Table 43 lists the various diseases observed during the survey. It is technically easy today to produce citrus plant material free of all known virus and virus-like diseases by the technique of shoot-tip grafting. Hence modern citrus orchards should be free of all virus and virus-like pathogens that are not spread by insect vectors (the agents of gummy bark, cachexia, scaly bark psorosis, concave gum-blind pocket, cristacortis, impietratura, exocortis, etc.). These diseases should never be seen again in newly planted citrus orchards if certified, healthy budwood is used.
TABLE 43 Summary of citrus diseases observed during survey in Pakistan (NWFP and Punjab)
| Location | Disease |
|||||||||||
DE |
GR |
TV |
EX |
GB |
CA |
BU |
YV |
SB |
WB |
PG |
CK |
|
| Tarnab Agric. Res. Inst. | + | + | + | + | + | + | + | |||||
| Khanpur Hayad Khan orchard | + | + | ||||||||||
| Sihala Perwaiz Khan orchard | + | + | ||||||||||
| Sargodha Hod. Res. Substation | + | + | + | |||||||||
| Sargodha Nasar Ullah Khan orchard | + | ? | + | + | + | |||||||
| Faisalabad Agric. Res. Station | + | + | ? | + | + | |||||||
| Sahiwal Hort. Res. Station | + | + | + | + | ? | + | + | + | + | + | ||
| Okara M. M. Shafi orchard | + | + | + | + | + | |||||||
| Renala Khurd Mitchell's fruit farm | + | + | ||||||||||
| Lahore Naizbeg Thooker | + | |||||||||||
| Lahore Fort | + | |||||||||||
Key:
DE=Decline; GR=Greening BLO; TV=Tristeza virus; EX=Exocortis;
GB=Gummy bark; CA=Cachexia; BU=Bud-union crease of sweet orange
on rough lemon; YV=Yellow vein clearing; SB=Sour orange bark
pegging and gumming; WB=Witches' broom of calamondin;
PG=Phytophthora gummosis; CK=Canker.
The exocortis viroid is mechanically transmissible and household bleach should be used, diluted to give a 5 percent sodium hypochlorite solution, to disinfect pruning tools and prevent transmission.
There is, however, a second category of citrus pathogens: those that are transmitted by insect vectors. This second category includes tristeza, greening, stubborn, vein enation-woody gall and any BLO or MLO diseases. A tree initially free from these pathogens can become infected in the nursery or the orchard if the agent and its insect vector are both present in the area. Two such pathogens are present in Pakistan: the BLO of citrus greening, transmitted by the citrus psyllid, D. citri, an insect present throughout the citrus areas of Pakistan, and CTV, transmitted by several aphid species present in Pakistan.
The greening organism is probably the cause of citrus decline in Pakistan. In India it is now well established that decline and greening are one and the same disease. Greening is a very serious disease and will continue to affect citrus orchards severely if not controlled. Most commercial citrus cultivars are susceptible to greening irrespective of rootstock. Because of this, greening should be of much greater concern to Pakistan than tristeza disease, although the survey has clearly proved the presence of CTV in Pakistan. Fortunately, T. citricida, the most efficient aphid vector of CTV, is not present in NWFP and Punjab, and most citrus orchards are grafted on rough lemon, a CTV-tolerant rootstock. One exception is in the Peshawar districts where trees grafted on sour orange are susceptible to CTV. The sour orange should therefore be replaced by a CTV-tolerant rootstock such as rough lemon. Kagzi lime trees are susceptible to CTV irrespective of rootstock. The CTV strains present in Pakistan are probably mild in comparison with some of the Asian strains of the virus. However, they may be more severe than Mediterranean strains. In any case, the distribution and the severity of the CTV strains present in Pakistan must be established.
Table 43 shows that agricultural and horticultural research stations are often good places to find evidence of virus and virus-like diseases. This is so because these stations have rich collections of citrus material obtained not only from local sources but also from foreign introductions. They have the duty to carry out rootstock experiments and they have plant pathologists on site. It has been found all over the world that rootstock experiments become "virus" experiments when old-line, non-certified citrus material is used on these rootstocks. This is the case for the sweet orange lines found to be infected with exocortis at the Sarnab, Sargodha and Sahiwal stations. Citrus cultivars of unknown health status should never be used for experimentation.
In 1975, a great number of certified virus-free varieties of sweet orange (26), grapefruit (8), mandarin (11), tangerine (5), tangelo (4) and lemon (18), as well as rootstocks (22), were imported from California to Sahiwal. Since the viroids of exocortis and cachexia are mechanically transmissible by pruning tools (scissors, knives) it is important to check that the plant material from California has not subsequently become accidentally infected.
