![]()
![]()
![]()
REPARTITION DES ESPECES DEMERSALES DU PLATEAU CONTINENTAL
IVOIRIEN (ET DU GOLFE DE GUINEE)
par
CAVERIVIERE Alain
Chargé de Recherches de l'ORSTOM
On trouve dans d'assez nombreux ouvrages des renseignements sur la répartition et l'abondance des espèces démersales peuplant le plateau continental de l'Afrique de l'Ouest, qui constituent une grande unité faunistique appelée province équatoriale ou guinéenne. Les limites de cette province sont assez variables selon les auteurs cités par Postel (1968) et nous considérerons qu'elle s'étend du Sénégal au Congo, suivant en cela Ekman (1963) et Longhurst (1965), ce dernier écrit qu'il n'y a pas d'importantes différences dans la composition spécifique des pêches côtières effectuées dans cette vaste zone.
Cependant, la plupart des études n'ont porté que sur une ou quelques espèces particulières et les travaux d'ensemble sont plus rares. Le plus complet - et qui porte sur une zone d'upwellings saisonniers bien marqués avec deux saisons marines chaudes et froides comme sur le plateau continental ivoirien - concerne le Congo (Durand, 1967) et servira souvent de base de comparaison.
Les communautés de la province guinéenne ont été définies par Longhurst (1969) dont nous utilisons la classification, que nous pourrons quelquefois moduler à partir de nos données. Nous ne nous intéresserons qu'à la répartition des espèces quantitativement les plus importantes et laisserons de côté certains peuplements dont les espèces sont très marginales dans les pêches ivoiriennes.
L'étude de la répartition des espèces est essentiellement basée sur les résultats de la radiale de Grand Bassam visitée régulièrement entre mars 1966 et février 1967 et également sur ceux des campagnes de chalutage CHALCI 78.01, 79.01 et 80.01. A l'intérieur de chaque communauté les espèces seront citées dans l'ordre de la systématique, tel que suivi par Blache et al. (1970).
1. PRESENTATION DE L'ETUDE
1.1 Les données
1.1.1 Les radiales
Des pêches ont eu lieu toutes les trois semaines, de mars 1966 à février 1967 (18 sorties), sur la radiale de Grand Bassam (Troadec et al., 1969) située un peu à l'est d'Abidjan et sur 1l immersions : 15, 20, 25, 30, 35, 40, 50, 60, 80, 100, 200 m. Les pêches, d'une durée d'une heure à une vitesse moyenne légèrement supérieure à 3 noeuds, ont toujours été effectuees de jour et d'ouest en est. On considèrera que les répartitions bathymétriques des espèces sur cette radiale représentent assez bien les répartitions diurnes pour l'ensemble de la Côte d'Ivoire. Le chalut de fond employé, de type Le Drezen, avait 24 m de corde de dos et un maillage du cul de 40 mm (vide de maille), il était tire par le N/O REINE POKOU, chalutier classique de 300 CV pêchant par le côte. Au total 196 traits de chalut ont été effectués en 1966–67 sur cette radiale, dont 126 entre 15 et 50 m et 54 entre 60 et 200 m.
Nous utiliserons aussi parfois les résultats des pêches effectuées - avec la même méthodologie et à une périodicité de deux mois - sur des radiales assez régulièrement espacées le long du littoral ivoirien et situées d'est en ouest près des villes de Jacqueville, Grand Lahou, Fresco et Sassandra (Bouillon et al., 1969). Sur ces radiales les profondeurs de pêche ne furent comprises qu'entre 15 et 45 m.
La nature des fonds pour toutes les radiales, de Grand Bassam à Sassandra, est donnée sur la figure 1 d'après les cartes de Martin (1973). Les conditions physico-chimiques au dessus de la radiale de Grand Bassam sont représentées sur la figure 2 (A à C). Les conditions de température et de salinité au niveau du fond, lors des traits de chalut effectués sur la radiale de Grand Bassam (fonds de 15 à 100 m), peuvent être reparties dans des classes d'eau dont les limites et les appellations sont données sur la figure 3. Ces classes sont proches de celles utilisées par Troadec (1971) pour l'étude du Sciaenidae Pseudotolithus seneqalensis. La principale modification consiste en la séparation à 19°C des eaux 4 et 5, toutes deux appelées eaux de la thermocline par cet auteur, alors que les eaux 5 sont nommées subtropicales dans la présente étude. En effet, le point bas de la thermocline, qui n'est jamais très net, se situe vers 18°–20° quand celle-ci existe.
La superposition de notre grille des différentes classes d'eau sur celles des abondances en fonction de la température et de la salinité (carrés de 1°C et de 0,1°/oo), données par Troadec et al. (1969) pour les principales espèces capturées lors des radiales Grand Bassam, nous permettra de commenter rapidement la répartition des espèces en fonction des types d'eau. Les mensurations effectuées pendant les radiales (Caverivière, 1976) pourront également être utilisées dans l'étude des répartitions.
1.1.2 Les campagnes de chalutage
Les résultats des campagnes CHALCI 78.01, CHALCI 79.01 et CHALCI 80.01 pourront être également utilisés pour préciser certains aspects de la répartition des espèces qui n'apparaissent pas à partir des résultats de la radiale de Grand Bassam. Ces campagnes ont consisté en un échantillonnage stratifié de tout le plateau ivoirien entre 10 et 120 m (Caverivièe et Champagnat, 1978 et 1979; Caverivière, 1980 ). Elles ont eu lieu en février 1978, mars 1979 et août-septembre 1980 à partir du N/O ANDRE NIZERY, chalutier arrière de 425 CV utilisant un filet de 24,6 m de corde de dos avec un vide de maille de 39 mm au niveau du cul. La duree standard des traits est de 1/2 heure. Au total plus de 200 traits de chalut pourront ëtre analysés à partir des campagnes CHALCI pour l'étude des répartitions spécifiques. Notons que les deux premiéres ont eu lieu en saison chaude et la derniére en saison froide, ce qui correspond proportionnellement assez bien à la dure de ces saisons.
Le chalut employé par le N/O ANDRE NIZERY est proche de celui utilisé par le N/O REINE POKOU, avec une vitesse en pëche similaire, de ce fait, il ne devrait pas y avoir de modifications notables dans les proportions des prises spécifiques dues au train de pëche. Cette remarque s'applique egalement pour les comparaisons que nous serons amenés á faire avec les résultats du N/O OMBANGO au Togo, Dahomey, Cameroun, Congo (Crosnier, 1964; Crosnier et Berrit, 1966; Durand, 1967).
Les rendements utilisés not été obtenus à partir de traits diurnes. Nous savons (Caverivière et al., 1982) que les variations nycthémérales modifient considérablement les abondances apparentes pour la plupart des espéces et pourraient entraîner des changements notables dans la répartition bathymétrique de certaines d'entre elles.
1.2 Importance des espèces
Le tableau 1 indique le pourcentage moyen des prises (sup. à 0,5 %) des principales espèces capturés de 15 à 100 m sur la radiale de Grand Bassam. Les prises ont été pondérées par des facteurs K (Tableau 2) calculés à partir des cartes de Martin (1973), qui permettent de tenir compte de l'écartement des stations (*). Dix espèces ont un pourcentage supérieur à 2 % et représentent 76,4 % du total, dix autres sont comprises entre 2 et 0,5 % et représentent 12,5 % du total. Les deux premières espèces (Brachydeuterus auritus et Ilisha africana), qui forment le tiers des prises, sont semi-pélagiques. Rappelons que nous nommons ainsi les spèces pèchées à la fois en bonne quantité par les chalutiers et les sardiniers, c'est-à-dire que ces espéces ont un lien assez làche avec le fond et peuvent se trouver régulièrement entre celui-ci et la surface en formant en général des bancs plus ou moins denses.
