![]()
![]()
![]()
LE REGIME ALIMENTAIRE DES PRINCIPALES ESPECES DEMERSALES DE
CÔTE D'IVOIRE (ET DU GOLFE DE GUINEE)
par
CAVERIVIERE Alain
Chargé Recherches de l'ORSTOM
Les régimes alimentaires des espèces démersales n'ont guèe été etudies en Cote d'Ivoire. Citons 21 espèces examiinées par le Loeuff et Intes (1973) mais le travail a porte sur peu d'individus récoltés à une seule époque de l'année d'ailleurs non précisés. Seykes quelques recherches ont eu lieu sur un nombre relativement élevé d'individus : il s'agit des études de Troadec (1968,1971) sur Pseudotolithus senegalensis et Pseudotolithus typus, de Barro (1976) sur Brachydeuterus auritus, de Intes et Le Loeuff (1976) sur Dentex angolensis. Nous pourrons compléter et comparer les résultats obtenus en Côte dI'voire avec ceux Concernant d'autres régions de la côte ouest africaine : étude rares elle aussi, parmi lesquelles il faut citer en particulier les travaux de Longhurst (1957, 1960a) qui portent sur 25 espéces de l'estuaire de la Sierra Léone et 71 espéces récoltées des Iles du Cap Vert àFernado Po.
Avant de décrire les régimes alimentaires des espéces dé mersales pour lesqulles nous disposons de renseignements, il nous semble préféable de parler tout d'abord des peuplemnets d'invertées benthiques qui sont d'une grande importance dans l'alimentation de ces poissons, en partculier les crustacés.
1. LA FAUNE BENTHIQUE
Aprés un résumé des connaissances générales ontenues sur la fune benthique de Côte d'Ivoire, nous envisagerons plus en détail la répartition et l'abondance de deux crustacés le Caridae Palaemon hastatus et le Penaeidae Parapenaeopsis atantica, qui peuvent étre particuliérement abondants dans les nols alimentaires des espéces démersales côtiéres de Côtr d'Ivoire.
1.1 Etude générale
Rijavec (1971) indique au Ghana une bonne corrélation significative à 99 %, entre la prise moyenne de poissons démersaux et la quantité d'invertébrés benthiques ramenée dans le filet, et prise comme indicateur de l'abondance sur le fond (le vide de maille du culde chalut étant de 32 mm).
En Côte d'Ivoire une étude de Le Loeuff et Intés (1968) sur la faune benthique, basée sur le méme type d'échantillonnage, a été effectuée à partir des inverébrés en 1966–67 lors des nombreux traits de chalut (vide d maile du cul de 40 mm) effectués sur les radiales de sassandra, Fresco, Grand Lahou, Jacqueville et surtout GrandBassam . Du fait de l'engin utilisé les prises ne concernent que l'épifaune de grande taille, et neuf traits effectués avec un chalut dont les mailles du cul mesuraient 16 mm étirées ont montré l'inadéquation du filet de 40 mm dans I'échantillonnage des espéces benthiques du point de vue du nombre des espéces récoltées et, plus gravement, de leur abondance. L. corrélation obtenue par Rijavec montre cependant la valeur glaonale de la méthode vis-a-vis de la densité pondérale des especes démersales ; on peut méme peser que la correlation obtenue avec ce type d'échantillonnage doit etre melleure que celle que l'on pourrait obtenir à partir d'engins plus classiques de récdolte du benthos téles ceux utilisés par Longhur st 1958 de la Ganble à la Sierra Leone et Buchanan 1958) au Ghana. Ces engins é talent principalement des bennes et qes dragues qui ne pernmettent pas d,écdhantillonner la faune vagile.
Le Loeuff et Intés montrent les variations, en fonction de la profondeur, du nombre des espéces et de l'abondance totale en individus (Figure 1). Ces variations correspondent á des faunes differents ; le nombre d'espéces et dindividus est:
assez important entre 15 et 30m.
elevé en especes et surtout en individus entre 35 et 50 m.Il est à noter que dans cette zone bathymétrique les récoltes peuvent varier beaucoup d'une sortie à l'autre et d'une profondeur à l'autre.
faible dans les deux cas - mais surtout en individus - à partir de 60 m, notamment à cette profondeur qui correspond à l'extension limite des influences littorales et à charniére entre deux types de faunes.
Ces auteurs indiquent égalemaent une augmentation, dans les deux premiéres aries bathymétriques des captures d.onvertébrés benthiques en salson foide et pendant les upwelling éspiosidques . Cette augmentation n'est pas trés visible sur les garaphes mais est bien réelle, tout au moins pour la premiére zone, si l'on considére que l,abondance de Palaemon hastauts, espéce côtiére de saison froide. est trés mal représentée dans les échantillons obtenus par chalutage. La crevette còtiére de plus grade taille Paraopenaeopsis atantica voit également son abondance augmenter du début de la grande saison froide au début de la petite saison chaude . L'augmentation des captures d'invertébrés benthiques en saison froide paraft une conséquence logique des poussées plantiques induite par les remontees d'eaux froides.
Les variations saisonnéres s'acciompagnent chez certaines espéces de migrationverticales, cependant pour la majorité des espéces la répartition des poplutions n'est pas affectée par les variations des condition hydrologiques méme pour les espéces les plus vaqgiles. Le mode de répartiton des organismes sur le fond doit jouer un role trés important dans la considérable variabilité des échantillons.
Pour la zone qui nous intéresse dans cette étude, de la côte a la rupture de pente du plateau continental, Le Loeuff et Intés regroupent les invertébrés benthiques en deux peuplements d'aprés les relations organismes-conditions hydrologiques.
Le peuplement littoral est situés au dessus de l'isobathe 60m, les organismes qui le constituent ne se rencontrent jamais dans des eaux dont la température est inférieur à 17°C. Cette zone est caractérisée par l'instabilité des conditions hydrologiques. A l'intérieur de ce peuplement une distinction est faite entre la faune côtiére, dont les espéces se nmaintiennent au dessus de la thermocline et supportent de grandes variations de température et de salinité, et la faune subcôtiére dont les espéces se rencontrent à partir de 25-30 m, évitant ainsi les eaux chaaudes et dessalées.
Le peuplement profond àpartir de 60 m, avec la faune de borure du plateaum est séparé du peuplment littoral par une limite bien marquée, il y a une brusque diminution de la richesse en benthos, les especes de transition sont peu nombreuses.
Ces peuplements ont des relations certaines avec les communutés benthiques de Longhurst (1958) et de Buchanan (1958), Leuns limites corresopondent assex bien avec celles des principaux peupleents ou communautés de vertébres démeersaux décrits par Durand (1967) et Longhurst (1969).
L'étude de Le Loeuff et Intés ne porte sur aucun fond dur, ce type de substra n'étant cependant pas rare en cote d'Ivoire du delà de 50-60 m. Au Ghana , ou ils sont majoritaires, les fonds durs (à gogones) sont donnés par Rigavec (1973) comme plus riches en organismes benthiques de grande taille que les fonds mous correspondants, ce qui paraft étre u cas trés général dans les océans. Longhurst (1959) montre que les vases et sables purs sont pauvres en benthos et nous noterons que d'aprés Rijacvec les fonds mous ds radiales effectuées au Ghana sont consitutés de telles vases, Signalons aussi que c'est à 60m, progondeur ou la biomasse bebnthique est la plus faible , que la teneur des élment fins du sédiment atteint son maximum sur la radiale de Grand Bassam.
