![]()
![]()
![]()
LES VARIATIONS SAISONNIERES D'ABONDANCE DES ESPECES
DEMERSALES DU PLATEAU CONTINENTAL IVOIRIEN
par
CAVERIVIERE Alain
Chargé Recherches de l'ORSTOM
Nous distinguerons deux types de variations qui sont liées toutes deux à la présence d'upwellings côtiers périodiques se produisant lors de l'hiver austral. Les premières sont en rapport apparent direct avec le refroidissement de l'eau tandis que les secondes - plus breves - correspondent à un phénomene trés particulier : une diminution importante de la quantite d'oxygène dissous à la fin de la saison froide, qui peut être considérée comme un effet indirect du refroidissement. D'autres variations saisonniéres non directement liées à la température, comme celles dues au recrutement et aux migrations de reproduction, peuvent également intervenir. Elles contribuent aux variations saisonniéres observées entre saison chaude et saison froide, mais dans l'ensemble leur rôle dans celles-ci est faible.
1. LES VARIATIONS SAISONNIERES EN RAPPORT APPARENT DIRECT AVEC LES REMONTEES D'EAUX FROIDES
1.1. Les données et leur traitement
L'étude utilise les données de la radiale de Grand Bassam (Troadec et al., 1969). Lorsque les données concernant plusieurs profondeurs de chalutage sont regroupées on a pondéré les rendements par l'écartement des stations (cf. annexe III du présent rapport). Nous avons également utilise les rendements standardisés moyens mensuels des chalutiers ivoiriens sur 10 ans (1968 à 1977) pour diverses strates bathymétriques. Les données de la radiale Grand Bassam peuvent présenter des variations quelque peu différentes de celles ayant lieu en moyenne sur le plateau continental, du fait du manque de répétitions à chaque période de traits de chaluts dont nous savons que les résultats présentent un caractère aléatoire important. Les rendements moyens mensuels des chalutiers permettent de suivre l'évolution saisonniére de leur pêche en Côte d'Ivoire avec une certaine sôrete, mais peuvent présenter certains biais si on veut considérer leurs variations comme représentatives de celles des abondances spécifiques au niveau du fond; en effet, les chalutiers cherchent les meilleurs rendements commerciaux possibles et les diverses rubriques de vente recouvrent généralement plus d'une seule espéce (Caveriviére et Barbe, 1977). Un bon accord entre les deux types de données (radiale Grand Bassam et rendements des chalutiers) nous permettra de penser que les variations saisonnières mises en évidence sont bien représentatives des variations moyennes qui ont lieu sur l'ensemble du plateau. Nous disposons enfin de deux sources de renseignements complémentaires:
Les rendements moyens mensuels pour quelques espéces démersales (Caveriviére, 1975) des sardiniers ayant travaillé sur le plateau continental ivoirien de 1974 à 1978, Ces valeurs devront être utilisées avec circonspection du fait de leur variabilité interannuelle, de ce que la pêche s'effectue toujours en deçà des 50 m et plutôt entre 25 m et la côte, des possibles reports des efforts d'une espéce à une autre (particuliérement quand la pêche de Sardinella aurita est faible, soit pendant les saisons des pluies et saisons froides marquées d'aprés Marchal, 1966).
Les données des campagnes CHALCI (Caverivière et Champagnat, 1978, 1979; Caverivière, 1980) qui permettent de comparer des rendements de saison chaude (CHALCI 78.01 et 79.01) et saison froide (CHALCI 80.01). Du fait que ces données n'ont pas été obtenues la même année et du fait qu'elles ne couvrent qu'une partie des différentes saisons, il ne sera fait appel à elles que pour confirmer des variations saisonnières qui n'ont été observées qu'a partir des radiales de Grand Bassam, mais aussi et surtout pour fourrir un élément d'appréciation supplémentaire quand ces dernières variations ne concordent pas avec celles observées par ailleurs. Le Tableau 1 donne les rendements par campagne et par grande strate bathymétrique pour les principales espèces, ainsi que pour celles moins importantes pour lesquelles ce type de donnée a été pris en compte.
Pour mieux appréhender les variations saisonnières d'abondance, l'évolution des redements des chalutiers et sardiniers sera représentée en pourcentages (indices d'abondance) par rapport à la moyenne annuelle (ou pluriannuelle) dans les tableaux et figures correspondantes. La séparation entre saison chaude et saison froide a été définie par l'apparition de l'isotherme 24°C en surface. La saison froide englobe, en 1966, les radiales Grand Bassam n° 10 à 13 (27 juillet au 30 septembre) et les 14 autres radiales seront considérées comme étant de saison chaude. En ce qui concerne les données statistiques des chalutiers et des sardiniers, les mois de juillet, aoôt et septembre représentent la saison froide.
1.2. Communauté des Sciaenidés
Dasyatis marqarita (Dasyatidae). Sur les fonds de 15 m de la radiale Grand Bassam -où se trouve le maximum d'abondance - le rendement moyen est plus élevé en saison froide qu'en saison chaude, alors que c'est l'inverse pour les fonds de 20 à 35 m (Tableau 2). Il semble qu'il y aurait donc un regroupement de l'espèce près de la côte lors de la saison froide. Le rapport global SC/SF, pondéré par les surfaces 15-30 m,est proche de 2.
Ilisha africana (Clupeidae). Les rendements horaires de saison chaude, pour chaque immersion de la radiale Grand Bassam où l'espèce est présente, sont toujours supérieurs à ceux obtenus en saison froide (Tableau 2), ce qui pourraît signifier qu'un évitement des eaux froides par ce clupeidé s'effectuerait probablement par un déplacement vers la surface. Les données statistiques des chalutiers ivoiriens pour la strate 10-50 m (de 1968 à 1973 seulement car par la suite les rendements sont devenus très faibles et imprécis) semblent montrer que l'abondance sur le fond serait la plus élevée de décembre à février (Tableau 3 et Figure 1a), la variabilité étant très élevée.
Le rapport des rendements entre saison chaude et saison froide pour la strate bathymétrique 10-50 m est de 1,4 à partir des radiales Grand Bassam et de 1,2 à partir des données statistiques des chalutiers. Durand (1967) trouve au Congo un rapport élevé proche de 11 sur la radiale de Pointe-Noire.
L'examen des statistiques des sardiniers (Tableau 4) paraît montrer que les rendements les plus élevés sont plutôt obtenus, à l'inverse des chalutiers, en saison froide (bien qu'ici aussi la variabilité interannuelle des rendements mensuels soit forte), ce qui corrobore l'hypothèse que l'évitement des eaux froides augmenterait le caractère pélagique de l'espèce. Le rapport saison chaude/saison froide pour les sardiniers est de 0,5 ; ce dernier chiffre ne doit être pris que comme l'indication d'une tendance qui devra être vérifiée avec plus de données.
Lagocephalus laevigatus (Tetraodontidae). D'après les rendements moyens du tableau 2, l'espèce est moins abondante en saison froide dans les prises de la radiale Grand Bassam.
Sphyraena spp. (Sphyraenidae). Les rendements (Tableau 2) de saison chaude sont nettement supérieurs à ceux obtenus en saison froide; le rapport SC/SF est proche de 21. Les rendements des sardiniers sont très variables (Tableau 4): de fortes prises peuvent avoir lieu aussi bien en saison chaude qu'en saison froide. Rijavec (1971) signale au Ghana une montée à la côte en saison froide.
Galeoides decadactylus (Polynemidae). Les rendements obtenus lors des radiales Grand Bassam sont plus élevés en saison chaude, et ceci pour toutes les profondeurs (Tableau 2). Le rapport(zone 15-50m) SC/SF = 3,4 alors qu'il est de 1,7 pour les données des prises des chalutiers ivoiriens. Le rapport trouvé par Durand au Congo est supérieur à 4. L'évolution des indices moyens de rendement des chalutiers est très régulière (Tableau 3 et Figure 1b). L'indice est le plus élevé en janvier et atteint son minimum au mois de septembre; il remonte ensuite. Nous noterons que les 10 indices annuels (dont la moyenne est seule indiquée dans le tableau 2) sont supérieurs à 100 pour les mois de janvier, février, mars et toujours inférieurs à cette valeur pour les mois de juillet, août, septembre.