Nous donnons dans le tableau 3 la liste de toutes les espèces capturées de 15 à 100 m lors des radiales Grand Bassam de 1966–67, avec les prises horaires moyennes obtenues à chaque profondeur. Nous ne nous intéresserons, par la suite, qu'aux espèces des communautés des Sciaenidés, des Sparidés (élément còtier et élément profond) et celles dites “eurybathes ou de la thermocline”. La communauté des Lutjanidés décrite par Longhurst (1969) existe bien en côte d'Ivoire, mais elle ne représente qu'une trés faible proportion des mises à terre de la pèche industrielle du fait que les substrats préférentiels au dessus desquels se tiennent les espèces de cette communauté ne sont pas favorables au chalutage. Pour la méme raison elle n'est pas représentée dans les radiales. Les communautés profondes du plateau et de la pente continentale ne sont pas exploitées.
1.3 La variabilité intrinsèque
Avant d'examiner les espéces une par une, nous rappellerons brièvement que le chalut n'est pas un instrument parfait pour échantillonner une population et qu'il existe des variations aléatoires de l'abondance toutes conditions égales par ailleurs (dans la limite des observations). Ces variations sont en général fonction du caractère plus ou moins grégaire des espèces et on pourra se reporter plus particulièrement à Durand (1967) pour plus de détails. Cet auteur estime en première approximation à 30–40 % la marge d'erreur possible pour un trait de chalut du fait des variations aléatoires. Nous pouvons illustrer cette variabilité à partir de 24 traits de chalut effectués avec une périodicité de 2 heures au même endroit (fonds de 35 m ± 1 m) lors de la campagne CHALCI de mars 1979. Les prises globales sont présentées sur le tableau 4 avec les températures au niveau du fond. A 24 heures d'intervalle ces prises ont varié de 0 à 82 % pour des conditions assez similaires : la différence de température peut atteindre 2,5°C, mais un écart de 63 % est quand mëme enregistré pour une différence inférieure à 0,5°C. Nous verrons par la suite (cf. Annexe VI du présent volume) qu'une partie de cette variabilité peut s'expliquer par l'action d'ondes internes à courtes périodes, la part de la variabilité considérée comme aléatoire restant cependant importante. A l'intérieur des prises globales, les écarts élevés proviennent surtout d'espéces gregaires à caractere semi-pélagique telle Brachydeuterus auritus.
Notons comme facteur pouvant ëtre considéré comme aléatoire la présence ou non de quantités importantes de méduses sur les petits fonds. Une baisse nette de l'abondance a été observée dans le premier cas lors des campagnes CHALCI. Ce phénoméne avait déjá été noté par B audiri-Laurencin (1967) au Congo et Troades (1971) en Cöte d'Ivoire, plus récemment Franqueville (1980) le signale au Sénégal.
Nous concluerons au sujet de la variabilité intrinsèque qui, bien sôr, ne touche pas que l'étude des répartitions mais toutes celles utilisant la pèche comme moyen d'échantillonnage, que le calcul de moyennes doit cependant permettre d'accorder une certaine valeur aux résultats obtenus à partir de series suffisamment longues.
2. COMMUNAUTE DES SCIAENIDAE
Dasyatis margarita (Dasyatidae). Cette raie, dite “raie perlée”, été rencontrée de 15 à 35 m ; elle est particulierement abondante sur les petits fonds. Son pourcentage d'occurrence (*) à 15 m approche 90 % avec une moyenne de 15 kg par trait. Elle a été trouvée de 18 à 30°C et de 33,8°/00 à 35,7 °/00 de salinité. Sa présence a donc été observée jusqu'au point bas de la thermocline, mais les fortes abondances n'apparaissent qu'á partir de 21°C. Sa distribution est analogue au Bénin, Cameroun, Congo (Crosnier, 1964 ; Crosnier et Berrit, 1966 ; Durand, 1967).
Ilisha africana (Clupeidae). Ce Clupeidae cotier d'assez petite taille et de forme aplatie est surnomme “rasoir” du fait que le bord de l'abdomen présente des écailles en saillies. Deuxième espece en importance sur la radiale avec 15,7 % des prises, elle est assez grégaire (810 kg ont été pris à 25 m lors de la radiale Grand Bassam 15) et présente un caractére semi-pélagique marqué : Ilisha africana représente environ 1 % des prises des sardiniers ivoiriens, mais il faut noter que ceux-ci ne la recherchent pas et la rejettent souvent quand elle est capturée.
Le rasoir a été récolté de 15 à 50 m avec une abondance maximale à 25 m (Figure 4). Son pourcentage d'occurrence est trés proche de 100 % sur les fonds de 15 à 30 m. Les captures s'échelonnent de 17 à 30°C et de 33,8 à 35,8 °/oo et montrent une préférence de l'espéce pour les eaux chaudes (diminution nette au delà de la partie haute de la thermocline). C'est une espéce typique des eaux guinéennes. Elle est moins abondante au Congo qu'à l'intérieur du golfe de Guinée.
Laqocephalus laeviqatus (Tetraodontidae). C'est le représentant le plus abondant de cette famille. Il a été pèché 70 fois de 15 à 100 m. Assez commun de 15 à 50 m, la prise maximale a été de 15 kg en une heure sur les fonds de 40 m. Il est aussi classé dans la communauté des Sparidés par Longhurst (1969), ce qui dénote un caractére eurytherme.
Sphyraena spp. (Sphyraenidae). Les brochets ou barracudas sont représentés de 15 à 100 m dans les prises de la radiale. Le pourcentage d'occurrence est supérieur à 50 % de 15 à 60 m. Des prises horaires maximales de 82 et 95 kg ont été effectuées à 40 et 50 m. Avec 2,5 % des prises l'espèce est classée au dixième rang des abondances sur l'ensemble de la radiale. Ces prédateurs sont également capturés en quantité non négligeable par les sardiniers. L'espéce principale est Sphyraena dubia. La famille est également classée dans la communauté des Sparidés par Longhurst.
Galeoides decadactylus (Polynemidae). Les “petits capitaines” représentent 2,7 % des prises totales et sont présents sur les fonds de 15 à 30 m de la radiale Grand Bassam avec une abondance moyenne relativement uniforme. Ils décroissent ensuite assez rapidement (Figure 4) et quelques spécimens ont encore pu ètre pèchés à 60 m.
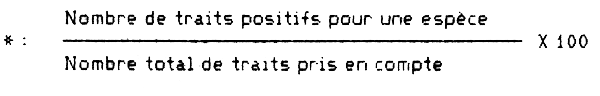
Les prises ont été effectuées pour des températures variant de 16 à 30°C et des salinités de 33,8 à 35,8 °/oo, principalement dans les eaux chaudes et la partie haute des eaux de la thermocline. D'après les résultats obtenus au Ghana, Rijavec (1971) indique que l'espèce préfèrerait les fonds mous, mais les diverses radiales effectuées en Coête d'Ivoire (Tableau 5) montrent également de bonnes abondances sur fonds sableux et sablo-vaseux, les vases putrides seraient peut-bèvitées (Grand Lahou). La taille maximale observée sur les radiales en 1966–67 fut de 46 cm de longueur totale (Jacqueville). Lors des campagnes CHALCI de 1978–1980, le plus grand individu mesurait 38 cm de longueur à la fourche. La répartition moyenne annuelle des tailles sur la radiale de Grand Bassam en 1966–67, pour trois bandes bathymétriques (15–20 m, 25–35 m, 40–60 m), a été représentée sur la figure 5. Il n'y a guère de différence dans les histogrammes, cependant, à la lecture du tableau 6, il apparait que les individus les plus gros se tiennent plutòt à la còte et que les plus petits sont proportionnellement plus fréquents en profondeur. Des individus de petite taille, jamais en grand nombre, sont réguliérement capturés en lagune Ebrié.