It is not advisable to keep certified and non-certified citrus plant material side by side or close to each other. This must be kept in mind when citrus blocks or experiments are laid out in the future. Finally, a few well-kept experiments with a limited number of scion and rootstock cultivars are more fruitful than over-ambitious and unmanageable projects.
The numbers before each reference are the keys to the sources used for Table 41.
1 Ahlawat, Y.S. 1975. Leathery leaf - a new virus disease isolated from greening affected mandarin trees. Indian Phytopathol., 28: 146. (abstract)
2 Ahlawat, Y.S. 1987. Association of mycoplasma-like bodies with citrus rubberywood disease. In 3rd Regional Workshop on Mycopl., p. 12.
3 Ahlawat, Y.S. & Chenulu, V.V. 1985. Rubberywood: a hitherto unrecorded disease of citrus. Curr. Sci., 54: 580-581.
4 Ahlawat, Y.S., Chenulu, V.V., Chakraborty, N.K. & Viswanath, S.M. 1984. Occurrence of impietratura disease of citrus in India. Curr. Sci., 53: 384-385.
5 Ahlawat, Y.S., Chenulu, V.V., Gumpf, D.J. & Pandey, P.K. 1986. Some newly discovered virus and mycoplasma diseases of citrus in India. In Proc. 10th Conf: IOCV, Riverside, Univ. Calif. (abstract)
6 Ahlawat, Y.S., Chenulu, V.V., Viswanath, S.M. & Pandey, P.K. 1985. Studies on a mosaic disease of citrus in India. Curr. Sci., 54: 873-874.
7 Ahlawat, Y.S., Chenulu, V.V., Viswanath, S.M., Pandey, P.K. & Bhagabati, K.N. 1985. Mosaic disease of citrus in India. Curr. Sci., 54: 873-874.
8 Ahlawat, Y.S. & Raychaudhuri, S.P. 1988. Status of citrus tristeza and dieback diseases in India and their detection. In Citriculture. Proc. 6th Int. Citrus Cong., p. 871-879. Tel Aviv, Israel, 6-11 March 1988, Rehovot, Balaban.
9 Ahlawat, Y.S. & Sardar, K.K. 1976a. A note on the lemon crinkly leaf virus in India. Indian J. Hort., 33: 168-171.
10 Ahlawat, Y.S. & Sardar, K.K.1976b. Distribution of greening and tristeza diseases in various blocks of Darjeeling district. Sci. & Cult., 42: 275-277.
11 Anonymous. 1977a. Preliminary report on transmission of citrus huanglungbin by psylla. Guangdong Agric. Sci., 6: 50-5 1.
12 Anonymous. 1977b. Citrus psylla and citrus huanglungbin. Ganju Keji Tungxun, 3-4: 23-24.
13 Balaraman, K. & Ramakrishnan, K. 1977. Studies on strains and strain interaction in citrus tristeza virus. USA Tech. Univ. Agric. Sciences [Bangalore], 19: 62.
14 Balaraman, K. & Ramakrishnan, K. 1980. Strain variation and cross protection in citrus tristeza virus on acid lime. In Proc. 8th Conf: IOCV, p. 60-68. Riverside, Univ. Calif.
15 Bhutani, V.P., Bakhshi, J.C. & Knorr, L.C. 1972. Biochemical changes in healthy and decline sweet orange trees associated with bud-union crease. In Proc. 5th Conf: IOCV, p. 229-233. Gainesville, Univ. Fla. Press.
16 Bové, J.M., Calavan, E.C., Capoor, S.P., Cortez, R.E. & Schwarz, R.E. 1974. Influence of temperature on symptoms of California stubborn, South Africa greening, India citrus decline and Philippine leaf mottling disease. In Proc. 6th Conf. IOCV, p. 12- 15. Berkeley, Div. Agric. Sci., Univ. Calif.
17 Bové, J.M., Garnier, M., Mjeni, A.M. & Khayrallah, A. 1988. Witches' broom disease of small-fruited acid lime trees in Oman: first MLO disease of citrus. In Proc. 10th Conf: IOCV, p. 307-309. Riverside, Univ. Calif.
18 Bové, J.M. & Saglio, P. 1974. Stubborn and greening: a review, 1969- 1972. In Proc. 6th Conf. IOCV, p. 1-11. Berkeley, Div. Agric. Sci., Univ. Calif.
19 Capoor, S.P. 1963. Decline of citrus trees in India. Bull. Nat. Inst. Sci. India, 24: 48-64.
20 Capoor, S.P. 1965. Presence of seedling yellows complex in the citrus of south India. In Proc. 3rd Conf. IOCV, p. 30-35. Gainesville, Univ. Fla. Press.
21 Capoor, S.P., Rao, D.G. & Viswanath, S.M. 1967. Diaphorina citri Kuwayama, a vector of the greening disease of citrus in India. Indian J. Agric. Sci., 37: 572576.
