![]()
![]()
![]()
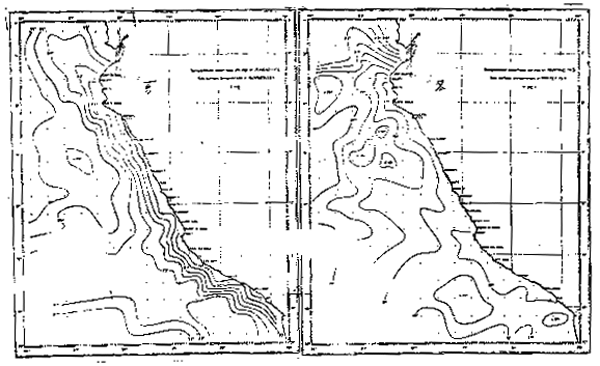
Fig. 4.- Temperatura promedio a Om en verano e invierno (de Zuta y Guillén, 1970).
Las temperaturas promedio frente al Ecuador, Colombia y Panamá (Enfield, 1976) presentan a lo largo de la costa un aumento de la temperatura hacia el norte, dando lugar a una zona de gradiente de temperatura o frente ecuatorial situado al norte de los 5°S. En invierno presenta una distribución casi zonal de temperatura máxima para las latitudes de 5°–8°N entre los meses de diciembre y abril y una distribución longitudinal de tempe– raturas mínimas para los meses de marzo y abril entre el Golfo de Panamá y 2°N.
Al sur de las Islas Galápagos se halla un área de baja temperatura debido a la surgencia ecuatorial, así como a la extensión hacia el oeste de la Corriente Peruana. Las variaciones de la temperatura son causadas principalmente por la variación estacional de la posición de la Zona de la Convergencia Intertropical (ZCIT) la cual puede hallarse frente a Panamá en el verano del hemisferio norte, o cerca al límite Colombia-Ecuador durante el invierno.
Salinidad
En el Pacífico Sudeste la salinidad máxima pasa de los 36.5 o/oo y la salinidad mínima se halla al noreste de las Galápagos con salinidades alrededor de 32 o/oo y frente a la costa meridional de Chile con salinidades alrededor de 33 o/oo.
La salinidad a lo largo de la costa (Inostroza, 1972) de Chile va aumentando hacia el norte (Fig.3) desde valores de 33 o/oo a 40°S a 35 o/oo al norte. La salinidad promedio al norte de los 34°S es de 35.2 a 33.9 o/oo y de 35.0 a 33.8 o/oo en verano e invierno respectivamente. Al sur de los 37°S se hallan los valores mí nimos de salinidad (<34 o/oo). En el verano además se observa una lengua de agua salina (>35 o/oo) al norte de los 23°S la cual se repliega en el invierno al norte de los 20°S.
Frente a las costas del Perú (Zuta y Guillén, 1970) la salinidad en la superficie del mar (Fig. 5) presenta gradientes zonales al sur de los 6°S, en donde la salinidad aumenta lejos de la costa. Al norte de los 6°S presenta gradientes longitudinales pronunciados, debido al efecto de las Aguas Ecuatoriales Superficiales y Aguas Tropicales Superficiales. La salinidad en la parte netamente costera es de 35.1 a 34.8 o/oo. El rango promedio de salinidad en el verano e invierno es de 35.6 a 33.7 o/oo y de 35.5 a 32.0 o/oo, respec respectivamente.
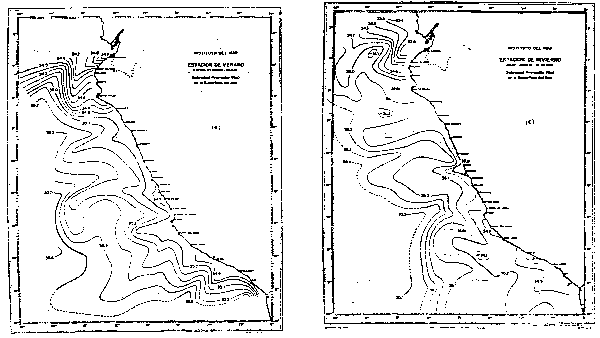
Fig. 5.- Salinidad promedio a Om en verano e invierno (de Zuta y Guillén, 1970).
