Chapter 3 : Aspects of fat digestion and metabolism
Digestion, absorption and transport
Metabolism
of fatty acids
Essential
fatty acids
Biological
roles
Oxidation
of fatty acids
Digestion, absorption and transport
Most dietary fat is supplied in the form of triacylglycerols which must be hydrolyzed to fatty acids and monoacylglycerols before they can be absorbed. In children and adults, fat digestion is efficient and is nearly completed in the small intestine. In the newborn, the pancreatic secretion of lipase is low. The digestion of fat in babies is augmented by the lipases secreted from the glands of the tongue (lingual lipase) and a lipase present in human milk. The stomach is part of the process of fat digestion because of its churning action which helps to create an emulsion. Fat entering the intestine is mixed with bile and is further emulsified. The emulsion is then acted upon by lipases secreted by the pancreas. Pancreatic lipase catalyzes the hydrolysis of fatty acids from positions 1 and 3 to yield 2-monoacylglycerols (Tso, 1985). Phospholipids are hydrolysed by phospholipase A2 and the major products are lysophospholipids and free fatty acids (Borgstrom, 1974). Cholesterol esters are hydrolyzed by pancreatic cholesterol ester hydrolase.
The free fatty acids and monoglycerides are absorbed by the enterocytes of the intestinal wall. In general, fatty acids which have a chain length of less than 14 carbons enter directly into the portal vein system and are transported to the liver. Fatty acids with 14 or more carbons are re-esterified within the enterocyte and enter the circulation via the Iymphatic route as chylomicrons. However, the portal route has been described as an absorptive route for dietary long chain fatty acids as well (McDonald et al., 1980). Fat soluble vitamins (vitamins A, D, E and K) and cholesterol are delivered directly to the liver as part of the chylomicron remnants.
Diseases that impair the secretion of bile, such as biliary obstruction or liver diseases, lead to severe fat malabsorption, as do diseases that influence the secretion of lipase enzymes from the pancreas, such as cystic fibrosis. As a result, medium-chain triglycerides can be better tolerated in individuals with fat malabsorption and these are often used as a source of dietary energy. Complete absorption of lipids from the intestine may be marginally affected by a high amount of fibre in the diet. Fat absorption is illustrated in Figure 3.1.
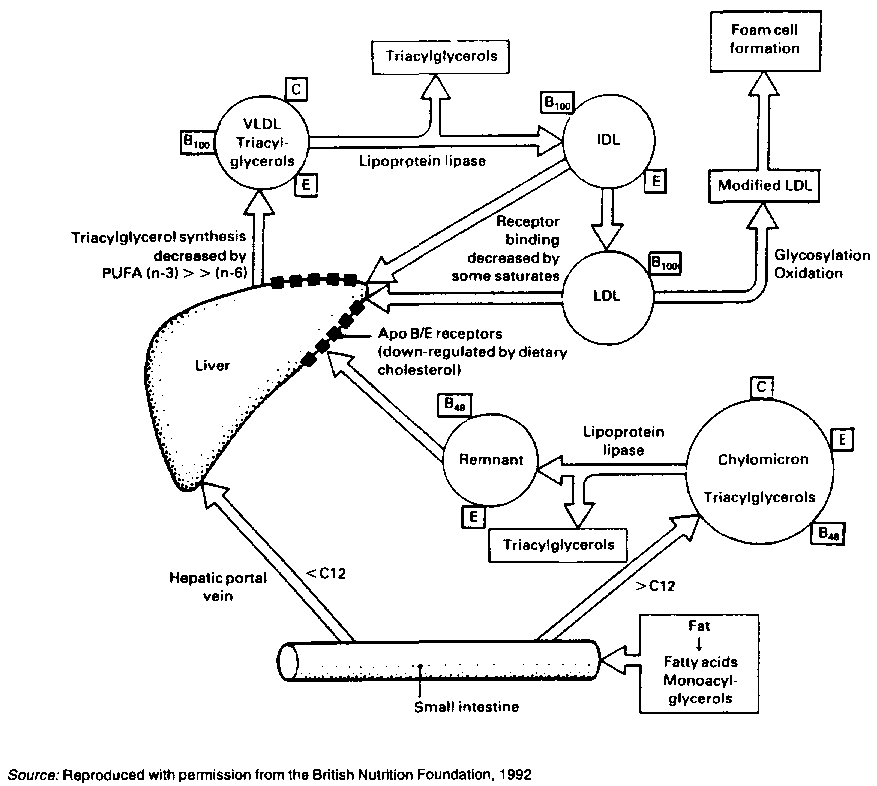
Fatty acids are transported in the blood as complexes with albumin or as esterified lipids in lipoproteins. These consist of a core of triacylglycerols and fatty acid esters of cholesterol, and a shell of a single layer of phospholipids interspersed with unesterified cholesterol. Coiled chains of one or more apolipoproteins extend over the surface and, with the amphipatic phospholipids, enable the lipids in the core to be carried in the blood. They also regulate the reaction of the lipid package with specific enzymes or bind the particle to cell surface receptors.
