![]()
![]()
![]()
Foster (1982) utilized legion analysis (Pope, 1980) in a preliminary examination of a selected group of species on Georges Bank to estimate abundance and mortality. Although the overall conclusion of this preliminary analysis indicated that including the feeding interactions in these analyses did not greatly alter the conclusions drawn from cohort analysis of individual species, there were some significant indications of extensive feeding on the very early juvenile fish and thus a possibility that further analysis might be promising, particularly if a wider range of species could be included.
Despite the failure of the analyses of Sissenwine et al. (1982) and Foster (1982) to clearly demonstrate species interactions there is considerable circumstantial evidence for species interaction similar to that in the analysis of Brown et al. (1976). A plot of the catches on Georges Bank from 1955 to 1979 by species and total indicates a greater relative stability in the total compared to the individual species (Fig. 19; Hennemuth, 1979). Similar conclusions can be gathered from examining the trends in trawl survey estimates of population indices as illustrated in Clark and Brown (1977).
Sissenwine 3 examined the variance of individual species catches from NAFO Subareas 5 and 6 during 1966-80 and compared the variance of the sum with the sum of the variances. He found the latter to be smaller than the former for the total catches of all species, but not for demersal species alone. Thus, the statistical evidence for stability rested with the herring and mackerel components.
Environmental Interactions
The influence of temperature has been suggested as a principal factor in controlling abundance levels of fish stocks on the continental shelves of the Northwest Atlantic off the Canadian Maritimes and the northeastern United States (Dow, 1977; Sutcliffe et al. 1977). The reductions in abundance of herring and mackerel in both the Northeast and Northwest Atlantic in response to heavy fishing mortality, followed by increases in Ammodytes and other small, fast-growing fish (e.g., Fig. 16) do not support the hypothesis that the changes are due mainly to environmental factors as they are unlikely to be the same in both areas.
Zooplankton sampling for five years over an area of 260,000 km2on the continental shelf off the northeast coast of the US has indicated different seasonal pulses in abundance in four areas that are temporally coherent. Unlike the 30-year decline of zooplankton reported for the North Atlantic and North Sea, NEFC observations indicate that zooplankton off the northeast coast of the US has not undergone any large-scale change in abundance or species composition during the past 70 years. It appears that greater perturbations have been imposed on fish populations in this area by fishing mortality than from any changes in the abundance levels of zooplankton (Sherman et al., in press).
RECRUITMENT AND ESTABLISHMENT OF YEAR CLASS STRENGTH
Recruitment is the major source of uncertainty in all of the major fisheries that have been discussed in this paper. Recruitment is affected by both the biotic and abiotic environment. Obviously, at some level there has to be a relationship to stock size. Much effort in fisheries research has gone into examining the relationships between stock and recruitment. The early pioneering work was done by Ricker, Beverton and Holt. The basic discussion can be found in their texts (Beverton and Holt, 1957; Ricker, 1958). Attempts have been made to fit relationships between stock and recruitment for most of the stocks of the Northwest Atlantic. None of these have proven statistically satisfactory. Realizing that only very minor changes in the survival in the early life history stages can cause very large differences in the number of survivors to adult sizes, it is not surprising that there is great variability about any stock recruitment function and attempts to use such a function as a predictor are not satisfactory over a wide range of stock biomasses. Frequently, advice to fisheries managers is that over a wide range of stock sizes above some minimum, recruitment can be viewed as varying independently of stock size.
3 Michael P. Sissenwine, NEFC, Woods Hole Laboratory, Woods Hole, MA, personal communication.
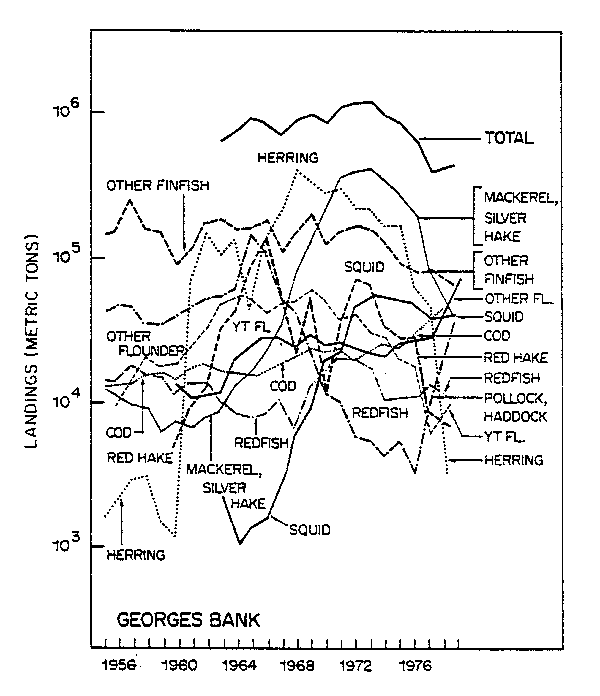
Fig. 19. Nominal catch for total and major species on Georges Bank, 1955-79.
Hennemuth et al. (1980) examined recruitment in 18 fish stocks for which they could obtain data and fit statistical distributions functions to these numbers. The stocks they examined included several in the Northwest Atlantic. In general, a lognormal distribution describe these stock recruitment observations. Further efforts to refine the statistical fitting procedures have been produced by (Taille and Patil, 1982). In almost all cases such distribution functions need to account for infrequent occurrence of very large year classes, a modal group consisting of much smaller year classes and with a few year classes being very small. The latter is most obvious in very heavily fished stocks with reduced spawning stock biomass. These data would suggest that the real problem in fishery prediction is understanding the occurrence of the very large year classes which, historically, have been observed to drive fisheries.
Serial correlations between adjacent years recruitment estimates were found by Hennemuth et al. (1980) indicating that in most cases, recruitment in one year correlated significantly with that of the previous year. However, this does not hold true for the occurrence of the very large year classes. Serial correlations are not surprising if recruitment is related to stock size which tends to be similar in adjacent years or to climatic conditions which also tend to be serially correlated.
Although certain forms of stock recruitment relationships, particularly those developed by Ricker (1954) indicate a negative effect of a very large spawning stock size, this has not been conclusively observed for the species discussed here in the Northwest Atlantic or for similar species in the Northeast Atlantic. On the other hand, there have been strong indications that at very low spawning stock sizes there is a reduction in the number of large year classes and an increase in the proportion of very small year classes although cause and effect are not clear given the paucity of data and its confounding with the lack of independence of adjacent years observations. Thus, there is a concern to provide advice to managers of stock sizes which cause potential problems. Sometimes this has been done empirically as for mackerel, in which case scientists recommend, because of the lack of specific information, that stock sizes not be reduced below those that existed prior to the production of the large year classes. From a practical standpoint this did prove to be a stock size from which in later years the mackerel stock began to rebuild. However, there is no evidence to indicate that the stock would not have begun to rebuild even if stock size had been driven to a lower level prior to the reduction of fishing effort.