Les indices de rendement des sardiniers indiquent également que les plus fortes prises ont lieu en saison chaude (Tableau 4). Notons également que la pêche artisanale utilise des filets maillants adaptés à la capture de gros capitaines (filet “Kuatchinfo” et “Yakur”) pendant la saison chaude (Bouberi et al. 1980). Ces variations saisonnières des rendements s'expliqueraient (Samba, 1974) par une remontée à la côte des Galeoides decadactylus en saison froide sur des fonds très faibles, ainsi cet auteur écrit: “Dans la baie de Pointe-Noire, les pêcheurs ont capturé des capitaines, soit avec leurs filets droits maillants à grandes mailles,... soit à la senne de plage,... sur les fonds inférieurs à 15 mètres” (loc.cit.p.28). Au Ghana, les rendements obtenus avec un chalut à grande ouverture verticale sont les plus élevés en saison froide, il y aurait donc un certain décollement du fond en cette période.
Pentanemus quinquarius (Polynemidae). Les rendements de saison chaude sont toujours supérieurs à ceux de saison froide sur la radiale Grand Bassam (Tableau 2), leur rapport est de 4,3. La différence entre la saison chaude et la saison froide est moins marquée au Congo.
Pomadasys jubelini (Pomadasyidae). Les prises de saison froide sont en moyenne supérieures à celles de saison chaude pour les deux premiéres stations, 15 et 20 m, de la radiale Grand Bassam (Tableau 2). Le rapport global des rendements moyens entre les deux saison est proche de 1 (0,94). Les données sur 10 ans concernant la rubrique “carpes” (composée en majorité de Pomadasys jubelini) des chalutiers commerciaux pour la strate bathymétrique 10-50 m montrent que les meilleurs rendements sont obtenus en saison froide (Tableau 3 et Figure 1), le rapport SC/SF = 0,8.
C'est l'inverse pour les sardiniers dont les prises sont faibles à nulles en période froide (Tableau 4). En lagune Ebrié, les plus fortes abondances au niveau du fond prendraient place en saison froide. Les données de la radiale Grand Bassam paraissent indiquer une remontée à la côte en période, mais un autre phénomène saisonnier jouerait un rôle important dans les variations des rendements des chalutiers et sardiniers, qui sont à l'opposé de celles auxquelles on aurait pu s'attendre pour une espèce côtière de la communauté dés Sciaenidés, et qui sont d'ailleurs décalées par rapport aux saisons climatiques. Il s'agit de la reproduction qui aurait plutôt lieu en saison froide (d'avril à août) comme cela a été observé au Sénégal (Alberdi, 1971), période que les quelques données dont nous disposons semblent confirmer pour la Côte d'Ivoire. Pendant l'époque de la reproduction les Pomadasys jubelini pourraient être plus proches du fond, cette hypothèse permettrait d'expliquer les variations de rendement observées.
Brachydeuterus auritus (Pomadasyidae). Lors des radiales Grand Bassam de 1966–67 les rendements de 15 à 80 m ont été beacoup plus faibles (8,7 fois pour l'ensemble de la strate) en saison froide qu'en saison chaude (Tableau 2). Durand trouve un rapport inverse (0,7) au Congo, mais note un resserrement de la répartition à la côte en période froide. Les rendements des chalutiers ivoiriens pour la strate 20-50 mètres montrent également une diminution en saison froide (Tableau 3 et Figure 1d), qui est cependant moins marquée (rapport SC/SF =1,4), la variabilité est élevée. Les rendements moyens obtenus pendant les campagnes CHALCI confirment (Tableau 1) les variations qui ont déjà été observées en Côte d'Ivoire par chalutage de fond. Pour les sardiniers, dont 10 à 25% des mises à terre annuelles sont composées de friture, les rendements sur 12 ans (1966–1977) présentent des indices moyens dont la variabilité est très forte et qui ont des valeurs élevées pendant les mois de novembre à février, l'indice le plus bas est obtenu en avril (Tableau 3 et Figure 1e).
Rijavec (1971) note qu'au Ghana les Brachydeuterus auritus sont particulièrement abondants par petits fonds (18–27m)durant la saison froide. Ces rendements élevés ont été obtenus par comparaison avec les rendements obtenus en Côte d'Ivoire et sa distribution bathymétrique en saison chaude, on peut supposer qu'il y a, en période froide, un décollement plus important du fond couplé avec une remontée vers la côte. Nous ne pouvons cependant formuler, pour le moment, aucune hyptothèse explicative concernant les indices moyens très importants obtenus par les sardiniers en décembre et janvier, et qui doivent être représentatifs de la pêche bien que les prises de friture puissent ne pas être bonnes chaque année durant ces mois.
Pseudotolithus typus (Sciaenidae). Les prises moyennes de saison froide sont plus faibles qu'en saison chaude, sauf pour la station la plus côtière de la radiale de Grand Bassam (Tableau 2). Le rapport SC/SF - 2,7. Au Congo, Durand trouve un rapport inférieur à 1 (0,7).
Pseudotolithus senegalensis (Sciaenidae). Les prises de saison chaude sont généralement plus fortes qu'en saison froide. Comme pour Pseudotolithus typus les fonds de 15 m font exception (Tableau 2); les deux radiales de novembre ont fourni les rendements globaux les plus élevés. Le rapport SC/SF = 1,4. Troadec (1971), qui a étudié en détail la répartition de l'espèce en Côte d'Ivoire, trouve une bonne relation entre la température moyenne sur le fond et les indices d'abondance; la petite saison froide aurait également un effet négatif sur les rendements.
Troadec montre une remontée sur les fonds de 15 à 20 m quand les eaux sont froides, ainsi qu'un déplacement vers la surface. Fontana (1979) confirme l'analyse de Troadec en indiquant qu'il a pu observer des concentrations importantes de Pseudotolithus seneqalensis à la côte, “chassant” des bancs de petites sardinelles. Ces prédateurs peuvent alors être capturés à la trafne en utilisant une cuillère comme leurre. Notons que le rapport SC/SF a été trouvé très proche de 1 sur la radiale de Pointe-Noire au Congo avec, à l'inverse de la radiale de Grand Bassam, des valeurs à 15 m nettement plus élevées en saison chaude qu'en saison froide et que Durand relie à la concentration des adultes sur les petits fonds pour la reproduction.
Les indices moyens de rendement (10-50 m) sur 10 ans des chalutiers opérant en Côte d'Ivoire montrent de fortes valeurs en début de saison chaude, qui diminuent par la suite pour atteindre un palier minimum de mai à septembre (Tableau 3 et Figure 1f). Les rendements des chalutiers congolais sont également plus élevés en saison chaude qu'en saison froide (Poinsard, 1969, Fontana, 1979). Les sardiniers présentent des indices moyens importants de décembre à février (Tableau 4), mais de fortes prises peuvent entre relisées pour une année donnée pendant un mois ou l'autre de saison froide.
Pteroscion peli (Sciaenidae). Sur la radiale Grand Bassam cette espèce a montré un rendement moyen 1,5 fois plus élevé en saison chaude qu'en saison froide, les rapports SC/SF étant supérieurs à 1 pour toutes les profondeurs, sauf pour 15 m (Tableau 2). Il n'y a apparemment pas eu de variations saisonnières au Congo à l'époque des radiales Pointe-Noire. Les résultats des campagnes CHALCI (Tableau 1) confirment la baisse observée en saison froide au large de Grand Bassam.
Selene dorsalis (Carangidae). Les rendements sont toujours plus faibles en saison froide (Tableau 2), excepté pour la profondeur 60 m dont la valeur moyenne élevée par rapport à ses voisines est principalement due à la pêche (30 kg) effectuée le 29.9.66 qui a peut-être pu se produire lors de la remontée du chalut. Le rapport global SC/SF = 2,5. Au Congo, les prises sur la radiale de Pointe-Noire ont été insignifiantes. La baisse des rendements en saison froide s'observe également à partir des campagnes CHALCI (Tableau 1).