Chylomicrons are lipoprotein particles derived from dietary fat and packaged by the mucosa cells. They enter the blood stream via the Iymph vessels. Lipoprotein lipase located on the interior walls of the capillary blood vessels hydrolyses the triacylglycerols, releasing fatty acids. These enter the adipose tissue where they are stored, and the muscles where they serve as fuel. The remnants of chylomicrons are cleared by the liver within a few hours of the ingestion of a fat-containing meal.
Very-low density lipoproteins (VLDL) are large triacylglcerol-rich particles produced in the liver from endogenous fat, as opposed to chylomicrons which transport exogenous fat. VLDLS are the main carriers of triacylglycerols which are also processed by lipoprotein lipase and supply fatty acids to adipose and muscle tissues.
Low-density lipoproteins (LDL) are the end products of VLDL metabolism. The core consists mainly of cholesterol esters and its surface has only one type of apolipoprotein, apoB. About 60-80 percent of cholesterol in plasma is carried by LDLs. Average LDL values vary among populations because of genetic and environmental factors, however, diet is probably a major determinant of these values.
FIGURE 3.1 : The influence of dietary fat on lipoprotein metabolism
High-density lipoproteins (HDL) carry 15-40 percent of plasma cholesterol. They are probably formed in the circulation from precursors made in the liver and in the gut. The major apolipoprotein of HDL is apoA-1. In humans, LDL delivers cholesterol to the liver and HDL may transfer cholesterol to other lipoprotein particles. There is evidence that HDL actively protects vessel walls (NIH Consensus, 1993). It is not known whether manipulation of HDL levels by diet affects the development of atherosclerosis.
Lipoprotein(a) or Lp(a) is a complex of LDL with apolipoprotein(a). This apoprotein has a sequence homology with the pro-enzyme plasminogen which is involved with the dissolution of blood clots (Scanu and Scandiani, 1991). The concentration of Lp(a) is largely determined by genetics.