22 Capoor, S.P., Rao, D.G. & Viswanath, S.M. 1974. Greening disease of citrus in the Deccan Trap country and its relationship with the vector, Diaphorina citri Kuwayama. In Proc. 6th Conf: IOCV, p. 43-49. Berkeley, Div. Agric. Sci', Univ. Calif.
23 Celino, C.S., Salibe, A.A. & Cortez, R.E. 1966. Diaphorina citri Kuway., the insect vector for the leaf mottle virus of citrus in the Philippines. Manila, Bureau of Plant Industry.
24 Chapot, H. 1970. Les problémes de la production des agrumes au Proche-Orient et en Afrique du Nord. UNDP/ FAO Report AT 2870. Rome, FAO.
25 Cochran, L.C. 1976. The occurrence of greening disease in Pakistan. In Proc. 7th Conf. IOCV, p. 21. Riverside, Univ. Calif.
26 Dakshinamurty, V. & Reddy, G.S. 1975. Mosaic - a transmissible disorder of sweet oranges. Indian Phytopathol., 28: 398-399.
27 Desjardins, P.R., Drake, R.J. & French, J.Y. 1969. Transmission of citrus ringspot virus to citrus and non-citrus hosts by dodder (Cuscuta subinclusa Dur. & Hilg.). Plant Dis. Rep., 53: 947-948.
28 Fawcett, H.S. & Klotz, L.J. 1939. Infectious variegation of citrus. Phytopathol., 29: 911-912.
29 Fraser, L.R. 1961. Lemon crinkly leaf virus. In Proc. 2nd Conf: IOCV, p. 205210. Gainesville, Univ. Fla. Press.
30 Fraser, L.R. & Singh, D. 1966. Greening virus, a threat to citrus industry. Indian Hort., 10: 21 -22.
31 Fraser, L.R. & Singh, D. 1968. Citrus dieback in India - the contribution of greening virus. In Proc. 4th Conf: IOCV, p. 141-144. Gainesville, Univ. Fla. Press.
32 Fraser, L.R., Singh, D., Capoor, S.P. & Nariani, T.K. 1966. Greening virus the likely cause of citrus dieback in India. FAO Plant Prot. Bull., 14: 127- 130.
33 Garnier, M., Danel, N. & Bové, J.M. 1984a. Aetiology of citrus greening disease. Ann. Inst. Pasteur/Microbiol., 135A: 169-179.
34 Garnier, M., Danel, N. & Bové, J.M. 1984b. The greening organism is a Gram-negative bacterium. In Proc. 9th Conf IOCV, p. 115-125. Riverside, Univ. Calif.
35 Gupta, O.P., Nauriyal, J.P. & Knorr, L.C. 1972. Citrus greening in the Indian Punjab. In Proc. 5th Conf: IOCV, p. 14-18. Gainesville, Univ. Fla. Press.
36 Igwegbe, E.C.K. & Calavan, E.C. 1970. Occurrence of mycoplasma-like bodies in phloem of stubborn-infected citrus seedlings. Phytopathol., 60: 1525-1526.
37 Ke, C., Chen, H. & Lin, S.C. 1980. Study on transmission of rickettsia-like organism associated with citrus yellow shoot by psylla. Fulian Agric. Sci. Tech., 10: 10-11.
38 Laflèche, D. & Bové, J.M. 1970a. Structures de type mycoplasme dans les feuilles d'orangers atteints de la maladie du greening. C.R. Acad. Sci., Ser. D, 270: 1915-1917.
39 Laflèche, D. & Bové, J.M. 1970b. Mycoplasmes dans les agrumes atteints de "Greening", de "Stubborn" ou de maladies similaires. Fruits, 25: 455-465.
40 Lallemand, J., Fos, A. & Bové, J.M. 1986. Transmission de la bactèrie associée à la forme africaine de la maladie du "Greening" par le psylle asiatique Diaphorina citri Kuwayama. Fruits, 41: 341 -343.
41 Mali, V.R., Chaudhuri, K.G. & Rama, S.D. 1975. Vein enation and woody gall diseases of citrus. FAO Plant Prot. Bull., 23: 190-191.
42 Mali, V.R., Chaudhuri, K.G. & Rama, S.D. 1976. Leaf curl virus disease of citrus in India. Sci. & Cult., 42: 525-527.
43 Martinez, A.L. & Wallace, J.M. 1967. Citrus leaf-mottle yellows disease in the Philippines and transmission of the causal virus by a psyllid: Diaphorina citri. Plant Dis. Rep., 51: 692-695.
44 Massonie, G., Garnier, M. & Bové, J.M. 1976. Transmission of India citrus decline by Trioza erytreae (Del Guercio), the vector of South Africa greening. In Proc. 7th Conf: IOCV, p. 18-20. Riverside, Univ. Calif.