El rango de salinidades (Stevenson et al., 1970) a través del límite septentrional de la corriente del Perú varía estacionalmente y de año a año, hallándose los valores más altos de salinidad de julio a septiembre cuando la Corriente Peruana es más fuerte y los menores valores de enero a marzo, con valores de 33.5 a 35.0 o/oo, y 33.0 a 34.7 o/oo respectivamente.
Frente a las costas de Ecuador, Colombia y Panamá (Fig. 6) se encuentran las salinidades más bajas en los meses de octubre a diciembre, con los valores más bajos en noviembre, inferiores a 24 o/oo frente a Panamá Bight y salinidades menores de 33 o/oo a lo largo de la costa de Colombia y Ecuador hasta 1°S. Las salinidades máximas se hallan en marzo y abril (mayor de 33 o/oo) en el Panamá Bight y (mayor de 34 o/oo) a lo largo de las costas de Colombia y Ecuador (Bennett, 1966). El mes de marzo quizás no es muy representativo para mostrar la distribución de la salinidad frente a la costa del Ecuador y sur de Colombia, ya que en este tiempo las lluvias son muy fuertes al norte del Ecuador y sur de Colombia debido al desplazamiento de la ZCIT hacia el sur. La celda de baja salinidad en la región de la Isla de Cocos se debe a un movimiento mar afuera de las aguas de baja salinidad provenientes del Panamá Bight.
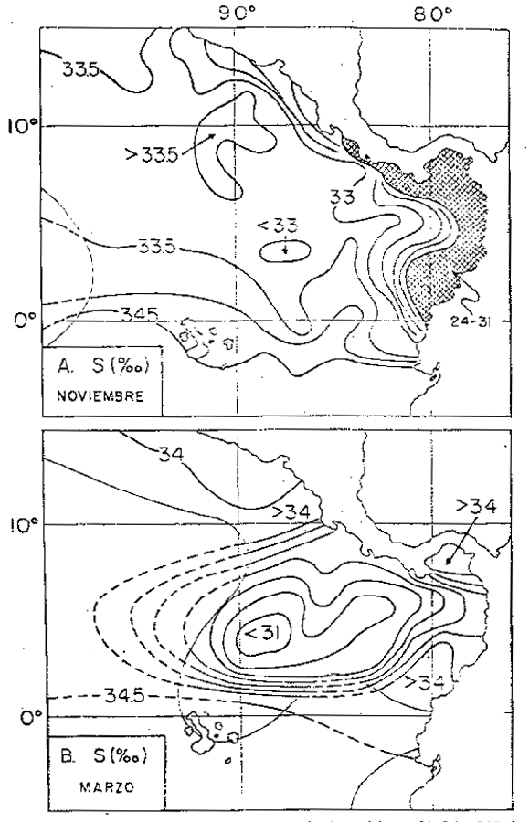
Fig. 6.- Salinidad promedio a Om en marzo y noviembre (de Enfield, 1976).
La zona de convergencia intertropical se halla situada al norte del Panamá Bight de mayo a diciembre, cuando los vientos alisios SE son más fuertes, ocasionando intensas lluvias y descargas en el Bight. En esta época no hay afloramiento en el Golfo de Panamá, la corriente de Colombia es fuerte y el remal occidental del remolino ciclónico que fluye hacia el sur del Bight es débil. En diciembre la zona de convergencia intertropical comienza a desplazarse hacia el sur.
