![]()
![]()
![]()
El herbívoro juega un papel muy importante dentro del ecosistema del pastizal. Al consumir el forraje existente promueve un mayor crecimiento de las especies que conlleva a una mayor productividad de la sabana inundable.
El herbívoro es el reciclador de nitrógeno más efficiente de todos los animales, ya que en cuestión de horas, a través de la orina principalmente, hace soluble muy importantes cantidades de nitrógeno, que regresan de nuevo al pastizal y mantienen su fertilidad. Las grandes extensiones de sabanas tropicales y pastizales de los altiplanos y climas templados no existirían sin la presencia de los herbívoros.
Los carbohidratos estructurales de las plantas (celulosa, hemicelulosa, etc.) constituyen la fuente más abundante de la producciuón primaria de los ecosistemas. Los animales superiores no pueden utilizar estos compuestos directamente, debido a que no disponen de las enzimas requeridas para su hidrólisis o descomposición, y solamente mediante su asociación con bacterias que viven en su tracto digestivo es que los pueden utilizar. Todos los animales probablemente mantienen microorganismos en su tracto digestivo en una relación simbiótica provechosa. El beneficio para el animal depende de la estructura del tracto digestivo y de la naturaleza del alimento consumido (Parra, 1977).
La dentición de las diferentes especies animales varía de acuerdo a los hábitos alimenticios y otras funciones que tienen que desempeñar.
El capibara, al igual que otros roedores, se caracteriza por posser cuatro incisivos muy desarrollados y la ausencia de caninos. Posee a cada lado cuatro dientes molariformes. La fórmula dentaria es la siguiente:
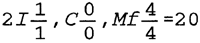
Los incisivos, de crecimiento continuo, son fuertes, largos y afilados, y cada par está íntimamente unido. El ancho del corte es de 2 a 3 cm (Escobar y González-Jiménez, 1971).
Entre los incisivos y el primer alveolo molar existe un espacio desprovisto de dientes (el diastema o espacio interalveolar), de 2 cm de ancho en el adulto. Los dientes molariformes se presentan en número de 4 en cada lado de las arcadas dentarias. Se componen de prismas transversales en el caso del último molar superior, que es tan largo como los tres molariformes anteriores juntos. Las caras masticatorias son aplanadas y unen a los prismas en toda su extensión y van dispuestos en plano inclinado con pendiente negativa.
Por este carácter, los molariformes superiores e interiores realizan un contacto muy íntimo de las caras masticatorias.
La articulación mandibular es muy peculiar; además de articular con el temporal lo hace también con el hueso molar. Las caras articulares del temporal y del molar forman un canal en el cual se desplaza con mucha libertad en el sentido horizontal el cóndilo de la mandibula, lo que permite la predominancia de los movimientos de propulsión y retropropulsión por el gran desarrollo de los músculos masticatorios: digástrico y maseteros. Esto permite a la articulación mandibular realizar los largos movimientos anteroposteriores de masticación, molienda, desmenuzamiento y deglución que realiza este animal (Mendoza, 1977).
La molienda es muy efectiva por su potente capacidad de masticación, reduciendo los forrajes a partículas finas de 0, 001 a 0,3 mm2, Ojasti (1973), contribuyendo a la eficiencia digestiva de este roedor herbívoro. Baldizan et al. (1983) estudiaron este aspecto y reportaron que el 46, 5% de las particulas de alimento en el estómago tenían un tamaño menor a 0,15 mm2, demostrando su capacidad masticatoria. Este desmenuzamiento tan efectivo permite una mayor eficiencia en la extracción de substancias solubles y en el ataque microbiano.
En la lengua predominan de glándulas serosas (de Von Ebner), cuya finalidad es la de diluir las sustancias que llegan al surco circunvalador de las papilas donde se encuentran las cápsulas gustativas. Esto según Leal-Medina (1978) puede ser una de las razones por las cuales el capibara es un animal de palatabilidad selectiva. Tal vez dicha secreción sirva también para ayudar a la masticación tan minuciosa de este animal, la cual también sería ayudada por las papilas filiformes, cuya estructura está formada casi exclusivamente por la capa cornea. El capibara tiene una elevada producción salival que se explica por ser el parenquima de la parótida rico en acinis y numerosos conductos excretores.
El esófago tiene un epitelio cornificado. Esto obedece a que la mucosa tiende a protegerse del paso de los alimentos toscos como son los forrajes y más aún cuando están secos. Este órgano no presenta glándulas ni siquiera en la submucosa. Así que su lubricación, la cual normalmente debe existir para el deslizamiento del bolo alimenticio, estaría asegurada por la excesiva secreción de las glándulas salivales y linguales ya mencionadas.
En el estómago se observan las regiones que caracterizan los estómagos de los monogástricos, donde la porción cardial es muy pequeña, la fúndica muy desarrollada y la pilórica un poco mayor que la cardial.