Shepherd (1982) described a generalized stock recruitment relationship, which by varying parameters can represent non-asymtotic, asymtotic, and dome stock recruitment relationships but more importantly addressed its practical applications. From the practical management standpoint he suggested that given the data available that the fits of the critical parameters could be just as accurately obtained from crude procedures as from sophisticated nonlinear curve fitting. Shephard states that the important question is the left hand slope of the line where according to stock recruitment hypotheses average recruitment would drop rapidly as stock size decreased. This left hand limb can be determined by drawing a straight line through the origin just to the left of the bulk of the data available. The slope of this line gives the parameter a of Shephard's stock recruitment function and allows the estimate of the left hand limb. This was done for the North Sea herring and provides a practical way of obtaining a minimum spawning stock size to recommend to managers.
A variation of the stochastic approach of Hennemuth et al. (1980) would assign probabilities to given levels of recruitment by segmenting the data into components of stock size groups and calculating probabilities of recruitment size for different ranges of spawning stock sizes. However, frequently the data bases are too small to allow such analyses.
Brown and Halliday (1983) reviewed the observed recruitment/population trends for a number of stocks in the Northwest Atlantic here and indicated that in a number of cases there was circumstantial evidence of lower recruitment from low stock sizes. This was observed not only for Georges Bank haddock where only one strong year class occurred during a long period of low spawning stock sizes under a wide variety of environmental conditions but for other stocks as well. For herring, no year class of any strength has recruited to the Georges Bank since 1970 during a period when the spawning stocks except for 1974, were considerably below (1/10) those of the early 1960's. In contrast to Georges Bank, however, the Gulf of Maine spawning stock rebuilt in 1974 and, although subsequently reduced by fishing, did not reach extremely low levels (it stayed above 1/5th of 1960-70 peaks) and strong year classes were produced in 1976, 1977, and 1979. The present contrast is that the herring stock in the Gulf of Maine is in a robust condition while the Georges Bank spawning stock, fished to very low levels, has suffered complete collapse.
Silver hake similarly shows such a contrast. The Georges Bank stock, now less than 10% of the 1960's, has been producing only poor year classes since 1970 while in the southern New England - Middle Atlantic silver hake spawning stock, now about 20% of its peak in 1965, has been producing year classes significantly larger than those on Georges Bank. The mackerel stock as discussed earlier was stabilized at about 20% of its peak size and has initiated a recovery trend.
Except for the question of the very small stock sizes examination of stock recruitment relationships has not been of help in understanding year class variability. This multispecies recruitment problem was identified within the Northwest Atlantic as the most important scientific problem more than two decades ago (Galtsoff, 1962).
Statistical studies such as Sissenwine's (1977) study of yellowtail flounder have been used to relate environmental conditions to recruitment. However, like spawning stock size, environmental correlations tend not to be precise predictors and often do not hold up over time. Thus, evidence is accumulating that recruitment is probably a multiplicative function of a highly variable process occupying various life stages during the first year of life.
Traditionally, fisheries ecologists have hypothesized that variability in year class strength reflects relatively small changes in very high mortality rates during the ichthyoplankton stage and thus have focused on the earlier life history stages. Extensive programs have been carried out in the Northwest Atlantic, both field surveys and experimental studies. Recently, Sissenwine et al. (in press) demonstrated that on Georges Bank predation by a few key predators (e.g. silver hake and cod) probably results in high postlarval fish mortality. Furthermore, preliminary examination of larval abundance data and estimates of egg production in relation to recruitment indicate mortality at the late larval and postlarval stages of several Northwest Atlantic species (e.g. Georges Bank haddock, herring) is of equal magnitude to mortality rates of eggs and larvae. Furthermore, virtually none of the variability in year class strength has been explained by the abundance of larvae. Thus the evidence however limited points to the postlarvae and early juvenile period where the effects of predation can be expected to be significant to be critical to the determinations of year class variability as observed at juvenile 1-2 year ages.
DISCUSSION
During the past 25 years, fisheries in NAFO Subareas 5 and 6 responded to the occurrence of strong year classes. The 1960 and 1961 year classes of herring established the abundance on which the offshore fishery developed. A very large 1963 year class of haddock attracted additional US and Canadian effort and new USSR effort. Given the mixed species nature of bottom trawl fisheries, this resulted in increased mortality to cod and other demersal species. The distant water fleets (primarily the USSR) next turned to silver hake which was highly abundant as a result of strong year classes which entered the population in the early 1960's (the exact years differ between stocks). The silver hake fishery used small mesh nets fishing on or close to the bottom which were capable of catching demersal fish as young as age 1, in contrast with the traditional US and Canadian trawl fisheries which used larger mesh, most effective for demersal species 2.5 years and older. As a result of the small mesh, the silver hake fishery contributed significantly to mortality in young fish of many species. After the silver hake fishery developed the next major resource subjected to greatly increased fishing pressure was mackerel, where the large 1966 and 1967 year classes attracted the distant water fleets.
The direct effect of fishing on these resources is relatively easily measured. The larger year classes that attracted these increases in effort would have sustained populations of considerably larger size for much longer periods of time had removals been kept at modest levels. There is no question that if fisheries managers desired a more stable catch and catch per unit effort, a moderate fishing level would have achieved that goal for the above resources.
The question, however, obviously arises whether the extremely heavy fishing pressure with its depression of spawning stock size had any effect on the length of time between strong year classes. The evidence is inconclusive. Nevertheless, as pointed out earlier in the species discussions and in the stock recruitment section, there is strong circumstantial evidence for the effect of reduction in biomass to extremely low levels (less than 20 and in some cases lower than 10% of peak abundance levels) being followed by less frequent occurrence of strong year classes.
In addition to the single species effect of fishing, either directly on the fishery resource yields or on future recruitment, there is the multispecies question. The evidence for a multispecies relationship again is both circumstantial and difficult to explain biologically. Nevertheless, the circumstantial evidence points to a greater stability in total biomass than in individual species, particularly when the pelagic species are considered. It also points to maintaining high levels of total biomass through shifts from originally herring, to mackerel, to squid and then to Ammodytes. There has been an increase in recent years of dogfish and bluefish after a decline in silver hake, resulting in total fish predators maintaining a relatively stable total predatory biomass. This shift has had a qualitative change toward predators able to feed on larger fish. The mackerel-herring substitution appears not only in this recent period, but there is some indication of similar situations in the past century as discussed in the review of Skud's (1982) paper. However, this must still be considered circumstantial as no biological hypothesis can yet be substantiated.