| Masas de Agua Aguas Superficiales: Wyrtki (1967) en la superficie del mar (Fig. 7) distingue las siguientes masas de aguas: Aguas Tropicales Superficiales, que se caracterizan por su alta temperatura y baja salinidad y se presentan al norte de los 4°S, con salinidades menores de 34 o/oo y temperaturas mayores de 25°C. Las más bajas salinidades se hallan en el Golfo de Panamá y frente a las costas de Colombia, donde las salinidades varían de 34 o/oo a menos de 30 o/oo al final de la estación de lluvias (Bennett, 1965). Aguas Subtropicales Superficiales, de alta salinidad y de un rango variable en temperatura (28° a 15°C) y con salinidades mayores de 35 o/oo, debido al exceso de la evaporación sobre la precipitación, salinidades mayores de 36 o/oo se hallan entre las latitudes 12°S y 25°S y longitudes 100°W y 150°W. |
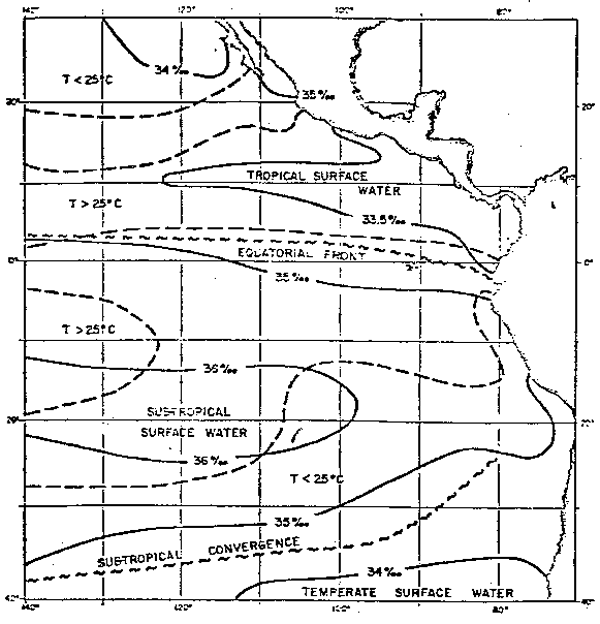 |
Fig. 7.- Principales masas de aguas superficiales en el Océano Pacífico Oriental: -Salinidad superficial, --- isoterma de 25°C: principales frentes oceánicos (de Wyrtki, 1966).
El límite norte de las Aguas Subtropicales Superficiales no coincide con el límite sur de las Aguas Tropicales Superficiales, excepto cerca a la costa del Ecuador. Entre estas dos masas de agua se encuentran las Aguas Ecuatoriales Superficiales, como producto de mezcla de dichas masas de aguas y se presenta generalmente al norte de los 6°S, con salinidades<34.8 o/oo (Zuta y Guillén, 1970).
El flujo hacia el norte de las aguas frías de la Corriente Peruana se opone a la tendencia natural de las Aguas Tropicales Superficiales que fluyen hacia el sur, dando como resultado la convergencia de las dos masas de agua y la formación del Frente Ecuatorial. Dicho frente ocupa una banda cuasizonal de cerca de 3° de latitud de ancho localizado entre los 0° y 5°S cerca del continente y se extiende en sentido oeste-noreste cerca de las Islas Galápagos, hallándose entre los 0° y 3°N. El frente se distingue con temperaturas desde 19° hasta 25°C y salinidades desde 35 o/oo a 33.5 o/oo de sur a norte, respectivamente.
Las aguas temperadas subantártidas del Pacífico Sur que fluyen hacia el este bajo la Corriente del Pacífico Sur, al aproximarse a la costa de Chile cerca de 50°S se dividen en dos brazos, uno hacia el sur y otro hacia el norte; este último fluye a lo largo de la costa de Sudamérica como parte del Sistema de Corrientes del Perú y Chile. Entre las latitudes 40°–50°S, la salinidad del agua superficial es reducida por las intensas lluvias y a la descarga de los ríos alcanzando valores menores de 33 o/oo. A medida que avanza hacia el norte su temperatura y salinidad incrementan lentamente alcanzando una salinidad superficial de 35 o/oo cerca de los 23°S.