45 McClean, A.P.D. 1974. Abnormal bud union between some sweet oranges and rough lemon rootstock: evidence of cause by a transmissible pathogen. In Proc. 6th Conf: IOCV, p. 203-210. Berkeley, Div. Agric. Sci., Univ. Calif.
46 McClean, A.P.D. & Oberholzer, P.C.J. 1965. Citrus psylla, a vector of the greening disease of sweet orange. S. Afr. J. Agric. Sci., 8: 297-298.
47 Nagpal, R.L. 1959. Tristeza found in Bombay state. Calif: Citrog., 44: 392, 402-405.
48 Naidu, R. & Govindu, H.C. 1981. Association of bacterium-like organism with citrus greening in India. Curr. Sci., 50: 242-244.
49 Nariani, T.K. & Raychaudhuri, S.P. 1968. Occurrence of tristeza and greening viruses in Bihar, West Bengal and Sikkim. Indian Phytopathol., 21: 343344.
50 Nariani, T.K., Raychaudhuri, S.P. & Bhalla, R.B. 1966. Citrus tristeza virus in northern and central India. Indian Phytopathol., 19: 397-399.
51 Nariani, T.K., Raychaudhuri, S.P. & Bhalla, R.B. 1967. Greening virus of citrus in India. Indian Phytopathol., 20: 146150.
52 Nariani, T.K., Raychaudhuri, S.P. & Sharma, B.C. 1960. Citrus viruses in Assam and Orissa. Indian Phytopathol., 23: 141-143.
53 Nariani, T.K., Raychaudhuri, S.P. & Sharma, B.C. 1968. Exocortis in citrus in India. Plant Dis. Rep., 52: 834.
54 Nariani, T.K., Sahambi, H.S. & Chona, B.L. 1965. Occurrence of tristeza virus in citrus in northern India. Indian Phytopathol., 18: 220.
55 Nariani, T.K., Viswanath, S.M. & Menon, M.R. 1970. Occurrence of tristeza and greening diseases of citrus in Kerala. Agric. Res. J. Kerala, 8: 123-124.
56 Nour-Eldin, F. 1959. Citrus virus disease research in Egypt. In J.M. Wallace, ed. Citrus virus diseases, p. 219-227. Riverside, Univ. Calif.
57 Oberholzer, P.C.J. 1959. Host reactions of citrus to tristeza virus in South Africa. In J.M. Wallace, ed. Citrus virus diseases, p. 35-43. Riverside, Univ. Calif.
58 Patil, B.P. & Warke, D.C. 1968. A note on existence of exocortis virus in India. Curr. Sci., 37: 469-470.
59 Raychaudhuri, S.P., Nariani, T.K. & Ahlawat, Y.S. 1977. Dieback of citrus in India. Proc. Int. Soc. Citricult., 3: 914918.
60 Raychaudburi, S.P., Nariani, T.K., Lele, V.C. & Singh, G.R. 1972. Greening and citrus decline in India. In Proc. 5th Conf: IOCV, p. 35-37. Gainesville, Univ. Fla. Press.
61 Reddy, G.S. 1965. Citrus decline in south India. In Proc. 3rd Conf: IOCV, p. 225. Gainesville, Univ. Fla. Press.
62 Reddy, G.S. & Govinda Rao, P. 1961. Is there tristeza in Andhra Pradesh, India? In Proc. 2nd Conf: IOCV, p. 132-135. Gainesville, Univ. Fla. Press.
63 Reichert, I. & Perlberger, J. 1934. Xyloporosis, the new citrus disease. Agric. Exp. Sta., Rehoboth, Palestine, Bull., 12: 49.
64 Roistacher, C.N. 1988. The cachexia and xyloporosis diseases of citrus - a review. In Proc. 10th Conf: IOCV, p. 116-124. Riverside, Univ. Calif.
65 Saglio, P., Laflèche, D., Bonissol, C. & Bové, J.M. 1971 a. Isolement et culture in vitro des mycoplasmes associés au "stubborn" des agrumes et leur observation au microscope électronique. C.R. Acad. Sci. Paris, 272: 1387-1390.
66 Saglio, P., Laflèche, D., Bonissol, C. & Bové, J.M. 1971b. Isolement, culture et observation au microscope électronique des structures de type mycoplasme associées à la maladie du stubborn des agrumes et leur comparaison avec des structures observées dans le cas de la maladie du greening des agrumes. Physiol. Veg, 9: 569-582.
67 Schwarz, R.E. 1970. A multiple sprouting disease of citrus. Plant Dis. Rep., 54: 1003-1007.
68 Varma, P.M., Rao, D.G. & Capoor, S.P. 1965. Transmission of tristeza virus by Aphis craccivora (Koch) and Dactynotus jaceae L. Indian J. Entomol., 27: 67-71.