1.3 Communauté des Sparidés
Mustelus mustelus (Triakidae). Bien que les rendements soient trop faibles pour se prononcer avec certitude, cette espèce paraît présenter d'intéressantes variations saisonnières d'abondance (Tableau 5). Les rendements moyens sont les plus élevés en saison froide (rapport SC/SF = 0,2) et il y aurait à cette époque un déplacement de l'espèe vers des fonds moindres, qui se traduit par le fait que les seules prises significatives à 25 et 30 m ont été effectuées en saison froide tandis que la présence de l'espèce sur les fonds de 100 et 200 m n'est signalée qu'en saison chaude. Les données des campagnes CHALCI montrent également (Tableau 1) une nette augmentation des rendements en saison froide.
Priacanthus arenatus (Priacanthidae). L'abondance est généralement plus élevée en saison chaude qu'en saison froide (Tableau 5). Le rapport SC/SF est proche de 0.4. Notons cependant que cette variation n'est pas confirmée par les resultats des campagnes CHALCI.
Pentheroscion m'bizi (Sciaenidae). L'abondance de cette espèce (Tableau 5) est plus élevée en saison froide qu'en saison chaude sur les petits fonds de sa distribution (50 et 60 m); le rapport paraît inversé pour les stations les plus profondes sil'on considère que la forte yaleur moyenne obtenue pour les fonds de 100 m en période froide est due à une prise exceptionnelle de 216 kg le 29.9.66.II y auraitici aussi une remontée de l'espèce en saison froide modifiant l'abondance sur les fonds. Cette “montée” de l'espèce à également été observée au Congo. Le rapport moyen des rendements SC/SF est proche de 0,7 pour la radiale de Grand Bassam et de 0,5 pour calle de Pointe-Noire.
Trachurus trecae (Carangidae). Les rendements moyens à Grand Bassam sont beaucoup plus élevés en saison froide qu'en saison chaude (Tableau 5). L'observation station par station des données de la radiale (Troadec et al., 1969) montre en fait qu'un pic marqué des prises à eu lieu de septembre à la mi-octobre, soit pendant la deuxième moitié de la saison froide. Au Congo, où son abondance est plus élevée, les rendements sur la radiale de Pointe-Noire n'ont pas montré de changements significatifs d'une saison à l'autre, par contre, une remontée assez nette à été observée en saison froide. Notons enfin que les prises effectuées loan de la campagne CHALCI 80.01 (période froide) ont été très faibles (Tableau 1).
Dentex angolensis (Sparidae). Durand indique qu'au Congo les rendements doublent en saison froide et note également une remontée sensible du poisson pendant cette période. Le rapport global moyen SC/SF proche de 0,6 lors des radiales Grand Bassam (Tableau 5) est en accord avec la première observation.
Dentex canariensis et Pagrus ehrenbergi (Sparidae). Les prises en Côte d'Ivoire sont insuffisantes pour pouvoir déceler des variations saisonnières pour ces deux espèces. Rijavec (1973) note au Ghana une augmentation apparente de leur abondance en saison froide, accompagnée de déplacement vers la côte.
Pagellus coupei (Sparidae). Lors des radiales Grand Bassam le rapport global moyen SC/SF a été proche de 0,7. Rijavec (1973) note à partir des rendements d'un chalut à grande ouverture - comme pour Dentex canariensis et Pagrus ehrenberqi -une augmentation apparente de l'abondance ainsi qu'une migration vers la côte en saison froide. Ce déplacement peut être décelé à partir des données de la radiale Grand Bassam (Tableau 5); 82% des prises de saison froide ont lieu avant la station 60 m, contre 66% en saison chaude.
Les statistiques de pêche des chalutiers ivoiriens, qui regroupent la majorité des prises de sparidés dans l'appellation “pageot”, confirment de manière globale les résultats que nous venons d'indiquer pour Dentex angolensis et Pagellus coupei. Le tableau 6 et la figure 2 présentent les indices moyens mensuels de rendement pour les pageots capturés sur les fonds de 21 à 50 m, ainsi que des indices similaires - calculés différemment de la manière jusqu'ici utilisée* - pour ceux pêchés dans la strate 50–120 m. Il apparaît nettement que les rendements de pageot dans les deux strates sont les plus élevés en saison froide.
Notons cependant pour Pagellus coupei des indications contradictoires à une telle variation saisonnière d'abondance : elles sont fournies par les résultats de la radiale de Pointe-Noire au Congo (où Durand note - avec précaution - un rapport SC/SF nettement favorable à la saison chaude) et par ceux des campagnes CHALCI (cf. Tableau 1)
Brotula barbata (Brotulidae). Les rendements de saison froide ont été en moyenne de près du double de ceux de saison chaude sur la radiale de Grand Bassam (Tableau 5). Cette évolution est confirmée par les résultats des campagnes CHALCI (Tableau 1).
Balistes carolinensis (Balistidae). Cette espèce, dont la biomasse s'est récemment développée, présente de faibles abondances sur le fond en saison froide (Caverivière, 1979; Tableau 1).
Paracubiceps ledanoisi (Stromateidae). Son abondance diminuerait au niveau du fond en saison froide. Le rapport SC/SF = 4,6; il dénoterait une accentuation du caractère pélagique de l'espèce en saison froide. La baisse des rendements en période d'upwelling s'observe également à partir des campagnes CHALCI.
1.4 Espèces eurybathes ou de la thermocline
Raja miraletus (Rajidae). Des variations saisonnières de l'abondance peuvent être suivies à partir des radiales Grand Bassam (Tableau 7). L'espèce est plus abondante en saison froide qu'en saison chaude sur les fonds de 15 à 35 mètres, un équilibre s'établit à 40 m et à partir de 50 m c'est l'inverse qui se produit. Il y aurait donc un mouvement de l'espèce vers la côte en saison froide. Le rapport moyen SC/SF = 0,7. L'observation des données détaillées de la radiale Grand Bassam (Troadec et al., 1969) permet de dire qu'il y a eu également une remontée de l'espèce pendant la petite saison froide. Les données du Congo concordent avec celles de Côte d'Ivoire.
Trichiurus lepturus (Trichiuridae). Les rendements moyens de “ceintures” se sont montrés toujours plus élevés en saison froide qu'en saison chaude sur les fonds de 15 à 60 mètres de la radiale Grand Bassam, l'inversion notée plus bas ne peut être considérée comme significative mais traduit peut-être une certaine remontée de l'espèce en période froide. Une observation par radiale des données (Figure 3) montre cependant que les deux radiales de saison froide effectuées en juillet présentent des abondances très faibles alors que de forts rendements sont obtenus en début de saison chaude. Le rapport moyen global des rendements SC/SF avoisine 0,5.
Les indices moyens mensuels de rendement des chalutiers pour les fonds de 21 à 50 m présentent un pic très marqué au niveau des trois derniers mois de l'année (Tableau 8 et Figure 4); de plus ils ont toujours été pendant 10 ans supérieurs à 100 en novembre et décembre, ce qui donne à ce pic un haut degré de signification. Les indices moyens sur 5 ans des sardiniers (Tableau 8) ont une grande variabilité d'une année à l'autre pour un mois donné, avec ici aussi les plus forts indices observés pendant la première moitié de la saison chaude. L'ensemble des données à notre disposition montrerait donc une augmentation générale de la disponibilité de l'espèce aux engins de pêche plus ou moins étalée de part et d'autre du mois de novembre. La cause exacte de cette augmentation est actuellement inconnue.
Des études biologiques sur Trichiurus lepturus actuellement en cours en Côte d'Ivoire indiquerait que le maximum de la reproduction coinciderait avec le pic des rendements (Bouberi, com. pers.). Une certaine variabilité interannuelle dans la position ou l'étalement du pic d'abondance pourrait peut-être expliqùer que les prises moyennes en 1966–67 sur la radiale de Grand Bassam ont été plus élevées en saison froide qu'en saison chaude, alors que les données 1963–65 de la radiale Pointe-Noire au Congo donnent un rapport inverse (Durand, 1967), ainsi d'ailleurs que les données des campagnes CHALCI (Tableau 1).