1.2 Répartition et abondance de Palaemon hastaus et Parapenaeopsis atlantica
1.2.1 Le Caridae Palemon hastatus (Aurivillius)
Cette petite crevette transparente, d'une taille maximale de 7 cm, est une espéce côtiére abondante lors du maximum de productivité planctonique en saison froide . Elle n'a njsque à ce jour, et à notyre connnaissance, pas fait l'objet d'etudes particuliéres en Afrique Occidentale. Elle est présente de juillet à octobre-novembre devant Grand Bassam,. Traoadec (1971) note que des prises attengant hjusqu'à 10 kg par heure de péche ont pu étre obtebnus malgre l'inadéquatuib des naulles du cdhalut à la péche ont pu étre obtenues malgré (1968) l'ont récoltéses sur les fonds de 15 et 20m , aucun renseignement n'étant ét effectuées sur les fonds de 15 et 20m, aucun renseignement n'étent disponible pour les fonds plus côtiers. L'espéce a été éga;ement rencoterées lors des radiales de Sassandra et Grand Lahou, mais sa présence n'apas été mais sa présence n'a pas été constatée lors des radiales de Fresco et Jacqueville, ce qui laisseà penser que sa presence est lie à la proximité d'eaux dessalées. Ce caridé est d'ailleurs aussi connu comme une espéce d;eaux esssalees. Troadec (1971) fait état d'une communication personnelle de Longhurst selon laqulle Palaemon hastatus abonde le long des' côtes nigérianes en période de dessalure de la couche superdficielle. Le Loeuff et Intés indiqunet qu'un certain nombre de femelles étaient ovigéres.
1.2.2 Le Penaeidae Parapenaeopsis atlantica (Balss)
Paraqoenaeopsis atlantica n'a encore été que peu étudiée . Crosnier (1967) lui a consacré une coutre monographie au Conogo, dont les conclusions sont reprises par Crosiner et De Bondy (1967). Cette crevette côtere, qui ne se rencontre jamais en lagune, parait commune sur les fonds vaseux ou vaso-sableux. Les longueurs totales des individus captureés au chalut sont comprise entre 6 et 14 cm.
Son abondance dans le chalut sur la radiale de Grand Bassam n'a jamais été trés forte (max. 20 kg/h). Elle a été pévhée de 15 à 50m, plus particukiérement de 15 à 30 m avec un maximum à 25 m. Elle se rencontre toute l,année mais les variations saisonniéres d'abondance sont importantes. Troadec (1971) donne, pour le poids de l'ensemble des individus et le nombre de Parapenaeopsis atlantica de longueur totale inférieure à 8 cm, les indices d'abondance sur la radiale de Grand Bassam (Figure 2) les meilleures prises ont lieu pendant la petite saison des pluies et la cure des fleuves, en fin de saison froide - début de saison chaude.
1.2.3 Comparaison entre le Côte d Ivoire et le Congo - Essai d'interpretation
Crosnier et De Bondy (1967) pour Parapenaeopsis atlantica et Samba (1974) pour Palaemon nastatus, indiquet qu'au Congo les maxima d'abondance se rencontrent en salsons chaudes, qul correspondent aux crues des fleuves. Nous avons vu qu'en Côte d'Ivoire les abondances maximales pour ces deux especes prenaient place en saison froide et au début de la precipitation à la côte et de décharge correspondent periodes des maxima de prévipitation à la cdote de decharge des fleuves. Les apports d'eaux douces joueraient donc, plutot que la tempèrature, un rôle important sur la variations d'abondance de ces deux crevettes dans les zones de pêche des poissons dèmersaux côtiers.
La dessalure en elle méme ne paraft cependant pas avoir une influence directe sur l'augmentation d'abondance car en Côte d'Ivoire c'est pendant les périodes de crues que, a part une mince couche d'eau dessaléee en surface, la salinite sur le fond a son maximum du fait de la remontée par des phénoménes d'upwelling de l'eau centrale sud atlantique.
Dandonneau (1973) et Binet (1976) ont montré sur le plateau continental ivoirien que les apports de sels nutritifs par les eaux continentales auraient un rôle prépondérant sur l'accroissement des biovolumes planctoniques. Binet (1970) ne signale pas de phénoméne comparable au Congo, les biovolumes de zooplancton montreraient méme une baisse important au large de Pointe-Noire pendant la grande crue du Congo, quand les prises de Parapenaeopsis atlantica sont les plus importantes. L'augmentation d'abondance des deux crustacés ne parait donc pas liée une richesse planctonique simultanée des eaux.
Binet (1970) indique qu'au Congo les larves de décapodes Natantia - essentiellement des larves de Caridae et, moins nombreuses, de Penaeidae(*) - sontabondantes en saison chaude au contraire de la majorite des autres taxons du zooplancton; les maxima se situent pendant les deux crues.
Un phénomène assez analogue (période d'anondance en saison chaude, mais des maxima pendant la grande saison des pluies et la crue des fleuves) a lieu en Côte d'Ivoire pour les larves de décapodes (Binet, 1976a). Cependant la différenciation n'est pas suffisante à l'intérieur de cet ordre pour l'attribuer à Palaemon hastatus et Parapenaeopsis atlantica.
(*) : Qui sont problement pour la plupart des larves de Parapenaeopsis atlantica
De l'ensemble des observations precédentes il semble se dessiner que les augmentations d'abondance de Palaemon hastatus et Parapenaeopsis atlantica sur les fonds de pêche des poissons demersaux côtiers (augmentations apparaissant dans le bol alimentaire de ces denniers) seraient en liaison avec de forts apports d'eaux continentales. sans que les variations de facteurs tels que la température, la salinite, l'abondance planctonique sur ces fonds, paraissent entrer en ligne de compte. Les frotes abondances observees coincideraient avec la période de reproduction de ces espéces, comme l'indiquerait l'observation de femelles ovigéres de Palaemon hastatus en Côte d'Ivoire et l'augmentation du nombre de larves de Caridae et de Penaeidae au Congo. En périodes de reproduction les adultes pourraient avoir une répartition plus profonde ou étre plus vulenérables. Nous ne formulerons pas d'hypotheses sur les mécanismes directs ou indirects qui induiraient la reproduction des deux espéces etudiées pendant les saisons de décharge. Le Reste (1973) et Garcia (1977) ont deja signale l'importance des précepitations et des crues des fleuves sur le déclenchement des pontes de Penaeus indicus á Nosy-Be (Madagascar) et de Penaeus duorarum en Côte d'Ivoire Le premier auteur ecrit que l'on peut supposer que “les variations pluviométriques sont l'horloge de la reproduction”.
2. LES REGIMES ALIMENTATIRES
2.1. Moyens d'etude et presentation des donnees commentees
2.1.1. Moyens d'étude
Le principal moyen est l'analyse des contenus stomacaux. La méthode la plus simple pour ce faire consiste á denombrer (occurence) les estomacs contenant tel ou tel groupe de proies, le pourcentage d'occurrence étant la proportion des estomacs non-vides contenant l'espéce ou la famille considérée. Ce procédé ne tient pas compte des quantités ingerees, mais Jones (1954) a montre une bonne corrélation entre l'augmentation du poucentage d'occurrence et celle du poids de chaque type de nourriture. “En effet, si pour une proie donnee la quantite totale consommée individuellement par un lot de predateurs croît, la possibilite de rencontrer cette proie dans un plus grand nombre d'estomacs augmente” (Troadec, 1971, loc. cit. p.152). Les pourcentages d'occurrence ne sont pas cumulables car dans un méme estomac les divers groupes peuvent étre presents et la somme depasser largement 100%.