Llama poderosamente la atención la gran cantidad de células parietales en las glándulas fúndicas, lo cual indica una gran producción de ácido clorhídrico. Este ácido, como ha sido demonstrado, actúa como agente bacteriostático y también dando actividad al pepsinógeno para transformarlo en pepsina.
El intestino delgado presenta en la región del duodeno vellosidades cortas muy parecidas a las de los rumiantes.
Las vellosidades conniventes son muy desarrolladas, indicio de un alto poder de absorción, al igual que ocurre en los rumiantes. Las glándulas de Bruner no existen ni en el duodeno ni en el yeyuno. Como las células de estas glándulas son de tipo mucoso, podría ser que su secreción sea sustituida por la excesiva producción de mucus en el intestino grueso por parte de las glándulas de Lieberkuhn (Leal-Medina, 1978).
El yeyuno se presenta formado por vellosidades sumamente cortas que contrastan con las de los demás mamíferos. En apariencia la capacidad de absorción se vería compensada por su grán número y por el desarrollo de las glándulas conniventes del duodeno.
El intestino grueso es semejante al de los otros mamíferos, con la diferencia de que el ciego presenta pliegues, es allí donde se efectúa la absorción del agua y todos aquellos componentes hidrolizados tal como sucede en los otros roedores y herbívoros con fermentación bacteriana. Por último, en las observaciones al microscopio electrónico realizadas por Leal-Medina (1978) encontró que las glándilas fúndicas de este animal son muy parecidas a las de los otros animales, presentando sólo pequeñas diferencias en cuanto a la unión con las células vecinas, donde predominan las interdigitaciones.
El sistema digestivo en términos de tamaño, capacidad y volumen fue estudiado por Parra y González-Jiménez (1970). Sus pesos (Cuadro 4) muestran que el ciego es el mayor de los órganos de este roedor (37.7%), constituyendo los intestinos delgado y grueso casi el 70% de todo el tracto. Fernandez (1981) en sus estudios anatómicos sobre el abdomen de esta especie hace un analisis topográfico de estos y estudia su irrigación. Mendoza (1984) estudia ademas de la topografía de organos los trayectos vasculonerviosos.
CUADRO 4
Peso de los órganos del tracto digestivo
| Peso (g) | % del peso vivo | % del peso del tracto digestivo total | |
| Estomago | 223±33 | 0,55 | 10,4 |
| Intestino delgado | 650±152 | 1,61 | 30,7 |
| Ciego | 798±132 | 1,97 | 37,7 |
| Colon | 273±97 | 0,68 | 12,9 |
| Recto | 175±43 | 0,43 | 8,3 |
| Total | 2188±37* | 5,24 | 100,0 |
* Promedio ± desviación estandar.
* Peso vivo promedio: 40,5±78
Fuente: Parra y González-Jiménez (1972).
En la figura 2, se pueden ver las longitudes de los componentes del tracto. Si se compara su longitud relativa con otros herbívoros se nota cuán importante es el ciego en esta especie. Esto se evidencia cuando se pesó el contenidos del tracto digestivo del capibara (Cuadro 5), de donde se concluye que el 74% de la ingesta total de este animal se encuentra en el ciego.
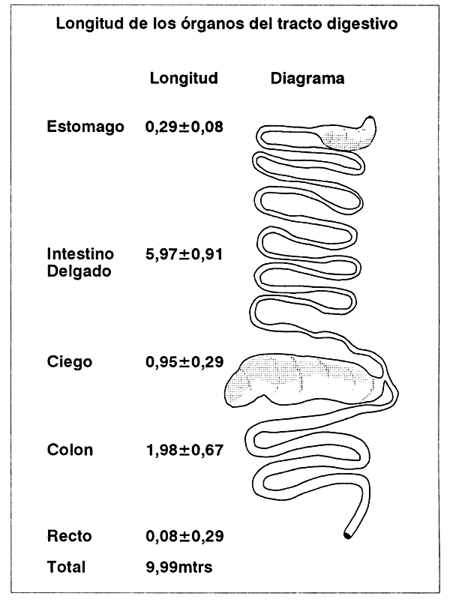
Fuente: Para y González Jiménez (1972).
CUADRO 5
Peso de los contenidos del tracto digestivo
del capibara (g)
| Ingesta fresca* | Materia Seca | en % del total (MS) | |
| Estómago | 747 ± 275 | 113 ± 58 | 9,6 |
| Intestino delgado | 403 ± 250 | 38 ± 16 | 3,2 |
| Ciego | 4746 ± 1027 | 869 ± 274 | 74,0 |
| Colon | 386 ± 95 | 78 ± 31 | 6,6 |
| Recto | 231 ± 138 | 76 ± 59 | 6,5 |
| Total | 6672 ± 1391* | 1175 ± 315 | 100,0 |
* promedio ± desviación estandar.
Fuente: Parra y González-Jiménez (1972).
Cuando se compara con otros animales la capacidad relativa de los diferentes comportamientos del tracto digestivo (Cuadro 6), definitivamente se concluye que el capibara es un herbívoro de digestión cecal y es el monogástrico que presenta la mayor capacidad relativa para este órgano.