Cynoglossus canariensis (Cynoglossidae). Les rendements moyens de saison chaude et de saison froide obtenus lors des radiales Grand Bassam sont proches, leur rapport pour l'ensemble de la radiale est égal à 1. Les rendements des chalutiers ivoiriens pour la strate 21–50 m montrent que leurs meilleurs résultats sur cette espèce sont obtenus en saison froide (Tableau 8 et Figure 5). Les distributions de fréquence ne montrent pas de période de recrutement particulièrement élevée. La baisse des rendements 21–50 m en saison chaude correspondrait à la période de ponte (Chauvet, 1970) pendant laquelle les reproducteurs viendraient à la côte. Il pourrait peut-être également y avoir, à ce stade, une diminution de la vulnérabité par un accroissement de l'enfouissement.
2. VARIATION SAISONNIERES CONSECUTIVES A LA DIMINUTION DU TAUX D'OXYGENE DISSOUS
Nous savons que les variations de la teneur en oxygène sont très sensibles sur les fonds de 15 à 50 m et jusqu'à 80 mètres (Troadec, 1971). En règle générale, la quantité d'oxygène dissous décroît à partir de début juin pour atteindre un minimum vers septembre - début octobre, fonction de la durée et de la force de l'upwelling (verstraete, 1970). Les teneurs en oxygène peuvent alors être très faibles (inférieures à 1 m1/1 et 20 % de saturation), elles sont dues à une importante réduction bactérienne consécutive à une forte production phytoplanctonique, la thermocline qui se reconstitue à la fin de l'upwelling faisant de plus un barrage à la diffusion de l'oxygène atmospherique.
Troadec (1971) montre, après traitement des données de la radiale Grand Bassam, que les rendements de Pseudotolithus senegalensis paraissent, de manière très probable, diminuer nettement du fait de la teneur en oxygène dissous quand celle-ci est inférieure à 2,0 ml/l. Il note que le traitement des données pourrait masquer un effet des teneurs en oxygène pour des concentrations légèrement supérieures. Le tableau 9 montre qu'une raréfaction très importante de l'espèce coincide avec l'apparition d'un minimum d'oxygène bien marqué au niveau des fonds de 15 à 25 mètres. Troadec indique également que l'effet négatif des faibles concentrations en oxygène agirait sur la plupart des espèces de la communauté des Sciaenidés, seuls Pteroscion peli et Ilisha africana ont été pris pendant cette période en quantités encore appréciables, ainsi que l'espèce eurybathe Trichiurus lepturus.
Les données des campagnes CHALCI ne permettent pas d'analyser les variations des captures en fonction de la teneur en oxygène dissous. Cependant, pendant toute la durée de la campagne CHALCI 80.01 (20.08 au 07.09.80), nous avons observé sur les fonds côtiers (15-40 m) des taux le plus souvent inférieurs à 2,0 ml; en particulier sur les fonds 20–40 m des deux radiales Grand Bassam qui ont été effectuées au début et en fin de campagne (1,7 à 1,9 ml/l pour la première et 1,4 à 1,6 ml/l pour la seconde). Or, de fortes prises de Pseudotolithus senegalensis ont été réalisées pendant la première radiale sur les fonds de 20–25 m (83 et 86 kg) et les captures de la seconde sont d'un bon niveau (28 et 22 kg). Ces prises, ainsi d'ailleurs que celles de Pomadasys jubelini (23,13,26 et 25,5 kg), nous conduisent à penser que l'effet de la diminution du taux d'oxygène dissous n'est peut-être pas aussi net que ce qu'il paraissait à partir des travaux de Troadec, tout au moins pour les teneurs comprises entre 1,5 et 2,0 ml/l.
3. DISCUSSIONS ET CONCLUSIONS
Lors des radiales Grand Bassam l'abondance moyenne sur les fonds a montré, pour presque toutes les espèces étudiées appartenant aux communautés des Sciaenidés et des Sparidés, des variations saisonnières allant dans le même sens à l'intérieur de chaque groupe, mais de sens opposés entre eux : rendements plus élevés en saison chaude pour la communauté des Sciaenidés (sauf Pomadasys jubelini qui montre un rapport SC/SF proche de 1) et rendements plus élevés en saison froide pour la communauté des Sparidés (sauf Priacanthus arenatus et Paracubiceps ledanoisi).
Ces variations de sens inverses apparaissent également quand l'on considère les prises toutes espèces pour les fonds de 15 à 50 m de la radiale (peuplés en majorité par les espèces de la communauté des Sciaenidés) et pour les fonds de 60 à 100 m (communauté des Sparidés(*)). Ainsi on peut observer sur la figure 6- malgré une importante variabilité intra-saisonnière - une diminution globale des rendements horaires sur les fonds de 15 à 50 m en saison froide et une augmentation de ceux obtenus sur les fonds de 60 à 100 m, les rendements sur ces derniers devenant alors supérieurs aux premiers. On notera que les plus mauvais rendements sur les fonds côtiers ont été obtenus à la mi-juin pendant une période de mauvais temps et alors que la température était encore élevée; cette baisse est donc plus nette que celle que correspond au minimum d'oxygène, qui n'est pas très marquée pour l'ensemble de la strate. En ce qui concerne la strate profonde, le pic obtenu à la mi-avril n'est pas représentatif de la communauté des Sparidés car dû à une prise importante de Brachydeuterus auritus (500 kg) à 60 m. Les variations de rendement pour l'ensemble du plateau (Figure 6) sont plus faibles - du fait des évolutions opposées de l'abondance des deux principales communautés - et montreraient au total une légère diminution en saison froide.
* : La pondération suivant l'écartement des stations a été effectuée.
Les indices mensuels moyens de rendement (toutes espèces) des chalutiers présentent de faibles variations pour l'ensemble du plateau et pour les fonds 10–50 m (Tableau 10), strates dont les indices moyens sur 10 ans et leurs évolution peuvent être considérés comme représentatifs malgré des coefficients de variation le plus souvent supérieurs à 10%. Une diminution des rendements en saison froide est peut-être à peine perceptible pour les fonds de 10 à 50 m et ne l'est pas du tout pour l'ensemble du plateau. En ce qui concerne la strate 50–120 m nous avons effectué le calcul des indices mensueals comme pour les captures de “pageot” des chalutiers (cf. paragraphe. 1.3). Il y aurait une certaine augmentation en saison froide (Tableau 10), bien que les indices pour ces fonds soient peu sûrs.
La diminution d'amplitude de la variabilité inter-mensuelle quand on considère les rendements des chalutiers, par rapport à ceux obtenus sur la radiale de Grand Bassam par un navire de recherche, n'est pas anormale. Il y a en effet un certain effet tampon des statistiques de pêche. Par ailleurs les chalutiers recherchent les meilleurs rendements: un navire ne continuera à travailler dans une strate peu habituelle pour la saison (mais qui est toujours plus ou moins prospectée) que tant qu'il obtient des rendements supposés aussi bons que dans la zone saisonnière habituelle. L'évolution des rendements obtenus sur la radiale Grand Bassam donne donc une idée plus exacte des variations saisonnières d'abondance que l'évolution des indices mensuels calculés pour les chalutiers.
Les variations observées sur cette radiale expliquent pourquoi 74% de l'effort des chalutiers sur les fonds supérieurs à 50 m(Tableau 11) est appliqué d'août à octobre : c'est la période pour laquelle les rendements sur ces fonds sont supérieurs à ceux obtenus dans la zone plus côtière.
Remarquons sur le tableau 11 que l'effort optimum dans la strate profonde a lieu en septembre et octobre, ce qui nous paraît correspondre à une augmentation réelle de l'abondance sur ces fonds. Ce maximum est décentre par rapport au creux formé en saison froide par l'evolution des températures moyennes qui est un peu plus précoce. L'abondance dans cette strate serait donc plus élevée en fin de saison froide qu'en début, sans que la température puisse être mise en cause; cette augmentation a été décelable en 1966 sur la radiale Grand Bassam (Figure 6). L'accroissement du nombre de copépodes zooplantoniques dont les effectifs suivent une évolution parallèle (Figure 7) est peut-être en relation avec ce décalage.