Attempts to find environmental correlates that hold true over time have not yet proven very successful. Yet, like the multispecies interactions, there are obviously relationships. The lesson here is that these relationships are difficult to hypothesize and interpret based on empirical data, just as there has been failure to date to directly demonstrate multispecies interactions.
The strongest lesson learned to date is the importance of looking at the total fishery resource ecosystem to the fullest extent possible, and understanding the wide variability that is normal. Table 3 and Figure 20 summarize the current and historical biomass conditions for major Northwest Atlantic fisheries (Serchuk et al., 1982b). This figure also indicates where in the current judgement of the Northeast Fisheries Center's fishery scientists concern should be expressed about low stock sizes relative to the immediate future productivity of these resources, as well as indications where evidence exists for low spawning stock sizes having actually had an effect on this productivity. Several general, but important, relationships are discernable from this summary.
1. For most all stocks the maximum observed biomass is generally no larger than four to five times the median, while the lower observed level may be proportionally much loweroften as low as 5 to 10 percent of the median. This pattern suggests that biological and ecological factors probably constrain the upper limit of stock biomass, but that exploitation can have a marked effect at reducing stock levels to their lowest bounds.
| Stock | Biomass | Current | Biomass | Current Fishery | ||||
|---|---|---|---|---|---|---|---|---|
| Median | Maximum | Minimum | Value | Units | Index | Primary data source | Management Status | |
| Gulf of Maine | 8.2 | 33.6 | 2,0 | -.8 | kg/tow | Autumn | Clark et al. 1982a | Managed under Fishery |
| Haddock | (1963-1981) | (1963) | (1972) | (1981) | (0+) | survey | Management Plan (FMP) | |
| Gulf of Maine | 9.4 | 14.1 | 4.2 | 13.1 | kg/tow | Autumn | Serchuk et al. 1982a | Managed under FMP |
| Cod | (1963-1980) | (1964) | (1976) | (1980) | (0+) | survey | ||
| Gulf of Maine | 75 | 194 | 16 | 30 | 10-3tons | VPA | Almeida and Anderson | No USA management |
| Silver hake | (1955-1981) | (1955) | (1971) | (1981) | (2+) | 1981a | FMP in preparation | |
| Gulf of Maine | 4.3 | 11.1 | 2.2 | 10.0 | kg/tow | Autumn | Serchuk 1982 | No USA management FMP |
| American plaice | (1963-1981) | (1980) | (1972) | (1981) | (0+) | survey | Unpublished data | in preparation (ADF Plan) |
| Gulf of Maine | 2.5 | 4.5 | 1.0 | 2.5 | kg/tow | Autumn | Clark 1982 | No USA management FMP |
| with flounder | (1963-1980) | (1966) | (1976) | (1980) | (0+) | survey | Unpublished data | in preparation (ADF Plan) |
| Pollock | 4.8 | 6.8 | 2.7 | 6.3 | kg/tow | Spring | Clark et al. 1981 a | No USA management FMP |
| (1968-1981) | (1967) | (1969) | (1981) | (0+) | survey | in preparation (ADF Plan) | ||
| Redfish | 23.5 | 54.6 | 12.2 | 12.2 | kg/tow | Autumn | Mayo 1980 | No USA management FMP |
| (1963-1981) | (1964) | (1981) | (1981) | (0+) | survey | in preparation (ADF Plan) | ||
| Georges Bank | 153 | 427 | 14 | 67 | 10-3tons | VPA | Clark et al. 1982a | Managed under FMP |
| Haddock | (1933-1960) | (1965) | (1973) | (1982) | (2+) | |||
| Georges Bank | 8.7 | 21.5 | 4.9 | 17.5 | kg/tow | Autumn | Serchuk et al. 1982 a | Managed under FMP |
| Cod | (1963-1981) | (1978) | (1969) | (1981) | (0+) | survey | ||
| Georges Bank | 4.9 | 10.6 | 1.2 | 2.3 | kg/tow | Autumn | Clark et al. 1981b | Managed under FMP |
| Yellowtail flounder | (1963-1981) | (1964) | (1976) | (1981) | (0+) | survey | ||
| Georges Bank | 143 | 595 | 26 | 32 | 10-3 tons | VPA | Almeida and Anderson | Managed under preliminary |
| Silver hake | (1955-1981) | (1964) | (1975) | (1981) | (2+) | 1981a | Fishery Management Plan | |
| (PMP) | ||||||||
| Georges Bank | 0.8 | 2.4 | 0.1 | 0.1 | kg/tow | Spring | Almeida and Anderson | Managed under PMP |
| Red hake | (1970-1982) | (1971) | (1982) | (1982) | (0+) | survey | 1981b | |
| So. New England | 9.2 | 20.1 | 0.7 | 3.8 | kg/tow | Autumn | Clark et al. 1981b | Managed under FMP |
| Yellowtail flounder | (1963-1981) | (1972) | (1975) | (1981) | (0+) | survey | ||
| So. New England- | ||||||||
| Mid-Atlantic | 93 | 375 | 57 | 61 | 10-3 tons | VPA | Almeida and Anderson | Managed under PMP |
| Silver hake | (1955-1981) | (1965) | (1958) | (1981) | (2+) | 1981a | ||
| So. New England- | ||||||||
| Mid-Atlantic | 94 | 174 | 30 | 38 | 10-3tons | VPA | Almeida and Anderson | Managed under PMP |
| Red hake | (1963-1981) | (1965) | (1977) | (1981) | (2+) | 1981b | ||
| So. New England- | ||||||||
| Mid-Atlantic | 6.2 | 12.4 | 1.1 | 3.2 | kg/tow | Spring | Mayo 1982 | No USA management |
| Scup | (1968-1981) | (1979) | (1969) | (1981) | (0+) | survey | FMP planned | |
| Summer flounder | 0.3 | 1.0 | 0.05 | 0.4 | kg/tow | Spring | Fogarty 1981 | No USA management |
| (1968-1980) | (1976) | (1970) | (1980) | (0+) | survey | FMP in preparation | ||
| Bluefish | 0.7 | 1.5 | 0.01 | 0.6 | kg/tow | Autumn | Anderson 1980 | No USA management |