Pentanemus quinquarius (Polynemidae). La “friture à moustache”, ou “barbillon”, est une espèce très còtière de petite taille et de faible valeur marchande qui a été rencontrée de 15 à 40 m (Figure 4), avec un pourcentage d'occurrence supérieur à 50 % (moyenne 89 %) de 15 à 25 m. La prise maximale d'un trait a été de 32 kg sur les fonds de 20 m, qui présentent également la plus forte prise moyenne annuelle. La distribution en fonction de la température et de la salinité est similaire à celle de Galeoides decadactylus (de 18 à 30°C et de 33,9 à 35,8 °/oo), il a été péché en quantités notables de 23 à 28°C. Les répartitions observées au Bénin, Cameroun, Congo sont proches, mais les maxima sont obtenus sur des fonds un peu plus faibles (15 m). Cette différence proviendrait sans doute de ce que l'espèce éviterait les fonds purement sableux, comme le suggeère Durand (1967) à partir des résultats obtenus par Crosnier et Berrit (1966) au Bénin-Togo où des fonds sableux font suite à des fonds sablo-vaseux à partir de 15–17 m : l'espèce n'a pas été trouvée au delà de cette profondeur, alors que des prises plus profondes ont eu lieu sur les fonds à fraction vaseuse du Cameroun et du Congo. Cette supposition paraft confirmée par les résultats obtenus en còte d'Ivoire ou des fonds sableux cotiers plus ou moins larges précèdent les sables vaseux. Les radiales où les sables montrent la plus grande extension (Jacqueville et Fresco) ne présentent que très peu de Pentanemus Quinquarius (Tableau 7). Notons aussi que sur la radiale de Pointe- Noire, ou les fonds cotiers sont plus vaseux qu'en Còte d'Ivoire, les rendements sont nettement supérieurs (55 kg en moyenne à 15 m en saison chaude).
Pomadasys jubelini (Pomadasyidae). Cette espèce, surnommée “carp blanche”, est régulièrement présente sur les fonds còtiers de la radiale de Grand Bassam ainsi que Pomadasys peroteti, une autre carpe blanche, dont l'abondance a été 17 fois plus faible. Pomadasys jubelini, a été capturé de 15 à 40 m, son pourcentage d'occurrence a été supérieur à 75 % de 15 à 30 m et nettement inférieur à 50 % ensuite. Cette espèce a représenté 0,6 % des prises sur la radiale (15–100 m). Elle a été capturée pour des températures allant de 18 à 30°C et pour des salinités de 33,9 à 35,7 °/00, principalement dans les eaux chaudes et la partie haute de la thermocline. L'espèce évite très nettement les eaux subtropicales. Le plus grand individu pèché lors des radiales de 1966–67 mesurait 55 cm de longueur totale ; 57 cm de longueur à la fourche lors des campagnes CHALCI de 1978–80.
Nous avons représenté dans le tableau 8 les prises moyennes horaires 10–40 m pour les cinq radiales ivoiriennes. Pour les quatre radiales situées à l'ouest d'Abidjan ces moyennes doivent ètre considées avec précaution du fait du faible nombre de traits de chalut et du caractère assez grégaire de l'espèce qui peut entrainer une grande variabilité dans les prises. Ce caractère grégaire se manifeste dans des captures parfois importantes des sardiniers ivoiriens. Au Sénégal, Alberdi (1971) signale qu'ils peuvent, en saison chaude, “se rassembler sur le fond en bancs de grandes dimensions qui, en raison de leur extension verticale, sont accessibles aux filets tournants” (loc.cit.p.9). Les prises du tableau 8 sont importantes lors des radiales Sassandra et Grand Lahou (l'ajout des Pomadasys peroteti aux autres radiales ne modifierait guère les chiffres) qui sont situées près du débouché de deux grands gleuves.
Ceci paraft en rapport avec l'importance de Pomadasys jubelini dans la faune estuarienne et lagunaire, notee par Gras (1961) au Bénin, Longhurst (1963) en Sierra Leone. Cette espèce est pèchee trèe régulièrement dans la lagune Ebrie près d'Abidjan, ou les tailles ne sont jamais elevées par rapport à celles pouvant étre observées en mer.
Brachydeuterus auritus (Pomadasyidae). Surnommée “–friture” ou “friture à ecailles” en Còte d'Ivoire, c'est l'espèce la plus abondante sur le plateau continental avec 17,6 % des prises sur la radiale de Grand Bassam (15–100m). Elle a un caractère semi—pélagique marqué et constitue environ 15 % des mises à terre des sardiniers ivoiriens. L'agrégation fréquente des individus en bancs plus ou moins denses entraine une importante variabilité des prises. Ainsi une prise horaire de 1120 kg a été réalisée sur les fonds de 30 m lors de la radiale Grand Bassam 19, représentant 93 % de la prise totale. L'espece a été rencontrée de 15 à 80 m à Grand Bassam (Figure 4) avec une abondance maximale située sur les fonds de 30 à 50 m, ce qui est très comparable aux résultats obtenus au Congo. Le pourcentage d'occurrence est supérieur à 60 % de 15 à 60 m. Brachydeuterus auritus a été capturé sur la radiale de Grand Bassam pour des températures de 16 à 30°C et des salinités de 33,8 °/00; elle évite apparemment les eaux subtropicales et peut-ëtre également les eaux supérieures à 27°C.
Les tailles maximales relevées en Còte d'Ivoire ont été, en 1966–67, de 25 cm LT sur la radiale de Grand Bassam, et deux individus d'une longueur exceptionnelle de 33 cm LT ont été observés sur les fonds de 18 m à Fresco ; en 1978–80 le maximum a été de 24 cm LF (correspondant à 26 cm LT) lors des campagnes CHALCI. Les jeunes individus se tiennent plutòt en profondeur comme nous le montrons sur la figure 6 qui représente les distributions de taille obtenues sur la radiale de Grand Bassam pour les fonds 15 à 25 m, 30 à 40 m et 50 à 80 m (*): dans les deux premieres distributions 12% et 17 % de la population mesurent moins de 13 cm, et dans la derniére, près de 36% . D'apres Crosnier (1966), la répartition bathymétrique des tailles paraît varier avec la saison, les gros individus présenteraient des déplacements en relation probable avec la reproduction.
Crosnier (1966) et Durand (1967) indiquent que l'espèce semble avoir une certaine préférence pour les fonds de vase ou de vase-sableuse. Nous avons utilisé pour le vérifier les résultats des campagnes CHALCI 78.01 et 79.01 (**), à partir desquels nous avons calculé pour chaque campagne (Tableau 9), la prise moyenne de friture suivant les typoes de sédiment pour les fonds de 25 à 65 m (aire de répartition préférentielle). Au vu des résultats, la friture semble préfèrer les fonds vaso-sableux puis sablo-vaseux, mais évite les vases et les sables purs. Nous indiquerons en conclusion quelles sont, à notre avis, les explications à cette répartition préférentielle, qui peut etre notée de manière plus ou moins nette pour plusieurs espèces de la communauté des Sciaenidés. Remarquons, à propos de la variabilité des prises dont nous avons parlé plus haut, que les estimations de l'abondance moyenne entre les deux campagnes présentent une différence importante (51,8 kg/0,5 h contre 84,6 kg/0,5 h).
Pseudotolithus typus (Sciaenidae). Ce “bar” est commun sur les petits fonds de la radiale de Grand Bassam avec des pourcentages d'occurrence nettement supérieurs à 50 % de 15 à 25 m. Les prises ont é effectuées pour des températures de 18 à 30°C et des salinités de 33,9 à 35,8 °/oo ; l'espèce n'est relativement abondante qu'à partir de 23°C. La taille maximale enregistrée en Côte d'Ivoire a été de 107 cm lors de la campagne CHALCI d'aoôt-septembre 1980. Le tableau 10 représente l'abondance moyenne observée en 1966–67 sur les différentes radiales. Les résultats sont en accord avec les observations faites par Crosnier (1964). Longhurst (1966) et Durand (1967), qui indiquent que l'espèce est abondante dans les eaux dessalées côtières (estuaires et leur voisinage) et qu'elle éviterait les fonds sableux.