Plusieurs réserves peuvent étre faites concernant la précision des études sur les régimes alimentaires et elles sont bien développées par Troadec (1971). La principale provient de ce que la vitesse de digestion varie suivant les proies. de ce fait, les animaux qui ont des piéces anatomiques dures (squelettes des vertébrés, tests calcaires) pourront étre déterminés plus longtemps aprés leur ingestion que dautres, tels que les petites crevettes et les polychétes.
Deux autres indices sont aussi par fois utilises dans l'étude de l'alimentation des poissons: il s'agit de l'indice de repletion et du facteur de condition. Ils peuvent fournir des informations uniquement quantitatives sur les variations saisonniéres du régime alimentaire.
L'indice de ráplátion est souvent exprimé en quarts de volume stomacal (empli au quart, á la moitié, au trois-quart et plein).
Le facteur de condition exprime la relation entre le poids et la longeur du poisson, il traduit son embonpoint. Il est souvent de la forme (poids/longueur au cube). Les variations de ce rapport sont indépendates de la longueur des poissons quand l'exposant de la relation longueur-poids est egal ou tres proche de 3. Dans le cas contraire des biais apparaissent (Freon, 1979) et il taudra, pour les eviter, modifier l'eposant de la longueur od effectuer une correction (Troadec, 1971). Pour que le facteur de condition donne une meilleure mesure de l'embonpoint, il sera bon d'eliminer du poids du poisson celui du contenu stomacal. La valeur moyenne du facteur de condition n'est souvent pas directement représentative de l'abondance de la nouriture á un moment donné. En effet le facteur de condition ne dépend pas que de l'alimentation et, entre autres, pourna traduire les variations de poids des gonades. D'autre part, méme sil est principalement assujetti aux quantites de nourriture ingérées, il sena, encore faible au début d'une période d'alimentation intense et encore fort quelques temps aprés sa fin; c'est le sens de variation du facteur qui devra, plutot étre considére. Notons aussi qu'en période de reproduction une diminution de l'alimentation pourra agin sur le facteur de condition sans que l'abondance des proies soit en cause.
2.1.2 Présentation des données commentées
Dans cet apercu nous ne mentionnerons que les espéces du plateau continental qui ont une certaine importance quantitative ou qui présent un type d'alimentation particulier. Les espéces relativement abondantes dont le rôle de prédateur est bien connu ne seront pas mentionnées : requins, mérous, barracudas. Pour la plupart des autres espéces du golfe de Guinée, nous renvoyons aux auteurs cités en introduction et á Cadenat (1954). Ce dernier a examiné les contenus stomacaux d'individus pêchés au Sénégal et appartenant á 120 espéces démersales et pélagiques.
Nous avons regroupe sous une forme synoptique dans les tableaux 1 á 3, á partir des donées de Le Loeuff et Intés (1973), Longhurst (1960) et de celle d'autres auteurs ayant etudié une espéce particuliére, les informations concernant l'alimentation des principales espéces démersales de la region. Les indications fournies par Cadenat (1954) ne seront utilisees dans le texte que dans le cas ou, pour l'espéce considéree, les renseignements obtenus á partir des aauteurs précedents manqueraient ou ne proteraient que sur un faible nombre d'estomacs; ceci een supposant que l'alimentation de l'espece dans le golfe de Guinee (sensu stricto) ne doit pas etre, en ce qui concerne la répartition entre les grands groupes zoologiques composant le bol alimentaire, tres differente de celle du Sénégal.
Du fait des pourcentages d'occurrence dont le total est namene á une valeur proche de 100 chez Longhurst, les valeurs indiquees par cet auteur et celles tournies par Le Loeuff et Intes ne sont pas directement comparables, aussi utiliserons-nous dans les tableaux i á 3 quatre symboles pour indiquer les niveaux de presence, éleves á faibles, des grands groupes zoologiques. L'ordre de classement de gauche á droite de ces groupes dans les tableaux correspond grossomodo au passage des espéces les plus vagiles á celles qui sont pratiquement fixes.
Dans son travail de 1957, Longhurst regroupe la plupart des especes en trois classes suivant que leur nourriture est principalement composee de:
poissons (ichtyophages);
epifaune active et poissons;
epifaune sedentaire et endofaune.
Nous conservons cette classification, malgre le caractere parfois trop schematique de ces ensembles, en ajoutant une classe pour les especes dont l'alimentation est trés variee.
2.2 Communaute des Scianenidae
Dasyatis margarita ou “raie perlee” (Dasyatidae), D'apres les donnees de Longhurst (1960a), ceette espece a un regime compose essentiellement de petits crustaces, Les polychetes et echiuriens occupent une place importante dans l'alimentation. Contrairement á la raie Raja miraletus, accur poisson ou céphalopode n'a éte trouvé dans les contenus stomacaux.
Ilisha africana ou “rasoir” (Clupeidae). Cette petite espece cotiere semi-pelagique a une nourriture principalement constituee de crustaces et de poissons. Le Loeuff et Intes (1973) trouvent, sur sept estomacs non vides, un regime á base de poissons avec une seule crevette de l'espéce Palaemon hastatus. Longhurst (1960a) indique un regime plutot a base de crustaces pour un plus grand nombre d'individus.
Pentanemus quinquarius ou “petit capitaine” (Polynemidae). Samba (1974) a examine 950 estomacs de cette espece au Congo, ou les conditions du miulieu sont proches de celles de Cote d'Ivoire. Cet auteur indique que l'estomac de Galeoides decadactylus a une paroi trés musculeuse, qui broie les carapaces des crustaces et les coquilles des mollusques, ainsi que des sucs digestits tres puissants. D'aprés les pourcentages d'occurrence, cette espéce a un regime alimentaire varie, mais elle se nourrit surtout de crustaces parmi lesquels les crevettes Palaemon hastatus et Parapenaeopsis atlantica sont les plus representees en nombre et en occurrence. Les poissons constituent egalement un apport important dans l'alimentation de cette espéce. Les proies sont generalement de petite taille par rapport au predateur et, d'aprés Samba, leur taille moyenne evoluerait correlativement á celle des individus etudies. Au large du Congo le nombre d'estomacs vides serait plus important en saison froide qu'en saison chaude; rappelons qu'au contraire de la Côte d'Ivoire, c'est en saison chaude que les crevettes Palaemon hastatus et Parapenaeopsis atlantica sont les plus abondantes.
Le Loeuff et Intés trouvent pour le littoral ivoirien un regime alimentaire assez equivalent, avec des valeurs relatives un peu plus faibles pour les poissons et un peu plus fortes pour les espéces vagiles. Parmi les crustaces Parapenaeopsis atlantic a la meilleure representativite. Cette nourriture variee á base de crustacés est confirmée par Longhrust (1960a).
Pomadasys jubelini ou “carpe blanche” (Pomadasyidae). Nous n'avons que peu de renseignements sur cette espéce. Les travaux de Longhrust (1960a) indiquent un regime alimentaire trés varie s'étendant des polychetes aux poissons. Notons que l'espéce est malacophage en lagune Ebrié, prés d'Abidjan (Albaret, comm.pers.).