Les comparaisons des résultats des campagnes CHALCI (Tableau 12) concordent bien dans l'ensemble avec ce que nous venons d'écrire sur les rendements globaux. Ainsi les abondances apparentes pour la strate 10–50 m pendant les campagnes de saison chaude (CHALCI 78.01 et 79.01) ont toujours été supérieures à celles de saison froide (CHALCI 80.01), que l'on considére les prises totales ou des seuls téléostéens, diminuées ou non des balistes et fritures (Brachydeuterus auritus) qui ne sont pas ou peu recherchés par les chalutiers. L'accord n'est bon pour la strate 50–120 mètres que si l'on ne considère que les prises totales diminuées de celles des balistes (dont la biomasse êtait insignifiante avant 1971–72) ou, mieux, de celles des balistes plus fritures.
Nous nous sommes posés la question de savoir sides variations saisonnières d'abondance sont d'autant plus accentuées d'une année sur l'autre que le sont les variations de la température (qui dépendent de l'intensité des remontées d'eau froide). Pour ce faire, nous avons calculé plusieurs indices d'upwelling. Pour l'année et pour la période juillet-octobre, au quels nous avons comparé les rendements moyens totaum et de “pageots” obtenus par les chalutiers sur les fonds 10–50 m et supérieurs à 50 m pour les mémes périodes. Aucune relation entre les valeurs des indices d'upwelling et les rendements n'a pu etre mise en évidence, ce qui aurait été le cas, malgré les efforts tampons sur les données statistiques et les possibilités de biais trendements dépendant des efforts des années précédentes, changements d'abondance dus à des modifications de l'écosystème,…), siles relations pouvant exister étaient bien marquées au niveau du fond.
Les variations saisonniéres d'abondance correspondent à des changements dans l'abondance apparente au niveau du fond, qui peuvent étre dus à des déplacements horizontaux ou verticaux ainsi qu'à l'arrivée de classes nouvellement recrutées. Les migrations verticales existent apparemment pour de nombreuses espèces. particulièrement pour celles de la communauté des Scianidés qui évitent, en se déplaçant vers la surface, les remontées d'eaux trop froides en periodes d'upwellings.
Les migrations horizontales vers la côte peuvent obeir au même phénomène d'évitement - ou au contraine à la montée des sparidés en saison froide - et peuvent aussi étre liées de manière directe à la reproduction. Ces migrations verticales et horizontales vers la côte sont de plus en plus confirmeées en Côte d'Ivoire par l'étude des péches artisanales (traine, palangrotte, filtes maillants). Des migrations horizontales latérales hors des eaux de Côte d'Ivoire ne nous paraissent guère avoir lieu pour les espèces démersales, quoique seuls quelques marquages de Pseudotolithus senegalensis aient été effectués dans cette région (Troadec, 1971). Par contre elles pourraient exister pour les espèces semi-pélagiques capturées par les chalutiers et il a été supposé qu'elles sont sans doute importantes pourBalistes carolinensis. L'arrivée des recrues dans la pêcherie - soit par sélectivité des chaluts, soit par migration des jeunes sur les fonds de péche - ne modifie apparemment jamais grandement l'abondance des espèces dans les captures et leur répartition sur les fonds, bien que les nombres de classes d'âge contribuant significativement aux prises soient très généralement faibles; ceci sans doute du fait d'une répartition assez étalée des périodes de ponte. Cette absence d'effet notable du recrutement est caractéristique des eaux tropicales de l'Afrique de l'Ouest, même quand les saisons sont bien marquées et plus généralement de toutes les eaux tropicales.
BIBLIOGRAPHIE
Albendi,P.G.,1971 - Biologie et pêche du “sompat” Pomadasys Ubelini (Curier 1830) des eaux sénégambiennes (apençu préliminaire). Centre de Pech Océanogr. Dakar-Thiaroye,Doc.Scient.Prov.n°30:38p.
Binet, D.,1977 - Grands traits de Méologie des principaux tanos du zooplancton ivoirien. Cah. CRSTOM, ser. Océanogr., vol.XV, o° 2 : 89–110.
Boubeni, D., Hié Dané,J.P. et Konan, J.,1983 - La pêche artisanale maritime en Côte d'Ivoine. Le potentiel de pêche. Centre de Pech.Océanogr. Abidjan, Arch.Scient., vol.IX,n° 1:11–29.
Caverivière, A., 1975 - Traitement des données de prises des sardiniers ivoiriens pour “19 espèces”. Centre de Rech.Océanogn.Abidjan, Rapp. interne.5p., miméo.
Caverivière, A., 1979-Estimation des potentiels de pêche des stocks dimersaux ivoiriers par les modèles globaux, Effets de la prolifénation du baliste (Balistes capriscus). Centre de Rech.Océanogr.Abidjan, vol.X, n° 2:5–164.
Caverivière.A., 1980 - Campagne CHALCI 80.01 (20.8.80–10.9.80).Résultats des chalutages. Centre de Rech.Océanogr.Abidjan, Arch.Scient., vol.6,n° 3 :198 p.
Caverivière, A.et Barbe, F., 1977 - Traitements statistiques des données de la pêche au chalut des poissons démersaux en Côte d'Ivoire. Prises - Efforts - Distribution de fréquence. Centre de Rech.Océanogr. Abidjan,Arch.Scient.,vol.2,n° 1 : 35p.
Caverivière, A.et Champagnat, C., 1978 - Campagne CHALCI 78.01 (30.1.78 au 12.2.78). Résultats des chalutages. Centre de Rech.Océanogr.Abidjan, Arch.Scient., vol.4, n° 1 : 118p.
Caverivière : A. et Champagnat, C., 1979 - Campagne CHALCI 79.01 (13.3.79 au 31.3.79). Résultats des chalutages. Centre de Rech. Océanogr.Abidjan. Arch.Scient., vol.5, n° 4 : 198p.
Chauvet, C., 1970 - Etude de Cynoglossus canariensis (STEIN) du plateau continental de la Côte d'Ivoire : lecture d'âge, étude de la croissance en longueur et en poids - observations sur leur biologie et leur dynamique. Thèse 3° cycle, Fac.Scien.Aix-Marseille, 102p.
Durand, J.R., 1967 - Etude des poissons benthiques du plateau continental congolais. Troisième partie : étude de la répartition, de l'abondance et des variations saisonnières. Cah.ORSTOM, sér. Océanogr., vol. V, n° 2 :3–68.
Fontana, A., 1979 - Etude du stock démersal côtier congolais. Biologie et dynamique des principales espèces exploitées. Propositions d'aménagement de la pêcherie. Thèse Doct. Etat Sciences Nat., Univers. Pierre et Marie Curie - Paris VI : 300p.
Poinsard, F., 1969 - La pêche au chalut à Pointe-Noire. UNESCO, Actes symp. océanogr. et ress. halieut. Atl.trop., Rapp.de Synthèse et Comm., Abidjan 1966 : pp. 381–390.
Rijavec, L., 1971 - A survey of the demersal fish resources of Ghana. Rapp. FAO/UNDP, 40p. multigr. - Aussi FAO, 1980, CECAF/TECH/80/25.
Rijavec, L., 1973 - Biology and dynamics of Pagellus coupei (Dieuz. 1960), Pagrus ehrenbergi (val. 1830) and Dentex canariensis (poll.1954) in Ghana waters. Centre Rech.Océanogr.Abidjan, Doc.Sci., vol 4, n°3 : 43–97.
Samba, G., 1974 - Contribution à l'étude de la biologie et de la dynamique d'un Polynemidae quest-africain, Galeoides decadactylus (Bloch 1793). Thèse 3° cycle, Universe, Bordeaux I'n° d'ordre 1141 :122p.
Troadec, J.P., 1971 - Biologie et dynamique d'un Sciaenidae quest africain, Pseudotolithus senegalensis (V.). Centre de Rech. Océanogr.Abidjan, Doc.Scient., vol.II,n° 3 : 225p.
Troadec, J.P., Barro, M. et Bouillon, P., 1969 - Pêche au chalut sur la radiale de Grand Bassam (Côte d'Ivoire) (mars 1966-février 1967). Centre de Rech.Océanogr.Abidjan, Doc.Scient.Prov., n° 33 : 14p. + 11 tab.+ 89p.