Pseudotolithus seneqalensis (Sciaenidae). Cette “ombrine” est l'espèce commercialement la plus importante de Côte d'Ivoire, quoique sixième en ordre d'importance sur la radiale de Grand Bassam avec 5,6 % des prises. Sa distribution va de 15 à 50 m sur cette radiale (Figure 4) où son pourcentage d'occurrence est de 100 % de 15 à 25 m. Les fonds de 25–30 m présentent l'abondance maximale. La prise horaire la plus importante de la radiale a été de 139 kg (fonds de 25 m) et une prise de 202 kg a été effectuée à Sassandra (fonds de 22 m). Les prises à Grand Bassam ont eu lieu de 17° à 30°C et de 33,8 à 35,8 °/oo ; Troadec (1971) a particulièrement bien démontré que l'espèce évitait, dans la mesure du possible, les eaux froides. Pseudotolithus senegalensis n'a jamais été capturé à plus de 50 m de profondeur en Côte d'Ivoire ainsi qu'au Nigéria (Longhurst, 1965), alors qu'on l'a rencontré jusqu'à 70 m au Congo.
L'évitement par l'espèce des fonds durs et sableux a été noté par la plupart des auteurs qui ont étudiés sa répartition. Longhurst (1966) est celui qui le montre le mineux en comparant les pourcentages d'occurrence de ce sciaenidé au dessus de six types de substrats (Tableau 11) d'après les résultats des campagnes du Guinean Trawling Survey qui ont eu lieu en 1963–64 entre le Sud du Sénégal et le Congo. La lecture du tableau 11 montrerait qu'en plus des fonds durs, Pseudotolithus seneqalensis éviterait probablement aussi les vases putrides.
Comme indiqué par Troadec (1971) en Côte d'Ivoire et Fontana (1979) au Congo, les jeunes individus sont plutôt côtiers, ce qui ressort du traitement des mensurations des radiales Grand Bassam selon trois bandes bathymétriques (Figure 7). Les gros individus surviendraient, eux, de manière apparemment uniforme (Tableau 12). Le plus grand individu mesuré en Côte d'Ivoire a atteint 63 cm (campagne CHALCI de mars 1979 - fonds de 20 m), pour 57 cm au Congo.
Pteroscion peli (Sciaenidae). La “friture blanche” est une espèce abondantetroisième en importance avec 7,7 % des prises sur le plateau continental au niveau de la radiale de Grand Bassam - qui est assez généralement rejetée par les chalutiers. Elle a été observée de 15 à 60 m (Figure 4) sur la radiale avec un pourcentage d'occurrence égal ou proche de 100 % de 15 à 30 m. L'abondance maximale se situe à 25 m où la meilleure prise horaire a été enregistrée (211 kg). Pteroscion peli a été capturé à Grand Bassam de 16 à 30°C et de 34,5 à 35,8 °/oo de salinité, principalement dans les eaux chaudes et celles du niveau supérieur de la thermocline. Les prises horaires moyennes sur les différentes radiales (Tableau 13) semblent montrer que l'espèce éviterait les fonds sableux (Jacqueville 25 et 30 m, Fresco 30 m, Sassandra 22 m) ; cela pourrait expliquer la différence de répartition observée entre la radiale de Grand Bassam et la radiale de Pointe-Noire, l'abondance apparente pour cette dernière étant la plus élevée sur les fonds de 15 m.
Vomer setapinnis (Carangidae). Le “plat-plat Mussolini” a été observé de 15 à 80 m sur la radiale de Grand Bassam, avec des pourcentages d'occurrence supérieurs à 50 % de 15 à 40 m. L'espèce a représenté 1,6 % des prises du plateau au niveau de la radiale et la prise maximale observée pour une heure a été de 64 kg à 20 m le 10 novembre 1966. Les prises ont eu lieu de 16 à 30°C et de 34,5 à 35,8 °/oo de salinité ; comme pour l'espèce précédente, elle se trouve surtout dans les eaux chaudes et celles du niveau supérieur de la thermocline. Sa répartition est similaire au Congo (où il a pu cependant être observé à 100 m) et dans les autres régions du golfe de Guinée.
Une autre espèce de “plat-plat” est plus côtière (15 à 35 m) et moins abondante (maximum 28 kg à 30 m le 25 février 1967), il s'agit de Chloroscombrus chrysurus, appelé “médaille” quand il est de petite taille. Les deux espèces se rencontrent dans la partie de type estuarien de la lagune Ebrié.
3. COMMUNAUTE DES SPARIDES
Mustelus mustelus (Triakidae). C'est le plus abondant des requins. Il a été pêché 36 fois, de 25 à 200 m, sur la radiale de Grand Bassam. Son abondance maximale semble être sur les fonds de 35 à 60 m. Au Congo ses captures sur la radiale de Pointe-Noire ont eu lieu de 15 à 200 m avec une dominante à 70–100 m. On reprendra pour ce sélacien sa classification antérieure par Durand (1967) dans les espèces eurybathes.
Squatina oculata (Squatinidae). “L'ange de mer” a été récolté 23 fois de 60 à 200 m.
Raja miraletus (Rajidae). Cette espèce, surnommée “raie zéro” à cause de ses ocelles, a été rencontrée de 15 à 100 m lors des radiales Grand Bassam, avec une abondance maximale sur les fonds de 30 à 60 m (Figure 12). Capturée entre 15 et 29°C de température et 34,5–35,8 °/oo de salinité, elle préfère très nettement les eaux de la thermocline et les eaux subtropicales supérieures à 17°C, les plus fortes abondances se situent entre cette valeur et 24°C. L'examen de sa distribution nous entraine à modifier le dernier classement de Longhurst (1969) et à la classer dans la communauté des espèces eurybathes ou de la thermocline. Les distributions observées au Bénin et au Congo sont tout à fait similaires.
Fistularia villosa (Fistulariidae). Ce “poisson-flûte” a été pêché à Grand Bassam, de 30 à 100 m, et toujours en petite quantité.
Epinephelus aeneus (Serranidae). Ce “mérou”, très apprécié des consommateurs, est assez fréquent en Côte d'Ivoire. Il a été capturé de 20 à 80 m sur la radiale de Grand Bassam, avec des pourcentages d'occurrence supérieurs à 50 % de 30 à 50 m. Les pêches ont eu lieu pour des tempérieurs variant de 15 à 25°C et des salinités comprises entre 34,7 et 35,8 °/oo ; l'espèce évite très nettement les eaux chaudes. Sa répartition sur la radiale de Grand Bassam n'est pas très représentative de l'ensemble du plateau continentahivoirien et nous avons utilisé les résultats des trois campagnes d'échantillonnage stratifié de 1978–1980. Si le pourcentage d'occurrence est bien le plus élevé dans la strate 30–49 m (Tableau 14), la prise moyenne est similaire pour les fonds supérieurs à 80 m, et nettement plus forte dans la strate 50–80 m.
Les gros individus, non capturés pendant la radiale Grand Bassam, se trouvent sur les fonds de 50 à 100 m et plus particulièrement sur ceux de 50 à 80 m. Ils ont été généralement pris sur des fonds assez durs ou proches d'affleurements rocheux. La répartition bathymétrique d'Epinephelus aeneus sur la radiale de Pointe-Noire au Congo est très proche de celle des campagnes CHALCI,qui est également en accord avec les indications fournies par Rijavec (1971) pour le Ghana. Cet auteur note également la proximité préférentielle de fonds durs non chalutables lors des bonnes prises.
Priacanthus arenatus (Priacanthidae). Cette espéce appeleée “motard” à cause de la grosseur de ses yeux a été trouvée de 25 à 200 m. Elle occupe le quinzième rang sur le plateau continental au niveau de la radiale de Grand Bassam avec 13 % des captures. Son aire de répartition préférentielle va de 50 à 100 m, avec des pourcentages d'occurrence compris entre 66 et 94 %. Présente dans des eaux de 15 à 25°C et de 34,8 à 35,8°/oo de salinité (aucune prise n'a été effectuée dans les eaux guinéennes et tropicales), l'espèce se tient dans la partie basse de la thermocline et dans les eaux subtropicales. La capture maximale observée a été de 44 kg (fonds de 100 m).