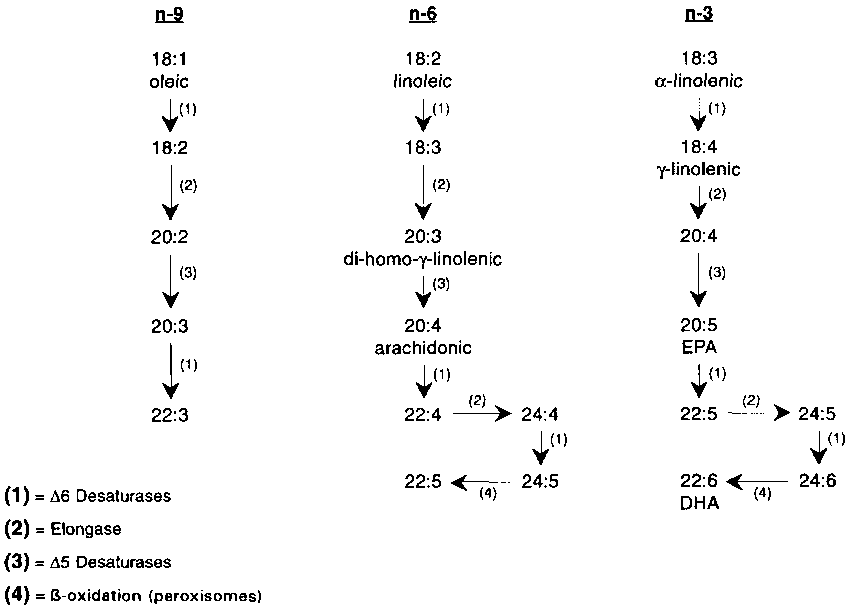
Saturated fatty acids and monounsaturated fatty acids can be biosynthesized from carbohydrates and proteins. Unsaturated fatty acids may be substrates for desaturases and elongases, as shown in the scheme for conversion in the n-9, n-6 and n-3 families of fatty acids (Figure 3.2).
FIGURE 3.2 : Sequence of conversions of unsaturated fatty acids
The first members of each fatty acid family (oleic, linoleic and a -linolenic acid) compete for the same 6-desaturase, with rates of conversion increasing with the number of double bonds. This ratelimiting enzyme is under the control of many dietary and hormonal factors (Brenner, 1989; 1990) and is believed to be important in the synthesis of 22:6 n-3. Such an effect may explain why high intakes of linoleic acid reduce the level of 22:6 n-3. Similarly, the 5-desaturase is modulated by dietary and hormonal factors. The C20 and C22 members of the n-6 and n-3 families can inhibit desaturation earlier in the sequence of fatty acid conversion (Fisher, 1989).
It appears that the 4-desaturation does not involve another specific desaturase, but an elongation, then 6-desaturase, both microsomal processes, followed by retroconversion by the peroxisomal ,8oxidation pathway (doss et al., 1991). The desaturation is thus a process based on intracellular cooperation. The nutritional significance of this additional role for 6-desaturase has yet to be evaluated. The activities of the elongases appear to be greater than those of desaturases (Sprecher, 1989).
The essential fatty acids are those that must be supplied in the diet and they include members of both the n-6 and n-3 series. Under conditions of a dietary deficiency of linoleic acid, the most abundant unsaturated fatty acid in tissue, oleic, is desaturated and elongated to eicosatrienoic, n-9, which is normally found in trace amounts only. The accumulation of this fatty acid is considered to be a marker of essential fatty acid deficiency (Holman, 1970). When the diet has a low n-3 fatty acid content, compared to the n-6, there is a reduction of 22:6 n-3 associated with a compensatory accumulation of 22:5 n-6 in tissues (Gall), Agradi and Paoletti, 1974).
The most abundant product of the n-6 pathway is arachidonic acid found in glycerophospholipids where it is selectively esterified in the 2-position. Diets containing fats of animal origin provide appreciable amounts of preformed arachidonic acid.
The efficiency of the n-3 pathway in humans has been open to question. However, human studies on the conversion of deuterated linoleic and a -linolenic acids to their long chain derivatives showed that the efficiency for the conversion of linoleic to arachidonic was 2.3 percent, while the conversion of a -linolenic to its long chain derivatives was 18.5 percent (Emken et al., 1992). These conversion values, measured as the accumulation of products in plasma, were obtained in subjects with relatively low intakes of linoleic acid (around 5 percent). When the diet contained around 9 percent linoleic acid, the conversions were reduced to 1 percent for linoleate and 11 percent for linolenate. These data do indicate that the synthesis of n-3 metabolites from 18:3 occurs in adults.
Fish oil, which is rich in eicosapentaenoic acid compared to linseed oil, greatly enhances the concentration of the long-chain derivatives in human blood (Sanders and Roshanai, 1983). High levels of eicosapentaenoic acid and docosapentaenoic acid in blood platelets or other cells are attained only when they are provided as such in the diet. The n-3 fatty acids, like the n-6 fatty acids, are incorporated into the 2-position of membrane phospholipids.