CUADRO 6
Comparación del volumen gastrointestinal del capibara con otras especies
| Especies | Contenido total | Retículo Rumen | Estómago (abomaso) | I.D.* | Ciego | C+R** |
| (en % del peso corporal) | ||||||
| Vacuno | 13–18 | 9–13 | 0.5 | 0.8 | 0.8 | 0.8–1.5 |
| Bovino | 12–19 | 9–13 | 0.7–1.6 | 0.9 | 0.9–1.6 | 0.5–0.7 |
| Capibara | 16–25 | - | 1.8 | 1.0 | 11.0 | 1.3 |
| Conejo | 7–8 | - | 2–7 | 0.6–1.8 | 2.5–7.8 | 0.7–1.3 |
* Intestino delgado
** Colon y recto
Los herbívoros pueden ser clasificados en dos grandes categorías de acuerdo a la fisiología digestiva y al órgano principal de la digestión microbial (Parra, 1977). Los rumiantes tienen un comportamiento pregástrico muy desarrollado donde se efectúa la digestión microbial, es decir, donde esta digestión microbial precede a la gástrica. El otro grupo lo constituyen aquellos animales con un compartimiento postgástrico grande en los cuales la digestión gástrica e intestinal ocurre antes de la fermentación microbial, donde ubican un gran número de animales como el capibara, los equinos, los conejos y los elefantes entre otros.
La característica fundamental del herbívoro es la gran capacidad del tracto digestivo para almacenar alimentos si se compara con los omnívoros y carnívoros.
Esto se debe a la dieta tan voluminosa que ingieren y a la necesidad de mantener por largo tiempo la digesta, para que el lento proceso fermentativo de la digestión bacterial pueda realizarse. Parra (1977) mostró que para rumiantes y no rumiantes (herbívoros monogástricos) cuando aumenta el tamaño corporal aumenta proporcionalmente la capacidad del tracto digestivo (como % del peso vivo del animal).
En el caso del capibara cuando se le compara con el rumiante, los compartimientos fermentativos (como el ciego) son tan importantes relativamente como el rumen de los herbívoros con digestión microbial pregástrica.
La velocidad de paso es otro importante componente de la fisiología digestiva a considerar en los herbívoros. Los componentes fibrosos (celulosa, lignocelulosa, hemicelulosa, etc.) son muy refractarios a la digestión, de allí que sea la fermentación un proceso lento. Esta digestión microbial requiere de la retención de los alimentos en los órganos o depósitos fermentativos como el omaso, ciego, colon, etc. del herbívoro. De allí que la velocidad de paso de los alimentos sea mucho más lenta en los herbívoros que en los omnívoros y los carnívoros. Blaxter (1962) claramente define que la utilización mayor se presenta cuando los alimentos tienen suficiente tiempo de digestión en los comportamientos donde se efectúa la digestión microbial. Es evidente que el efecto de la tasa de paso sobre la digestibilidad de los alimentos mayormente se explica por el lento proceso de digestión de los componentes fibrosos. Es por eso que el grado de desmenuzamiento del bolo alimenticio es tan importante para la digestión.
El proceso digestivo en los rumiantes, tal y como su nombre lo indica, consiste en desmenuzar el alimento fibroso mediante la rumia que no es más que una masticación sucesiva mediante regurgitación del bolo alimenticio, y la de los herbívoros no rumiantes mediante un fino molido a través de una eficiente molienda en la boca, este es el caso del capibara. Algunos comentarios de observadores del comportamiento alimentario del capibara sugieren que esta práctica puede ser otra causa de la eficiente molienda que hace del forraje.
Si se compara la velocidad de paso de la ingesta en el tracto digestivo del capibara con otras especies (mediante el tiempo para la excreción del 5% y del 95% del marcador, ver Cuadro 7), se constata que el tránsito de los alimentos disminuye del herbívoro al carnívoro, siendo intermedio el omnívoro. Los herbívoros monogástricos tienen velocidad de paso mayor que los rumiantes. Esto podría explicarse por la molienda muy eficiente que debe ejecutar el monogástrico para hacer buen uso del alimento, Escobar y González Jiménez (1972).
CUADRO 7
Velocidad de paso de la ingesta (horas) para la excreción
del 5% y el 95% del marcador.
| Animal | para 5% | horas | 95% |
| Rumiantes | |||
Bovino | 20 | 120 | |
Ovino | 18 | 93 | |
| Monogástricos Herbívoros | |||
Caballo | 15 | 60 | |
Elefante | 22 | 60 | |
Capibara | 8 | 84* | |
Capibara | 12 | 88** | |
Conejo | 5 | 50 | |
| Omnívoros | |||
Cerdo | 10 | 40 | |
Gallina | 2 | 14 | |
Rata | 4 | 24 | |
| Carnívoro | |||
Zorro | 3 | 12 |
Fuente: Parra (1978)
* Calculado a partir de los datos de Ojasti (1973).