Pentheroscion m'bizi (Sciaenidae). Il ressemble au Sciaenidae côtier Pteroscion peli en forme et en taille. Sa présence - notée à partir de 50 m - est importante de 60 à 200 m (Figure 8) avec un pourcentage d'occurrence très proche ou égal à 100 %. Sa répartition en fonction de la température et de la salinité (de 15 à 22°C et de 35,5 à 35,8 °/oo) est similaire à celle de l'espèce examinée précédemment, avec une préférence encore plus marquée, pour les eaux froides. Durand (1967) note que la température optimale en saison froide parait etre 16–17°C, 83 % des captures en cette saison ayant été effectuées entre ces deux températures. La prise maximale en une heure sur la radiale Grand Bassam fut de 216 kg sur fonds de 100 m. Elle représente 6,9 % des prises sur cette radiale (15–100 m), mais elle est rejetée en grande majorité par les chalutiers. La taille du plus grand individu mesuré est de 33 cm, le maximum le plus courant étant 27–29 cm. Il y a une nette répartition bathymétrique des tailles (Figure 9) ; les individus les plus petits sont sur les fonds de moins de 100 m et les plus gros sont plus profonds. La répartition de l'espèce est comparable dans d'autres régions du golfe de Guinée et au Congo. Il semble, d'après Durand (1967), que l'espèce évite les fonds durs, ce qui est confirmé par Rijavec (1971) au Ghana.
Trachurus trecae (Carangidae). Ce “chinchard” peut parfois se montrer assez abondant de 50 à 100 m. Il a été capturé sur la radiale Grand Bassam de 15 à 26°C et de 35,0 à 35,8 °/oo de salinité ; il se trouve dans toutes les eaux de la thermocline, mais son abondance est de beaucoup la plus forte dans les eaux subtropicales. La prise maximum fut de 89 kg à 80 m. Sa distribution est similaire au Congo.
Pseudupeneus prayensis (Mullidae). Le “rouget-barbet” a été capturé de 15 à 80 m sur la radiale de Grand Bassam avec un pourcentage d'occurrence supérieur à 50 % de 30 à 60 m. Les prises ont eu lieu pour des températures de 16 à 26°C et des salinités de 34,7 à 35,8 ° /oo ; l'espèce se trouve surtout dans les eaux de la thermocline et les eaux subtropicales supérieures à 17°C. Il nous semble qu'elle serait vraisemblablement mieux classée parmi les espèces de la thermocline, ou dans l'élément côtier de la communauté des Sparidés, plutôt que dans l'élément profond de cette dernière (Longhurst, 1969).
La repartition paraft assez similaire dans les autres région du golfe de Guinee et au Congo. L'espèce représente 3,3 % des prises entre 18 et 73 m au Ghana (Rijavec, 1971). Le rouget-barbet fait l'objet d'une importante pèche au Sénégal sur les fonds sableux et durs ; ils sont trouvés sur la còte à 75 m de profondeur avec un maximum de 30 à 50 m (Garcia et al., 1978). Cette préférence pour les fonds à fraction sableuse importante se manifeste en Còte d'Ivoire dans les résultats des campagnes CHALCI de 1978–1980 : 14 prises supérieures à 10 kg (pour 0.5 heure de trait) ont été enregistrées pour les 94 chalutages effectués de 20 à 80 m sur des fonds dont la teneur an particules inférieures à 50 μ n'excède pas 50 %, contre aucune pour les 66 traits sur fonds plus vaseux.
Dentex anqolensis (Sparidae). Présent de 40 à 200 m, le “pagect rouge” est particulièrement abondant à 80–100 m (Figure 8). Il occupe le quatrième rang sur le plateau continental au niveau de la radiale de Grand Bassam, avec 7.2 % des prises pondérées 15–100 m. Capturée de 15 à 24°C et de 35,2 à 35,8 ° /oo de salinité sur la radiale, l'espece est parfois présente dans les eaux de la thermocline mais se trouve surtout dans les eaux subtropicales. La distribution est similaire au Congo et les tailles maximales enregistrées sur la radiale de Pointe-Noire et celle de Grand Bassam sont identiques (33 cm de longueur totale). Comme noté par Crosnier (1966), Durand (1967) et Konan (1977), les petits individus sont dans la frange còtière de la distribution et les grands plutot dans la frange profonde. Cela est nettement visible après traitement des mensurations effectuées lors des radiales Grand Bassam de 1966–67 (Figure 10). D'après les resultats des campagnes CHALCI de 1978–1980 il ne semble pas y avoir de variations de l'abondance suivant le type de fond, sauf peut-être une diminution sur les fonds durs, mais le nombre de chalutages supérieurs à 50 m sur ces derniers fonds est insuffisant pour se prononcer valablement.
Dentex congoensis (Sparidae). Cette espèce, qui ressemble beaucoup à la précédente, n'a été signalée qu'une fois (à 200 m) au cours des radiales Grand Bassam de 1966–67. Nous l'avons cependant rencontrée assez fréquemment lors des campagnes CHALCI sur toute la longueur du plateau continental ivoirien, y compris au niveau de la radiale Grand Bassam, et l'on peut se demander si un problème de détermination spécifique n'a pu se poser en 1966–67. Dentex conqoensis est très rare au Congo mais commun dans les autres régions du golfe de Guinée.
Au Ghana cette espèce est plus abondante que Dentex angolensis (Rijavec, 1971) et cela paraît également être le cas au Nigéria et au Cameroun. Dentex congoensis a une distribution un peu plus profonde que Dentex angolensis survient pas avant 70 m lors de CHALCI 78.01 alors que Dentex angolensis apparaît dès 45 m. Les profondeurs d'apparition respectives sont de 50 et 40 m lors des campagnes de mars 1979 et d'août-septembre 1980. Dentex congoensis est un peu plus petit que Dentex angolensis (max. 24 cm LF contre 29 cm). Crosnier (1966) indique que l'augmentation des tailles avec la profondeur est moins marquée que pour Dentex angolensis.
Dentex canariensis et Pagrus ehrenbergi (Sparidae). Ces deux esèces sont présentes, en très faibles quantités et occurrences, de 15 à 50 m sur la radiale de Grand Bassam. Elles ont été un peu plus communes lors des campagnes CHALCI de 1978–1980, où des prises ont pu ètre effectuées sur les fonds de 15–80 m et un individu de la première espèce exceptionnellement pèché à 120 m.
Ces deux especes sont beaucoup plus abondantes sur les fonds durs du Ghana, où Rijavec (1971) indique qu'elles ont respectivement représenté 6,5 et 5,1 % des prises 18–73 m sur l'ensemble des radiales. Rijavec (1973) observe une abondance maximale à 30 brasses (55m) et des prises moyennes considérablement plus basses sur les fonds mous que sur les fonds durs, il note également une augmentation de la taille moyenne avec la profondeur. Dentex canariensis et Pagrus ehrenbergi sont deux espèces caractéristiques de l'élément côtier de la communauté des Sparidés, dont les composants peuvent survenir sur les fonds durs situés au dessus de la thermocline.