| (1967-1981) | (1974) | (1968) | (1981) | (0+) | Survey | FMP developed | ||
| Gulf of Maine | 103 | 175 | 34 | 83 | 10-3tons | VPA | Fogarty & Anthony 1982 | Managed under FMP |
| Herring | (1963-1981) | (1968) | (1973) | (1981) | (4+) | through mid-1982. | ||
| Georges Bank | 342 | 1143 | Near 0 | Near 0 | 10-3tons | VPA | Anthony & Waring 1980 a | Managed under FMP |
| Herring | 1961-1977) | (1967) | (1981) | (1981) | (4+) | through mid-1982. | ||
| Atlantic | 915 | 2827 | 567 | 780 | 10-3tons | VPA | Anderson 1981 | Managed under FMP |
| mackerel | (1962-1981) | (1970) | (1962) | (1981) | (1+) | |||
| Butterfish | 5.5 | 20.6 | 2.9 | 5.2 | kg/tow | Autumn | Waring & Anderson 1981 | Managed under FMP |
| (1968-1982) | (1980) | (1975) | (1982) | (0+) | survey | |||
| Loligo squid | 28.3 | 51.4 | 14.1 | 24.3 | 10-3tons | Autumn | Lange 1982 | Managed under FMP |
| (1968-1981) | (1976) | (1971) | (1981) | (0+) | survey | |||
| Illex squid | 5.4 | 68.6 | 0.4 | 68.6 | 10-3tons | Autumn | Lange 1982 | Managed under FMP |
| (1968-1981) | (1981) | (1969) | (1981) | (0+) | survey | |||
| Total Finfish | ||||||||
| and squids | 3416 | 8012 | 1934 | 3358 | 10-3tons | VPAS and | Clark and Brown 1979 | No USA management as a unit. |
| Gulf of Maine- | (1965-1981) | (1968) | (1975) | (1981) | Surveys | |||
| Cape Hatteras | ||||||||
| Georges Bank | 1.2 | 2.5 | 0.6 | 0.6 | kg/tow | Scallop | Serchuk et al. 1982 c | Managed under FMP |
| Sea scallops | (1975-1982) | (1978) | (1982) | (meat) | survey | |||
| (>70 mm) | ||||||||
| Mid-Atlantic | 0.3 | 0.7 | 0.18 | 0.2 | kg/tow | Scallop | Serchuk et al. 1982c | Managed under FMP |
| Sea scaliops | (1975-1982) | (1978) | (1981) | (1982) | (meat) | survey | ||
| (>70 mm) | ||||||||
| Mid-Atlantic | 2.5 | 4.5 | 0.5 | 4.3 | kg/tow | Surf | Murawski & Serchuk 1981 | Managed Under FMP |
| Surf Clam | (1965-1982) | (1982) | (1977) | (1982) | (meat) | clam | ||
| (0+) | survey | |||||||
| Offshore American | 0.8 | 1.3 | 0.5 | 0.9 | kg/tow | Autumn | Fogarty et al. 1982 | FMP prepared |
| Lobster | (1963-1981) | (1964) | (1970) | (1981) | (0+) | survey | ||
| Gulf of Maine | 5.8 | 45.8 | 1.6 | 3.0 | kg/tow | Maine | Clark 1982 | Managed by States |
| Northern Shrimp | (1972-1981) | (1968) | (1977) | (1981) | (0+) | summer | under ASMFC | |
| survey | ||||||||
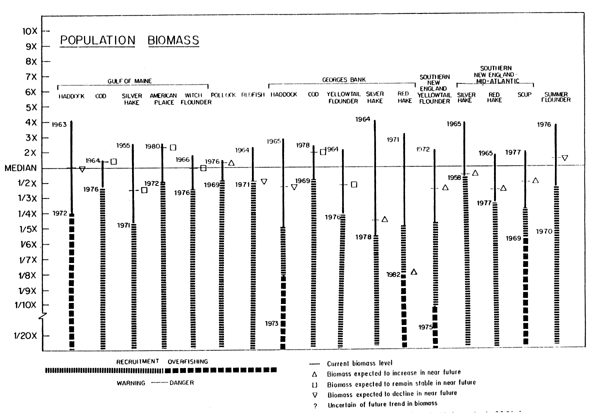
Fig. 20. Graphical summary of biomass patterns for major Northwest Atlantic finfish and shellfish ressources (Serchuk et al., 1982b).
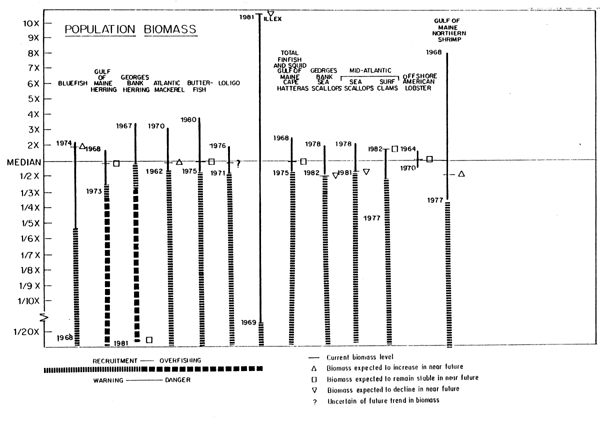
Fig. 20. Contimued
2. The range of variability of biomass appears to be as large in demersal as it is in pelagic species. Relatively short-lived species such as squid, are probably more affected by environmental conditions causing large variations of biomass than longerlived species like haddock or herring where biomass variations are also influenced by the number of age groups in the stock. When fishing activity reduces the number of age groups in the stock, as was the case for the Georges Bank haddock in the late 1960's and Georges Bank herring in the 1970's, large variations in biomass predominantly reflect recruitment success for single year classes, a situation normally observed in short-life species. It should be noted as we discussed in the species discussion that the expressed variations in Illex squid biomass is also a function of availability as this species often migrates out of the geographical range covered by Northeast Fisheries Center assessment surveys.
3. Currently more stocks appear to be increasing in biomass than decreasing. This is probably the result of the substantial reduction in foreign fishing effort in the past six years. Under more moderate fishing mortality circumstances, one might expect as many stocks decreasing in biomass as increasing. The difference among stocks in the time (years) at which maximum/minimum biomass levels were observed suggests such an expected pattern of biomass changes and the unlikelihood of all stocks being high at the same time.
4. For stocks harvested together in single fisheries, the relationship of current biomass and the expected direction of the biomass change with respect to the median and the warning and danger zones appear quite different among stocks. This suggests that simultaneous increases of biomasses of all components in the mixed fisheries is unlikely due to differential fishing mortalities generated by the current fishery on each component stock.
IMPLICATIONS FOR RESEARCH
Galtsoff (1962) over 20 years ago wrote, "the most important scientific phase of this research is concerned with the causes of the great fluctuations in abundance of fisheries stocks. Life in the ocean is in a state of unstable equilibrium in which the struggle for existence gives temporary predominance to one group which in turn may be replaced by another the intricate picture of life in the open ocean cannot be illustrated by a single short-term observation of restricted area---Long and well planned observations are needed." This approach has been the basis for the research of the Northeast Fisheries Center for a quarter of a century and the results of this research form the basis for the paper presented here.