** Determinados por González-Jiménez, et al. (1976).
Los procesos microbiológicos de los reservorios digestivos dependen de las condiciones fisiólogicas de éstos: pH, temperatura, capacidad tampón, grado de anaerobiosis y electrolitos presentes.
El capibara mantiene en el estómago un pH óptimo de 3,5 (cuadro 8), indicativo de una digestión ácida; para luego llegar a la neutralidad en el intestino delgado y demás compartimientos. Es evidente que el ciego corresponde al componente más importante del sistema digestivo del capibara debido a su capacidad y pH y es en este órgano donde se desarrolla un activo proceso de fermentación en el que son degradados los carbohidratos estructurales (celulosa y hemicelulosa) que componen la mayor parte de los alimentos que ingiere. Al mismo tiempo, juega un papel muy importante en la economía del nitrógeno de este animal y en la síntesis de proteína microbial y sustancias hidrosolubles necesarias en su nutrición (González-Jiménez y parra, 1971).
CUADRO 8
pH DEL CONTENIDO DEL TRACTO DIGESTIVO DEL CAPIBARA
| Forrajes+Concentrados (promedio±D.E.) | Baldizan et al. (1983) | ||
| Forrajes | Concentrados | ||
| Estomago | 3,50 ± 0,29 | 2,5 | 3,4 |
| Intestino Delgado | 6,86 ± 0,22 | ||
| Ciego | 6,03 ± 0,29 | 6,6 | 6,3 |
| Colon | 6,32 ± 0,17 | 7,0 | 6,5 |
| Recto | 6,73 ± 0,32 | ||
La fermentación microbiana depende fundamentalmente de las condiciones del reservorio fermentativo, óptimas en el capibara. Por otro lado se tiene la población microbial. Existen muchas investigaciones sobre las poblaciones microbiales de los rumiantes y sin embargo pocas en herbívoros silvestres. El trabajo de McLure (1970) sobre los ciliados del ciego del capibara demostró que existe una población abundante y heterogénea. Sin embargo, los primeros trabajos sobre estos microorganismos fueron hechos por Da Cunha (1915, 1917 y 1938), Da Fonseca (1939) y recientemente por Hollande y Batisse (1959) y Batisse (1965, 1966). Los trabajos mencionados comprenden 19 especies de ciliados, de las cuales 17 se han descrito únicamente en el capibara.
Sin embargo se puede decir que todos los herbívoros albergan poblaciones muy similares de microbios en sus reservorios de fermentación y en cantidades muy parecidas, pues esta población está mayormente definida por la dieta que ingiere tanto en número como en composición de los microorganismos que la integran (Parra 1977), ya que la tasa muy elevada de recambio de esta inmensa población microbial hace que tenga un gran potencial de cambio y adaptación al sustrato que digiere. Baldizan (1983) identificó una activa fermentación en el ciego, pero de intensidad menor que la del rumen.
Se puede decir que los productos finales de esta dígestión en proporción de ácido acético, ácido butírico y ácido propiónico, para sustratos de forrajes y alimento concentrado, es muy similar a la de los rumiantes.
Pero la concentración de estos ácidos grasos de cadena corta en el ciego y el colon (41–44 mM) (ver cuadro 9), fueron menores que las conseguidas comúnmente en el rumen de la oveja consumiendo forrajes. Esto podría explicarse en parte por la baja calidad del sustrato fermentado y a la digestión gástrica de los componentes nitrogenados y carbohidratos solubles en las porciones de tracto anteriores al ciego. Por eso las proporciones de acetato (81–83%) son mayores y las de butirato (2–4%) menores a las conseguidas en el rumen de bovinos y ovinos, debido a la digestión gastrica e intestinal de los componentes solubles de la dieta.
| Raciones | Comportamiento | total de AGV (mM) | Acetato | Propionato | Proporciones (%) Butirato |
| Forrajes | |||||
| Estómago | 6 | 73 | 21 | 6 | |
| Ciego | 44 | 83 | 15 | 2 | |
| Colon | 26 | 80 | 17 | 3 | |
| Concentrados | |||||
| Estómago | 6 | 65 | 28 | 7 | |
| Ciego | 41 | 81 | 14 | 4 | |
| Colon | 25 | 77 | 16 | 7 |
Fuente: Baldizan, Dixon y Parra (1983)
La economía del nitrógeno es sin duda un punto clave de la fisiología digestiva de esta especie y como ya se mencionó, tiene una significación ecológica de gran relevancia.
La proteína ingerida por el capibara como parte de los forrajes se digiere y absorve principalmente en el duodeno, iniciándose su digestión en el estómago, al igual que los otros nutrientes. Sin embargo, es necesario estudiar con mayor profundidad la digestión microbial a nivel del ciego, porque de éste depende la economía del nitrógeno del capibara.
Si se analiza la figura 3 del metabolismo del nitrógeno en el capibara propuesto por Parra (1977), modificado de Slade et al. (1970), se resume que este animal es capaz de utilizar la proteína micronbiana sintetizada a partir de nitrógeno amoniacal, y así suplir parte de las necesidades de aminoácidos esenciales que el animal necesita para su adecuada nutrición.