Aguas Subsuperficiales:
Wyrtki (1967), ha realizado un estudio detallado sobre las masas de agua en la sub-superficie y señala que las Aguas Subtropicales Subsuperficiales y las de la capa mínima de oxígeno disuelto son las únicas formadas en la región. Las otras masas de agua son formadas fuera de la región y penetran dentro de ellas por medio del flujo horizontal y en gran escala debido a las mezclas horizontales.
Las Aguas Subtropicales, formadas en el medio del Pacífico Ecuatorial, se encuentran a lo largo del Ecuador como una lengua de máxima salinidad centrada aproximadamente entre los 100 m - 200 m de profundidad, las que son atrapadas por la Sub-corriente Ecuatorial (Corriente de Cromwell) y llevadas hacia el este. Cerca de las Islas Galápagos presenta un máximo cerrado de salinidad, el cual hace contraste con el agua subsuperficial menos salina del norte y sur del Ecuador (Stevenson y Taft, 1971).
La capa mínima de oxígeno disuelto frente al Perú se encuentra entre 15°C y 50°C, y tiene casi las mismas características de temperatura y salinidad señaladas por Sverdrup (1946) para la masa del Pacífico Ecuatorial. Esta misma agua fue llamada por Wyrtki (1963) Agua Ecuatorial Subsuperficial (Fig. 8). Frente al Perú, Zuta y Guillén (1970) hacen una diferencia denominando Aguas Ecuatoriales Subsuperficiales a las aguas asociadas con valores relativamente altos de salinidad (34.9–35.1 0/00) y de oxígeno, vinculados a la extensión de la Corriente de Cromwell y Aguas Ecuatoriales Profundas a las aguas con salinidades de 34.9 – 34.0 0/00 y que coinciden con el mínimo de oxígeno y fluyen hacia el sur.
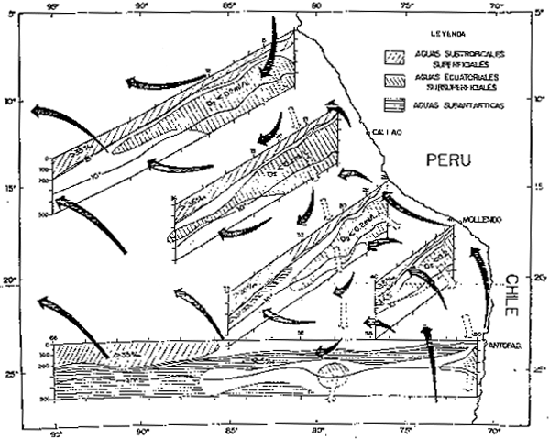
Fig. 8-Masas de agua frente al Perú y Chile (de Wyrtki, 1967).
La capa superior de las Aguas Ecuatoriales Subsuperficiales presenta dos máximos de salinidad con sus correspondientes mínimos de oxígeno: uno costero asociado a la Contra- corriente Subsuperficial Peruano-Chilena, la cual avanza hasta más allá de 41°S (Robles, 1968; Alarcón y Pineda, 1969; y Wooster y Gilmartin, 1961), y otro oceánico más atenuado relacionado con la Contracorriente Oceánica Peruana descrita por Wyrtki (1963) y llega más allá de los 33°S.
Las aguas con salinidades menores de 34 0/00 y temperaturas menores de 15°C que fluyen hacia el norte de las costas de Chile cerca de la Convergencia Subtropical (Latitud 35°S) se hunden por debajo de las Aguas Subtropicales Superficiales que tienen salinidad y temperatura ligeramente mayor. Dicho flujo puede ser seguido por la salinidad mínima a 100 m de profundidad que se dirige hacia el norte de Chile y sur del Perú hasta cerca de los 15°S.
El Agua Antártica Intermedia se forma cerca del Frente Antártico Polar y se extiende hacia el norte entre las profundidades de 500 y 1500 m de profundidad (Wyrtki, 1967; y Robles, 1966) y se caracteriza por su mínima salinidad. Al sur de los 15°S la salinidad es menor de 34.5 o/oo y el contenido de oxígeno es alto. Al sur de 20°S la salinidad mínima coincide más o menos con la máxima de oxígeno (1.0 - 2.0 ml/L).