Brachydeuterus auritus ou “friture áécailles” (Pomadasyidae). Barro (1976) ecrit a partir de l'examen de 2316 estomacs, dont 1633 contenaient de la nourriture: “Il est remarquable que la polyphagie de ce poisson se limite essentiellement aux elements de la faune vagile composee principalement de zooplancton microscopique, de crevettes, de poissons juveniles, de cephalopodes, de crustacés benthiques et, en cas de besoin, de polychetes”. Les crustacés planctoniques forment la majorite du regime alimentaire de Brachydeuterus auritus, principalement les copépodes et, parmi eux, ceux qui ont les plus grandes tailles, en particulier les calanoides qui sont nombreux au voisinage des fonds. Brachydeuterus auritus se nourriratures press du fond le jour et si oecessaire, dans les couches d'eaux intermediares la need.
D'aprés les variations du facteur de condition. Barro indique que l'alimentation serait importante en saison froide quand l'abondance du zooplancton est élevée. Le régime alimentaire presenterait des variations saisonniéres entre la grande saison froide et les saisons chaudes, la petite saison froide n'ayant pas un régime nettement different des deux saisons chaudes. En ete l'alimentation est plus variee du fait que l'abondance dess crustacés planctoniques ne serait pas suffisante. En petite saison chaude et en petite saison troide les mysidacés predomineraient dans les contenus stomacaux.
Longhurst (1960a) trouve pour cette spece un regime trés varie oú les petits curstacés dominent aussi, mais de facon moins importante; l'abondance des polychétes et des poissons est beaucoup plus grande. On peut penser de ce qui precede, compte tenu egalement de l'importance des estomacss vides (80%), que les individus examines par cet auteur ont été capturés en saison chaude.
Pteroscion peli ou “friture blanche” (Sciaenidae). A partur d'un petit echantillon Le Loeuff et Intes (1973) observent un regime alimentaire base essentiellement sur les petits crustaces epibenthiques ou Palaemon bastatus domine. Pour Longhrust (1960a) la nourriture est un peu plus variee avec la présence de poissons, cephalopodes, mollusques. La encore, comme pour beaucoup d'espéces etudieees par cet auteur, le pourcentage eleve d'estomacs vides (84%) pourrait signifier que la plupart de ses echentillons auraient ete captures en saison chaude.
Pseudotolithus (Fonticulus) elongatus ou “bossu” (Sciaenidae). La representation du tableau 34 nous a été aimablement communiquée par Le Guen (comm.pers.)á partir de ses observations propres et de celles de Poll (in Le Guen. 1971).
Pseudotolithus seneqalensis ou “ombrine” (Sciaenidae). Les differents auteurs ne montrant pas de desaccord important, nous ne parlerons que des travaux de Troadec(1971) qui decrit l'alimentation de cette espéce dans les eaux ivoiriennes et qui est le seul á traiter de ses variations saisonniéres.
Cet auteur a étudie le regime alimentaire de Pseudotolithus senegalensis á partir de 3437 estomacs dont 2814 avaient un contenu. Au total quatre estomacs sur cinq renfermaient des crevettes (un estomac sur deux contenait le caridé Palaemon hastatus et un sur cing le penaeidé Parapenaeopsis atlantica). Il y a donc choix des proies, cette sélection descendant méme au niveau de l'espéce. Troadec montre de maniére trés nette que Palaemon hastatus, espéce de saison froide, est l'aliment préféré des Pseudotolithus senefalensis et que Parapenaeopsis atlantica consitute une nourriture de remplacement. Cela se déduit de la superposition des figures 57 et 58 : d'aprés les graphiques la consommation de Parapenaeopsis atlantica n'est liée ni á leur abondance totale ni á celle des individus de petite taille, leur occurrence augmente dans les estomacs quand celle de Palaemon hastatus diminue du fait de leur abondance dans le milieu en début de petite saison chaude.
En grande saison chaude le régime alimentaire se diversifie nettement du fait des baisses d'abondance sur les fonds de peche de Palaemon hastatus et de Parapenaeopsis atlantica.
Les variations de l'indice de répletion, avec des valeurs faibles en grande saison chaude, sont en accord avec le régime alimentaire. Les baisses de l'indice observées en fin d'upwelling correspondraient au minimum d'oxygéne qui apparait á la méme epoque et qui, selon Troadec, agirait en chassant des fonds la pluparit des espéces de la faune vagile et en particulier la nourriture des ombrines. D'aprés les observations du méme auteur au Congo, les variations du facteur de condition diminution en saison chaude, puis augmentation progressive en saison froide), dépendraient largement de l'alimentation (*).
Les proies consommees par Pseudotolithus senegalensis sont de petite taille, seuls de jeunes individus d'espéces pouvant atteindre des tailles relativement importantes par rapport au predateur ont été notés dans les contenus stomacaux. Le volume individuel des proies augmente avec la longueur des Pseudotolithus senegalensis et, parallelement, la composition de la diéte varie. Toutefois, Palaemon hastatus et Farapenaeopsis atlantica seraient des aliments adéquats pendant une longue periode.
Pseudotolithus typus ou “ombrine sosso” (Sciaenidae). Trodec (1968) indique à partir de 1645 estomacs non vides que l'alimentation de cette espéce cotiere est tres proche de celle de Pseudotolithus senegalensis, avec une occurrence encore plus forte de Palaemon hastatus.
Chloroscombrus chrysurus ou “plat-plat” (Carangidae). Nous ne possedons comme donnees quantifiees pour cette espece semi-pelagique que les résultats des quelques examens effectués par Le Loeuff et Intés (1973). L'alimentation est a base de zooplancton et de micronecton.
Vomer setapinnis ou “plat-plat Mussolini” (Carangidae). La nourriture de cet autre carangide n'a été étudiee par laes auteurs cites précedemment que sur trois estomacs pleins. L'important occurrrence des juveniles de poissons est confirmée par Cadenat (1954) qui défnit cette espéce comme ichthyophage.
Cynoglossus browni et goreensis ou “soles langues” (Cynoglossidae). Le regime alimentaire de ces deux especes tres voisines a été respectivement examiné par Le Loeuff et Intes (1973) et Longhurst (1960a). Les regimes sont du méme type, axes principalement sur l'epifaune sedentaire et sur l'endofaune, avec des pourcentages d'occurrence assez variables d'un groupe zoologique á un autre.
2.3 Communauté des Sparidae
Fistularia villosa ou “poisson trompette” (Fistulariidae). Ces poissons, pourvus d'une petite bouche á l'extrémité d'un long tube, sont de redoutables prédateurs, Le Loeuff et Intés (1973) ont trouvé sept estomacs sur huit “littéralement bourrés” de jeunes poissons. Cadenat (1974) donne une alimentation semblable pur Fistuliaria tabaccaria au Sénégal. Les deux espéces sont actuellement mises en synonymie (Seret, comm. pers.). Nous avons personnellement observé un tel régime, pour de nombreaux individus, lors de campagnes de chalutage effectuées sur le plateau continental ivoirien.
Priacanthus arenatus ou “motard” (Priacanthidae). Les deux estomacs examinés par Le Loeuff et Intès (1973) contenaient des crustaces, des poissons et un cephalopode.
Pentheroscion m'bizi (Sciaenidae). D'apres Longhurst (1960a) cette estpéces serait un prèdateur de jeunes poissons et cephalopodes. Du fait de fort pourcentage d'estomacs vides, ses observations re portent capendant que sur trois bois alimentaires.
Pseudupeneus orayensis ou “rouget” (Mullidae). A partir de 22 individus, Le Loeuff et Intès (1973) indiquent une alimentation très varièe ou les crustacés dominent, suivis par les poissons et les polychetes. Longhurst (1960a) trouve ègalement une nourriture très varièe avec des differences sensibles entre les pourcentages relatives d'occurrence, particulierement pour les mollusques qui sont beaucoup mieux reprèsentès. Le Loeuff et Intès ècrivent á ce sujet qu'il est probable que le règime alimentaire de ce mullidè varie en fonction de la disponibilite oes proies.