The guidelines of Galtsoff remain true today. It is vital to have a multipronged multidisciplinary research approach that addresses the total resource ecosystem if one is going to address the multispecies variability as it relates to the resource productivity. It is useful to examine the research program of the NEFC which has followed these guidelines and indicate what aspects have proven most valuable and what aspects require targeting at the present time in addition to maintaining a basic monitoring program.
This total program includes monitoring the catch, not only the total amount but also measures of quality whether by size, age, or both. It is also extremely valuable to have a regular measure of stock abundance of older juveniles and adults independent of the fishery. NEFC trawl surveys provide these indices. The precision of these indices certainly varies among species, but major trends can be determined for an extremely wide variety of species whether pelagic, semipelagic or demersal. It is important that survey coverage encompasses a fairly significant portion of the range of the resource to allow regular interpretation.
The NEFC ecosystem research program also monitored the basic physical oceanographic conditions, the icthyoplankton and the food habits of the fish taken. Zooplankton studies have indicated relative stability over long periods of time (Sherman et al., in press). Within the sampling network of the multispecies ichthyoplankton surveys, studies are conducted of the factors controlling growth and survival of target species, e.g., cod and haddock. Laboratory studies are now underway to confirm laboratory determinations of optimal prey densities with at-sea experiments on Georges Bank on the availability and abundance of suitable densities of zooplankton prey of cod and haddock. Plans are also being prepared to conduct predation experiments on eggs and larvae in large enclosures. Preliminary observations indicate that larval growth and survival is very high in predatorfree, large, flow-through net mesh enclosures placed in a highly productive estuarine environment (Laurence et al., 1979).
Studies have also begun to monitor the incidence of disease. We are not yet at the stage where one can-indicate their importance in the monitoring of natural mortality except, of course, in the classical incidences of massive die-offs from epizootics.
Pollution is an important concern in heavily populated areas of the northeastern United States. Recent studies have been conducted on cytogenic abnormalities in mackerel embryos in relation to concentrations of certain pollutants in the New York Bight region (Longwell, 1976, 1978; Longwell and Hughes, 1980). The egg stage may be most critical from the standpoint of the impact of man's pollutants which fish have not evolved to, as opposed to the variation in natural environmental factors. In cooperation with the U.S. Fish and wildlife Service, studies were made by the NEFC (Buckley4) of the viability of striped bass (Morone saxatilis) larvae from parents exposed to heavy metals and other toxins. Initial results indicate significant impact of the exposure to larvae hatched from the Hudson River parent stock.
Other pollution-related studies underway by NEFC in cooperation with the U.S. Environmental Protection Agency are focused on the impacts of exposures of larvae to urban sludge compounds identical to those now being disposed of on the continental shelf by the State of New York. One of the encouraging advances to study these effects at the NEFC is the use of RNA/DNA analysis for determining the growth potential of larvae collected routinely during the MARMAP surveys. High ratios indicate a "healthy" physiological condition for the larvae, while low values indicate that the growth potential of the larvae is impaired (Buckley, 1980, 1982).
Oceanographic studies have attempted to look at the possibility of major events such as advection of shelf water and associated shelf icthyoplankton. Warm core rings from the Gulf Stream have been observed entraining large volumes of shelf water across the shelf slope front into a nutrient-poor environment. It has been hypothesized that eggs and larvae of shelf species advected off the highly productive shelf in an entrainment feature would not survive in the prey-poor environment. Collections made in an entrainment experiment on the survival of ichthyoplankton in an entrainment feature revealed that no shelf species were in the entrainment, suggesting that warm core rings are not responsible for advective mortality of shelf larvae. Replicate experiments are planned to confirm these preliminary results (Laurence and Burns, 1982). A number of correlation studies have examined year classes on Georges Bank with environmental factors, such as offshore winds, slope frontal water and warm core rings and so far no relationship has been found that holds up over time. This may mean that significant larval mortality may occur at various times throughout the first year of life.
Although the survival of an individual larva is effected primarily by its own microenvironment, the extremely large differences between year class size (regularly 10 to 1 for many species) means that for environmental effects to be significant they must operate over the entire area that the larvae occupy. The accumulation of independent factors from microenvironments would average out over the area and thus not give the wide fluctuations with the regularity that they are observed. Further theoretical statistical studies could be of use in evaluating this hypothesis and to determine the relative sizes of areas for which it would be useful to evaluate environmental conditions if one is to examine year class survival.
The intensive efforts that have been made in the Northwest Atlantic to relate egg and larval work to predictions of recruits indicate that this procedure is not likely to be successful. These surveys are useful for other purposes such as independent estimates of spawning stock size in some situations and for indicating areas and times of spawning.
4 Laurence J. Buckley, NEFC, Narragansett Laboratory, Narragansett, RI, personal communication.
The results to date would indicate that the next step in NEFC research will be to target on postlarval fish and to focus more resources on the actual recruitment processes. This understanding is most important for elucidating general principles that can be used for determining strategies of management. Logistical problems make monitoring unlikely except in very unusual situations for use in tactical management. This research effort will involve focusing on sampling at the postlarval and early juvenile stage and in evaluating the effects of predation. Here again this means extensive continuation of food habits studies of species most likely to prey on young fish. This does not mean that it is not necessary to continue at some level the background work of egg and larval surveys, oceanographic conditions, and even increased efforts on factors that affect the production of eggs by adult fishes such as changes in fecundity with stock size. It does, however, mean a concentration of NEFC resources on the postlarval period and the effects of fish predation thereon in the next period of years with reduced efforts in the other areas. An important aspect of monitoring mortality at this stage is that it should give strategic information concerning ranges of variability and the potential year class fluctuations under a variety of species compositions. Such efforts involve both field studies and modeling efforts. In long range research considerations, in order for further development of an ecosystem approach to the management of fishery resources, it will be necessary to overcome present deficiencies in:
(1) Identifying the linkages between primary, secondary, and fish production;
(2) Quantifying predator-prey dynamics, and
(3) Understanding the relationship between stock-size and recruitment.
IMPLICATIONS FOR MANAGEMENT
Rarely in a fisheries management situation is the information available that would be required to provide very precise advice to managers. Precise advice would allow managers to weigh the risks involved in the decisions they are required to make and to have a high confidence in the risks being taken. Traditionally, fisheries scientists have often given relatively conservative advice based on their knowledge of other situations where fisheries have collapsed under increasing and intense fishing pressure. The decision-makers in industry and government, on the other hand, have observed situations where despite increases in fishing, productivity was maintained and thus have tended towards an optimistic view. Frequently management decisions have turned out to be a compromise between these two positions.
