![]()
![]()
![]()
In the previous sections we have dealt with the collection of data and the estimation of the parameters of growth and total mortality. These are essential steps in most assessment work, but they do not in themselves provide assessments in the sense of statements about the state of the fish stocks, the impact on them of exploitation, and the effects of alternative policies for the future development or management of the fishery. This statement of the nature of assessment suggests that the process can be divided into three stages; first, the simple determination that fishing is (or is not) having an impact on the stock, with some indication of the extent of that impact; second, putting this qualitative statement into quantitative terms, such as the determination of the relation between yield-per-recruit and fishing effort, which will lead to the formulation of long-term strategic advice; and third, the provision of tactical advice, for example on the level of the Total Allowable Catch for next year.
The first stage is a matter of detecting some signal imposed on the stock by changing amounts of fishing. This can be of two kinds - changes in abundance, or changes in composition. The first may be detected in changed cpue in the commercial fishery, or less usually from surveys of one kind or another. In either case information on length is not relevant. The second kind of signal will arise because increased fishing will cause increased mortality, and hence result in relatively fewer old fish, and also (unless the fish do not grow much after recruiting to the fishery) fewer big fish.
In most fisheries, therefore, a simple sign of heavy fishing is a change in the size of fish. Figures 4.1 and 4.2 show the changes in the size composition of catches of plaice in the North Sea, and of hake to the westward of the British Isles as a result of the almost complete cessation of fishing during the two world wars. These are typical of the effects, though perhaps showing them more clearly than most data sets, because of the big changes in the amount of fishing over a short period. They are also typical in that there are two quite distinct changes. As expected, the post-war length-frequencies show a more extended right-hand limb, indicating a better survival. There is also a change in the left-hand limb, with the very small fish being much scarcer in the catches than in the pre-war periods of heavy fishing.
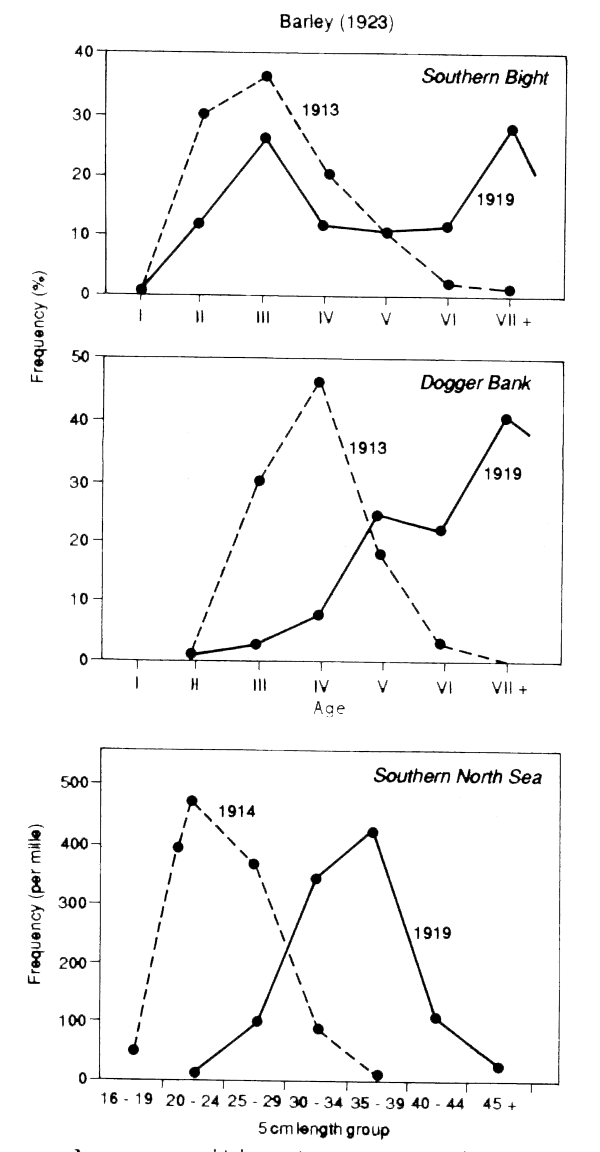
Figure 4.1 Age-composition (upper two fiqures) and length-composition of North Sea plaice (lower figure) sampled just before and just after the first world war (data from Bannister, 1977). Note that there is a shift to the right after the war and not just an increase in older and bigger fish
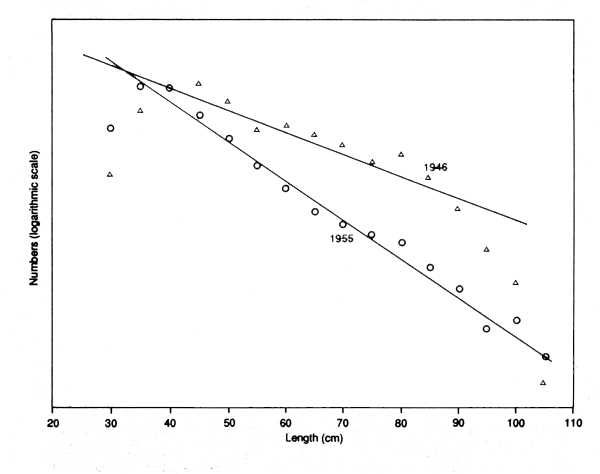
Figure 4.2 Length-composition of hake landed by English trawlers in 1946, just after the second world war, and in 1955 after a period of renewed heavy fishing. The difference in slope was taken as an index of the increase in total mortality (data from Gulland, 1956)