En los trabajos realizados por González-Jiménez y Escobar (1973), se constató que el capibara digería mejor la proteína del forraje y menos la de alimentos concentrados comparado con un ovino y un conejo. Estos autores calcularon el requimiento de nitrógeno para el capibara de 414 mg de N y para el ovino de 367 mg de N por kg peso metabólico. El requerimiento diario de proteína en g por kg de peso metabólico fué determinado en 2.5, 2.0 y 2.3 para el capibara, ovino y conejo, respectivamente. Corroborando lo dicho por Parra (1977), en cuanto a la similitud de los valores de excreción endógena, de origen bacterial principalmente, en herbívoros rumiantes y no rumiantes.
Gran parte de la fracción endógena es de origen bacterial, por lo tanto la fracción endógena de nitrógeno es similar para rumiantes y otros herbívoros no rumiantes.
Parra (1977) propone dos hipótesis para explicar el metabolismo del nitrógeno en el capibara: la autolisis de la microflora y la cecotrofía. La lisis de las bacterias componentes de la microflora aún no ha sido muy bien explicada, pero Slade et al. (1971), inyectando nitrógeno marcado (N15) al contenido líquido del ciego de un caballo, obtuvieron dos picos de radioactividad en la proteína sanguínea. Uno a los 60 minutos, sugiriendo una absorción muy rápida de aminoácidos y amonio desde el ciego; y otro a las cuatro horas, indicando la hidrólisis y absorción de la proteína microbial.
Metabolismo del Nitrogeno en el Capibara
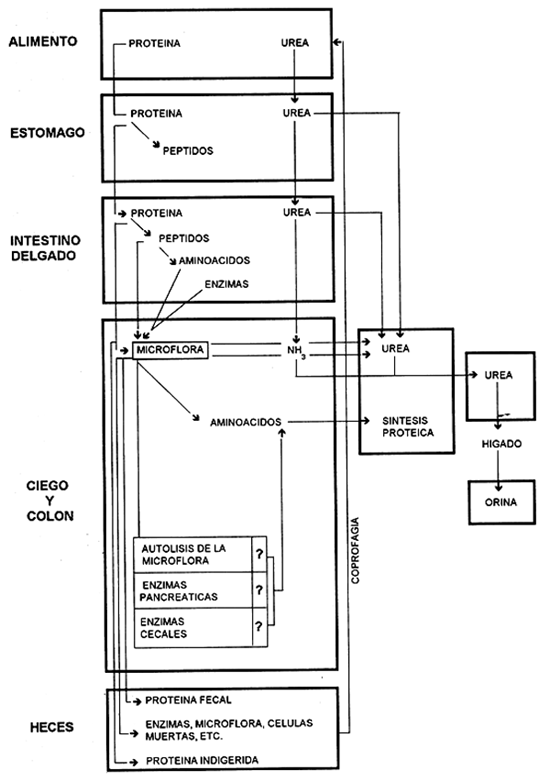
Fuente: PARRA (1977)
En observaciones realizadas por González-Jiménez y Parra (1972), sobre la estructura del ciego del capibara, corroboradas por los estudios histológicos de Leal Medina (1978), se pudo constatar a la entrada del ciego un área más vascularizada y de pared más gruesa, donde pudiera efectuarse la lisis y absorción de la proteína bacterial.
Los herbívoros monogástricos que practican la cecotrofía tienen un excelente mecanismo para utilizar tanto vitaminas como la proteína bacterial sintetizada por microorganismos del aparato digestivo (Hintz, 1969). Según Parra (1977), el capibara como animal herbívoro de talla mediana, con requerimientos energéticos relativamente bajos, con una alta ingestión de forraje y proteína, y con un desmenuzamiento muy eficiente del forraje consumido, no necesita practicar la cecotrofía.
González-Jiménez et al. (1976) compararon la excreción fecal de capibaras, conejos y ovinos utilizando una dosis única de óxido de cromo. La curva de excreción fecal del conejo fué escalonada debido a las reingestiones diarias mediante la cecotrofía. En el capibara y el ovino mostraron curvas similares.
Sin embargo, Herrera (1985), observó en el campo un comportamiento indicativo de cecotrofía y argumenta que en las mediciones realizadas por González-Jiménez et al. (1976) no se manifestó esta práctica al estar los animales en cajas metabólicas.
Muchos piensan que mediante la cecotrofía se incrementa la eficiencia de utilización de la energía y de algunos nutrientes como la proteína y las vitaminas.
Davidson, Laycock y Woods (1984) argumentan que existe una posible vía para que el capibara no necesite practicar la cecotrofía. Esta puede ser la de disponer de un mecanismo de reflujo cólico-caecal, el cual se realizaría mediante un reflujo de los contenidos cecales hacia el colon en la parte próxima de éste, mezclando ambos contenidos casi completamente mediante movimientos peristálticos. Efectuándose una separación que permite a ciertos componentes circular hacia adelante (líquido) o hacia atrás del ciego (sólidos) cuando el resto de los componentes circula caudalmente.