69 Varma, P.M., Rao, D.G. & Vasudeva, R.S. 1960. Additional vector of tristeza disease of citrus in India. Curr. Sci., 29: 359.
70 Vasudeva, R.S. & Capoor, S.P. 1958. Citrus decline in Bombay State. FAO Plant Prot. Bull., 6: 91-92.
71 Vasudeva, R.S., Varma, P.M. & Rao, D.G. 1959. Transmission of citrus decline virus by Toxoptera citricida (Kirk) in India. Curr. Sci., 28: 418-419.
72 Wallace, J.M. & Drake, R.J. 1968. Citrange stunt and ringspot, two previously undescribed virus diseases of citrus. In Proc. 4th Conf: IOCV, p. 177-183. Gainesville, Univ. Fla. Press.
73 Weathers, L.C. 1960. Yellow-vein disease of citrus and studies of interactions between yellow-vein and other viruses of citrus. Virology, I 1: 753-764.
74 Weathers, L.G. 1961. Responses of citrus to concurrent infection with two or more unrelated viruses. In Proc. 2nd Conf: IOCV, p. 187- 196. Gainesville, Univ. Fla. Press.
75 Xu, C.F., Li, K.B. & Liao, J.Z. 1985. On the transmission of citrus yellow shoot by psylla and observation with electron microscopy. Acta Phytopathologica Sinica, 15(4): 241-245.
76 Yora, K.Y., Doi, Y., Raychaudhuri, S.P. & Ahlawat, Y.S. 1977. Infectious variegation in India. Calif: Citrog., 62: 342.
Greening disease and its psyllid vectors
Other virus and virus-like diseases
Phytophthora gummosisa-fungal-disease
Bacterial disease: Citrus canker
Bibliography
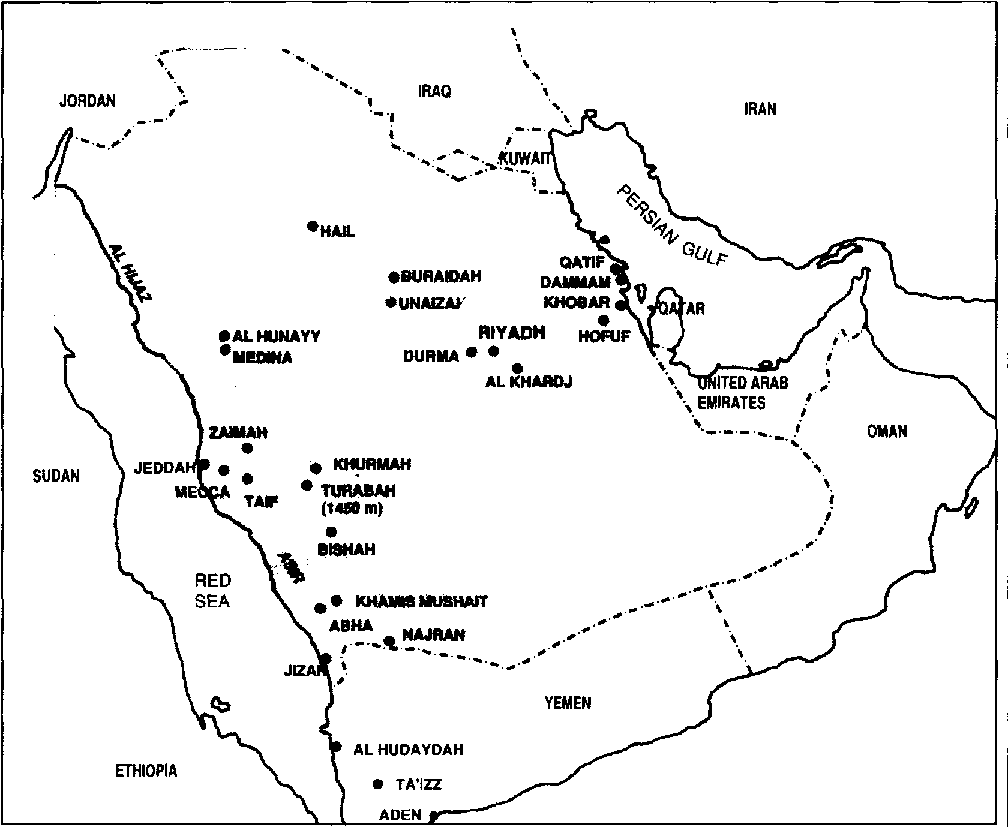
Traditionally, agriculture in Saudi Arabia has been centred around oases and wadi channels scattered throughout the Kingdom where springs and shallowground water are available, or where rainfall alone is sufficient for the cultivation of crops such as wheat, barley, millet and some vegetables and fruit (for example, in the Asir region where the annual rainfall exceeds 300 mm).