The change among the smaller fish is not easily explained by changes in mortality alone, and there must be some change in the pattern of entry to the catches, or perhaps more accurately the landings. The hake data, and probably the plaice data, are of commercial landings, and the relative shortage of small fish might, in part, reflect increased discarding of small fish when bigger fish are abundant. It may also reflect a shift in fishing grounds towards the areas where larger fish are abundant, and which were not attractive during periods of heavy fishing. Whatever the precise explanation, the result has been that the behaviour of the fishermen has magnified the signal (the change in size-composition) in the stock produced by the change in the amount of fishing. Thus the size-composition in the landings has become more sensitive to the effect of fishing, though it may provide a biased estimate of the extent of the effect.
Care should be used in interpreting the signal, especially in stocks that may be subject to natural variations in recruitment, and when observations extend over only a short period. Garcia and Josse (1988) point out that in such fisheries, when the most recent recruitment has been poor, fishing may be concentrated on the older fish (the survivors of earlier good recruitment) and the length-frequency will include many large fish, and indicate low mortality, and a healthy fishery. Conversely, if recent recruitment is good, fishing will concentrate on small fish, and the length-frequency will suggest high mortality and a fishery in a poor condition. The result is length-frequencies during the increasing and decreasing phases of an oscillating fishery that are very like those of heavy and lightly fished stocks (see Figure 4.3, and compare it with Figures 4.1 and 4.2). As Garcia and Josse point out, few fisheries are in equilibrium and a single snapshot, whether of age- or length-composition or any other characteristic of the fishery, is not necessarily an indication of equilibrium conditions. In practice it should be easy on the basis of the history of the fishery (trends in catch, and in available indices of fishing activity - sophisticated measures of fishing effort are not necessary for this purpose) to determine whether the difference in the two length-compositions reflects the effects of fishing or of natural fluctuations.
Even a very simple examination of changes in the length-composition over a period when the amount of fishing is changing can therefore be very valuable. In these examples the length-data confirm, in a quite independent way, the impression from the catch statistics that the stocks had become, at the beginning of each war, very heavily fished. In the case of hake the length- data also suggest that that the fishing pressure of hake had, by 1955, reached a level surpassing that of the inter-war period. The length-data also indicate that there had been changes in fishing practice involving a shift away from small fish. These may not be assessments in the quantitative sense, but they are still valuable conclusions in themselves, and in pointing the way for quantitative assessments.