The polyunsaturated fatty acids for membrane phospholipids come from the diet or endogenous sources. The availability is influenced by deacylation-reacylation, particularly at position-2 of phospholipids (Lands et al., 1982), the specificity of diacylglycerol kineses in phospholipid synthesis (McDonald et al., 1988), and remodelling of donor and acceptor phospholipids. In all of these processes, competition exists between the fatty acids of the n-6 and n-3 series.
The essentiality of linoleic acid was demonstrated in studies of rats. When rats were fed a fat-free diet, they developed specific symptoms that were prevented by the provision of linoleic acid. Linoleic acid deficiency, a rare condition in humans, was described in children given fat-free diets (Hansen et al., 1963). The children developed skin disorders similar to those produced in rats that were cured by the administration of linoleic acid. To prevent the accumulation of ecosatrienoic acid (20:3 n-9), the amount of linoleic acid required in the diet is at least 1 percent of the energy intake. This applies to all the animal species tested. Intakes between 4 and 10 percent of the energy are currently consumed by different population groups, and these appear to be compatible with optimal health status.
Linoleic acid deficiency may develop as a secondary condition in other disorders, such as proteinenergy malnutrition and fat malabsorption, or as a consequence of total parenteral nutrition with inadequate linoleic acid intakes. Under conditions of adequate linoleic acid intake, the ratio of triene to tetraene is below 0.2 (Mohrhauer and Holman, 1963). A better assessment of linoleic acid deficiency can be obtained with a profile of all polyunsaturated fatty acids in serum (Holman and Johnson, 1981).
The n-3 fatty acid, docosahexaenoic acid (DHA, 22:6), is present in high concentrations in the central nervous system, in the cell membranes, and the visual system (Tinoco, 1982; Neuringer et al., 1984; Bourre et a/., 1989). The role of DHA for optimal neuronal function and visual acuity is discussed in relation to early development later in this report.
The balance between n-6 and n-3 fatty acids in the diet is important because of their competitive nature and their essential and different biological roles. It is suggested that the relative amounts of linoleic and a -linolenic acids in the diet should be below 10:1.
Membrane structure. Unsaturated fatty acids in membrane lipids play an important role in maintaining fluidity (Lynch and Thompson, 1984). In the skin, linoleic acid plays a specific role where it is linked to some very long chain fatty acids (C30-C34) in the acyl ceramides. These form an intercellular matrix to maintain the epidermal permeability barrier (Hansen and Jensen, 1985).
In membranes, interactions between lipids and proteins may depend upon a specific polyunsaturated fatty acid. This appears to be the case for mammalian rod outer segments which are very rich in docosahexaenoic acid.
Three examples of lipid-protein interactions that control the metabolic functions of membranes were noted: first, the catalytic properties of transporter proteins; second, the activities of enzymes such as the Ca/Mg ATPase of sarcoplasmic reticulum, adenyl cyclase, and 5-nucleotidase which are influenced by the levels of n-6 and n-3 fatty acids in the membrane lipids (Kinsella, 1990); and third, enzymes involved in the phosphoinositide cycle. This cycle, associated with the responses of many cells to a range of hormones, neurotransmitters and cell growth factors, gives rise upon activation of a specific phospholipase C to two important second messengers: inositol 1,4,5 trisphosphate (IP3) and diacylglycerol. IP3 is responsible for the modulation of cytosolic calcium ions. Diacylglycerol, together with calcium ions and phosphatidyl serine, is involved in the activation of a protein kinase C which phosphorylates a number of intracellular proteins. Since polyphosphoinositides are very rich in arachidonic acid in position 2, diacylglycerol arising from them is also rich in this fatty acid. A lipase acting on diacylglycerol may subsequently release arachidonic acid for conversion to eicosanoids.