Pagellus coupei (Sparidae). Le “pageot blanc” a été rencontré de 15 à 200 m sur la radiale de Grand Bassam avec une abondance maximale à 50 m (Figure 8), cependant l'abondance moyenne peut ètre considérée comme nulle à 200 m et au dessus de 30 m. Capturée de 15 à 26°C et de 35,0 à 35,8° /oo de salinité, l'espèce fréquente les eaux de la thermocline et le haut des eaux subtropicales, elle est abondante de 17 à 24°C. La distribution est similaire au Togo-Bénin ainsi qu'au Ghana et est assez proche au Congo, Cameroun, Nigéria. Comparée à celles des campagnes CHALCI de 1978–1980, l'abondance nous paralt faible sur la radiale Grand Bassam où Pagellus coupei représente 1,2 % des prises. De mème la prise maximale horaire observée, 60 kg à 50 m, est faible comparée à 1978–1980 où des prises supérieures à 100 kg par demi heure de trait n'ont pas été exceptionnelles (max. = 248 kg). Une explication à ces faibles prises sur la radiale de Grand Bassam pourrait provenir d'une préférence de Pagellus coupei pour les fonds à importante fraction sableuse, préférence qui est signalée par Crosnier (1966) et Rijavec (1973). Ce dernier auteur observe au Ghana des rendements moyens 3,6 fois plus èlevès sur fonds durs que sur fonds mous. Ceux obtenus en Côte d'Ivoire pendant les campagnes CHALCI de 1978–1980, pour l'aire de répartition préférentielle 40–70 m, sont en accord avec une telle préférence (Tableau 15) ; nous noterons cependant que quelques forts rendements ont été obtenus sur des fonds à dominante vaseuse. Cette prèdilection pour les substrats durs, qui nous semble plus nettement visible pour les petits fonds, ainsi que la répartition bathymétrique d'abondance de l'espèce, nous permettent de dire que Pagellus coupei est particulièrement proche des espèces classées par Longhurst (1969) comme appartenant à l'élément côtier de la communauté des Sparidés.
La taille maximale observée en Côte d'Ivoire en 1966–67 a été de 37 cm de longueur totale (plus communément 32–33 cm), pour 30 cm au Ghana. Crosnier (1966) écrit que la répartition bathymétrique en fonction de la taille n'est pas très nette, les jeunes paraissant cependant plus abondants au large que dans la frange côtière de la répartition. Les conclusions de Rijavec (1973), qui se base sur un nombre important de poissons échantillonnés (20071), ne sont pas très claires ; les plus gros poissons éviteraient les eaux côtières (moins de 20 brasses, soit environ 37 m). En Côte d'Ivoire les distributions moyennes annuelles en fonction de la profondeur sont assez similaires (Figure 11) ; compte tenu du nombre relativement faible de poissons échantillonnés, il semblerait que les plus gros individus seraient plutôt dans la partie profonde de la distribution. Une étude récente et détaillée effectuée au Sénégal sur les nurseries de Pagellus coupei (Franqueville, 1980) montre une concentration en saison froide des juvéniles sur les petits fonds de 5–30 m, alors que les adultes se trouvent de 15 à 65 m. Cet auteur indique également que les juvéniles auraient une préférence pour les zones d'affleurements rocheux et de sables grossiers, et que l'emplacement des nurseries correspond aux zones d'upwellings les plus intenses.
Uranoscopus albesca (Uranoscopidae). Cette espèce d'assez petite taille est actuellement rejetée par les chalutiers ivoiriens, alors qu'elle est souvent conservée par certains chalutiers étrangers operant au large de l'Afrique. Sa répartition sur la radiale de Grand Bassam va de 60 à 200 m, avec un pourcentage d'occurrence supérieur à 80% de 80à 100 m, contre moins de 15% à 60 et 200 m. Pendant les campagnes CHALCI de 1978–1980, elle n'a jamais été rencontrée à moins de 45 m. Les prises horaires sur la radiale de Grand Bassam furent souvent de l'ordre de 5 à 13 kg. La répartition en Côte d'Ivoire est identique à celle qui à été observée au Congo. Crosnier au Bénin signale avoir pêché Uranoscopus albesca à partir de 12 m.
Brotula barbata (Brotulidae). La “brotule” a été capturée à Grand Bassam de 35 à 200 m, avec un maximum à 80–100 m (Figure 8). Le pourcentage d'occurrence est supérieur à 75 % de 50 à 200 m et la plus forte prise a été effectuée a 100 m (139 Kg). L'espèce a été pêchee de 15 à 24°C et de 35,2 à 35,8°/00 de salinite ; elle n'est abondante que dans les eaux subtropicales. En Cote d'Ivoire elle représentait 3,6 % des prises de la radiale Grand Bassam (15–100 m). La répartition bathymétrique est identique au Congo et apparemment aussi au Togo-Bénin, mais l'abondance est nettement plus faible dans ces régions.
Balistes carolinensis (Balistidae). Cette appellation doit etre preférée à celle de Balistes capriscus qui a souvent été utilisée en Afrique de l'Ouest. L'espèce s'est avérée rare en 1966–67 ; elle a été pêche 11 fois de 20 à 50 m, pour un poids total de 3,1 kg, sur la radiale de Grand Bassam. Son abondance s'est considérablement accrue au début des années 1970 et sa répartition bathymétrique en Côte d'Ivoire va de 15 à 100 m, avec le maximum entre 30 et 60 m,d'après Caverivière (1979). Nous renvoyons à cet article pour plus de détails.
Paracubiceps ledanoisi (Stromateidae). Le “sardineau” apparait dans les prises de la radiale Grand Bassam de 50 à 200 m, et en quantities non negligeables de 80 à 200 m, avec un pic à100 m (Figure 8). L'espèce, capturée de 15 à 22°C et pour des salinités supérieures à à 35,5 ° /oo. se trouve quasi exclusivement dans les eaux subtropicales (plus fortes abondances en dessous de 19°C). Avec 2,0% des prises 15–100 m, c'est la onzième espèce par ordre d'importance du plateau continental ivoirien au niveau de la radiale de Grand Bassam. La prise horaire maximale a été de 121 kg à 100 m en avril 66. La taille maximale enregistrée pour les 9430 mensurations effectuées en 1966–67 est de 21 cm de longueur totale.
Lepidotrigla cadmani, Lepidotrigla carolae, Trigla lineata, Trigla gabonensis (Triglidae). Plusieurs espèces de “grondins” - qui ne sont pas commercialisées en Côte d'Ivoire - ont pu etre pêchées entre 35 et 200 m lors de la radiale Grand Bassam. Les prises horaires n'ont généralement pas dépassé 4 kg et la principale espèce est Lepidotrigla cadmani, qui a son abondance maximale à 100 m. Lors des campagnes CHALCI de 1978–1980 les prises (ramenées à une heure de chalutage) se sont sovent montrées plus importantes qu'en 1966–67 à Grand Bassam. Nous avons également note, lors de ces campagnes, une répartition bathymétrique différente entre Lepidotriqla cadmani et Triqla spp., les espèces de ce dernier genre (presque exclusivement Triqla gabonensis) étant plus côtieres (Tableau 16). La taille maximale enregistrée en 1979 pour Lepidotrigla cadmani est de 21 cm (L.F.) pour 744 individus mesurés, elle est de 26 cm (L.F.) pour le genre Trigla (193 mensurations).
4. ESPECES EURYBATHES OU DE LA THERMOCLINE
Scoliodon terranovae (Carcharinidae). Ce requin a été rencontré quelques fois de 15 à 35 m. Durand le signale de 15 à 70 m au Congo.
Torpedo torpedo (Torpedinidae). Cette “torpille” a été pêche 35 fois de 20 à 100 m - en quantitiés toujours faibles - sur la radiale de Grand Bassam. Son pourcentage d'occurrence et les prises moyennes sont maxima de 35 à 60 m, en accord avec les données de Crosnier au Bénin. Durand et Crosnier l'ont également trouvée en quantité relativement abondante sur les petits fonds du Cameroun et de la radiale de Pointe-Noire.L'occurrence relative est plus élevée au Congo.
Raja miraletus (Rajidae). Nous avons classé cette espèce - considérée comme appartenant à la communauté des Sparidés par Longhurst - dans ce groupement (cf.Figure 12 et communauté des Sparidés).