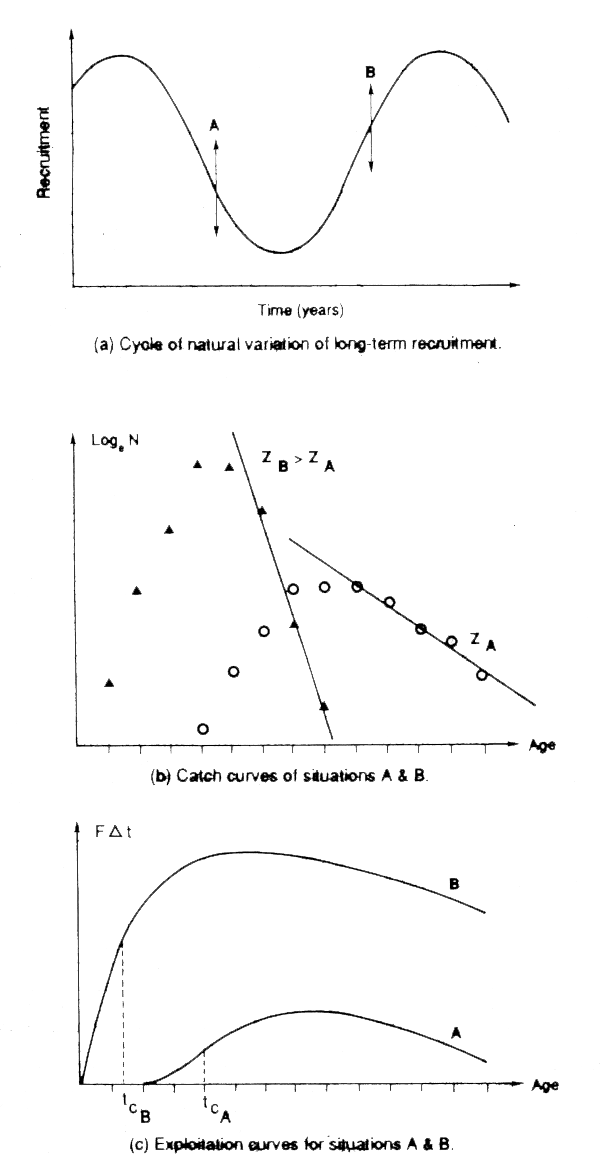
Figure 4.3 The danger of shot assessments in a fluctuating fishery. Note the difference in catch-curves (middle figures) taken at different points in the cycle (from Garcia and Josse, 1988)
A simple measure for monitoring the stocks, in the absence of more detailed analysis, is provided by the mean length of fish in the catch. An early example is that of Henderson (1972), who examined data for a trout fishery in Colombia (Figure 4.4). During the nearly twenty years of data there was a steady decrease in mean length, possible with two periods of relatively little change, as indicated by broken lines. This strongly suggests increasingly heavy fishing, and this is a conclusion that could not have been obtained from other available sources. Examination of mean length (based on regular samples, even if they are not large) should be done to monitor the “health” of all fisheries, even when there is no time, or resources, to set up a proper stock assessment programme.
A long-term assessment, aimed at producing advice on the average steady-state relations between the amount and selectivity of fishing and the catches, will be based on one or other of the standard families of models (production models, or dynamic pool models). Length-data (like age-data) are not normally of direct use in applying production models, but there are exceptions. Csirke and Caddy (1983) noted that the standard relation between fishing effort and long-term catch could be rewritten to relate catch to total mortality. That is, from the relation
| C = af + bf2 | (4.1) |
where C = catch, f = effort and a and b are constants, and writing Z = F+M, and F = qf, we have
| C = (a/q) (Z-M) + (b/q2) (Z-M)2, | (4.2) |
Thus, if the catch in any period is plotted against the estimated total mortality in that period the result should be a parabola, similar to that obtained from the more usual plot of catch against fishing effort, but displaced to one side so that it cuts the x-axis where Z = M, as well as at some high value of Z. This approach can be applied to any set of length-data from which values of Z/K have been obtained by any of the methods described earlier.
This is a deceptively attractive method, but there are problems. Practical experience of those stocks in which effort has varied over a wide range suggests that the relation between catch and effort is seldom a parabola, but descends more gradually at high levels of effort. The relation, whether parabolic or otherwise, only holds for stocks in equilibrium, so that the method can strictly only be applied to a number of periods during each of which the fishing effort has remained constant, whereas on most stocks fishing effort is changing continuously. If it has been increasing, the catch in the current year is likely to be higher than it would be in a steady state with the current level of effort. The latter problem may be in part balanced by the fact that the mortality rate estimated from the length-data in say 1987, is likely to measure the mean mortality over the last few years, rather than in 1987, but the balance is unlikely to be exact. The same problem occurs in the normal method of applying the production model, but various methods, from Gulland's (1961) method of considering average effort over some earlier period, onwards have been developed to deal with this.