Circulación
Circulación superficial:
La circulación en la capa superficial en el Océano Pacífico (Fig. 9) está sometida al sistema de los vientos y a los movimientos giratorios anticiclónicos del Océano Pacífico del norte y del sur. Entre estos dos movimientos se encuentra la Contracorriente Ecuatorial.
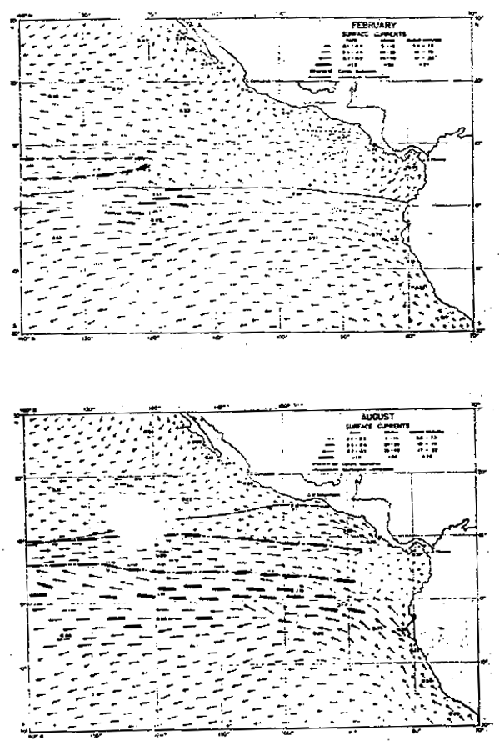
Fig.9-Corrientes uperficiales en febrero y agosto (de Wyrtki, 1965).
Como parte de la circulación del Pacífico Sur las Aguas Subantárticas fluyen hacia el este alcanzando las costas de Chile entre las latitudes de 44° y 48°S y en el verano algo más hacia el sur; luego se dividen en dos ramales, uno que se dirige a lo largo de la costa hacia el sur, la Corriente de Cabo de Hornos; y la otra que se dirige hacia el norte, la Corriente de Humboldt o Chileno-Peruana, la cual tiene dos ramales, uno oceánico y otro costero. Frente a las costas de Chile, la rama oceánica es denominada Corriente de Chile (Wooster, 1970) o Corriente Oceánica Chileno-Peruana (Robles et al ., 1976), la cual fluye hacia el norte con velocidades de 12 cm/seg. Esta corriente transporta principalmente Aguas Subantárticas de relativamente baja temperatura y salinidad. La mayor parte de este flujo gira hacia el oeste antes de alcanzar los 20°S. La rama costera de la corriente Chileno-Peruana es denominada Corriente Costera Chilena cuyo flujo es subantartico o netamente costero con velocidades de 25 cm/seg aproximadamente, y es observada a partir de los 28°S llegando algunas veces más allá de la Península de Mejillones en el invierno. Sievers y Silva (1976) dicen que la Contracorriente del Perú tiene presumiblemente dos ramas: la primera, oceanica, situada al oeste de los 77°W la cual avanza hasta los 27°S, coincidiendo con la ubicación dada por Wyrtki (1963). La segunda, costera, que avanza hacia el sur por los 75°W, extendiéndose hasta Valparaíso. Robles et al. (1976) señalan además una contracorriente que aparece inmediatamente hacia la costa, denominada Contracorriente Costera de Chile, que fluye hacia el sur alcanzando en el verano la latitud de 37°S, la cual está sujeta a variaciones estacionales alcanzando velocidades mayores de 15 cm/seg en el invierno.