Trichiurus lepturus (Trichiuridae). La “ceinture” a été capturée de 15 à 200 m lors des radiales Grand Bassam de 1966–67 ; son pourcentage d'occurrence est supérieur à 50 % de 15 à 100 m. Sur ces fonds elle représente 6,9 % des prises. L'abondance moyenne (Figure 12) montre un pic bien marqué à 25 m (58 kg/h). Elle est de l'ordre de, ou supérieure à, 10 kg/h de 10 à 80 m avec une exception à 50 m. La forte variabilité des prises nous incite à ne pas attacher trop d'importance au “creux” observé à 50 m, la distribution bathymétrique moyenne calculée devant être considérée comme assez grossièrement indicative de l'abondance moyenne reelle. Cette espèce semi-pelagique est egàlement capturée par les sardiniers qui la rejettent parfois, comme d'ailleurs - mais dans une moindre mesure - les chalutiers. Les prises sur la radiale de Grand Bassam ont eu lieu pour des températures et des salinités comprises entre 15 et 30°C et 34,0 à 35,8°/oo. La répartition est relativement homogène dans ces eaux, l'espèce éviterait cependant les températures inférieures à 17°C et peut-etre celles supérieures à 28°C. La distribution est assez similaire au Congo - où la “ceinture” paraìt moins abondante - ainsi qu'au Bénin, Nigéria, Cameroun. Elle est présente en lagune Ebrié, près du débouché sur la mer.
Cynoglossus canariensis (Cynoglossidae). Cette sole “langue de chien” représente 90 % des pêches de soles en Côte d'Ivoire (Chauvet, 1970). Elle a été récoltée de 25 a 100 m sur la radiale de Grand Bassam (Figure 12) avec des quantités notables et des pourcentages d'occurrence supérieurs à 75% de 30 à 60 m,sans pic très marqué dans cet intervalle (max.à 40 m). Les captures ont eu lieu de 15 à 29°C et de 34,5 à 35,8° /oo de salinité ; l'espeèce se trouve principalement dans les eaux de la thermocline et la partie haute des eaux subtropicales. Elle représente 1,5 % des prises (15–100 m) de la radiale. La prise maximale a été de 37 kg à 40 m et la plus grande taille enregistrée égalait 54 cm (taille maximale au Congo = 56 cm). Les répartitions bathymétriques observées au Togo-Bénin, Cameroun, Congo, Sénégal (Thiam, 1978)sont similaires.
La préférence de l'espèce pour des substrats contenant une fraction importante de vase a été notée par Crosnier (1966) au Togo-Bénin et Chauvet (1970) en Côte d'Ivoire. Thiam (1978) trouve au Sénégal que Cynoglossus canariensis a une abondance decroissante suivant les fonds sablo-vaseux,vaso-sableux, vaseux et sableux, avec cependant un pourcentage d'occurrence le plus eleve pour les vases et vases sableuses. Les chalutages des campagnes CHALCI de 1973–1980 dans l'aire préférentielle de répartition de l'espèce (30–60 m) paraissent indiquer une prédilection (Tableau 17) pour les vases-sableuses en saison chaude (CHALCI 78.01 et 79.01), qui n'est pas confirmée par la campagne de saison froide (CHALCI 80.01) ; l'évitement des fonds sableux semble réel malgré le faible nombre de traits realises sur ce substrat.
Il existe une repartition differentielle des tailles suivant la profondeur (Figure 13) ; les gros individus se trouvent plutôt sur les petits fonds et c'est l'inverse pour ceux de moins de 30 cm. Cette répartition va assez à l'inverse de ce que dit Chauvet (1970), qui ne s'est basé que sur la sortie effectuée devant Grand Lahou le 1.4.67. D'après Durand au Congo, les plus petits et les plus gros individus se trouvent aux profondeurs moyennes de l'aire de répartition.
5. DISCUSSION ET CONCLUSIONS
Après avoir observé la répartition bathymetrique moyenne des principales espèces, nous pouvons examiner celle des prises totales toutes espèces (Figure 14). Les rendements maximaux s'observent de 20 à 30 m et le pic de 25 m est bien marqué (plus de 500 kg/h). A partir de 35 m les rendements sont assez stables autour de 200 kg/h, il fault cependant noter une baisse à 40–50 m correspondant à la base de la thermocline et à la séparation entre les deux communautés principales. En effet, la limite la plus basse des espèces de la communauté des Sciaenidés correspond, d'après Longhurst (1966), à la plus basse limite de la thermocline. Ainsi cet auteur indique que dans la baie de Biafra, où la thermocline est moins profonde que dans le reste du golfe de Guinée, la séparation de faune est également plus côtière.
Plus tard, Troadec (1971) a montré que la limitation vers le bas de l'ombrine Pseudotolithus senegalensis serait plutôt due à un facteur trophique. Il se base sur :
les travaux de Le Loeuff et Intès (1968,1969) qui montrent un changement qualitativement et quantitativement important dans la faune benthique ivoirienne au dessous de 50 m, correspondant à un changement de peuplement ; ces auteurs distinguent de plus dans le peuplement littoral une faune côtière séparée à partir de 25–30 m d'une faune subcôtière ;
l'observation que les ombrines ne dépassent pas 50 m de profondeur et diminuent fortement d'abondance à partir de 30 m, même quand les conditions physico-chimiques et la nature du fond sont favorables.
Nous ajouterons que Le Loeuff et Intès notent que : “dans la majorité des cas, la distribution bathymétrique des organismes n'est pas modifiée par le changement des conditions hydrologiques sur les fonds où ils vivent, même pour les espèces les plus vagiles”.
A partir des observations précédentes, on pourra généraliser à l'ensemble des espèces de la communaute des Sciaenides l'action préponderante de facteurs trophiques dans la répartition ers le bas de ces espéces. La correspondance entre la limite de la communauté des Sciaenidés et la limite vers le bas de la thermocline n'est, dans l'ensemble, valable que dans la mesure où la profondeur moyenne de cette derniére déterminerait la limite de la faune littorale benthique. Le facteur trophique est prépondérant dans la distribution des espèces demersales quand les conditions physico-chimiques ne sont pas trop défavorables.
Nous avons vu que la plupart des espèces de la communauté des Sciaenidés pour lesquelles nous disposons de donnees sur ce sujet eviteraient les fonds trop sableux (Pentanemus guinguarius Brachydeuterus auritus, Pseudotolithus typus et senegalensis, Pteroscion peli), ou trop vaseux (Galeoides decadactylus, Brachydeuterus auritus, Pseudotolithus senegalensis). A partir des résultats des traits de chalut effectués lors des campagnes CHALCI de 1978–1980, nous pouvons observer une diminution de l'abondance globale des especes sur les fonds de sables et de vases compris entre 10 et 50 m (Tableau 18). Cette diminution parait également apparente si l'on enlève des prises totales celles des principales espèces semi-pélagiques. Une telle baisse globale de l'abondance des espèces démersales côtières sur les fonds de vases et sables purs est à mettre en relation avec une diminution de la biomasse benthique sur ces fonds (Longhurst, 1959). Nous ne disposons pas de suffisamment de données, au delà de 50 m, pour les différents types de fonds. Il semblerait cependant que les différences - si tant est qu'elles existent réellement -y soient moins marquées.
D'après l'étude des distributions bathymétriques en fonction de la taille des principales communautes, il semble que l'on puisse dégager une indication générale concernant un schéma de répartition différent entre les espèces des divers groupements. Les espèces de la communauté des Sparidés pour lesquelles nous disposons de données suffisantes ont en moyenne annuelle une proportion de gros individus plus importante vers le bas de leur aire de répartition que vers le haut (Epinephelus aeneus, Pentheroscion m'bizi, Dentex angolensis, Pagellus coupei) ; cela ne serait pas le cas, et même serait souvent l'inverse, pour les espèces de la communauté des Sciaenidés ou eurybathes (Galeoides decadactylus, Cynoqlossus canariensis), Brachydeuterus auritus.
BIBLIOGRAPHIE
Alberdi, P.G., 1971 - Biologie et pêche du “sompat” Pomadasys jubelini (Cuvier 1830) des eaux sénégambiennes (aperçu préliminaire). Centre Rech. Océanogr. Dakar-Thiaroye, Doc.Scient.Prov. n° 30 : 38p.