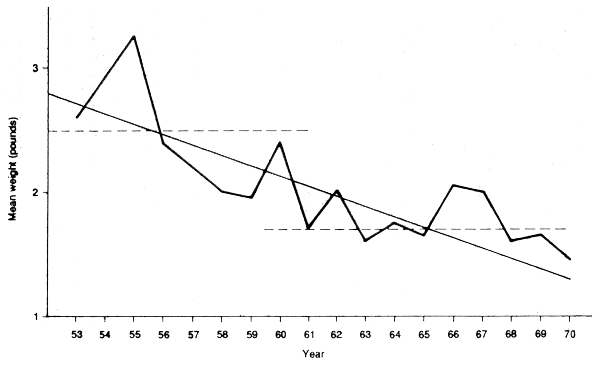
Figure 4.4 Changes in mean weight in the Colombian trout fishery as an index of the intensity of fishing (from Henderson, 1972)
The greatest difficulty in applying Csirke and Caddy's method is likely to be in fitting the theoretical curve to a set of observations. Fitting a parabola or any strongly non-linear relation is always difficult, and the statistical problems are increased by the fact that there is likely to be considerable error in the independent variable (Z/K), as well as natural variation in the catch. It is always worthwhile looking at alternative methods of assessment, but in this case the results should be treated with considerable caution unless the plot of catch against Z/K gives a very clear relation.
Length data are well suited to use in many forms of the dynamic pool models. The commonest formulation of the dynamic pool model is that of Beverton and Holt (1957). In its simplest form this assumes recruitment is constant, or is independent of adult stock, and the assessment is concerned with seeing how the yield-per-recruit (Y/R) varies with the amount of fishing and the size at the first capture. Their expression for yield-per-recruit can be rewritten in the following simplified form (Beverton and Holt, 1964);
| Y/R = aE (1 - c)M/K X Sum{Un(1 - c)n/[1 + n(K/M) (1 - E)]} | (4.3) |
where E is the exploitation rate F/Z
a is a constant
c is the relative size at first capture 1c/Linf
n is a dummy variable, the summation being over the values 0 - 3
and Un takes values of +1, -3, +3 and -1 for the n values between 0 and 3.
This expression makes it clear that the basic relations between Y/R and the amount of fishing or the size at first capture depend only on three ratios. One, M/K is characteristic of the fish stock, and the others E (=F/Z) and c (=1c/Linf) are determined by the intensity of fishing and the size of first capture. Since c relates directly to length, and most of the length-based methods of estimating mortality give the results in terms of ratios such as Z/K, it is seen that this formulation can be used almost immediately once the parameters of growth and mortality have been estimated as in section 3.
Apart from expressing the yield-per-recruit as a function of simple ratios, the revised Beverton and Holt formulation also makes it clear that the variety of Y/R relationships (given the assumption that fishing mortality is constant above some selection point) is not large. In fact there is a single family of relations, depending on the value of M/K, and for each member of the family there is a two-dimensional surface relating Y/R to the amount of fishing and to the size of first capture. The usual Y/R curves are then slices through this surface. Calculating a Y/R curve (or surface) involves nothing new, and should now be a matter of applying a simple computer programme, which can be done on spreadsheets (Gulland and Gibson, in press) or just by looking up the table in Beverton and Holt (1964) for the appropriate value of M/K. It should not in itself be considered as carrying out an assessment.
The simplest type of stock assessment, in the context of Y/R curves, consists of two phases: identifying the curve (or surface) corresponding to the biological characteristics of the stock, and locating the present position of the fishery on this curve. In both of these the estimation of M (or M/K), or the division of the total mortality between fishing and natural mortality is an important step and one that usually presents the biggest problem. If one has been lucky to start investigations on an unexploited stock then in that stock M = Z and any of the methods of the previous section to estimate Z (or Z/K) will produce an estimate of M (or M/K). Once a fishery starts, then mortality will increase, and the amount of the increase in Z/K will provide an estimate of F/K. This points to the advantages of studies carried out before fishing begins.
More usually studies do not begin until a fishery is well established. In these cases most of these methods will produce estimates of Z (or Z/K). There are then several ways of proceeding:
separating F and M by relating changes in Z to changes in the amount of fishing;
estimating F directly, e.g., by tagging, or from the ratio of catch to the biomass as estimated by some kind of survey;
estimating M directly e.g from the consumption by predators. This method is at best approximate, and likely to provide a lower bound, but some recent studies in the North Sea (ICES 1989) show that this method can provide interesting results.
estimating M by comparison with the values of M in other species.
The first method is the one that is most likely to give statistically sound results, and should be used when possible. Its typical age-based form consists of regressing Z on the fishing effort f. Exactly the same procedure can be used with length-based estimates, either of Z, or as occurs with some methods, of Z/K. In the latter case we obtain estimates of M/K and F/K.