Citrus is grown essentially in the highland oases of the Asir, from Taif to Najran, and in the Buraidah-Unaizah region (see Map 9).
Mandarin and sweet orange trees, mainly on sour orange rootstock, used to grow well in the Asir oases but, in the 1970s, they began to decline, and by the early 1980s they had been wiped out. The major result of the survey for virus and virus-like diseases in Saudi Arabia has been the discovery that citrus greening disease was the cause of this decline.
Today the less susceptible small-fruited acid lime (Citrus aurantifolia) is the only cultivar still grown. The distribution of the disease as well as of its insect vector, the psyllid Diaphorina citri, was determined by three surveys between November 1981 and December l 983. Greening and its psyllid vector were not encountered in the Buraidah-Unaizah region surveyed in 1981.
In the past, citrus has been introduced into Saudi Arabia from several countries, including Egypt, Jordan, Lebanon and Yemen. These introductions were not certified free of diseases, hence, it was to be expected that the graft-transmissible diseases affecting citrus in these countries would be present in Saudi Arabia. The survey has indeed shown this to be the case. Greening, however, was not introduced from the Mediterranean countries, which are still free of it.
No records are kept by farmers as to the origin of the trees, the nature of the rootstocks, or the scion varieties. This has often made it difficult to identify the diseases accurately. Sometimes, ironically, the type of symptoms present on rootstocks has led to identification of the rootstocks.
Greening disease and its psyllid vectors
Diagnosis
Diagnosis of greening was based on symptom expression and electron microscopical detection of the greening organism in the sieve tubes of leaves and/or fruits.
Leaves and fruits were kept in plastic bags for a few hours until they could be further processed. The leaf midrib was cut out with a razor blade and chopped into pieces 2 to 4 mm long. About five such pieces were fixed by immersion in a 5 ml screwcap tube filled with 2 percent glutaraldehyde in 0.1 M phosphate buffer at pH 7.4. For a given tree, midrib pieces from five to ten leaves were used and placed in the same tube.
The peduncular end of the fruit axis is rich in phloem tissue and represents choice material for the detection of the greening organism. The columella tissue was dissected from the fruit with a razor blade, chopped into 2 to 4 mm-long fragments and fixed in the same way as the leaf pieces.
The samples remained in the 2 percent glutaraldehyde solution for two to four weeks until they reached the Bordeaux laboratory, where they were post-fixed in I percent osmium tetroxide in the same phosphate buffer. After dehydration in ethanol, specimens were embedded in Epon and thin-sectioned. The ultra-thin sections were examined in a Siemens Elmiscope 101 electron microscope (Bové and Garnier, 1984).
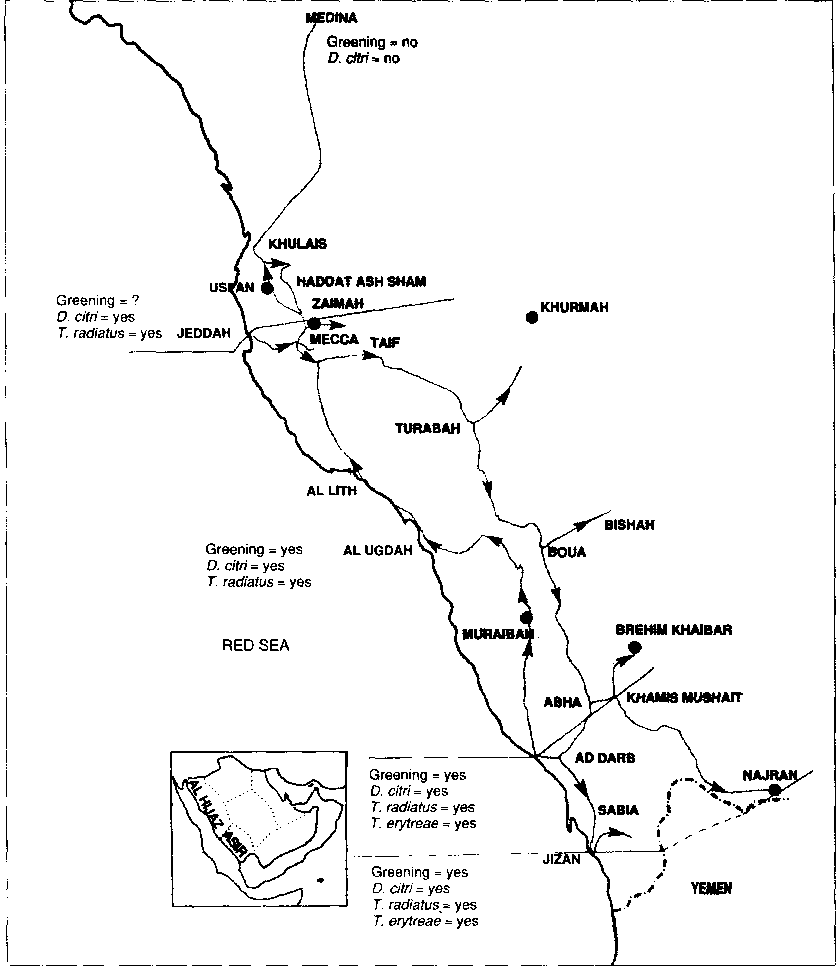
Areas where the Asian form of greening and the Asian psyllid vector, Diaphorina citri, are present: Asir
Clear-cut symptoms of greening disease (Figs 196 to 214) were observed in the highland oases of the Asir (see Map 9 and Table 44), where the Asian vector of the disease, D. citri, was also present. The greening organism was detected by electron microscopy in affected citrus material (Figs 7 to 18). These studies prove that the symptoms observed are indeed those of greening.