Frente a la costa del Perú, la Corriente de Humboldt o Chileno-Peruana toma el nombre de Corriente Peruana conformada por la Corriente Costera Peruana y Corriente Oceánica Peruana, las cuales fluyen hacia el noroeste abandonando la costa cerca de los 5°S para luego integrarse en la Corriente Surecuatorial como parte de la circulación anticiclónica del Océano Pacífico Sur. Estas dos corrientes están generalmente separadas por un flujo debil e irregular hacia el sur, la Contracorriente del Perú, que es subsuperficial y ocasionalmente llega a la superficie del mar (Wyrtki, 1963). La Corriente Costera Peruana que es fría y rica en nutrientes tiene una velocidad promedio de 5 a 15 cm/seg, alcanzando alqunas veces velocidades de 40 a 80 cm/seg cerca de los 7°S. Su flujo varía estacionalmente y es más intenso durante los meses de abril a septiembre, con un transports confinado a los primeros 200 m de profundidad. La Corriente Oceanica Peruana es más intensa qur la Corrietite Costera Peruana y llega hasta los 700 m de profundidad. Presenta variacones estacionales y durante los meses de julio a octubre forma un sólo flujo con la Corriente Costera Peruana hacia el oeste, integra-ndose luego en la Corriente Surecuatorial. la contracorriente en la superficie del mar es observada con mayor intensidad durante los meses de noviembre a febrero, estando ausente los meses de julio a octubre.
La Corriente Surecuatorial al oeste de las Islas Galápagos fluye hacia el oeste alcanzando las velocidades más altas en ambos lados del Ecuador, entre los 3°N y 8°S, aproximadamente. Su límite septentrional es la Contracorriente Ecuatorial cerca de los 4°N. Su velocidad promedio cambia poco durante el año al sur del Ecuador, mientras que al norte del Ecuador las velocidades son generalmente mayores y presentan una marcada variación estacional con velocidades mayores de 77 cm/seg de julio a noviembre.
La Contracorriente Ecuatorial (Wyrtki, 1965) al este de los 140°W, fluye hacia el este entre los 4° y los 11°N, cuya posición y anchura varía con la estación del año. de mayo a diciembre, la contracorriente se desarrolla claramente con una anchura que varía de los 30–5° de latitud, logrando en septiembre las mayores velocidades. Al este de los 90°W la contracorriente se bifurca: una parte fluye hacia el norte y el noroeste alrededor del Domo de Costa Rica, y otra parte se dirige hacia el sur y al oeste ingresando en el sistema de la corriente sur ecuatorial. En enero la contracorriente comienza la dividirse y existe solamente en secciones separadas. En febrero y marzo, la contracofriente sólo llega hasta los 120°W para luego en abril desaparecer totalmente.
Frente al Panamá Bight la circulación superficial es en el sentido contrario a las agujas del reloj y presenta dos ramales; uno oriental, que fluye hacia el norte a lo largo de la costa denominada Corriente de Colombia (Wooster, 1959) y que a menudo penetra dentro del Golfo de Panamá, y otro Occidental, formado por el derrame de agua procedente de la entrada del golfo y que a menudo se une a la corriente sudeste que fluye a lo largo de la península panameña. En el invierno (Stevenson, 1970; y Stevenson et al., 1970) del norte (febrero) el derrame superficial del Golfo de Panamá es más fuerte, alcanzando velocidades de 50 cm/seg. En esta época la Corriente de Colombia es estrecha y logra velocidades hasta de 150 cm/seg, mientras que en el Ecuador, época de lluvias, la circulación consiste de una serie de corrientes y de celdas que semejan remolinos, debido a la transición de la circulación del Panamá Bight a la del límite septentrional de la Corriente Peruana. En agosto la Corriente de Colombia, al norte de los 5°N, es más rápida que en las otras estaciones del año.
Circulación subsuperficial:
En la subsuperficie se tiene la Corriente Subsuperficial Peruano-Chilena, la Contra-corriente Surecuatorial, la Subcorriente Ecuatorial y la Corriente de Chile.