The experiences over the past 25 years in the southern area of the Northwest Atlantic add to the accumulative total of information which comprise the background of fisheries scientists. This accumulated information allows fishery scientists to be somewhat more explicit in their advice concerning the historically observed consequences of extremely heavy perturbations in fishing on highly variable resources. This leads to a greater refinement in the types of risk statements the fisheries managers can interpret and utilize in making their decisions.
In the Northwest Atlantic total fishing pressure has been demonstrated to be so large that resource productivity can be reduced on an overall basis. This occurred both because of the desire to harvest a large number of species and the use of non-selective fishing gear. The level of reduction of this resource from an unfished state was an estimated 70% (Clark and Brown, 1977).
Further implication for management comes from observing the recovery of this resource after fishing effort was drastically reduced. One of the areas that managers have to consider is the time-frame for recovery. For the fisheries off the northwest coast of the US there is approximately 10 years of history of intensive management, the last 5 being very minimal fishing for many of these stocks. In the period between 5 and 10 years after reaching bottom levels, the total biomass shows partial recovery. The recovery has been greater for the demersal fisheries than for the pelagic species. The latter should be tempered with the understanding that the very high peak observed in the mackerel stock in 1971 may occur only occasionally. Nevertheless, herring on Georges Bank are virtually non-existent and silver hake on Georges Bank are at a very low level despite a relatively long period of constraints.
The species composition of the demersal fish community resembles very much that which existed prior to heavy fishing, and one does not observe change in any major sense. Conversely, in the pelagic stocks major changes occurred with the silver hake component, the predator of largest biomass in the 1960's, being replaced primarily with dogfish and with bluefish to a lesser degree. Thus the total predator biomass remained more constant even though silver hake did not. In addition, the failure of herring to return and the explosion of sand lance makes the composition of the pelagic resources quite different than it was earlier. Therefore, managers should be warned that extremely heavy fishing not only can result in depression of the resources with relatively long time frames for recovery, but at least on a short-term basis, the recovery may be in resources that are somewhat different in composition than existed earlier.
In examining species individually, it can be seen that the greatest declines in herring nave taken place on Georges Bank in contrast with the Gulf of Maine, in silver hake on Georges Bank in contrast to Southern New England, in the haddock on Georges Bank in contrast to the cod stock. It should be noticed in all of these contrasts that the stocks that have reached the very lowest levels have not recovered, or have recovered to a moderate degree, such as haddock. In these cases, spawning stock was down to a very low level, approximately only 10% or less of that observed at the peak periods. Those herring and silver hake stocks which did reach such low levels but were reduced to only approximately 20 to 25% of peak levels are now recovering. Cod was reduced only moderately from peak levels but recovered quickly. Mackerel abundance was stabilized at the abundance level observed in the early 1960's which proved to be about 20% of that observed at the peak levels. These inferences are quite crude, but nevertheless, the histories of such disastrous reductions serve as warning flags for fishery managers.
Finally, we should note the situation with the haddock recovery in which industry demands, when recovery was fueled by a single year class (1957), created the situation in which the spawning stock size was not rebuilt up to a robust multiage group state. Therefore, a vigorous fishery removed most of this abundant year class and the year class (1978) which followed from the first spawning of the 1975 year class, so rapidly that recovery, although large, was brief, and the stock is once again declining to the lower levels of abundance observed earlier. This situation was complicated for fisheries managers by the mixed nature of the fishery, particularly the relatively abundant populations of cod. Small quotas in place for haddock were caught quickly and the large populations of haddock made it impossible to pursue a fishery for cod and other groundfish without greatly exceeding haddock quotas designed to allow only very low fishing mortality rates so that haddock spawning stock biomass could be rebuilt. Fishery scientists are now in a position of pointing out to managers and industry officials an historic case where observing a recovery and taking immediate advantage of it initially may result in that recovery being very short in duration.
REFERENCES
Almeida, F.P. and E.D. Anderson. 1981a. Status of the silver hake resource off the northeast coast of the United States - 1981. NMFS, NEFC, Woods Hole Lab.Ref.Doc. No. 81-36, 78 p. (Mimeo).
Almeida, F.P. and E.D. Anderson. 1981b. Status of the red hake resource off the northeast coast of the United States - 1981. NMFS, NEFC, Woods Hole Lab.Ref.Doc. No. 81-37, 49 p. (Mimeo).
Anderson, E.D. 1980. A preliminary assessment of the status of bluefish (Pomatomus saltatrix) along the Atlantic coast of the United States. NMFS, NEFC, Woods Hole Lab.Ref.Doc. No. 80-30, 29 p. (Mimeo).
Anderson, E.D. 1981. Status of the Northwest Atlantic mackerel stock - 1981. NMFS, NEFC, Woods Hole Lab.Ref.Doc.No. 81-38, 40 p. (Mimeo).
Anderson, E.D. 1982. Status of the Northwest Atlantic mackerel stock - 1982. NMFS, NEFC, Woods Hole Lab.Ref.Doc.No. 82-44, 45 p. (Mimeo).
Anderson, E.D., F.E. Lux and F.P. Almeida. 1980. The silver hake stocks and fishery off the Northeastern United States. Mar.Fish.Rev. 42(1):12-20.
Anderson, E.D. and A.J. Paciorkowski. 1980. A review of the Northwest Atlantic mackerel fishery. Rapp.P.-v.Reun.Cons.int.Explor.Mer, 177:175-211.
Anthony, V.C. 1972. Population dynamics of the Atlantic herring in the Gulf of Maine. Ph.D. thesis, Univ.Wash., 266 p.
Anthony, V.C. and G. Waring. 1980a. The assessment and management of the Georges Bank herring fishery. Rapp.P.-v.Reun.Cons.int.Explor.Mer. 177:72-111.
Anthony, V.C. and G.T. Waring. 1980b. A review of the herring fisheries, their assessment and management in the Georges Bank-Gulf of Maine area. p.115-178. In Proc. Alaska Herring Symposium. (B.R. Melteff and V.G. Wespestad, eds.) Alaska Sea Grant Rept. 80-4, 274 p.
Azarovitz, T.R. 1981. A brief historical review of the Woods Hole Laboratory trawl survey time series. p.62-67. In Bottom trawl surveys. (W.G. Doubleday and D. Rivard, eds.) Can.Spec.Publ.Fish.Aquat.Sci. 58, 273 p.
Beverton, R.J.H. and S.J. Holt. 1957. On the dynamics of exploited fish populations. Fishery Invest., London. Ser. 2, 19, 533 p.
Brown, B.E., J.A. Brennan, M.D. Grosslein, E.G. Heyerdahl and R.C. Hennemuth. 1976. The effect of fishing on the marine finfish biomass in the Northwest Atlantic from the Gulf of Maine to Cape Hatteras. Int.Comm.Northw.Atlant.Fish.Res.,Bull. 12:49-68.