The areas concerned are:
In these three areas, practically the only citrus species now grown is the small-fruited acid lime. According to citrus growers, sweet orange and mandarin trees used to do well, but have been wiped out within the last 15 years. Greening disease is very probably responsible for their destruction. Indeed, severe symptoms of greening were observed on moribund sweet orange (Fig. 197) and mandarin trees (Figs 196, 199 to 206) and electron microscopy showed the presence of the greening organism in these trees.
Lime has proved to be less susceptible to greening than sweet orange and mandarin. In affected areas, lime trees display greening symptoms (leaf mottle) (Fig. 214) but, although electron microscopy shows them to be infected by the greening organism, they achieve a respectable size (Fig. 206). An additional reason for relatively good growth is the fact that limes are not affected by citrus tristeza disease. This is a remarkable situation since, in Africa and Asia, greening is generally accompanied by tristeza, and acid lime is highly susceptible to this virus. Hence, in areas where both tristeza and greening are present, the lime trees decline not so much because of greening, but because of tristeza.
While in some areas only a few small lime orchards exist, in other areas, such as Turabah, Khurmah and Bishah, lime orchards cover many hectares and total thousands of relatively well-developed trees. In fact, the only adult, ten- to 15-year-old trees that can be seen in commercial orchards are lime.
MAP 10 Area covered by the 1983 survey for greening and its vectors in Saudi Arabia
TABLE 44 Greening. psyllids and psyllid parasites and citrus canker in Saudi Arabia as of 1983
In the above-mentioned areas, a high percentage of trees are affected. The reason for this high incidence of infection is the presence of the Asian vector of greening, D. citri (Figs 37 and 210).
This psylla was first observed in 1971 on citrus in the Jeddah, Wadi Khulais, Mecca and Haddat Ash Sham areas (FAO, 1972; Wooler, Badgham and Arafat, 1973). Although it has been known since 1966 that D. citri is a vector of greening (Capoor, Rao and Viswanath, 1967; Celino, Salibe and Cortez, 1966), no thought was given until 1981 to the possibility that the disease transmitted by this insect vector might be present in the areas where the vector occurred in Saudi Arabia.
In November 1981, Bové was struck by the magnitude of D. citri populations (Fig. 211). On lime trees the psyllas were so numerous in the areas visited (Taif and Turabah) and the damage so great (Figs 211 to 213), that it was almost impossible to realize that one was looking at lime trees.
The Agricultural Services had been aware of the problems since 1972 and had tried to control the psyllas with the use of various insecticides. From the situation as seen by Bové in 1981, it seems that these treatments were totally unsuccessful.
The primary eulophid hymenopterous parasite (ectoparasite) of D. citri is Tetrastichus radiatus Waterston. An ectoparasite identical to T. radiatus was collected in December 1983 by Aubert and Bové from parasitized nymphs of D. citri at Wadi Nahman and Turabah. Parasitized nymphs of D. citri were seen at Zaimah, but no T. radiatus individuals were obtained in the emergence boxes. It is, however, most probable that the ectoparasite is present. More sampling is needed. The Bishah area probably also harbours T. radiatus.
The strain of the Tetrastichus sp. ectoparasite obtained from parasitized nymphs of D. citri in Saudi Arabia was found by Panis (Valborne, France) to be morphologically indistinguishable from the species T. radiatus. However, Prinsloo (Pretoria, South Africa) found the Saudi Tetrastichus to be somewhat different from the strain of T. radiatus introduced from India (Bathinda area, Punjab) into Reunion, where it was used successfully to control D. citri. Further work is required to characterize the Saudi T. radiatus ectoparasite fully.
For the following reasons the form of greening present in the above areas is most probably the Asian form.