La chinchilla, el cuye y ciertos roedores más pequeños poseen este mecanismo de separación, que con el reflujo del material cecal permite la absorción de los aminoácidos esenciales y algunas vitaminas en la porción distal del tracto digestivo. Sin embargo, debieran realizarse estudios más detallados en el capibara para comprobar si este mecanismo existe.
La ingestión de alimentos en condiciones naturales no es fácil de medir, sobre todo en un ambiente con períodos tan marcados de inundación y extrema sequía que se alternan durante el año. Ojasti (1978) en sus estudios sobre capacidad de carga en las sabanas de Apure determinó un consumo diario de un grupo de 12 capibaras por ha de 203 kcal/m2 por año, que sólo equivale al 3.5% de la producción primaria neta de ese ecosistema.
El consumo de alimentos en condiciones experimentales fue realizado por González-Jiménez et al. (1976) al estudiar la eficiencia digestiva comparada del capibara, el conejo y el ovino. El forraje suministrado fué en forma comprimida para evitar la selección, y además se dió un alimento concentrado con 17% de proteína cruda. El consumo de materia seca expresado en porcentaje del peso vivo y al peso metabólico (Cuadro 10) indican que el capibara tiene una ingestión un poco menor que el rumiante y el conejo. Estos consumos, sin embargo, son bajos en relación a los que se consiguen con forrajes de buena calidad.
| Peso Vivo | Consumo | Consumo en | Consumo M.S. | |
| Especies | en Kg | M.S/dia | %Peso vivo | % P0.75 |
| Capibaras | 24.49±3.23 | 444 ± 61 | 2.57 ± 0.59 | 4.03 |
| Conejos | 3.41±0.15 | 94.9 ± 16.2 | 2.78 ± 0.50 | 3.78 |
| Ovinos | 46.88±2.23 | 1259 ±147 | 3.68 ± 0.29 | 7.00 |
Fuente: González-Jiménez, Escobar y Caires (1976)
Al calcular la digestibilidad (Cuadro 11), se aprecia que no existen diferencias para el forraje entre capibaras y ovinos, siendo significativamente más baja la del conejo (20%). Pero cuando consumieron exclusivamente concentrado, los herbívoros monogástricos tuvieron mayor digestibilidad.
| % Concentrado en la ración | Capibara | Ovino |
| 0 | 50,5 | 49,1 |
| 20 | 57,4 | 53,5 |
| 40 | 64,2 | 57,8 |
| 60 | 71,0 | 62,1 |
| 80 | 77,9 | 66,5 |
| 100 | 84,7 | 70,8 |
Los valores fueron calculados por las euaciones:
Y= 0,217X + 49,15 para los ovinos.
Y= 0,342X + 50,56 para los Capibaras.
Donde: Y es la digestabilidad en % de la materia seca.
X Proporción de concentrado en la ración.
Esto se hizo para obtener resultados comparables a porcentajes fijos de ingestión.
Baldizan et al. (1981), encontraron que en los ovinos cuando a una ración de heno de Cenchrus ciliaris se le añadió alimento concentrado, se incrementó la digestibilidad de la materia seca en 4% y se redujo la digestión de los carbohidratos estructurales en 7.5%. Pero en el capibara sólo redujo en 1.3% la digestibilidad de la celulosa e incrementó en 12% la de la ración. El disponer de la digestión gástrica previa a la microbial confiere a este animal una gran ventaja en la utilización de los alimentos concentrados y sobre todo en la utilización de la proteína.
El capibara tiene una tasa de recambio rápida, pero una velocidad de tránsito digestivo suficientemente lenta como para que se produzca una eficiente fermentación microbial. La eficiente molienda y el desmenuzamiento que realiza el capibara en la masticación, le confieren una doble ventaja. Por un lado disminuye la velocidad de paso de la ingesta en el ciego y por otro incrementa la superficie de ataque de las bacterias para una mejor fermentación microbial (Parra 1977). Además el sustrato que entra en el ciego del capibara, sufre en el estómago la digestión enzimática, mejorando la disponibilidad de los carbohidratos estructurales (celulosa y hemicelulosa) para el ataque microbial en este compartimiento.
El medio cecal donde la fermentación microbial se efectúa es más constante y menos influenciado por la ingestión continua de alimentos (pastoreo y rumia), pudiendo entonces realizarse una fermentación microbial más eficiente (Van Soest, 1966)). Esto último ha sido comprobado al utilizar un método in vitro para analizar varias muestras de pastos naturales, incubadas previamente con pepsina (González-Jiménez y Escobar, 1975). Incubando las mismas muestras, con licor ruminal o cecal, se constató un incremento de la digestabilidad de la materia seca de 14% y 17%, respectivamente. Este resultado se atribuyó al efecto de la digestión enzimática previa en el estómago, cuyo efecto sobre ciertos componentes de la pared celular de los forrajes (hemicelulosas y celulosa) permitían una mejor acción de las exoenzimas de las bacterias en los restos de la pared celular del sustrato fermentado. Además de lo ya mencionado sobre la selección que hace el capibara de los mejores y más ricos forrajes en proteína existentes en la sabana inundable, esto terminaría de explicar la gran adaptación y eficiencia digestiva de este herbívoro monogástrico. Su nivel de eficiencia es comparable a la que muestran los rumiantes para los mismos forrajes.