Even when it is not possible to apply the regression method exactly for some reason, e.g., because there is no exact measure of effort, an examination of less quantitative material can provide guidance on the relative values of F and M. Thus Gulland (1956) used the slope of the lines in Figure 4.1 as indices of Z. By noting that the fishing effort during the war was around one third of the pre-war level, he estimated that in the pre-war period fishing accounted for at least half the total mortality. Even in situations where the data are still less quantitative some approach of this kind should be made. Formal effort data may be completely lacking, but some idea of how the amount of fishing has been changing can usually be obtained from records of one kind or another - What is the impression of local fishermen? Have new markets opened up? In many cases these records will indicate that there have been changes in the amount of fishing, and then one needs to see whether there are any changes in the stock that might give some measure of the impact of fishing. Are large fish less frequent than they used to be? Have catch rates declined? If these signs have occurred, then it is reasonable to suppose that fishing is having an impact and that F is now a non- negligible proportion of Z. If they have not, F is probably still small.
A length-based version of examining the effects of fishing on the composition of the population, that does not require any direct estimate of fishing effort (or cpue), but relies on accurate statistics of total catch as an index of the amount of fishing, has been developed for sperm whales (Beddington and Cooke, 1981; Cooke and Beddington, 1982). The greater the change in length-composition that is observed following a catch of a given magnitude, the lower must have been the initial population and the higher the fishing mortality. The method requires that the total numbers caught in each length-group are known, and assumes either that the selection pattern of the fishery as a whole is constant, or that the catch data for each year can be broken down into small time/area strata, within each of which selection is the same from year to year. Alternative initial values of population numbers and growth parameters can be used to project the population forwards, and the set of parameter determined that give the best fit between observed and projected catches by length-group. The data requirements and assumptions about constant selectivity are rather stringent. Thus, although the method is equally applicable in principle to any fish stock in which there is appreciable post-recruitment growth, in practice its application to finfish is likely to be limited. Further details of the method are given in the International Whaling Commission reports, and a FORTRAN version of the program is available from the authors and from the IWC.
The second method, of estimating F as the ratio of catch to biomass does not formally involve length, and does not need discussion in this manual. However, it is worth noting that it is unlikely that the biomass, as measured by the survey gear, is precisely the same biomass as exploited by the fishery. Generally the survey will include smaller fish (for example, a trawl survey is usually done with a small mesh in order to collect information on pre-recruits and on small unexploited species). In that case the survey biomass should be adjusted downward to cover only the exploited part of the stock. This will be most easily done on the basis of length. If recruitment to the exploited stock is fairly abrupt, at some length lc, then all fish smaller than lc should be omitted from the survey data, and the biomass recalculated accordingly. If selection occurs over a wide range of sizes a value of lc may be difficult to determine. Then it may better to take some arbitrary value of lc above the selection range and calculate the weight of catch and stock above lc.
The third method, of directly estimating that part of M accounted for by predation is likely to be of value in fisheries with large research programmes, but is possibly of limited immediate relevance to many users of this manual. However, it is worth pointing out here that, because predation rates are largely determined by the relative sizes of predator and prey, a length-based approach to this method is likely to be fruitful. Pope and Yang (1987) describe how this may be done, using a multi-species extension to Jones' (1984) length cohort analysis. It is also worth noting here that this approach is providing much new insight into how natural mortality is determined. The North Sea results show that the predation mortality among the smaller sizes of fish is much greater than the values of natural mortality previously used for those species, and that M for a given species almost certainly can decrease quite substantially with size. To the extent that the abundance of the predator populations can vary from year to year, due for example to the impact of fishing, it is probable that natural mortality also changes, as suggested by Munro (1982) and others. Though such changes do not seem to have been clearly demonstrated, principally due to the problems of obtaining two good estimates of M, this is no reason for not believing that they can occur. To the extent that these changes with size and time occur it may be misleading to talk about “the natural mortality of mackerel” in a particular area as though it was a unique and unvarying quantity.