Areas where greening and the psyllid vectors, D. citri and Trioza erytreae, are present: Brehim Khaibar
In the oasis of Brehim Khaibar in the Abha-Khamis Mushait area (see Map 10), the two psyllid vectors of greening, D. citri and Trioza erytreae, both occur in the same orchards. Detection of T. erytreae was based on the presence of galls produced on the leaves by nesting nymphs (Fig. 207). These galls are very typical and bear witness to the presence of T. erytreae. Only empty galls were seen, indicating a normal emergence of adults, quickly followed by the shedding of the moult.
When psyllid parasites are present, punctured mummies remain attached inside the galls for ten months or more. Hence, the absence of punctured mummies may indicate that in the affected area T. erytreae is not parasitized, or at least not to a great extent. Obviously, more observations must be made over a longer period.
On the other hand, D. citri nymphs were parasitized and T. radiatus was obtained in the emergence box. Severe greening symptoms were observed on the only two moribund mandarin trees present and on the many acid lime trees (Fig. 206). The presence of the greening organism was confirmed by electron microscopy (Figs 17 and 18). The form of greening involved is probably the Asian form even though the African form may also have been introduced with T. erytreae.
Areas where the African form of greening and the African psyllid vector, T. erytreae, are present: Fayfa
Many T. erytreae-induced galls were seen on sweet orange leaves in the mountainous Fayfa area (altitude I 200 to 1 800 m) near the border with the northern part of Yemen, east of Jizan (see Map 10). Only one punctured mummy was seen in a gall, suggesting a low level of parasitism. D. citri was not observed.
Symptoms of zinc deficiency were severe on many trees (Figs 208 and 209), but the trees had not yet developed the typical "greening" aspect. However, the greening organism was detected by electron microscopy in sweet orange tissues, even though in very low numbers. In the absence of D. citri, it is highly likely that the form of greening involved is the African, and that the disease is in its early stages.
Areas where the African psyllid vector, T. erytreae, occurs, apparently in the absence of greening: Najran
While no evidence for the presence of T. erytreae was found during the November 1981 and December 1983 surveys in the Najran area (see Map 10), such evidence was obtained in January 1984 by the National Centre for Horticultural Research and Development in Najran. T. erytreae galls were observed in three different orchards on acid lime, lemon and mandarin leaves. The galls contained punctured mummies indicating parasitism. However, no symptoms of greening were observed.
Even though T. erytreae has definitely been found in Wadi Najran, the psylla populations seemed to be low and in a state of decline - at least in the period January to March 1984. The climatic conditions might not be favourable enough for high populations of T. erytreae to become established.
Eggs and young nymphs die when the temperature exceeds 32°C for several hours a day, especially when the relative humidity is low. Green and Catling (1971) have combined the mean daily maximum temperature and the mean minimum vapour pressure to characterize a "lethal day" for T. erytreae by the saturation deficit (SD) in millibars, given by the equation
SD = VPt°max(100 - RHmin)/100
where VPt°max is the saturation vapour pressure corresponding to the maximum temperature recorded during the day, and RHmin is the minimum relative humidity.
On a lethal day, the SD exceeds 35 mbar and causes 70 percent mortality of eggs and first instar nymphs. Mortality is 100 percent when the SD is higher than 48 mbar. These conditions occur in Najran from April to September, as shown in Table 45. The same is true for Bishah and Medina. However, at Khamis Mushait, no such conditions occur and this explains why nearby, at Brehim Khaibar, T. erytreae is present. Even at Taif, conditions might support the development of T. erytreae.
Areas where the Asian psyllid, D. citri, occurs, apparently in the absence of greening
Besides the areas mentioned above, D. citri is also present north of Jeddah and was seen at Haddat Ash Sham, Usfan and Khulais (see Map 11). The presence of the ectoparasite T. radiatus var. saudi and parasitized D. citri nymphs was also established in these areas (see Table 44). Furthermore, the encyrtid hyperparasite Cheiloneurus cyanonotus Waterston was obtained in the emergence box containing the leaf samples of a lime from Khulais.
This hyperparasite was the only one that was obtained during the December 1983 survey. This does not mean that hyperparasites are absent from other areas. More observations over longer periods are required.
No clear-cut symptoms of greening were seen in the area north of Jeddah and the greening organism was not detected by electron microscopy in the samples studied. The areas concerned are at low altitudes, 150-200 m, and it is possible that the very high temperatures that prevail in these areas in the summer are detrimental to the organism. Further study is required to confirm that greening is really absent in these coastal areas.
Areas tree of greening and psyllid vectors
Citrus in the following areas was found to be free of greening and psyllid vectors in 1983 (see Maps 9 and 10):
Summary and conclusions
{kind=link}
{kind=link}
{kind=link}