:59–94
Skud, B.E., 1982 Dominance in fishes: the relation between environment and abundance. Science, Wash., 216:144–9
Slobodkin,L.B., 1961 Growth and regulation of animal populations. Nueva York, Holt, Rinehart and Winston, 184 p.
Smith, C.L., 1975 Analysis of a coral reef fish community: size andrelative abundance. Hydro-lab J., 3:31–8
Smith, C.L. y J.C.Tyler, 1973 Population ecology of a Bahamian superbenthic shorefish assemblage. Am.Mus.Novit., (2528):38 p.
>Smith, C.L. y J.C.Tyler, 1975 Succession and stability in fish communities of Dome-shaped patch reefs in the West Indies. Am.Mus.Novit., (2572):18 p.
Soutar, A. y J.D. Isaacs, 1974 Abundance of pelagic fish during the 19th and 20th centuries as recorded in anaerobic sediment of the Californias. Fish.Bull.NOAA/NMFS, 72(2):275–94
Southward, A.J., E.I. Butter y L. Pennycuik, 1975 Recent cyclic changes in climate and in abundance of marine life. Nature, Lond., 253:714–7
Steel, R.G. y J.H. Torrier, 1980 Principles and procedures of statistics : a biometrical approach. Nueva York, McGraw-Hill Book Co.Ltd., 481 p. 2nd ed.
Steinbeck, J., 1958 The log from the “Sea of Cortez”. Londres, William Heinemann Ltd. Publicado también en 1974 por Pan Books Ltd., Londres, 220 p.
Still well, C.E. y N.E. Kohler, 1982 Food, feeding habits, and estimates of daily ration of the shortfin mako (Isurus oxyrinchus) in the northwest Atlantic. Can.J.Fish.Aquat.Sci., 39(3):407–14
Swartzman, G., 1979 Evaluation of ecological simulation models. In Contemporary quantitative ecology and related econometrics, editado por G.P. Patil y M.Rosenzweig. Fairland, Maryland, International Cooperative Publishing House, pp. 295–318
Swenson, W.A. y L.L. Smith Jr., 1973 Gastric digestion, food consumption, feeding periodicity, and food conversion efficiency in walleye (Stizostedion vitraeum vitreum). J.Fish.Res. Board Can., 30(9):1327–36
Taylor, C.C., 1953 Nature of variability in trawl catches. Fish.Bull.U.S.Fish Wildl.Serv., 54:145–66
Taylor, C.C., 1962 Growth equations with metabolic parameters. J.Cons.CIEM, 27:270–86
Thayer, C.W., S.M. Adams y M.W.La Croix, 1975 The importance of eelgrass beds in Puget Sound. Mar. Fish.Rev., 39:18–22
Theilacker, G. y K.T.Dorsey, 1980 Larval fish diversity, a summary of laboratory and field research. IOC Workshop Rep., (28):105–42
Thorhaug, A., 1985 Large-scale seagrass restoration in a damaged estuary. Mar.Pollut.Bull., 16(2): 55–62
Thorpe, J.E., 1977 Daily ration of adult perch, Perca fluviatilis L., during summer in Loch Leven, Scotland. J.Fish Biol., 11:55–68
Thorson, G., 1957 Bottom communities (sublittoral or shallow shelf). Mem.Geol.Soc.Am., (67):461–534
Tremblay, C. y F. Axelsen, 1980 Données sur la pêche, la biologie et 1’abondance du flétan du Groënland (Reinhardtius hippoglossoides) dans le Golfe du Saint-Laurent. CAFSAC Res.Doc./CSCPA Doc.Rech., Dartmouth, N.S., (80/34):26 p.