La Subcorriente Ecuatorial (Knauss, 1960) fluye hacia el este desde por lo menos 140°W hasta 92°W, con velocidades maximas de 100 a 150 cm/seg con su núcleo cerca de la base de la termoclina, la cual asciende desde los 100 m de profundidad en el oeste hasta cerca de 40 m en el este. Su flujo hacia el este ha sido encontrado al este de las Islas Galápagos por Knauss (1966) y Stevenson y Taft (1971), y se considera que es una extensión de la Subcorriente Ecuatorial con velocidades observadas de 10 a 37 cm/seg, hallándose la corriente al norte y sur del Ecuador. White (1969) halló que la subcorriente se bifurca tanto hacia el norte como hacia el sur de las Galápagos, encontrándose el ramal sur entre los 2°S y 4°S a los 84°W.
La Corriente Subsuperficial Peruano-Chilena (Wooster y Gilmartin, 1961) nace de la unión de la extensión de la Corriente de Cromwell con la Contracorriente Surecuatorial (Cochrane y Zuta, 1968 ; y White, 1969) un pocoal sur de los 6°S y lejos de la costa. Fluye hacia el sur próximo a las costas del Perú y Chile entre los 50 y 300 m de profundidad con velocidades de 4 a 10 cm/seg alcanzando 20 cm/seg frente a Punta Aguja. Su transporte y velocidad decrecen hacia el sur extendiéndose hasta cerca de los 40°S entre las profundidades de 100 y 300 m.
La Contracorriente del Perú fluye hacia el sur a lo largo de 80°W, principalmente, como una corriente subsuperficial llevando salinidad y bajo contenido de oxígeno entre los 100 y 500 m, siendo más fuerte cerca de los 100 m. Su transporte decrece hacia el sur de 10×1012 cm3/seg en 5°S, a 2 × 1012 cm3/seg en los 22°S.
La Contracorriente Surecuatorial fluye de oeste a este entre los 8° y 10°S, aproximadamente entre los 100 y 500 m de profundidad con velocidades de 8 a 15 cm/seg, transportando 10 millones de m3/seg. Esta contracorriente es más intensa entre julio y octubre y ha sido estudiada por Reid (1959 y 1961), Wooster (1961) y Wyrtki (1963). .bl. La Corriente Chilena fluye hacia el norte y noroeste entre los 50 y 150 m de profundidad en la parte costera del Perú.
Afloramiento
A lo largo de la costa el afloramiento es muy superficial y está restringido principalmente a profundidades menores de 100 m. Frente al Perú las aguas de afloramiento se caracterizan por su baja temperatura, bajo contenido de oxígeno disuelto y alto contenido de nutrientes. La velocidad promedio del afloramiento frente a las costas del Perú es de 20 × 10-5 cm/seg con un rango de 5 a 30×10-5 cm/seg.
El afloramiento típico y casi permanente se presenta entre las latitudes 140–16°S, luego le siguen las áreas 4°–6 ° S, 7°–6°S y 11°–13°S. Su intensidad varía de acuerdo a su posición geográfica y la estación del año, siendo más intenso en el invierno y mínima en el verano. Las aguas de afloramiento (Zuta y Guillén, 1970) a lo largo de la costa tienen tres orígenes: a) las aguas sde la extensión de la Corriente de Cromwell, las cuales afloran principalemente entre los 4°–6°S y que contribuyen en parte al afloramiento al norte de los 9°S; b) Las aguas de la Contracorriente Subsuperficial Peruano-Chilena que afloran principalmente al norte de los 12°S y que eventualmente en el verano y otono pueden contribuir al afloramiento más hacia el sur; y c) Las aguas temperadas subantárticas, las cuales afloran principalmente al sur de los 14°S. El promedio del área del afloramiento es de 25 000 millas cuadradas.
Frente a las costas de Chile las áreas principales de afloramiento se hallan frente a 18°30s' (Arica), 19°30'S (Antofagasta), luego le siguen las áreas situadas entre los 23°S. Al norte de Chile las temperaturas son bajas y las salinidades ususlmente menores; de 35 o/oo debido al afloramiento de las aguas temperadas subantarticas. Robles et al. (1976) hallaron un promedio de la profundidad de afloramiento del agua subantártica de 64 m y el promedio de la profundidad de afloramiento de las Aguas Ecuatoriales Subsuperficiales de 163 a 85 m.