In practice none of the first three methods may give usable numbers for M (or F, in the common case when Z can be estimated). If a long-term assessment is to be presented, some assumption about M, or M/K, has to be made if the dynamic pool approach is to be followed. It is natural in such cases to fall back on the comparative method. The commonest form to use is Pauly's (1980), which has been used so often in length based studies (e.g., in the ELEFAN packages) that it is sometimes thought of as a length-based method, which it is not. Pauly's equation is
| logM = -0.007 - 0.279 logLinf + 0.654 logK + 0.463 logT | (4.4) |
where T is the mean environmental temperature Celsius. This formula has potentially a great advantage in that confidence limits on the estimate are available. The regression on which this equation is based has a residual standard deviation about the regression of 0.245, or transforming from logarithms (these are to the base 10), the width of the approximate 95% confidence limits of 2.5 times the central estimate (i.e., if the estimate is 0.2 the confidence interval is approximately 0.07-0.57) about the central estimate given by equation 4.1. These are wide limits, so it is perhaps not surprising that they are rarely if ever used. It may be argued that they may be too wide, since they take into account the undoubted uncertainty in the estimates used by Pauly as well as the true variability from stock to stock. On the other hand the uncertainties in K, Linf and T for the stock being studied (which may be large) are not taken into account. They would make the true confidence limits for the estimates of M wider than given here. The degree of variability can often be reduced by considering only closely related species (e.g., clupeoids) since there is good evidence (Beverton and Holt, 1959; Beverton, 1963) that there are greater similarities in population parameters within groups of fishes than among fishes as a whole. Against this it can be argued that many of the estimates of mortality used by Pauly were published after the general pattern of values of M were known and authors tended, when their estimates were poor, only to publish those that fitted expectations.
Whatever the truth of these arguments it is clear that any figure for M produced from Pauly's equation must be subject to considerable uncertainty. Even if the standard deviations presented by Pauly may be too wide, the failure to take explicit account of substantial uncertainty is likely to be misleading. Possible confidence limits which reflect the likely spread might be ones extending half as far above the central estimate as the confidence limits suggested above. Then, if the central estimate of M from equation 4.4 is M = 0.36, our conclusion should be that the true value of M is somewhere between 0.21 and 0.61, and these values should be used as alternatives in the further assessments. In some circumstances, for example if our estimate of Z is 1.55, the range is narrow enough to show that the stock is heavily fished (M is at most just less than half Z), but more often the status of the stock will remain unclear. It was perhaps optimistic of Pauly (1987, p. 23) to claim that, when using ELEFAN II (which includes this method of estimating M), “it is in principle possible to assess whether or not a stock is over-fished, based solely on length frequency data obtained from that stock” - though the point is not so much that this is an unrealistic belief in the power of length-based methods, but that any method has difficulty in assessing the state of a stock unless there are observations of the stock and its size- or age-composition under different intensities of fishing.
The point is that, when there is uncertainty about the value of a parameter, subsequent computations should be done using a range of values applies to any parameter. It is stressed here because the uncertainties about M (or M/K or the ratio E = F/Z), whether based on Pauly's equation or other methods, are usually much greater than in other parameters.
Though length methods are considered an innovation in much assessment work, they have a long history of use in estimating the effect of selection changes, particularly in the North Atlantic (Gulland, 1961a; 1964). The principle used is simple; from the change in selectivity and the length-composition of the current catches the immediate change in the numbers caught following a change in selectivity can be calculated. In the long run, catches will increase when the fish released have grown enough to be caught by the new gear. There are two ways of estimating this increase; the original method of Gulland, which computes this increase in the numbers caught as the product of the numbers released times the exploitation ratio (F/Z), reduced by the natural mortality prior to the time when they are big enough to be retained by the larger mesh; and the method of Cadima (1978) which computes the change in fishing mortality.
The basic calculations are set out in Table 4.1, derived from the haddock data given by Gulland (1983). For each length group the number retained can be calculated as nk=n1p2/p1 where p1 and p2 are the proportion retained by the original and new gear respectively, as determined by selection experiments where n1 = numbers in the original catch, and nk = numbers retained with the new gear immediately after the change. The total numbers N and weight W caught before and immediately the change can then be calculated as the sums of n1, n1wL nk and nkwL.