Verstraete, J.M., 1970 - L'oxygène au large de Grand Bassam. Centre de Rech. Océanogr. Abidjan, Doc.Scient., vol.I, n° I : 19–35.
| CHALCI 78.01 | CHALCI 79.01 | CHALCI 80.01 | |||||||
|---|---|---|---|---|---|---|---|---|---|
| ESPECE | 10- 50m N=27 | 50- 120m N=25 | 10- 120m N=52 | 10- 50m N=26 | 50- 120 m N=40 | 10- 12 m N=66 | 10- 50 m N=49 | 50- 120 m N=39 | 10- 120 m N=88 |
| Mustelus mustelus | 1,0 | 2,2 | 1,7 | 1,8 | 1,6 | 1,7 | 4,6 | 4,1 | 4,3 |
| Raja miraletus Saleodes | 6,0 | 2,0 | 3,6 | 4,4 | 2,8 | 3,4 | 5,7 | 2,2 | 3,6 |
| decadactylus | 9,8 | 1,4 | 4,8 | 14,1 | 0 | 5,7 | 3,9 | 0 | 1,6 |
| Priacanthus anetus | 0,2 | 1,5 | 1,0 | 0,2 | 2,9 | 1,8 | 0,3 | 2,4 | 1,5 |
| Pomadasys jubelini | 5,8 | 0 | 2,4 | 14,6 | 0 | 5,9 | 6,0 | 0 | 2,4 |
| Brachydeuterus auritus | 42,1 | 20,7 | 29,4 | 59,0 | 28,2 | 41,0 | 29,7 | 0 | 12,1 |
| Fterosr peli | 8,7 | 0,1 | 3,6 | 11,3 | 0 | 4,6 | 5,5 | 0 | 2,2 |
| Pentherscion m'bizi | 0 | 6,3 | 3,7 | 0,1 | 8,4 | 5,0 | 1,0 | 3,1 | 2,2 |
| Pseudotolithus senegalensis | 11,2 | 0,1 | 4,6 | 21,8 | 0,4 | 9,1 | 8,8 | 0,2 | 3,7 |
| Trachurus trecae | 0,3 | 1,4 | 1,0 | 1,1 | 4,8 | 3,3 | 0,4 | 0,8 | 0,8 |
| Vomer setapinnis | 1,6 | 1,1 | 1,8 | 2,2 | 0 | 0,9 | 0,2 | 0 | 0,1 |
| Dentex congoesis | 0 | 2,1 | 1,2 | 0 | 9,3 | 5,5 | 0 | 11,2 | 6,7 |
| Dentex angolensis | 0 | 14,5 | 8,6 | 0,6 | 10,8 | 6,4 | 2,0 | 49,8 | 30,4 |
| Pagellus coupel | 25,7 | 28,9 | 27,6 | 37,0 | 28,8 | 32,1 | 17,9 | 9,7 | 13,0 |
| Brotula barbata | 0,1 | 4,4 | 2,7 | 0 | 1,7 | 1,0 | 1,9 | 6,9 | 4,9 |
| Baliates carclinensie | 60,6 | 54,0 | 56,7 | 26,7 | 7,2 | 15,1 | 13,9 | 0,6 | 6,0 |
| Trichiurus lepturus | 1,6 | 0,7 | 1,1 | 1,1 | 0,1 | 0,1 | 0,8 | 0,8 | 0,5 |
| Paracibiceps | |||||||||
| lendanos | 0 | 2,8 | 1,4 | 0 | 3,6 | 2,1 | 0 | 0,8 | 0,5 |
| Cynoglossus | |||||||||
| canariensis | 4,4 | 2,8 | 3,2 | 7,9 | 1,8 | 4,8 | 6,2 | 1,8 | 8,2 |
| TOTAL TELEOSTEENS | 206 | 175 | 188 | 246 | 146 | 187 | 184 | 145 | 141 |
| TOTAL GENERAL | 225 | 188 | 200 | 265 | 148 | 195 | 157 | 156 | 156 |
| Profondeur (m) Espèce | 15 | 20 | 25 | 30 | 35 | 40 | 50 | 60 | 80 | 100 | SC/SE(1) |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Dasyatis SC | 7,3 | 5,4 | 6,2 | 1,6 | x | 1,9 | |||||
| margarita SF | 17,3 | 2,1 | 1,2 | 0,5 | 0 | ||||||
| Ilisha SC | 50,6 | 131 | 214 | 103 | 20,1 | 13,9 | 1,9 | 1,4 | |||
| africana SF | 15,8 | 40,2 | 209 | 72,8 | 0,5 | 2,2 | 0 | ||||
| Lagocephalus SC | 1,6 | 0,6 | 0,2 | 0,4 | 1,1 | 2,0 | 0,6 | 0,1 | x | 0,2 | |
| laevigatus SF | x | 0,2 | x | 0 | X | X | X | 0,1 | 0 | 0 | |
| Sphynaena spp. SC | 1,5 | 2,5 | 3,1 | 2,6 | 7,3 | 11,5 | 19,8 | 20,1 | 0,5 | X | 20,9 |
| SF | 0,8 | 0,2 | 0 | X | X | 0,2 | 1,3 | 0,4 | 0,8 | 0 | |
| Galenides SC | 23,5 | 16,4 | 23,0 | 22,2 | 15,0 | 5,5 | 1,5 | X | 3,4 | ||
| decadactylus SF | 6,0 | 7,5 | 4,9 | 5,4 | 3,3 | 3,9 | 0,1 | X | |||
| Pentanemus SC | 3,1 | 13,8 | 8,0 | 1,9 | 0,6 | 4,8 | |||||
| quinquarius SF | 2,4 | 0,7 | 3,8 | X | 0 | ||||||
| Pomadasys SC | 4,8 | 6,8 | 6,5 | 2,8 | 0,9 | 0,2 | 0,9 | ||||
| jubelinii SF | 8,8 | 11,1 | 4,8 | 3,1 | 0 | 0 | |||||
| Brachydeuterus SC | 15,9 | 37,4 | 50,6 | 128 | 74,8 | 77,9 | 100 | 50,8 | 0,5 | 8,7 | |
| typus SF | 1,5 | X | 4,9 | 0,7 | 2,1 | 19,4 | 3,0 | 0,6 | 0 | ||
| Fseudotolithus SC | 1,6 | 6,9 | 8,4 | 0,7 | 2,7 | ||||||
| typus SF | 3,3 | 1,8 | 3,5 | 0 | |||||||
| Pseudotollthus SC | 16,4 | 35,2 | 53,0 | 44,2 | 23,6 | 7,8 | 2,6 | 1,4 | |||
| senegalensis SF | 27,0 | 23,0 | 40,1 | 38,0 | 9,0 | 3,6 | 3,0 | ||||
| Pteroscion SC | 25,0 | 70,6 | 81,5 | 51,8 | 25,6 | 7,8 | 2,2 | 0 | 1,5 | ||
| peli SF | 26,5 | 48,8 | 57,1 | 29,1 | 14,9 | 4,5 | 1,1 | 0,5 | |||
| Vomer SC | 8,4 | 13,3 | 11,2 | 8,8 | 7,8 | 5,7 | 0,8 | 1,8 | 0,2 | 2,5 | |
| setapinnis SF | 0,5 | 2,6 | 5,2 | 0 | 0,1 | 0,2 | 1,0 | 7,7 | 0,3 |