Dada la disponibilidad muy fluctuante de los forrajes en la sabana inundable y para estudiar la competencia alimenticia de los herbívoros mayores se emprendieron los estudios de productividad de la sabana en el Banco, el Bajío y el Estero. Esto se realizó para comprobar la hipótesis de que existe una superposición de los cuatro herbívoros mayores del llano: capibara, caballo, vacuno y venado.
Se midió el consumo de los diferentes pastos mediante el método ya citado en tres períodos clave de la sabana inundable: al final del período de lluvias (mayor abundancia), al final de la sequía (menor abundancia) y durante las lluvias. En el cuadro 12, se resumen las observaciones realizadas en los cuatro especies animales estudiadas de la ingestión que realizaron de los pastos de la sabana. Se realizaron más de doce mil observaciones microscópicas, determinándose que los capibaras cubren el 82% de su dieta consumiendo plantas del bajío y el estero, no así los caballos y vacunos que sólo hacen un 64%. Gran parte de la diferencia puede deberse al consumo de Ciperaceas (16% en total) que realiza el capibara, así como de la Eichornia (2%). Es interesante resaltar la baja frecuencia de la Parathenia prostata en el consumo de los vacunos, aun siendo muy abundante en el Bajío y el alto consumo que el capibara realiza de este forraje. El Caballo sí compite con él por esta gramínea. Estos forrajes cortos pueden ser consumidos por roedores gracias a sus incisivos y por el caballo, que con los labios los aprehenden facilmente.
| Especies forrajeras | Especies animales | ||
| Capibaras Bovinos | Equinos | Venados | |
| Hymenachne amplexicaulis | 25 | 20 | 222 |
| Paratheria prostata | 17 | <1 | 81 |
| Leersia hexandra | 15 | 19 | 211 |
| Sporobolus indicus | 9 | 13 | 6<1 |
| Panicum dichotomiflorum | 5 | 5 | 2<1 |
| Axonopus sp. | 4 | 14 | 200 |
| Luxiela sp. | 2 | <1 | 4<1 |
| Panicum laxum | 1 | 16 | 20 |
| Panicum junceum | <1 | 1 | <10 |
| Setaria geniculata | <1 | 2 | 4<1 |
| Oryza perennis | <1 | <1 | <10 |
| Panicum zizanioides | <1 | <1 | <1<1 |
| Eracrosti acutiflora | <1 | <1 | <10 |
| Paspalum chaffanjonil | <1 | 2 | <1<1 |
| Paspalum orbiculatum | <1 | <1 | <1<1 |
| Ciperacea | 16 | 3 | 4<1 |
| Eichhornia sp. | 2 | 0 | 00 |
| Otros y/o no identificados | 3 | 4 | 52 |
| Dicotiledoneas | 0 | 0 | 093 |
Los menos de 1% hacen: | 1 | 1 | 21 |
| TOTAL | 100 | 100 | 100100 |
El Bajío es la porción más grande de la sabana inundable y la que define hasta cierto punto la inexistencia de competencia alimenticia antes mencionada.
El Estero es la unidad fisiográfica donde abundan las especies más apetecidas, pues es donde los forrajes crecen todo el año ya que disponen de agua constantemente (tanto en la superficie como en el suelo).
En el Banco, con suelos más pobres, y comunmente pobladas por gramíneas anuales de crecimiento efímero y de lignificación muy rápida, la competencia interespecie as menor. Las especies vegetales aquí presentes son de porte alto y en macollas, como el Axonopus sp., Setaria geniculata, Paspalum chaffanjoni, consumidas preferencialmente por bovinos y equinos y poco por el capibara.
En verano, la competencia alimenticia, se hace crítica en el Estero, donde las plantas escogidas son muy similares pues son las que quedan aún verdes y hasta los venados las consumen. Era de esperarse que el capibara fuese un consumidor de la producción primaria del Estero, por su condición de animal semiacuático; pero no se pensó que todos los otros herbívoros hiciesen un uso tan exhaustivo de esta unidad. Sin duda esto se debe a que se conserva más la calidad del forraje en el Estero y a su mayor producción durante el año. En los estudios realizados por Brock (1970), las gramíneas identificadas por él en los tractos digestivos de los venados conseguidos en Apure correpondían a las gramíneas de esta unidad y más de 30 dicotiledóneas, correspondientes en su totalidad a los matorrales de los Bancos altos o a la selva de galerías adyacentes a los caños y los ríos.