Paqrus ehrenbergi ou “dorade” (Sparidae). D'après l etude de Rijavec (1973) ou Ghana, l'alimentation est varièe avec des proportions importantes de poissons, cèphalopodes, crustacès, mollusques, èchinodermes. Le pourcentage de l'èpifaune sèdentaire est plus impotant que pour les deux autres espéces de sparidès (cf. ci-dessous) ègalement ètudièes par cet auteur. L'ètude du pourcentage d'estomacs pleins et du facteur de condition montre que l'espéce 5'alimente plus pendant les saisons froides qu'á la grande saison chaude.
Longhurst (1960a) trouve également un régime varie ou 1'importance des mollusques est remplacée par celle des polychétes.
Dentex canariensis ou “pageot rouge” (Sparidae). Cette espéce, comme la précédente, se nourrit mleux en saison froide. Tout en étant variée, la nourriture se compose principalement d'espéces vagiles : poissons et céphalopodes (Rijavec, 1973).
Longhurst (1960a) sur la vingtaine d'estomacs examinés, n'a pas recontré un seul poisson ni céphalopode; les crevettes dominaient. La faibless de 1'échantillon de Longhurst, ainsi que le relativement faible pourcentage d'estomacs vides, peuvent laisser penser que les individus examinés par cet auteur proviendraient d'un méme échantillon ou auraient été capturés dans une meme saison, période pendant laquelle les crevettes-qui-pourraient étre une prole préférentielle-auraient été abondantes.
Rijavec note pour cette espéce et la précédente une corrélation positive entre leur abondance dans un trait de chalut et la richesse de la faune benthique également récoltée lors de ce trait.
Dentex anqolensis ou “pageot rouge” (Sparidae). Intés et Le Loeuff (1976) ont étuide en détail 1'alimentation de cette espéce en c&ote d'Ivoire . Les crustacés en constitient la base et sont recontrés dans 92% des bols alimentaires, les poissons vienment en seconde position avec prés de 40 % d'occurrence. Ces auteurs déduisent des observations sur les crustacés ingérés que Dentex anqolensis recherche sa nourriture au niveau de la surface du sédiment et qu'il évite de fouir. Les crevettes de 1'épifaune vagile constitueraint des proies appréciées.
Intés et Le Loeuff ne notemt pas de variations significatives des indices de réplétion en fonction des saisons et des profondeurs de prélevement. Cependant, le factur de condition dont les variations ont été suivies par Konan (1977), montre une valeur minimale en fin de grande saison chaude (juin), suivie de valeurs croissantes jusqu en fin de grande saison froide-début de petite saison chaude. Cette augmentation sertant en relation avec la nourriture de la distribution bathymétrique ou les individus seraient plus jenues et la norriture peut-étre plus abondante.
Paqellus coupei ou “Pageat blanc” (Sparidae). Les trois auteurs ayant étudié 1'alimentation de cette espéce dans le golfe de Guinee (Longhrust, 1960a; Rijavec, 1973 : Le Loeuff et Intés ,1973) donnet des régimes pouvant entre assez différents. II est a remarquer que Rijavec recontre dans les bols alimentaires des trois espéces de sparidés qu'ill a etudie des occurrence rewlative de une moindre mesure, de mollusques, plus élevées que celles donn5ees par les deux autres analyses; par contre les polychétes sont toujours en quantités plus faibles. Cela pourrait provenir de ce qu'au Ghana les fonds á sparidés sont plus “durs” qu'en Côte d'Ivoire et qu'au Nigéria.
Paqellus coupei peut donc avoir au total un régime alomentaire trés varié, les occurrences les plus élevées pouvant passer de la faune vagile á 1'endofaune, cependant, les pourventage de crustacés et de poissons sont toujours importants.
Uranoscopus Spp. (Uranoscopidae). Longhurst (1960a) indique pour un membre de cette famille, composée d'escpéces morphologiquement tres proches, une alimentation a base de jeune poissons et,en occurrence moindre, de crevettes. Cadenat (1954) confirme ce régime principalement ichthyophage.
Brotula barbata ou “loche” (Brotulidae). D'aprés les observations de Cadenat, cette espéce se norrirait essentiellement de crustacés et de jeunes poissons.
Triqlidae Spp. ou “grondins”. En ce qui concern 1'espéce Triqla lineate les données de Le Loeuff et Intés (1973), qui ne portent que sur trois individus, “suggérent un régime alimentaire a base de crustacés appartenant pour la plupart á 1'épifaune sédentasire ” . Les jeunes poissons entreraient également dans 1'alimentation. pour Longhurst (1960a) la norriture de Lepidotriqula cadman est variée, á base de poissons et de petits crustacés, il y aurait également occurrence d'annélides . Poissopns et crustacés composeraient donc 1'essentiel de 1'alimentations des grondins.
Céphalacanthus volitans ou “faux grondin” (Dactylopteridae). D'aprés les résultats de Longhurst (1960 a) et de Le Loeuff et Intés (1973) cette espéce a un régime varié avec une importante quantité de crustacés, principalement peu actifs.
2.4 Communauté des espéces eurybathes ou de la thermocline
Raja miraletus ou “raie zéro”(Rajidae). Son alimentation serait assez variée avec dominance des crustacés, principalement des crevettes, et dans une moindre mesure des poissons (Longhurst,1960a).
Trichiurus lepturus ou “ceinture” (Trichiuridae). Cette espéce semi-pélagique est connue un prédateur redoutable , ce que confirement Longhurst(1960a) et Le Loeuff et Intés (1973).
Cynoglossus canariensis ou “sole” (Cynoglossidae). Chauvet (1970), dans l'ètude qu'il a consacrèe á cette espéce au niveau du plateau continental ivoirien, n'a aborde qu'indirectement la question du régime alimentaire, Cet auteur montre que 1'abondance en fonction de la bath/métrie correspond a 1'abondance des peuplements d'invertebrés benthiques littoraux décrits par Le Loeuf et Intes (1968). Le facture de condition et la croissance diminueraient En saison frots.
L'étude des contenus stomacaux a été aboreée par Thiam (1978) au Sénégal, le régime alimentaire serait variá á base de mollusques et de crustacés, du même type que celui des deux especes de cynoglosses dont nous avons deja parle pracemment c'est a dire basé principalement sur 1'epifaune sédentaire et surtount 1'endofaune.
3. CONCLUSION - DISCUSSION
Un examen, meme rapide, des tableaux 1 a 3 montre la grande importance des crustacés dans le régime alimentaire des especes démersales du golfe de Guinée et de la Cote d'Ivoire en particulier, cette importance, déja signalée par plusieurs auterus, est notable pour les trois communautés examinées , En général les petits crustacés et les crevettes dominent. Cependant chez les espéces eurybathes et de la communauté des Sparidae le group des crabes et pagures a souvent un poutcentage d'occurrence élevé. Pour les espéces côtiéres, qui sont économiquement les plus importantes en côte d'Ivoire, l'importance du caridae palaemon hastatus et dans une moindre mesure du Penaeidae Parapenaeopsis antlantica a été souvent mise en évidence. Les juvéniles de poissons sont trés sovent representés en quantité plus ou moins grande , dans le bolm alimentaire des espéces étudiéces . Il semblerait que leur occurrence soit en général plus grande pour les espéces de la communauté des Sparidae que pour celles de la communauté des sciaenidae. Cela pourrait étre une conséquence de la diminution de la faune benthique en dessous dela la thermocline.