The phosphoinositide cycle controls processes in cell division. The modulation of the cycle, therefore, influences the rate of division of cells in the immune response and in tumour growth. Dietary polyunsaturated fatty acids and the proportion of n-6 and n-3 fatty acids appear to affect the phosphoinositide cycle.
Eicosanoid formation. The n-6 and n-3 fatty acids in the membrane phospholipids exert metabolic control through their role as precursors of eicosanoids. These highly active compounds of 20 carbon atoms are released in very small quantities to act rapidly in the immediate environment. After enzymatic degradation, the products from eicosanoids in the urine are an indication of the production by the body.
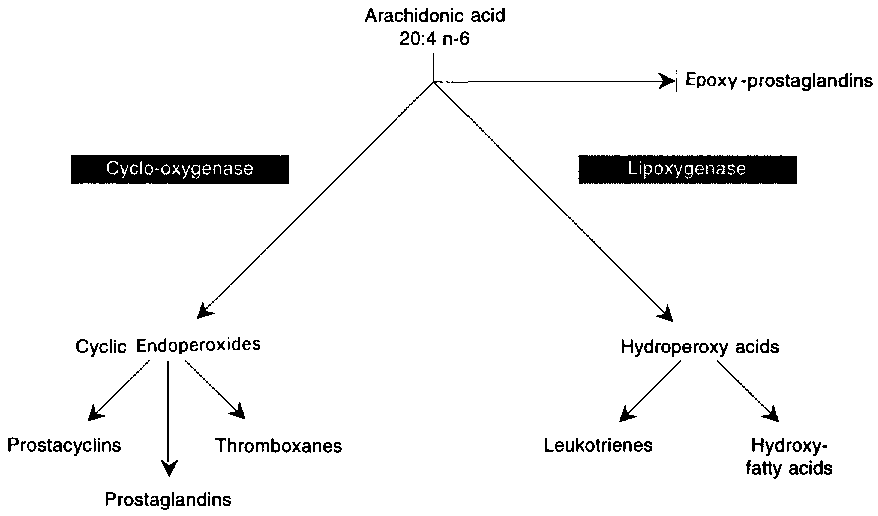
The first step in the biosynthesis of eicosanoids is the release of a 20 carbon polyunsaturated fatty acid from phospholipids by phospholipases, mainly a phospholipase A2, or from diacylglycerol produced in the inositol phosphate cycle. The eicosanoid cascade consists of hydroxylated derivatives of 20 carbon polyunsaturated fatty acids: (a) cyclic products, generated by a cyclo-oxygenase, which include prostaglandins, prostacyclin and thromboxane; (b) lipoxygenase products such as the 12-lipoxygenase derivatives, especially the products of the 5-lipoxygenase known as leukotrienes; and (c) products of cytochrome P450 activity (Figure 3.3). Eicosanoids, in general, are very potent, their effects are highly diverse, and the action of different eicosanoids often oppose each other. In addition, the patterns of eicosanoid production are different in various cells and tissues.
Among the most active eicosanoids, thromboxane A2(Tx A2) derived from arachidonic acid is produced by platelets and other cells through the cyclo-oxygenase pathway. This eicosanoid is a platelet proaggregatory and smooth muscle contracting agent, and is rapidly inactivated to thromboxane B2. Prostacyclin (PGI2), produced through the cyclo-oxygenase in cells of vessel walls, is a platelet anti-aggregatory and vasodilating agent. Other products of the cyclo-oxygenase pathway (for example, PGE2 and PGF2a ) exert various effects on smooth muscle cells, immune competent cells, and so forth. Among the products of the lipoxygenase pathway, the leukotrienes, produced mainly by the leukocytes, act on vascular parameters (permeability, contractility) and have chemotactic properties. They are involved in the modulation of inflammatory and immune processes.