| Length | Mean weight | % retained | Ratio | Numbers of fish | |||
|---|---|---|---|---|---|---|---|
| 90 mm | 120 mm | Initial | Retained | Released | |||
| w1 | p1 | p2 | n1 | nk | nR | ||
| 25 | 160 | 50 | 3 | .06 | 8 | 0.48 | 7.52 |
| 30 | 260 | 80 | 16 | .2 | 314 | 62.8 | 231.2 |
| 35 | 420 | 98 | 100 | .42 | 1084 | 435.5 | 630.5 |
| 40 | 630 | 100 | 100 | .72 | 1409 | 1014.5 | 394.5 |
| 45 | 880 | 100 | 100 | .94 | 1370 | 1287.8 | 82.2 |
| 50 | 1190 | 100 | 100 | 1 | 952 | 952 | 0 |
| 55 | 1570 | 100 | 100 | 1 | 465 | 465 | 0 |
| 60 | 1760 | 100 | 100 | 1 | 255 | 255 | 0 |
| 65 | 2530 | 100 | 100 | 1 | 124 | 124 | 0 |
| 70 | 3100 | 100 | 100 | 1 | 73 | 73 | 0 |
| 75 | 3800 | 100 | 100 | 1 | 30 | 30 | 0 |
| 80 | 4600 | 100 | 100 | 1 | 9 | 9 | 0 |
| Total numbers | 6093 | 4727 | 1365 | ||||
| Total weight | 5639 | 4986 | 652 | ||||
| % of initial numbers | 77.6 | 22.4 | |||||
| % of initial weight | 88.4 | 11.6 | |||||
From the table the immediate effect of the change is given as the ratio of the weight released to initial weight i.e. WR/W1 = 652/5639 = 11.6%. Using Gulland's approach, in the long run an additional number of fish, ENR, will be caught, where NR' = NRexp(-Mdt) where dt is the time taken by the fish to grow from the mean selection length of the two gears. The catches will then increase by a factor A = Nk/Nr'. The long term weight W2 is given by W2 = Wk (1 + A). Using Cadima's method the long term weight W2 is given by W2 = Wkexp(Fdt), and exp(Fdt) can be shown to be equal to (N1/Nk), where E' is the value of E = F/Z in the size range between the selection sizes of the two gears.
The method can be expanded to give the interim effects during the period between the change and the time when the new long-term equilibrium is reached (Gulland 1964). To do this we note that, if all fish grow at the same rate, then at any given time in the transition period, the older and larger fish (above length 1" say) will be unaffected by the change while the fish smaller than 1" will have reached the new equilibrium. The weight caught W" will then be given by
W" = WK,L + W1,L (1 + A) = Sum [WK,L (1+A)]
1>1" 1>1"
Like many length-based methods this method makes the assumption that the fishery is in equilibrium. Its particular advantage is that it can deal with a situation in which several gears exploit the same stock (e.g., trawlers and longliners), some of which may not alter their selectivity. In this case the values of N1, Nk, W1 and Wk are calculated for each gear separately. The long-term effects and the immediate effects can then be compiled after calculating NR as the sum of all the values of iNr for the individual gears.
Jones (1984) has pointed out that the procedures of cohort analysis (see section 3.2.4) can be extended to provide estimates of the changes in long-term catch as a result of changes in fishing mortality. For any length-group the numbers present in the new equilibrium state, 2Nl, can expressed as
2NL = 1NL × CF
where CF is a correction factor due to changed mortalities, given by CF = exp[sum(1Fdt) - sum(2Fdt)] and 1Nl are the numbers present in the original state, 1F and 2F are the original and new fishing mortalities, dt is the time taken to grow through the length interval, and the summations are taken from the recruitment size up to the middle of the lth length interval.
The new catches in each length interval, expressed as weight or numbers, will then be given as
2CL = 1CL × CF × 2FL/1FL and 2WL = 1WL × CF × 2FL/1FL.
It can be seen that there need be no assumption of F being the same for all age groups. It is thus possible to use the method for simultaneous changes in the amount of fishing and in selection. An example of this is given in Table 4.2 (taken from Table 7 of Jones, 1984). The caution to be noted in using the procedures in this table is that the summations of F (columns 7 and 8) are taken to the middle of each length-group, while most of the calculations in VPA and cohort analysis are to the boundaries of the groups.
The approach set out in Table 4.2 follows directly from Jones original form of length-based cohort analysis set out in Table 3.5. It is of course possible to use the cohort slicing approach as in Table 3.6. In that approach, once the slicing is done, then the individual slices can be treated as age-groups (actually half-year groups in Table 3.6, because the fish were short-lived). In that case the data can be analysed using standard age-based techniques, with the proviso that the results will become progressively less reliable among the larger fish as there is blurring of age-groups across slices.
Sparre et al. (1989, chapter 5) have suggested a very similar set of calculations, starting with the approach of Thompson and Bell (1934). It can be viewed as the reverse of cohort analysis (or VPA) in that instead of working back from the observed catches from a cohort to deduce the fishing mortality, it works forward from the numbers of a cohort recruiting to the fishery, to deduce (given the mortality in each length-interval) the numbers caught, and also the weights. An advantage of this method is that it incorporates information on the value of the catch. This can be important for those fisheries (e.g., on shrimp) where the price per kilogramme differs according to the size of the animal.