| STARTE | ESPECE MOIS | SAISON CHAUDE | SAISON FROIDE | SAISON CHAUDE | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Jan | Fév | Mars | Avr | Mai | Juin | Juil | Août | Sep | Oct | Nov | Dec | ||
| 1 | Ilisha africana (i) | 143 | 151 | 92 | 65 | 85 | 65 | 59 | 86 | 109 | 104 | 111 | 130 |
| 1 | Galeoides (i) | 166 | 149 | 132 | 119 | 89 | 68 | 68 | 65 | 61 | 74 | 86 | 122 |
| decadac. (s) | (41) | (27) | (21) | (28) | (19) | (22) | (15) | (16) | (17) | (42) | (18) | (28) | |
| 1 | Pomadasys (i) | 75 | 83 | 95 | 108 | 106 | 124 | 142 | 131 | 92 | 90 | 77 | 76 |
| jubelini (s) | (24) | (31) | (26) | (35) | (22) | (32) | (29) | (34) | (20) | (52) | (36) | (25) | |
| 2 | Brachyd. (i) | 115 | 114 | 111 | 102 | 102 | 109 | 77 | 71 | 88 | 102 | 123 | 87 |
| auritus. (s) | (34) | (26) | (17) | (27) | (21) | (45) | (17) | (25) | (35) | (39) | (32) | (20) | |
| 1 | Pseudoto (i) | 150 | 122 | 104 | 103 | 79 | 69 | 70 | 81 | 75 | 93 | 127 | 128 |
| senegal. (s) | 46 | 23 | 25 | 24 | 19 | 24 | 18 | 23 | 25 | 28 | 26 | 31 | |
| ESPECE MOIS | SAISON CHAUDE | SAISON FROIDE | SAISON CHAUDE | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Jan | Fév | Mars | Avr | Mai | Juin | Juil | Août | Sep | Oct | Nov | Dec | |
| Ilisha africana (i) | 78 | 75 | 28 | 10 | 135 | 204 | 265 | 136 | 77 | 80 | 53 | 83 |
| Sphyraena spp. (i) | 218 | 90 | 57 | 26 | 26 | 211 | 203 | 59 | 34 | 50 | 27 | 216 |
| Galeoides decadacty. (i) | 332 | 120 | 151 | 6 | 18 | 6 | 0 | 11 | 17 | 90 | 245 | 185 |
| Pomadasys jubelini (i) | 299 | 156 | 97 | 1 | 1 | 14 | 0 | 41 | 48 | 143 | 233 | 146 |
| Brachydeuterus auritus (i) | 187 | 110 | 54 | 37 | 73 | 67 | 88 | 80 | 97 | 96 | 116 | 193 |
| (s) | (95) | (61) | (44) | (29) | (65) | (75) | (96) | (73) | (80) | (72) | (60) | (111) |
| Pseudotolithus senegal.(i) | 221 | 190 | 33 | 0 | 18 | 0 | 80 | 17 | 111 | 97 | 60 | 352 |
| Profondeur (m)ESPECE(1) | 15 | 20 | 25 | 30 | 35 | 40 | 50 | 60 | 80 | 100 | 200 | SC/SF |
| Muetelus SC | 0 | 0,1 | 3,5 | 0,1 | 0,8 | 0,5 | 0,3 | 0,2 | 0,1 | |||
| mustelus SF | 1,8 | 6,4 | 3,4 | 3,0 | 9,6 | 3,4 | 1,4 | 0 | 0 | 0,2 | ||
| Priacanthus SC | 0,2 | 0,5 | 5,4 | 4,2 | 5,6 | 3,3 | 0,2 | |||||
| renetus | 0,2 | x | 0,1 | 5,3 | 14,4 | 11,5 | 117,8 | 0,3 | 0,4 | |||
| Pentheroscion SC | 2,9 | 39,4 | 46,6 | 33,3 | 48,1 | |||||||
| m'biz | 7,8 | 80,8 | 38,5 | 67,1 | 189,0 | 0,7 | ||||||
| Trachurus SC | x | 0,3 | 3,2 | 0,6 | 0,7 | |||||||
| trecae SF | 0,2 | 3,8 | 29,4 | 27,4 | 7,7 | 0,1 | ||||||
| Dentex SC | 0,4 | 10,7 | 53,1 | 58,3 | 5,0 | |||||||
| angolensis SF | x | 9,8 | 89,3 | 112,8 | 7,6 | 0,6 | ||||||
| Pagellus SC | x | 0,1 | 0,2 | 0,3 | 0,9 | 2,3 | 11,7 | 5,9 | 1,8 | 0,1 | x | |
| coupel SF | 0 | 0 | 0 | x | 0,9 | 4,8 | 21,6 | 5,1 | 0,,8 | 0 | 0 | 0,7 |
| Brotula SC | x | x | 0,6 | 8,9 | 19,7 | 26,3 | 9,8 | |||||
| barbata SF | 0 | 0 | 0,9 | 18,8 | 47,5 | 58,8 | 9,5 | 0,5 | ||||
| Paracubiceps SC | 0,5 | 0,7 | 17,8 | 31,0 | 20,3 | |||||||
| ledanoisi SF | x | 0 | 6,4 | 8,9 | x | 4,6 |
| ESPECE MOIS | SIASON CHAUDE | SAISON FROIDE | SAISON CHAUDE | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Jan | Fév | Mars | Avr | Mai | Juin | Juil | Août | Sep | Oct | Nov | Dec | |
| Pageots (i) | 33 | 39 | 27 | 26 | 39 | 94 | 236 | 209 | 181 | 220 | 60 | 38 |
| 21–50m (s) | (31) | (28) | (14) | (14) | (27) | (37) | (110) | (114) | (81) | (133) | (39) | (41) |
| Pageots (i) > 50 m | 120 | 86 | 19 | 26 | 60 | 93 | 130 | 189 | 207 | 158 | 79 | 33 |
| ESPECE (1) Profondeur (m) | 15 | 20 | 25 | 30 | 35 | 40 | 50 | 60 | 80 | 100 | 200 | SC/SF |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Raja SC | 0 | 0,1 | 0,9 | 2,8 | 7,0 | 8,1 | 11,5 | 5,4 | 0,9 | |||
| miraletus SF | 1,4 | 1,4 | 5,6 | 31,4 | 11,6 | 8,1 | 7,3 | 1,9 | 0 | 0,7 | ||
| Trichiurus SC | 8,6 | 18,6 | 44,0 | 14,6 | 15,0 | 13,7 | 4,1 | 6,5 | 17,5 | 4,0 | x | |
| lepturs SF | 13,5 | 20,5 | 93,0 | 39,1 | 44,0 | 27,6 | 5,0 | 32,1 | 3,1 | 1,0 | 0 | 0,5 |