Polyunsaturated fatty acids with 20 carbons and different degrees of unsaturation give rise to eicosanoids with different numbers and patterns of unsaturation, and with somewhat different biological activities. Since arachidonic acid (20:4 n-6) is the major polyunsaturated fatty acid in cells, the 2-series of eicosanoids is the most abundant and generally the most active. When a 20 carbon polyunsaturated fatty acid with a different number of double bonds, for example, di-homo-g linolenic (DGLA), 20:3 n-6, or eicosapentaenoic acid (EPA), 20:5 n-3, is incorporated into cell lipids, eicosanoids of the 1-series or the 3-series, respectively, are produced. These fatty acids also compete with arachidonic acid for the cycloxygenase, and thus reduce the formation of eicosanoids of the 2-series. EPA promotes the formation of eicosanoids of the 3 series and inhibits the formation of eicosanoids of the 2 series.
FIGURE 3.3 : The eicosanoid cascade
Effects on other lipid-derived mediators. Hydrolysis of selected cell phospholipids results in the formation of additional biologically active compounds such as the platelet activating agent, PAF. This compound arises from l-alkyl, 2-acyl phosphatidyl choline. PAF is an extremely potent pro-inflammatory agent, and a potent activator of several types of cells in addition to platelets. Although data on the effects of various polyunsaturated fatty acids on the PAF pathway are limited, incorporation of arachidonic acid into cell phospholipids was shown to potentiate PAF production (Suga et al., 1991). Different polyunsaturated fatty acids in position 2 of the precursor phospholipids can modify PAF formation, as was shown by the reduced production of PAF by monocytes of subjects receiving n-3 fatty acids (Sperling et al., 1987). The opposing effects of n-6 and n-3 fatty acids may explain some influences of polyunsaturated fatty acids on the function of certain cells (Ibid.).
Effects on other parameters. The n-3 fatty acids appear to affect various other processes, such as the production of cytokines and other factors. Cytokines are a family of proteins produced and released by cells involved in inflammatory processes and in the regulation of the immune system. These include the interleukins and tumour necrosis factors. The mechanism by which n-3 fatty acids affect cytokine synthesis is not clear, but some studies have shown an effect on mRNA levels, suggesting a pretranslational level of action.
The effects of fatty acids on the expression of genes encoding for enzymes which are involved in lipid metabolism, as well as on the expression of genes involved in cell growth regulation (immediate early response genes), represent an additional important aspect of the biological roles of polyunsaturated fatty acids. It is apparent that fatty acids can interact with a group of nuclear receptor proteins that bind to certain regions of DNA and thereby, alter transcription of regulatory genes.
Much of the literature on the n-3 long-chain fatty acids deals with pharmacological doses or is concerned with lipoproteins and their relationship to coronary heart disease. Additional clinical and metabolic studies should be encouraged.
Those fatty acids that are not used for synthesis of eicosanoids or incorporated into tissues are oxidized for energy. Fatty acids yield energy by beta oxidation in the mitochondria of all cells, except those in the brain and kidney. They enter the mitochondria as specific acyl carnitine derivatives. Saturated short, medium, and long-chain fatty acids undergo the first step of beta-oxidation with different dehydrogenases. The process yields successive acetylCoA molecules which enter the tricarboxylic acid cycle or other metabolic pathways. Acetate is the eventual product from fatty acids with an even number of carbon atoms. Unsaturated fatty acids require two more enzymatic steps than saturated fatty acids to change cis double bonds to trans and to move them from the a to beta position. Even so, oxidation of unsaturated fatty acids, including linoleic acid is as fast or faster than that of palmitic acid. The initial oxidative reaction is carried out by a different enzyme from that in mitochondria; the fatty acyl CoA enters directly into this organelle. The process does not proceed to the complete production of acetate but a shortened fatty acid is transferred to the mitochondria for complete oxidation. Long chain (>20C) fatty acids are preferentially oxidized by peroxisomes; also fatty acids with less than 14C are oxidized by this system. Peroxisomal oxidation is energetically less efficient than mitochondrial oxidation and yields more heat. This type of oxidation can be induced by diets which are high in fat as well as by a variety of xenobiotics.
{kind=link}
{kind=link}
{kind=link}