Baudin-Laurencin, F., 1967 - La sélectivité des chaluts et les variations nycthémérales des rendements dans la région de Pointe-Noire. Cah. ORSTOM, sér. Océanogr., vol. V,n° 1: 83–121.
Blache, J., Cadenat, J.et Stauch, A., 1970 - Clés de détermination des poissons de mer signalés dans l'Atlantique tropical (entre le 20° parallèle N. et le 15° parallèle S.). Faune Tropicale XVIII, ORSTOM Paris. 480p.
Bouillon, P., Troadec, J.P. et Barro, M. - Pêches au chalut sur les radiales de Jacqueville, Grand-Lahou, Fresco et Sassandra (mars 1966-février 1967). Centre Rech. Océanogr.Abidjan, Doc.Scient.Prov.,n°36 : 13p + 46pl.
Caverivière,A., 1976 - Mensurations effectuées devant la Côte d'Ivoire lors des radiales de chalutage de Grand Bassam, Jacqueville, Grand-Lahou, Fresco et Sassandra (1966–1967). Centre Rech. Océanogr. Abidjan, Arch. Scient., vol. 1, n° 3 : 124p.
Caverivière,A., 1980 - Campagne CHALCI 80.01 (20.8.80–10.9.80). Résultats des chalutages. Centre Rech. Océanogr. Abidjan, Arch.Scient., vol. 6, n° 3 : 198p.
Caverivière, A. et Champagnat, C., 1978 - Campagne CHALCI 78.01 (30.1.78 au 12.2.78). Résultats des chalutages. Centre Rech. Ocèanogr.Abidjan, Arch. Scient., vol.4, n° 1 : 118p.
Caverivière, A. et Champagnat, C., 1979 - Campagne CHALCI 79.01 (13.3.79 au 31.3.79). Résultats des chalutages. Centre Rech. Océanogr. Abidjan, Arch. Scient., vol.5,n°4 : 198p.
Caverivière, A., Konan, J. et Bouberi, D., 1982 - Etude des variations nycthémérales d'abondance des poissons de chalut en Côte d'Ivoire.Doc. Scient., Centre Rech.Océanogr.Abidjan, vol.XIII, n° 2 : 1–22.
Chauvet, C., 1970 - Etude de Cynoglossus canariensis (STEIN) du plateau continental de la Côte d'Ivoire : lecture d'age, étude de la croissance en longueur et en poids - observations sur leur biologie et leur dynamique. Thèse 3°cycle, Fac.Scien.Aix-Marseille, 102p.
Crosnier, A., 1964 - Fonds de pêche le long des côtes de la République Fédérale du Cameroun.Cah.ORSTOM,sér.Océanogr., n° spécial : 133p.
Crosnier, A. et Berrit, G.R., 1966 - Fonds de pêche le long des côtes des Républiques du Dahomey et du Togo.Cah.ORSTOM, sér.Océanogr., suppl.4 : 144p.
Durand, J.R., 1967 - Etude des poissons benthiques du plateau continental congolais. Troisième partie : étude de la répartition, de l'abondance et des variations saisonnières. Cah.ORSTOM,sér. Océanogr., vol. V,n° 2 : 3–68.
Eckman, S., 1953 - Zoogeography of the sea. Sidgwik and Jackson, London.
Fontana, A., 1979 - Etude du stock démersal côtier congolais. Biologie et dynamique des principales espèces exploitées. Propositions d'aménagement de la pêcherie. Thèse Doct. Etat Sciences Nat., Université Pierre et Marie Curie-Paris VI : 300p.
Franqueville, C., 1980 - Etude des nurseries de daurades Pagellus coupei le long de la côte ouest-africaine de 10° à 20°N. Centre Rech. Océanogr. Dakar-Thiaroye, Doc. Scient., n° 76 : 15p.
Garcia, S., Lhomme, F., Chabanne, J. et Franqueville, C., 1978 - La pêche démersale au Sénégal : historique et potentiel. FAO, COPACE/PACE SERIES/78/8 : 59–77.
Gras, R., 1961 - Liste des poissons du Bas Dahomey faisant partie de la collection du laboratoire d'hydrologie du Service des Eaux, Forêts et Chasses du Dahomey.Bull. IFAN, sér. A, T.23, n° 2 : 572–586.
Konan, J., 1977 - Biologie et dynamique de Dentex angolensis (Poll et Maul, 1953) du plateau continental ivoirien. Thèse Doct. 3° cycle Océanogr. biol., Université Pierre et Marie Curie, Paris, 87p.
Le Loeuff, P. et Intès, A., 1968 - La faune benthique du plateau continental de Côte d'Ivoire. Récoltes au chalut - Abondance - Répartition - Variations saisonnières. Centre Rech. Océanogr. Abidjan, Doc. Scient. Prov., n° 25 : 78p. + 30p. tab.
Le Loeuff, P. et Intès, A., 1969 - Premières observations sur la faune benthique du plateau continental de Côte d'Ivoire. Cah. ORSTOM, sér. Océanogr., vol. VII, n° 4: 61–66.
Longhurst, A.R., 1959 - Benthos densities off tropical West Africa. J. Cons. Intern.Explor.Mer, vol.25, n° 1 : 21–28.
Longhurst, A.R., 1963 - The bionomics of the fishery resources of the eastern tropical Atlantic.Fish.Publ.London, n° 20 : 65p.
Longhurst, A.R., 1965 - A survey of the Fish Resources of the Eastern Gulf of Guinea.J.Cons.Intern.Explor.Mer, vol.29, n° 3 : 302–334.
Longhurst, A.R., 1965 - The biology of West African Polynemid Fishes. J.Cons. Intern.Explor.Mer, vol.30, n° 1 : 58–74.
Longhurst, A.R., 1966 - Synopsis of biological data on West African croakers (Pseudotolithus typus, P. senegalensis and P. elongatus). FAO, Fish. Synopsis, n° 35 : 50p.
Longhurst, A.R., 1969 - Species assemblages in the tropical demersal fisheries. UNESCO. Actes du symp. sur l'océanogr. et les rèss. halieut. de l'Atl. Trop., Rapp. de synthèse et Comm., Abidjan 1966, pp. 147–170.
Martin, L., 1973 - Morphologie, sédimentologie et paléogéographie au quaternaire récent du plateau continental ivoirien. Thèse Doct. Etat Scien. Nat., Univarsité Paris IV, juin 1973, 340p. + 3 cartes H.T.
Postel, E., 1968 - Hydrologie et biogéographie marines dans l'ouest africain. in West African international Atlas (First instalment). OUA/IFAN, Dakar, 1968, pp. 13–17.
Rijavec, L., 1971 - A survey of the demersal fish resources of Ghana. Rapp. FAO/PNUD, 40p. multigr. - Aussi FAO, 1980, CECAF/TECH/80/25.
Rijavec, L., 1973 - Biology and dynamics of Pagellus coupei (Dieuz. 1960), Pagrus ehrenbergi (Val. 1830) et Dentex canariensis (Poll. 1954) in Ghana waters. Centre Rech.Océanogr.Abidjan, Doc.Scient., vol. 4, n° 3 :43–97.
Thiam, M., 1978 - Ecologie et dynamique des Cynoglosses du plateau continental sénégambien. Biologie de Cynoglossus canariensis (Steind. 1882). Thèse Doct. 3° cycle, Université Bretagne Occidentale, 180p.
Troadec. J.P., 1971 - Biologie et dynamique d'un Sciaenidae ouest-africain, Pseudotolithus senegalensis (V.). Centre Rech. Océanogr. Abidjan, Doc. Scient., vol. II, n° 3 : 225p.
Troadec, J.P., Barro, M. et Bouillon,. P., 1969 - Pêche au chalut sur la radiale de Grand-Bassam (Côte d'Ivoire) (mars 1966-février 1967). Centre Rech. Océanogr. Abidjan, Doc.Scient.Prov., n° 33 : 14p. + 11 tab. + 89 pl.
![]()
![]()
![]()