| Cynoglossus SC | 0,5 | 4,3 | 6,4 | 7,5 | 6,7 | 6,4 | 0 | x | ||||
| canrienaia SF | 0 | 1,7 | 8,3 | 9,6 | 6,9 | 6,11 | 0 | 1,0 |
| ESPECE MOIS | SAISON CHAUDE | SAISON FROIDE | SAISON CHAUDE | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Jan. | Fév. | Mars | Avr. | Mai | Juin. | Juil | Août | Sep. | Oct. | Nov. | Dec. | |
| T.lepturus | ||||||||||||
| (i) | 65 | 74 | 60 | 91 | 95 | 60 | 52 | 70 | 715 | 125 | 281 | 150 |
| chal. (s) | (41) | (28) | (46) | (46) | (71) | (31) | (24) | (27) | (54) | (102) | (134) | (50) |
| sard. (i) | 337 | 100 | 30 | 4 | 10 | 10 | 33 | 17 | 54 | 17 | 105 | 463 |
| C. Canariensis | ||||||||||||
| (i) | 88 | 76 | 94 | 98 | 96 | 123 | 153 | 142 | 129 | 79 | 55 | 69 |
| chal. (s) | (50) | (20) | (26) | (49) | (16) | (35) | (57) | (75) | (62) | (39) | (22) | (12) |
| RADIALE | Profondeur (mètres) | Oxygène (ml/l) | Poids Péché | Pourcentage par rapport à la moyenne des prises à cette profondeur |
|---|---|---|---|---|
| Grand Bassam 13 | 20 | 0,6 | 1,0 kg/h | 3% |
| (28–30/09/66) | 25 | 0,5 | 4,5 kg/h | 8 % |
| 15 | 1,1 | 0,1kg/h | 5% | |
| Grand Bassam 14 | 20 | 1,1 | 6,5 kg/h | 20 % |
| (17–19/10/66) | 25 | 1,7 | 11,0 kg/h | 21 % |
| SAISON CHAUDE | SAISON FROIDE | SAISON CHAUDE | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Jan. | Fév. | Mars | Avr. | Mai | Juin. | Juil | Août | Sep. | Oct. | Nov. | Dec. | |
| Ensamble (i) | 105 | 103 | 99 | 95 | 89 | 92 | 97 | 102 | 105 | 105 | 109 | 99 |
| du plateau (s) | (17) | (11) | (5) | (2) | (7) | (12) | (13) | (12) | (23) | (7) | (14) | (15) |
| (10–120m) | ||||||||||||
| Strate (i) | 110 | 103 | 97 | 95 | 89 | 92 | 99 | 99 | 93 | 110 | 112 | 100 |
| 10–50m (s) | (21) | (12) | (9) | (13) | (5) | (13) | (9) | (17) | (13) | (14) | (14) | (15) |
| Strate | ||||||||||||
| 50–120m (i) | 113 | 97 | 70 | 91 | 88 | 94 | 111 | 109 | 111 | 97 | 108 | 109 |
| SAISON CHAUDE | SAISON FROIDE | SAISON CHAUDE | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Jan. | Fév. | Mars | April | Mai | Juin | Juil. | Auût | Sept. | Oct. | Nov. | Déc. |
| 0,7 | 1,1 | 0,8 | 8,0 | 5,3 | 9,4 | 11,0 | 17,8 | 22,3 | 23,1 | 4,1 | 1,5 |
| CHALCT 78.01 (féurier 1978) | CHALCT 79.01 (mars 1979) | CHALCT 80.01 (août-sept.1980) | ||||
|---|---|---|---|---|---|---|
| 10–50m | 50–120m | 10–50m | 50–120m | 10–50m | 50–120m | |
| TOTAL GENERAL | 225,3 | 182,9 | 264,5 | 148,4 | 156,9 | 156,2 |
| TOTAL -Balistes | 164,7 | 128,9 | 237,8 | 141,2 | 143,0 | 155,6 |
| TOTAL -Belistes + Fritures | 122,6 | 108,2 | 178,8 | 122,5 | 133,3 | 155,6 |
| TOTAL TELEOSTEENS | 206,0 | 175,0 | 246,2 | 145,6 | 133,7 | 145,1 |
| TELEOSTEENS -Balistes | 145,4 | 121,0 | 219,5 | 138,4 | 119,8 | 144,5 |
| TELEOSTEENS -BalistestFritures | 103,3 | 100,3 | 160,5 | 109,7 | 90,1 | 144,5 |
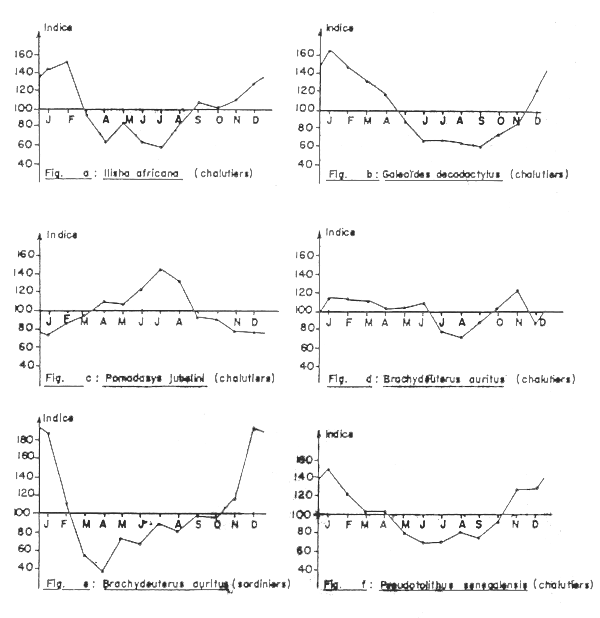
Fig. 1(a-b-c-d-e-f) - Evolution des indices mensuels moyens de rendement des chalutiers ou sardiniers en Côte d'Ivoire pour certaines des plus importantes espèces de la communauté des Sciaenidae (of.Tabl.22 et 23)
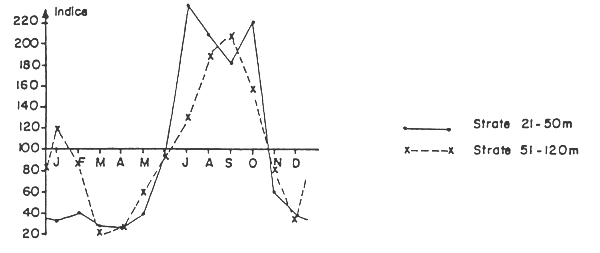
Fig. 2 Pageots (Sparidés) - Evolution des indices moyens mensuels de rendement.
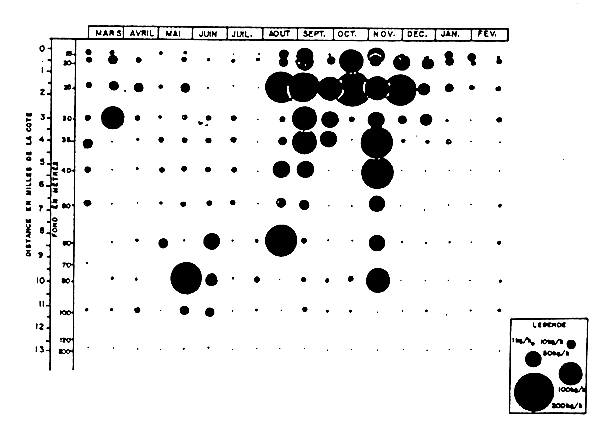
Fig. 3 Trichiurus lepturus - Rendements horaires lors des radiales Grand Bassam de 1966–67 )d'après Troadec et al., 1969)
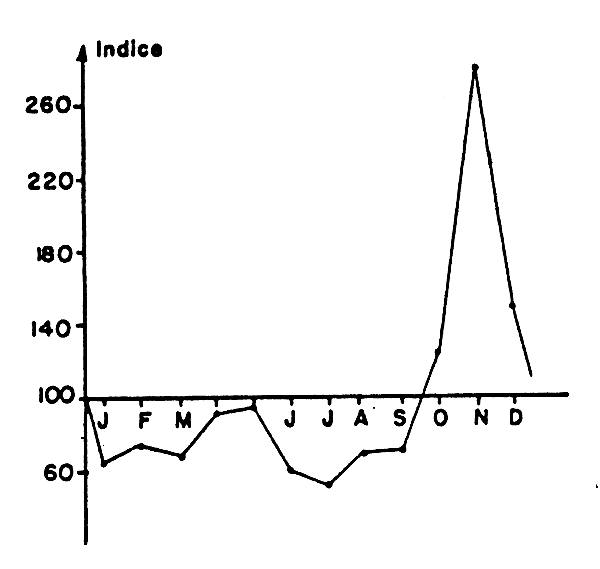
Fig. 4 Evolution des indices moyens mensuels des rendements de Trichiurus lepturus des chalutiers ivoiriens pour la strate 21–50 m.
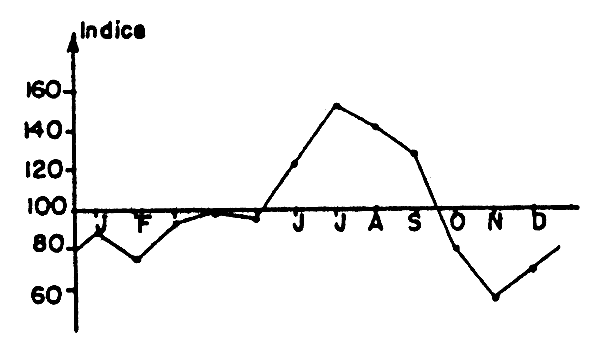
Fig. 5 Evolution des indices moyens mensuels des rendements de Cynoglossus canariensis des chalutiers ivoiriens pour la strate 21–50 m.
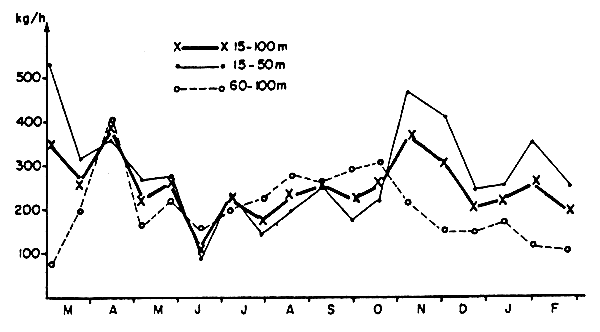
Fig. 6 Evolution des rendements horaires moyens par strate bathymétrique lors des radiales effectuées devant Grand Bassam en 1966–1967
![]()
![]()
![]()