Troadec, J.-P., W.G. Clark y J.A. Gulland, 1980 A review of some pelagic fish stocks in other areas. Rapp.P.-V. Réun.CIEM, 177:252–77
Tyler, A.V., 1971 Periodic y resident components in communities of Atlantic fishes. J. Fish.Res. Board Can., 28(7):935–46
Tyler, A.V., W.L. Gabriel y W.J. Overholtz, 1982 Adaptive management based on structure of fish assemblages of northern continental shelves. Can.Spec.Publ.Fish.Aquat.Sci., (59): 149–56
Ulanowicz, R.E., 1980 An hypothesis on the development of natural communities. J.Theor.Biol., 85:223–45
Ursin, E., 1982 Stability and variability in the marine ecosystem. Dana, 2:51–67
Vandermeer, J., 1981 Elementary mathematical ecology. Nueva York, John Wiley, 294 p.
Vinogradov, M.E., 1962 Feedings of the deep-sea zooplankton. Rapp.P.-V.Réun.Cons.CIEM, 153:114–20
Vlymen, W.J., 1970 Energy expenditure of swimming copepods. Limnol.Oceanogr., 15(3):348–56
Vlymen, W.J., 1974 Swimming energetics of the larval anchovy, Engraulis mordax. Fish.Bull.NOAA/NMFS, 72(4):885–99
Vlymen, W.J., 1977 A mathematical model of the relationship between larval anchovy (E. mordax) growth, prey microdistribution and larval behaviour. Environ.Biol.Fishes, 2:211–33
Walsh, J.J., 1978 The biological consequences of interaction of the climatic, El Niño, and event scales of variability in the Eastern Tropical Pacific. Rapp.P.-V-Réun.CIEM, 173:182–92
Walsh, J.J., 1981 A carbon budget for overfishing off Peru. Nature, Lond., 290:300–304
Walt, K.E.F., 1968 Ecology and resource management. Nueva York, McGraw-Hill Book Company, 450 p.
Ware, D.M., 1975 Growth, metabolism and optimal swimming speed of a pelagic fish. J.Fish.Res.Board Can., 32(1):33–41
Ware, D.M., 1980 Bioenergetics of stock and recruitment. Can.J.Fish.Aquat.Sci., 37(6):1012/24
Webb, P.W., 1975 Hydrodynamics and energetics of fish propulsion. Bull.Fish.Res.Board Can.,(190): 159 p.
Weihs, D., 1973 Optimal fish cruising speed. Nature, Lond., 245:48–50
Weihs, D., 1980 Energetic significance of changes in swimming modes during growth of larval anchovy, Engraulis mordax. Fish.Bull.NOAA/NMFS, 77:597–604
Welcomme, R.L., 1979 Fisheries ecology of floodplain rivers. London, Longman, 317 p.
Welcomme, R.L. y H.F. Henderson, 1976 Aspects of the management of inland waters for fisheries. FAO Fish.Tech.Pap., (161):40 p. Issued also in French and Spanish
Whittaker, R.H., 1967 Gradient analysis of vegetation. Biol.Rev.Camb.Philos.Soc., 49: 207–64
Wildish, D.J. y R.L. Phillips, 1974 An indentification strategy for benthos collected to assess marine and estuarine pollution. Tech.Rep.Fish.Res.Board.Can., (450):31 p.
Williams, F., 1966 Food of longline-caught yellowfin tuna from East Africa Waters. E. Afr. Agric. For. J., 31(4):375–82
Winberg, G.G., 1960 Rate of metabolism and food requirements of fishes. Transl.Ser.Fish.Res.Board Can., (194):202 p.
Windell, J.T., 1978 Digestion and the daily ration of fishes. In Ecology of freshwater fish produc tion, editado por S.D. Gerking. Nueva York, John Wiley and Sons, pp. 159–83
Yokota, T., 1961 et al., Studies on the feeding habit of fishes. Rep.Nankai Reg.Fish.Res.Lab.,(14): 234 p. (in Japanese, English summary)
Young, D.K.,1976 M.A. Buzas y M.W.Young, Species densities of macrobenthos associated with sea grass: a field experimental study of predation. J.Mar.Res., 34(4):577–92
Zaret, T.M., 1980 Predation and freshwater communities. New Haven, Connecticut, Yale University Press, 187 p.
Anon., 1981 Why bother with marine conservation? Conserv.Indon., 5(3)