Dans un peuplement donnè, tous les types d'alimentation peuvent être rencontrès avec une dominance des prèdateurs actifs. D'apres Longhurst (1957) le pourcentage d'estomacs vides diminuerait en passant des ichthyophages aux espéces se nourrissant principalement á partir de l'endofaune, la valeur calorique des poisson ingèrès ètant forte par rapport aux autres groupes de proies.
D'apres 1'ensemble des reseignements recueillis 1'alimentation de toutes les espéces démersales présenterait des variations saisonniéres en relation avec 1'hydroclimat, ce dernier induirait des variations d'abondance des groupes de la faune benthique composant 1'essentiel des proies. En Cote d'Ivoire les saisons froides serient les périodes favorabl;es pour la grande majorité des epéces ; il en serait de meme au Congo pour Pseudotolithus senegalensis et Pseudotolithus typus (Trodec, 1971; poinsard et Troadec, 1966). Deux exceptions sont actuellement connues:
Cynoqlossus canariensis marque un re;entissement de croissance et unm amaigrissem,ent en saison froide en Cote d'Ivoire (il en est également anisi au Sénégal). Cette espéce se nourrit principalement aux dépens de 1'endofaune qui ne semble guére marquer de variations saisonniéres d'abondance , la basisse de la température pourrait agir sur le métabolisme sans qu'il y ait compensations par une augmentation de la nourriture disponible.
Galeoides decadactylus ay Congo pcésenterait une alimentation et une croissance plus importantes en saison chaude qu'en saison foride. Cela pourrait provenir du fait que dans cette région 1'abondance de certaines proies apparemment liées aux apports d'eaux continentales (Palaemon hastatus, Parapenaeopsis atlantica) serait plus grande en saison chaude. Notons a ce sujet que dans la meme région le facterur de condition de Pseudotolithus elonqatus a ses maxima a 1'époque des crues, et que celui de Pseudotolithus senequalensis nous paraft presenter un decalage plus important que celui observé au Sénégal par rapport au saisons marines chaudes et froides, avec valeurs en augmentation pendant les grandes crues. En Cote d'Ivoire ou les deux phénomenes hydrologiques-saison froide et apponts d'eau douces-se superposent il sera interessant d'étudier les variations du régime alimentaite et de la croissance de Galeoides decadact/lus dans ce conteute.
Les proies sont en général de petite taille parrapport a celle du rédateur d'une maniére le leur taille monenne augmenterait pour une espéce donnée en founction de la longuer de 1'individu étuide importance des groupes zoologiques entrant dans le régime alimentaitre peut varner de ce fait. Le choix des proies, qui parait la régle , a blen mis en evidence par Troadec (1971) pour 1'espéce Pseudotolithus seneqalensis. Cet auteur a pu écrire : “La limite bathymtrique inferieure de ce peuplement (60m) sur lequel se nourrissent les Pseudotolithus senequlensispourrait expliquer que ces poisson ne gagnent pas périodoquement des profondeurs plus grandes lorsque les conditions hydrologiques correspondent a celles dans lesquelles on les recontre courammernt a des niveaux plus eleves”. La diminution de certaines proies entrainerait une variété plus grande du régime alimentaire quand la nourriture préférentielle se raréfie, cette diversification est , toutefois, souvent impuissante a éviter un amaigrissement saisonnier . L'adaptation, souvent certaine limite, des espéces démersales a la nourriture disponible expliquerait les quelques divergences parfois relevées entre différentes études en ce qui concerne le régime alimentaire d'une espéce; elles sont cependant toujours en accord sur ses grandes lignes.
Un bon exemple de cette faculté d'adaptation peut etre vu dans differences relevees par Longhurst (1957,1960a) entre les contenus stomacaux de poissons de memes espéces captures , d'une part dans 1'estuaire de la Sierra Leone River , et d'autre part en mer. Dans le premier millieu le premier milieu le nombre d'estomacs vides est plus faible que dans le second , ou le benthos serait moins abondant (Longhurst, 1959), et 1'occurrence relative des polychétes , specialement abondants dans la faune estuarienne,est en général plus élevée.
Ii y a peu de renseignements sur les variations nycthémérales de 1'alimentation des espéces démersales du golfe de Guinée. Longhurst (1960a) fournit des indications pour Pagrus ehrenberqi qui doivent pouvoir etre étendues a de nombreuses autres espéces. Un pic d'activité aurait lieu a 1'aube et au crépuscule. Les céphalopodes seraient ingérés principalement de jour ainsi que les organismes sédentaires; le pourcentage d'occurrence des invertébrés mobiles ne montre pas de tendance marquée sur 24 heures, la varibilité est grande.
Notons enfin, d'aprés mème auteur, que la chaline alimentaire condusiant aux poissons dèmersaux du plateau continental du golfe de Guinèe serait relativement longue.
BIBLIOGRAPHIE
BARRO, M., 1976 : Biologie et dynamique de Brachydeuterus auritus en Côte d'Ivoire. These d'Universitè Biol. animale. Universitè de Paris VI : 200 p.
BINET,D., 1970 : Apercu sur les variations saisonniéres du zooplacton et plus particuliérement des copépodes du plateau continental de pointe-Noire (congo). Centre Rech. Océanogr. pointe-Noire, Doc. Scient. (Nouv.Sér.),(8):109p.
BINET,D., 1976a : Contribution á 1'é cologie de quelques taxons du zooplancton de Cote d'Ivoire . III - Larves de décapodes et Lucifer. Centre Rech. Océnogr. Abidjan, Doc.Scient .,vol,n1:63–84.
BINET,D.,1976b:Biovolumes et poides secs zooplanctoniques en relation avec le milieu pélagique au dessus du plateau ivoirien. Cah. ORSTOM,sér. Océanogor . vol.XIV,n° 4 : 301–326.
BUCHANAN,J.B.,1958 : The bottom fauna community accross the continental shelf off Accra , Ghana (Gold Coast). Proc.Zool.Soc.Lond., 130(1) : 1–56.
CADENAT,J.,1954 : Notes d'ichthyiologie ouest—africaine - VII, Biologie, régime alimentaire. Bull.Inst.Afr.Noire,(A),16 (2) : 568–83.
CHAUVET,C.,1970 : Etude de Cynoqlossus canariensis (STEIN) du plateau continental de la Cote d'Ivoire : lecture d'age, étude de la croissance en longuer et en poinds -observations sur leur biologie et leur dynamique. Thése 3 cycle,Fac.Scien.Aix-Marseille,102p.
CROSINER,A.,1967 : Quelques données sur la biologie de la crevette penaeide Parapenaeopsis atlantica Balss dans la région de point Noire. Centre Rech.Océanogr. poinet-Noire, Doc,Ronéo n ° 376 S.R.:30p.+7pl.h.t.
CROSNIER, A. et DE BONDY,E.,1967 : Les crevettes commercialisables de la côte ouest de 1'Afrique inter—tropicale. ORSTOM, Paris, Doc.Techn.n° 7: 77p.
DANDONNEAU,Y.,1973 : Etude du phytoplanction sur le plateau continental de Côte d'Ivoire -III- Factures dynamiques et variations spatiotemporelles. Cah.ORSTOM,sér. Océanogr., vol. XI, n°4 : 431–545.
DURAND,J.R., 1967 : Etude des poissons benethiques du plateau continental congolais. Troisiéme parite : Etude de la répartition, de l'abondance et des variations saisonniéers. Cah.ORSTOM , sér Océanogr., vol.V, n°2 :3–68.