Midiendo la competencia alimentaria y superposición de dietas mediante el Indice de Disimilaridad de Hansen (Hansen et al. 1971) presentado en el cuadro 13, no se consiguió sobreposición de dietas entre capibara y ganado hasta el final de la estación de lluvias y período de inundación de casi 8 meses. Pero al final del período seco (sequía extrema en este ecosistema sabanero) una competencia muy marcada se desarrolla entre animales silvestres (capibaras y venados) y animales introducidos (vacunos y equinos). Tal hecho se debe simplemente a la baja disponibilidad forrajera para ese momento del año, la cual puede ser muy crítica durante ciertos lapsos y hasta catastrófica en años con largas sequías.
| Ch-va | Ch-Cab | Ch-Ve | Va-Cab | Va-Ve | Cab-Ve | |
| Totales | 0,09 | 0,07 | 0,61 | 0,06 | 0,53 | 0,54 |
| Estero | 0,11 | 0,13 | 0,14 | 0,04 | 0,11 | 0,07 |
| Bajio | 0,95 | 0,13 | 0,05 | 0,65 | 0,68 | 0,20 |
| Banco | 0,10 | 0,20 | 0,50 | 0,04 | 0,53 | 0,52 |
| Selva | 0,00 | 0,00 | 1,00 | 0,00 | 1,00 | 1,00 |
Fuera de su habitat el capibara, como harbíboro que es, consume diferentes alimentos, forrajes principalmente. Sin embargo, en algunos zoológicos los alimentan con raciones donde incorporan hasta carne y pescado (Zara, 1973). El naturalista Buffon (1844) indica que los vio consumir peces de los estanques de Fontaineblau en Francia. Osgood (1912) reportó que los contenidos estomacales de los capibaras del Lago de Maracaibo están formados por muchas algas del fondo del Lago. Mondolfi (1957) y Humboldt (1820) reportaron consumo de la paja chigüirera (Paspalum fasciculatun) de los capibaras que habitan los revalses de los ríos Orinoco y Apure. Nogueira (1973) reporta que los capibaras pueden dañar las plantaciones de caña de azúcar y arroz, por ello en Brasil en muchos sitios es considerado como una plaga y es sometido a exterminio por los propietarios de grandes plantaciones.
La alimentación en captividad es muy variada y exótica, en gran parte por desconocer que este animal es un estricto herbívoro y además porque aprende muy fácilmente a comer cualquier alimento, como monogástrico que es. Las dietas que le suministran en los jardines zoológicos son tan extravagantes como variadas; en cada una existe un preparado que va desde la mezcla de cubos para bovinos, primates, lagomorfos, monogástricos y hasta flamencos, pasando por frutas (manzanas), papas, zanahorias, remolachas, hasta una gama rica en ingenio y hasta, despilfarro se puede decir. Las sales minerales, bloques de diferentes contenidos, azules con sales de cobre, rosados con otros elementos, compuestos vitamínicos y hasta sobredosis de Vitamina E para mejorar la reproducción (Zara, 1973; Donaldson, 1975; Davidson et al., 1984). Un forraje de buena calidad compuesto fundamentalmente de gramíneas y un alimento concentrado balanceado (por ejemplo para vacas lecheras) constituirían una dieta más que adecuada.
Los pastos más frecuentemente ofrecidos son: Brachiaria mutica (Pasto Pará)), Hyparrhemia rufa, Melinis minutiflora, Panicum maximum, Paspalum plicatulum, Tripsacum laxum y algunos tallos y hojas de los cultivos como caña de azúcar (Saccharum officinarum) y arroz (Oriza sativa), según Mondolfi (1965), Otero de la Espriella (1973), Cruz (1974) y Piccini (1971). En Brasil, en el Zoológico Goeldi, Belen du Pará, le suministran repollos (Brassica oleracea) y lechuga (Lactuca sativa), raíces y tubérculos como batata (Ipomea batata), yuca (Manihot sculenta), papa (Solanum oficinarum) y ocumo (Colocasia sculenta), asi como patilla (Cucurbita pepo) y bananos (Musa paradisiaca). También le proveen residuos de molinería, como afrechos y granos partidos de trigo o arroz y hasta maíz (Zea maiz). Fuerbringer (1974), además de la mayoría de los anteriores alimentos, menciona la melaza de caña mezclada con alimentos harinosos para evitar la inhalación de polvo en los alimentos.
En los experimentos realizados por Parra et al. (1978) se le ofreció pasto elefante cortado (Pennisetum purpureum) y un concentrado 17% de P.C. para cerdos. Muchos piensan que como al capibara se le encuentra en sabanas pobres y desoladas en verano, éste puede alimentarse con cualquier cosa y en condiciones marginales. Olvidando que en su habitat seleccionan la mejor dieta, rica en forrajes muy nutritivos y que para estar en buenas condiciones de salud necesitan buena alimentación y óptimas condiciones de vida, áreas limpias con abundante agua corriente, alimentos variados como forrajes, raíces y tubérculos, frutas, hortalizas ofrecidas en comederos, rastrillos y bebederos para que no sean contaminadas por excretas u otros productos nocivos al animal.
![]()
![]()
![]()