Most of the assessment techniques discussed so far deal with steady-state conditions, and thus provide long-term strategic advice. Is it desirable to increase fishing effort by building new boats? Is the stock so heavily fished that measures to manage the fishery are necessary? What would be the long-term effect of increasing the mesh size by 10 mm? This is probably the most valuable type of advice, but increasingly, especially when management is being considered, short-term advice is needed. What is the immediate effect of using a bigger mesh? How long after reducing fishing effort on a heavily fished stock will it be before catches return to their original level (or better)? This type of advice has become particularly important in the parts of the North Atlantic and other areas where the amount of fishing is controlled by catch quotas or TACs.
Though some of the techniques described above, e.g., Gulland's mesh change method, can provide information on short-term interim effects, most of the length-based techniques deal with steady states, and age-based methods are usually better (because they have a simple time-scale) for studying short term effects. There may be exceptions.
One possible exception is the prediction of future catches in setting TACs. The stocks concerned are mostly assessed by cohort analysis or VPA. Future stock sizes and catches (in numbers) can be predicted by projecting the matrix of numbers at age forward one year. The total weight caught can then be estimated by multiplying the numbers by the average weight-at-age. This procedure is fine provided the average weights do not change. In practice there are often secular changes in mean weight which can sometimes lead to serious errors in predicting TACs.
Shepherd (1987) has pointed out that essentially the same technique can be used on length-data, though the projection is more complex. For age-data
Ni+1,j+1 = Ni,je-z
where Ni,j are the numbers at age j in year i
For length-data the projection is more complicated, since it is unlikely, even if all fish follow exactly the same growth pattern, that all the fish in one length-group in one year will all fall in a single length group in the next year. For example fish between 60 and 65 cm in one year may be between 67.5 and 71.7 cm in the next year, and thus are divided between two different 5-cm groups. Nevertheless, by considering the sizes, in the second year, of fish initially at the boundaries between adjacent length-groups (e.g., 10, 15, 20 cm, etc.) in the first year, and applying the expected mortality rate, it is possible to calculate expressions for the expected frequencies in each length-group in the second year, as a function of the first-year frequencies. The exceptions are the smallest sizes, which are occupied by recruits. To the extent that departures in the mean weights-at-age in the second year from the expected values are due to unusual growth in the earlier years, rather during the transition period, this approach should be less disrupted than the age-based approach.
So far this method has not been tested sufficiently to be sure that this is the case. Shepherd (1987) has applied the approach to data from surveys of cod in the North Sea. The results were promising, though the predictions were not very accurate, possibly because there was a lot of variability in the original data. Another possible advantage of the method is that, because the input data are simple, and there is no work involved in processing and reading otoliths and scales, there can be a shorter lead-time between making the observations and producing the predictions. This in itself could lead to greater precision. For the present though not enough has been done to apply the method in practice to be able to recommend it.
One big difference between age-data and length-data is that the latter can be collected in a much greater volume, and large numbers of different length samples of meaningful size (say 200 fish plus) can be collected from different places and times. In contrast the practical constraints of manpower mean that few age samples of comparable size can be collected. For age-data therefore we are usually constrained to look at pooled data, for the fishery as a whole during a month or even a year. For most of the methods described in this report this difference does not matter - most of the methods use pooled data. However, the act of pooling does throw away information that could be of use for other purposes.
One obvious use of detailed information on length composition in management is to identify those areas, times or gears in which small fish are unusually abundant. Closing fishing on nursery grounds, or at times when small fish are recruiting, or discouraging fishing by gears that take predominantly small fish is often the best way of improving the exploitation pattern, i.e., moving the effective size at first capture upwards and thus increasing the yield-per-recruit. This identification can be done in many ways. Any differences between fishing grounds are probably seen by plotting the mean length of each sample at the position of capture on a chart of the fishing grounds.
Looking at individual samples can also help in the interpretation of many of the techniques described earlier. For example in Figure 3.3a (p. 26) the length distribution for September clearly does not fit the predicted growth-curve. It would be helpful to know if this was a single sample, and if the fish were caught in the same area as the other samples. It would probably also be helpful to know whether the large number of fish in January (1 390) came from several samples with the same length distribution, or whether the observed broad distribution arises from the combination of several samples with different modal lengths.
It is not the purpose here to set out recommended ways to proceed with detailed length composition data. The point to be made is that these data probably contain much useful information and looking at and arranging the data in different ways is part of the essential process of understanding the data which should precede any application of the process of understanding the data which should precede any detailed data can be quite simple, but equally sophisticated methods can, and should, be used when appropriate. Thus McGlade and Smith (1983) applied principal component analysis to length data of Canadian commercial landings of pollock to provide insight into the patterns of movement and distribution of this species.
![]()
![]()
![]()