FREON, P.,1979 : Relation taill—poids , facteurs de condition et indices de maturitésexuellz : rappels bibliographiques, intetrprétations, remarques et applications. Centre Rech.Océnogr. Dakar-Thiatoye,Doc. Scient.,n°68 :144–171.
GARCIA,S., 1977 : Biologie et dynamique des populations de crevettes roses (Penaeus duorrum notialis peres) Farfante,1967) en Cote d'Ivoire. ORSTOM,Parise,Trav.et Doc.,n°79 : 274p.
INTES,a et LE LOEUFF,P.,1976 : Le régime alimentaire de Dentex anqolensis (poll et Maul 1953) (Sparidae). Centre Rech.Océanoger. Abidjan , Doc. Scient., vol.VII,n°1 : 91–99.
JONES,R., 1954 : The food of the whitiong and a compasrison wuith that of the haddock . Mar.Res.Scotl.Home Deot., n°2 : 1–34.
KONAN, J., 1977 : Biologie et dynamique de Dentex anqolensis (Poll et Maul 1953) du plateau continental ivoirien. Thèse Doct. 3° cycle Océanogr. bio., Université Pierre et Marie Curie, Paris, 87p.
LE GUEN, J.C.,1971 : Dynamique des populations de Pseudotolithus (Foncticulus) elonqatus (Bowd. 1825). Poissons-Sciaenidae. Cah. ORSTOM, vol. IX, n°1 : 3–84.
LE LOEUFF, P. eet INTES, A., 1968 : La faune benthique du plateau continental de Côte d'oire. Récoltes au chalut - Abondance - Répartition -Variations saissonniéres. Centre Rech. Océanogr.Abidjan, Doc. Scient. Prov., n°25 : 78p.+30p.tab.
Le LOEUFF, P. et INTES, A., 1973 : Note sur le régime alimentaire de quelques poissons démersaux de Côte d'Ivoire. Centre Rech.Océanogr.Abidjan, Doc.Scient., vol.Iv, n°2 : 17–44.
Le RESTE, L. 1973 : Etude de la répartition spatio-temporelle des larves et des jeunes postlarves de la crevette penaeus indicusH. Milne Edwards en baie d'Ambaro (Côte nord-ouest de Madagascar). Contribution á l'étude d'une baie eutrophiqu tropicale.Cah.ORSTOM sér. Océanogr.vol.XI n° 2 : 179–189
LONGHURST A.R. 1957 : The food of the demersal fish of West African estuary.J. Anim.Ecol., n°26 : 369–387.
LONGHURST A.R. 1958 : An ecological survey of the West African marine benthos. Colonial office, Fish. publ. n°1 : 101p.
LONGHURST, A.R., 1959 : Benthos densities off tropical West African. J. Cons. Intern-Explor. Mer, vol.25, n°1: 21–28.8
LONGHURST, A.R., 1960 : a SUMMARY SWRVEY OF THE FOOD OF wEST aFRICAN DEMERSAL FISH. bULL. iFAN, SÉR.a, t.xxii, n°1: 276–282.
LONGHUST, A.R., 1969 : Species assmblages in the tropical demersal fisheries. UNESCO. Actes du symp. sur l'océanogr.et les ress.halieut.de l'Atl. Trop., Rapp de synthése et Comm., Abidjan 1966, pp. 147–170.
POINSARD, F.et TROADEC, J.P., 1966: Détermination de l'áge par la lecture de otolithes chez deux espéces de sciaenidés Ouest-Africains (pseudotolithus senegalensis) C.V. et Pseudotolithus typus Blkr.). J. Cons. Intern.Explor.Mer., vol. 30, n°3 :291–307.
RIJAVEC,, L., 1971 : A survey of the demersal fish resources of Ghana. Rapp. FAO/PUND, 40p. multigr. -Aussi FAO, 1980, CECAF/TECH/80/25.
RAJAVEC, L., 1973 : Biology and dynamics of Pagellus coupei (Dieuz. 1960). Pagrus ehrenbergi (Val. 1830) and Dentex canariensis (Poll. 1954) in Ghana waters. Centre Rech. Océanogr. Abidjan, Doc. Sci., vol. 4, n°3 :43–97.
SAMBA, G., 1974 : Contribution à l'etude de la biologie et de dynamique d'un Polynemidae ouest-africain, Galeoides decadactylus (Bloch 1793). Thèse 3° cycle Univers. Bordeaux I'n° d'ordre 1141 : 122p.
THIAM, M., 1978 : Ecologie et dynamique des Cynoglosses du plateau continental sénégambien. Biologie de Cynoglossus canariensis (Steind. 1882). These Doct. 3° cycle, univers. Bretagne Occidentale, 180p.
TROADEC, J.P., 1968 : Le régime alimenaire de deux espèces de Sciaenidae ouest-africaines (Pseudotolithus senegalensis V. et Pseudotolithus typus Blkr.). Centre Rech. Océanogr. Abidjan, Doc. Scient. Prov., n°30 : 24p.
TROADEC, J.P., 1971 : Biologie et dynamique d'un sciaenidae ouest-africain, Pseudotolithus senegalensis (V.). Centr. Océanogr. Abidjan, Doc. Scient., vol. II. n°3 : 225 p.
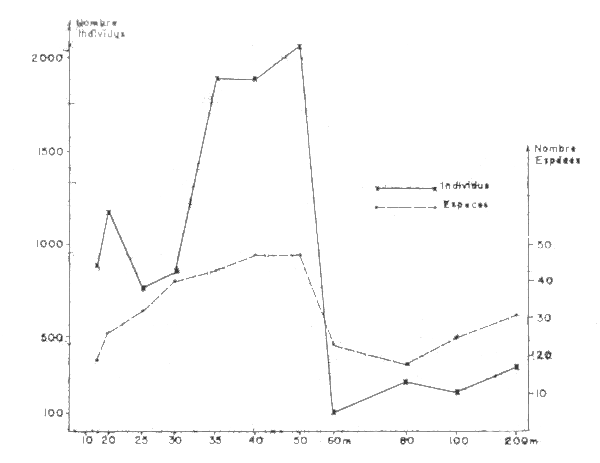
Fig. 1 Niveau de richesse en benthos suivant la profondeur sur la radiale de Grand Bassam (d'après Le Loeuff et Intès, 1968)
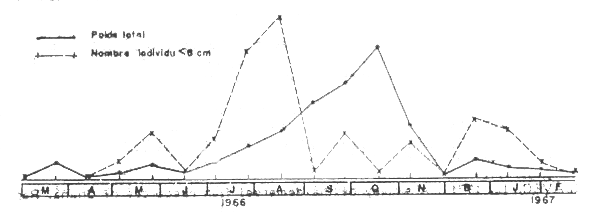
Fig. 2 Indices d'abondance de Parapenaeopsis atlantica sur la radiale de Grand Bassam (Troadec, 1971)
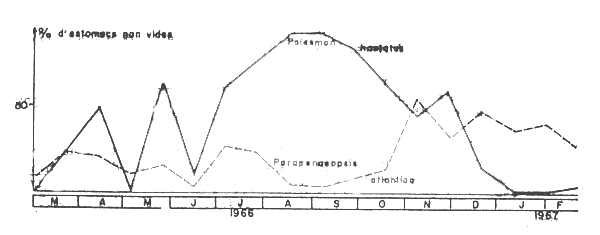
Fig 3 Abondance de Palaemon hastatus de Parapenaeopsis atlantica dans les estomacs de Pseudotolithus senegalensis (Troadec, 1971)
| ESPECE | ESTOMACS | POISSONS | CEPHALOPODES | PETITS CRUSTACES | CREVETTES | CRABES PAGURESS | STOMATOPODES THALASSINIDES | ENSEMBLE CRUSTACES | MOLLUSQUES | OPHIURES | POLYCHETES ECHIURIENS | CATEGORIE | AUTEURS | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| N | N' | |||||||||||||
| Dasyatis maragarita | 150 | 74 | · | D | D | O | · | O | ■ | 2–3 | Longhurst (1960a) | |||