Brown, B.E. and R.G. Halliday. 1983. Fisheries resources of the Northwest Atlantic - responses to extreme fishing perturbations. Can.Spec.Publ.Fish.Aquat.Sci.
Buckley, L.J. 1980. Changes in ribonucleic acid, deoxyribonucleic acid and protein content during ontogenesis in winter flounder, Pseudopleuronectes americanus, and effect of starvation. U.S.Fish.Bull. 77:1703-1708.
Buckley, L.J. 1982. Effects of temperature on growth and biochemical composition of larval winter flounder, Pseudopleuronectes americanus. Mar.Ecol. - Progr.Ser. 8:181-186.
Clark, S.H. 1981. Use of trawl survey data in assessments. p. 82-92. In Bottom trawl surveys. (W.G. Doubleday and D. Rivard, eds.) Can.Spec. Publ.Fish.Aquat.Sci. 58, 273 p.
Clark, S.H. 1982. Assessment and management of the Gulf of Maine northern shrimp (Pandalus borealis) fishery. Cons.int.Explor.Mer. CM 1982/K:13, 20 p. (Mimeo).
Clark, S.H. and B.E. Brown. 1977. Changes in biomass of finfishes and squids from the Gulf of Maine to Cape Hatteras, 1963-74, as determined from research vessel survey data. U.S.Fish.Bull. 75(1):1-21.
Clark, S.H. and B.E. Brown. 1979. Trends in biomass of finfishes and squids in ICNAF Subarea 5 and Statistical Area 6, 1964-1977, as determined from research vessel survey data. Inv.Pesq. 43(1):107-122.
Clark, S.H., L. O'Brien and R.K. Mayo. 1981a. Scotian Shelf, Gulf of Maine and Georges Bank pollock stock status - 1981. NMFS, NEFC, Woods Hole Lab.Ref.Doc.No. 81-32, 38 p. (Mimeo).
Clark, S.H., 1981b. Yellowtail flounder stock status - 1981. NMFS, NEFC, Woods Hole Lab.Ref.Doc.No. 81-10, 47 p. (Mimeo).
Clark, S.H., R.K. Mayo and A. Green. 1982a. Georges Bank and Gulf of Maine haddock stock status - 1982. NMFS, NEFC, Woods Hole Lab.Ref.Doc.No. 82-32, 39 p. (Mimeo).
Clark, S.H., W.J. Overholtz and R.C. Hennemuth. 1982b. Review and assessment of the Georges Bank and Gulf of Maine haddock fishery. J.Northw.Atl.Fish.Sci. 3:1-25.
Cohen, E.B., M.D. Grosslein, M.P. Sissenwine, F. Steimle and W.R. Wright. 1982. Energy budget of Georges Bank. p. 95-197. In Multispecies approaches to fisheries management advice. (M.C. Mercer, ed.) Can.Spec.Publ.Fish.Aquat.Sci. 59, 169 p.
Crisp, D.J. 1975. Secondary productivity in the sea. p. 71-90. In Productivity of world ecosystems. (D.E. Reichle, J.F. Franklin and D.W. Goodal, eds.) Nat.Acad.Sci., Wash. D.C.
Daan, N. 1980. A review of replacement of depleted stocks by other species and the mechanisms underlying such replacement. Rap.P.-v.Reun.Cons.int.Explor.Mer. 177:405-421.
Dow. R.L. 1977. Effects of climatic cycles on the relative abundance and availability of commercial marine and estuarine species. J.Cons. 37(3):274-280.
Fogarty, M.J. 1981. Review and assessment of the summer flounder (Paralichthys dentatus) in the Northwest Atlantic. NMFS, NEFC, Woods Hole Lab.Ref.Doc.No. 81-25, 57 p. (Mimeo).
Fogarty, M.J. and V.C. Anthony. 1982. Status of Atlantic herring resources in the Gulf of Maine region - 1982. NMFS, NEFC, Woods Hole Lab.Ref.Doc.No. 82-34, 35 p. (Mimeo).
Fogarty, M.J., R.A. Cooper, J.R. Uzmann and T. Burns. 1982. Assessment of the USA offshore American lobster (Homarus americanus) fishery. Cons.int.Explor.Mer. CM 1982/K14, 21 p. (Mimeo).
Foster, K.L. 1982. An application of a multispecies cohort analysis to six fish stocks located on Georges Bank. Cons.int.Explor.Mer. CM 1982/G:37, 21 p. (Mimeo).
Galstoff, P.S. 1962. The story of the Bureau of Commercial Fisheries Biological Laboratory, Woods Hole, Massachusetts. U.S.Fish.Wildl.Serv.Circ. 145, 121 p.
Grosslein, M.D. 1969. Groundfish survey program at BCF Woods Hole. Comm.Fish.Rev. 31(8-9):22-35.
Grosslein, M.D., R.W. Langton and M.P. Sissenwine. 1980. Recent fluctuations in pelagic fish stocks of the Northwest Atlantic Georges Bank region, in relationship to species interactions. Rapp.P.-v.Reun.Cons.int.Explor.Mer. 177:374-404.
Hennemuth, R.C. 1979. Man as predator. p. 507-532. In Contemporary quantitative ecology and related ecometrics. (G.P. Patil and M.Rosenzevig, ed.) Int.Co-op.Publ.House, Fairland, MD.
Hennemuth, R.C., J.E. Palmer and B.E. Brown. 1980. A statistical description of recruitment in eighteen selected fish stocks. J.Northw.Atl.Fish.Sci. 1:101-111.
Jones, R. and J. Richards. 1976. Some observations on the interrelationships between the major fish species in the North Sea. Cons.int.Explor.Mer. CM 1976/F:35, 10 p. (Mimeo).
Lange, A.M.T. 1982. Status of the squid (Loligo pealei and Illex illecebrosus) populations off the northeastern USA - October 1982. NMFS, NEFC, Woods Hole Lab.Ref.Doc.No. 82-27, 21 p. (Mimeo).
Lange, A.M.T. and M.P.Sissenwine. (in press). Squid resources of the Northwest Atlantic. UN Food Agric.Org., Fish.Tech.Rept.
Laurence, G.C. and B.R. Burns. 1982. Ichthyoplankton in shelf water entrained by warm-core rings. AGU/ASLO. San Francisco, CA. 5-7 Dec. 1982. Abstract No. 032A-07. EOS 63(45):998.
Laurence, G.C., T. Halavik, B. Burns and A. Smigielski. 1979. An environmental chamber for monitoring "in situ" growth and survival of larval fishes. Trans.Am.Fish.Soc. 108:197-203.
Longwell, A.C. 1976. Mutagenesis in developing mackerel eggs sampled from the New York Bight. NOAA Tech.Memo. ERL-MESA-7, 59 p.
Longwell, A.C. 1978. Field and laboratory measurements of stress responses at the chromosome and cell levels in planktonic fish eggs and the oil problem. In the wake of the Argo Merchant. Proceedings of a Symposium held January 11-13, 1978. Center Ocean Mgmt. Studies, Univ. Rhode Island.
Longwell, A.C. and J.B. Hughes. 1980. Cytologic, cytogenetic and developmental state of Atlantic mackerel eggs from sea surface waters of the New York Bight and prospects for biological effects monitoring with ichthyoplankton. Rapp.P.-v.Reun.Cons.int.Explor.Mer. 179:275-291.
Mayo, R.K. 1980. Exploitation of redfish, Sebastes marinus (L.), in the Gulf of Maine-Georges Bank region, with particular reference to the 1971 year class. J.Northw.Atl.Fish.Sci. 1:21-37.
Mayo, R.K. 1982. An assessment of the scup, Stenotomus chrysops (L.), population in the Southern New England and Middle Atlantic regions. NMFS, NEFC, Woods Hole Lab.Ref.Doc. No. 82-46, 61 p. (Mimeo).
McBride, M.M., M.P. Sissenwine, B.E. Brown and L.M. Kerr. 1980. Yellowtail flounder (Limanda ferruginea): status of the stocks, March 1980. "NMFS" NEFC, Woods Hole Lab.Ref.Doc.No. 80- 20, 27 p. (Mimeo).
Morse, W. 1982. Spawning stock biomass estimates of sand lance, Ammodytes sp. off northeastern United States determined from MARMAP plankton surveys, 1974-1980. Cons.int.Explor.Mer, CM 1982/G:59, 11 p. (Mimeo).
Murawski, S.A and F.M. Serchuk. 1981. Assessment and current status of offshore surf clam, Spisula solidissima, populations off the Middle Atlantic coast of the United States. NMFS, NEFC, Woods Hole Lab.Ref. Doc. No. 81-33, 50 p. (Mimeo).
Pope, J.G. 1979. A modified cohort analysis in which constant natural mortality is replaced. Cons.int.Explor.Mer. CM 1979/H:16, 7 p. (Mimeo).
Resource Assessment Division. 1982. Status of the fisheries resources off the Northeastern United States for 1981. NOAA Tech.Memo. NMFS-F/NEC-12, 114 p.
Ricker, W.E. 1954. Stock and recruitment. J.Fish.Res.Bd.Can. 11:559-623.
Ricker, W.E. 1958. Handbook of computations for biological statistics of fish populations. Bull.Fish.Res.Bd.Can. 119, 300 p.
Robins, C.R. (ed.). 1980. A list of common and scientific names of fishes from the United States and Canada. Am.Fish.Soc.Spec.Publ. 12, 174 p.
Schaefer, M.B. 1954. Some aspects of dynamics of populations important to the management of commercial marine fisheries. Bull.Inter-Am.Trop.Tuna Comm. 1(2):27-56.
Serchuk, F.M., R.S. Rak and J. Penttila. 1982a. Status of the Georges Bank and Gulf of Maine cod stocks - 1982. NMFS, NEFC, Woods Hole Lab.Ref.Doc. Noc. 82-32, 46 p. (Mimeo).
Serchuk, F.M., M.P. Sissenwine, R.C. Hennemuth and B.E. Brown. 1982b. Summary of biomass trends and conditions for major Northwest Atlantic fisheries resources: a graphical approach. NMFS, NEFC, Woods Hole Lab.Ref.Doc. No. 82-42, 12 p. (Mimeo).
Serchuk, F.M., P.W. Wood and R.S. Rak. 1982c. Review and assessment of the Georges Bank, Mid-Atlantic and Gulf of Maine Atlantic sea scallop (Placopecten magellanicus) resources. NMFS, NEFC, Woods Hole Lab.Ref.Doc.No. 82-06, 32 p. (Mimeo).
Sette, O.E. and A.W.H. Needler. 1934. Statistics of the mackerel fishery off the east coast of North America. 1804 to 1930. U.S.Bur.Fish.Invest,Rept. 19, 48 p.
Shepherd, J.G. 1982. A versatile new stock-recruitment relationship for fisheries and the construction of sustainable yield curves. J.Cons. 40(1):67-75.
Sherman, K. 1980. MARMAP, a fisheries ecosystem study in the NW Atlantic: Fluctuations in ichthyoplankton-zooplankton components and their potential for impact on the system. p. 9-37. In Advanced concepts on ocean measurements for marine biology. (F.P. Diemer, F.J. Vernberg, D.Z. Mirkes, eds.) Belle W. Baruch Institute for Marine Biology and Coastal Research. Univ.South Carolina Press.
Sherman, K., C. Jones, L. Sullivan, W. Smith, P. Berrien and L. Ejsymont. 1981. Congruent shifts in sand eel abundance in western and eastern North Atlantic ecosystems. Nature 291(5815):486- 489.
Sherman, K., J.R. Green, J.R. Goulet and L. Ejsymont. (in press). Zooplankton coherence in a large Northwest Atlantic ecosystem. Science.
Sissenwine, M.P. 1977. A compartmentalized model of the southern New England yellowtail flounder, Limanda ferruginea, fishery. U.S.Fish.Bull. 75(3):465-482.
Sissenwine, M.P., B.E. Brown, J.E. Palmer and R.J. Essig. 1982. Empirical examination of population interactions for the fishery resources off the Northeastern USA. p. 82-94. In Multispecies approaches to fisheries management advice. (M.C. Mercer, ed.) Can.Spec.Publ.Fish.Aquat.Sci. 59, 169 p.
Sissenwine, M.P., E.B. Cohen and M.D. Grosslein. (in press). Structure of the Georges Bank ecosystem. Rapp.P.-v.Reún.Cons.int.Explor.Mer.
Skud, B.E. 1982. Dominance in fishes: the relation between environment and abundance. Sci. 216:144-149.
Steele, J.H. 1974. The structure of marine ecosystems. Harvard Univ.Press, Cambridge, MA. 128 p.
Sutcliffe, W.H.J., K. Drinkwater and B.S. Muir. 1977. Correlations of fish catch and environmental factors in the Gulf of Maine. J.Fish.Res.Bd.Can.34:19-30.
Taille, C and G.P. Patil. 1982. Modeling and analysis of recruitment distributions. Rept. Contract: NA-30-FAC-6040. Stochastics and Statistics in Marine Fisheries Research and Management. Dept.Stat.Penn.State Univ., University Park, PA. 92 p.
Waring, G.T. and E.D. Anderson. 1981. Status of the Northwestern Atlantic butterfish stock. NMFS, NEFC, Woods Hole Lab.Ref.Doc.No. 81-27, 24 p. (Mimeo).
![]()
![]()
![]()