| Ilisha africana | 8 | 7 | · | D | D | 2 | Le Loeuff et Intès (1973) | |||||||
| 284 | 54 | D | · | D | · | 2 | Longhurst (1960a) | |||||||
| Pentanemus | 23 | 23 | ■ | · | · | D | · | O | 2 | Le Loeuff et Intès (1973) | ||||
| quinquarius | 154 | 39 | D | · | · | O | · | 2 | Longhurse (1960a) | |||||
| Galeoides decadactylus | 950 | 175 | ■ | · | · | · | D | · | D | O | D | 2 | Samba (1974) (Congo) | |
| 20 | 20 | D | · | · | · | D | · | D | O | D | 2 ®3 | Le Loeuff et Intès (1973) | ||
| 630 | 192 | D | · | · | · | O | · | O | O | D | 2Җ3 | Longhurust 1960a) | ||
| Pomadasys | ||||||||||||||
| jubelini | 349 | 70 | ■ | ■ | D | · | ■ | D | · | 4 | Longhurst (1960a) | |||
| Crustaces | ||||||||||||||
| Brachydeuterus | 2316 | 1633 | O | O | · | D | · | O | O | O | plancto- | Barro (1976) | ||
| auritus | niques | |||||||||||||
| 386 | 77 | ■ | · | O | · | D | ■ | 4 | Longhurst (1960a) | |||||
| Pteroscion | 47 | 32 | · | · | O | · | 2 | Le Loeuff et Intès (1973) | ||||||
| Peli | 248 | 39 | D | O | · | O | · | D | 2 | Longhurust (1960a) | ||||
| Pseudotolithus elongatus | ? | ? | D | · | D | O | O | · | O | 2 | Le Guen (1971 et com. pers) | |||
| Pseudotolithus senegalensis | 3437 | 2814 | D | O | ■ | · | O | O | · | O | 2 | Troadec (1968, 1971) | ||
| Pseudotolithus typus | 1864 | 1645 | D | O | D | · | · | O | 2 | Troadec (1968) | ||||
| Chloroscombrus chrysurus | 19 | 18 | · | · | D | · | 2 | Le Loeuff et Intès (1973) | ||||||
| Macroplancton | ||||||||||||||
| Vomer sstapinnis | 5 | 3 | · | ■ | · | · | 2 | Le Loeuff et Intès (1973) | ||||||
| Macroplancton | ||||||||||||||
| Cynoglossus browni | 14 | 14 | · | · | · | ■ | · | · | · | 3 | Le Loeuff et Intès (1973) | |||
| C. goreensis | 151 | 62 | O | D | D | ■ | O | · | 3 | Longhurst (1960a) | ||||
N = Estomacs examinés ; N' = Estomas non vides
1 = Inchthyophages ;
2 = Poissons et épifaune active ;
3 = Epifaune sédentaire et endofaune ;
4 = Très varié
| ESPECE | ESTOMACS | POISSONS | CEPHALOPODES | PETITS | CREVETTES | CRABES PAGURES | STOMATOPODES | ENSEMBLE | MOLLUSQUES | OPHIURES | POLYCHETES | CATEGORIE | AUTEURS | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| N | N | CRUSTACES | PAGURES | THALASSINIDES | CRUSTACES | ECHIURIENS | ||||||||
| Fistularia villosa | 8 | 7 | · | 1 | Le Loeuff et Intès (1973) | |||||||||
| Priacanthus arenatus | 2 | 2 | · | ■ | · | ■ | · | 2 | Le Loeuff et Intès (1973) | |||||
| Pentheroscion m'bizi | 77 | 3 | · | ■ | 1 | Longhurst (1960a) | ||||||||
| Pseudupeneus | 2 | 22 | · | ■ | · | · | ■ | · | O | ■ | 4 | Le Loeuff et Intès (1973) | ||
| Prayensis | 36 | 21 | ■ | · | · | · | ■ | 4 | Longhurust (1960a) | |||||
| Pagrus | 4268 | 134 | · | ■ | · | D | D | D | 2→3 | Rajavec (1973) (Ghana) | ||||
| ehrenbergi | 1148 | 373 | · | D | D | D | ■ | O | · | D | D | ■ | 2→3 | Longhurst (1960a) |
| Dentex | 4202 | 129 | · | ■ | D | D | D | ■ | 2 | Rijavec (1973) (Ghana) | ||||
| canariensis | 33 | 20 | · | · | · | ■ | 2→3 | Longhurust (1960a) | ||||||
| Dentex | 7 | 386 | · | ■ | · | · | D | · | D | D | 2→3 | Le Loeuff et Intès (1976) | ||
| angolensis | ||||||||||||||
| 4754 | 164 | · | ■ | ■ | D | D | D | 2 | Rijavec (1973) (Ghana) | |||||
| Pagellus | 33 | 24 | ■ | ■ | ■ | O | · | O | · | · | 4 | Le Loeuff et Intès (1973) | ||
| coupei | 291 | 105 | ■ | ■ | D | D | D | · | ■ | · | 4 | Longhurst (1960a) | ||
| Uranoscopus sp. | 27 | 8 | · | D | D | 1→2 | Longhurst (1960a) | |||||||
| Trigla lineata | 3 | 3 | ■ | · | · | ■ | · | 2→3 | Le Loeuff et Intès (1973) | |||||
| Lepidotrigla | 166 | 16 | · | ■ | D | D | ■ | ■ | 2®3 | Longhurst (1960a) | ||||
| cadmani | ||||||||||||||
| Cephalacantus | 7 | 7 | D | · | · | · | D | · | ■ | · | 4 | Le Loeuff et Intès (1973) | ||
| volitans | 51 | 39 | · | · | D | · | D | · | D | 4 | Longhurst (1960a) | |||
N = Estomacs examinés; N' = Estomacs non vides
1 = Ichthyophages ;
2 = Poissons et epifaune active
3 = Epifaune sédentaie et endofaune ;
4 = Très varié.
| ESPECE | ESTOMACS | POISSONS | CEPHALOPODES | PETITS | CREVETTES | CRABES | STOMATOPODES | ENSEMBLE | MOLLUSQUES | OPHIURES | POLYCHETES | CATERORIE | AUTEURS | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| N | N | CRUSTACES | PAGURES | THALASSINDES | CRUSTACES | OURSINS | ECHIURIENS | |||||||
| Raja miraletus | 182 | 123 | D | O | O | · | D | · | O | O | O | 2 | Longhurst (1960a) | |
| Trichiuris lepturus | 9 | 5 | · | 1 | Le Loeuff et Intès (1973) | |||||||||
| 83 | 27 | · | ■ | ■ | 1–2 | Longhurst (1960a) | ||||||||
| Cynoglossus | ? | 52 | · | D | ð | D | · | · | ■ | ■ | 3 | Thiam (1978) | ||
| canariensis | ||||||||||||||
1 = Ichthyophages ; 2 = Poissons et épifaune active ;
3 = Epifaune sédentaire et endofaune ; 4 = Très varié.
![]()
![]()
![]()