En el Pacífico Ecuatorial Oriental las aguas superficiales más frías y termoclina menos profunda son atribvuidas a la divergencia supervcicial y al afloramiento bajo la influencia de los alisios SE. Lejos de la costa al sur de las Islas Galápagos se observa un area de baja temperatura, kla cual refleja el afloramiento ecuatorial.
El afloramiento (Stevenson, 1970) se presenta más frecuentemente en el centreo del Panamá Bight y a lo largo de la mitad del este de la estrada del golfo, ambos con veclocidades de 45×10-4 cm/seg, siendo más intenso y fuerte durante febrero y marzo, aunque durante cada estación ocurren algunos afloramientos.
El Fenómeno El Niño
La designación de El Niño se aplica a las grandes anomalías oceánicas que occurren de tiempo en tiempo frente a la costa de Sudamérica. El Fenómeno El Niño es ocasional, irregular, aperiódico y de grandes repercusiones socio-económicas para el Perú. Su origen no es bien conocido y parece estar ligado al debilitamiento general de la circulación de los vientos alisios del hemisferio sur.
El Fenómeno El Niño se presenta con diferentes intensidades con intervalos de 5 a 16 años y su duración es variable, presentando su mayor intensidad en los meses de lebrero y marzo. Los eventos principales del fenómeno fueron los occurridos en 1891, 1925–26, 1940–41, 1957–58, 1965, 1972–73, 1976 y 1982–83, los cuales alteraron temporalpesguería, la agriculatura y el clima. Dentro de los últimos eventos secundarios del Fenomento El Niño se hallan los occurridos en 1953 y 1969.
Nutrientes y Oxígeno Disuelto
El oxígeno disuelto frente a las costas de Chile (Fig. 10) en la superfi ie del mar se halla con valores de 4.0 a 8.0 ml/L, predominando el rango de 5.0 a 6.0 ml/L (Forsbergh y Breeknow, 1965; Robles, 1966; e Inostroza, 1972).
Frente al Pereú (Zuta y Guillén, 1970). la distribución superficial del oxígenio disuelto presenta valores promedio de 2.0–7.0 mk/L, correspondiendo los mínimos a las areas de afloramiento. En el varano e invierno se hallan valores prodemio de 7.0 a 2.0 ml/L, respectivamente.
El oxígeno disuelto en la superficie del mar frente a las costas del Ecuador, colombia y Golfo de Panamá presentan valores promedio de 4.0 a 5.0 ml/L (Stevenson et al. 1970) Sin emvargo, puede observarse algunas veces valores menores de 4.0 ml/L y mayores de 5.0 ml/L. debido a los procesos de afloramiento y fotosíntesis, respectivamente. Parra (1977) hallo valores en la superficie del mar de 2.5 a 6.7 mol/L de oxígeno disuelto.
Las conventraciones de nutrientes en la capa supercficial decrecen debido a su utilización por el fitoplancgon y son abstecidas por la sescomposición de la materia orgánica in sityu y por los procesos de afloramiento, l9s cuales abastecen de nuytrientes a las aguas superciciales. Entre los nutrientes tenemos: los fostatos, silicat6os y nitratros,
Frente a las costas de Chile (Fig.10) los nutrientes presentan valores bajos en la supercieis del mar en comparacion con comparacion con las costas del Perú. Los fosfatos a lo largo de la costa presentatn un trango promedio de 2.0 a 0.25 ug-at/L (Reid, 1962; Forsbergh y Broenkow,, 1965; e Inostroza, 1972), correspondiendo los mayores vcalores a las áreas cercxa de la ostta. En la superficie a la latitud 23°S se observatron converntraciones menores de 1.0 ug-at/L, 5.0 ug-at/L y 10 ug-at/L de fosfatos, nitratos y silicatos, respectivamente; y a la latitud 33°S se hallaron valores menores de 0.4 ug-at/L de fosfatos y 1.09 ugat/L y 5.0 ug-at/L de silicatos y nitratos, respectivamente (Forsbergh y Broenkow, (1965).
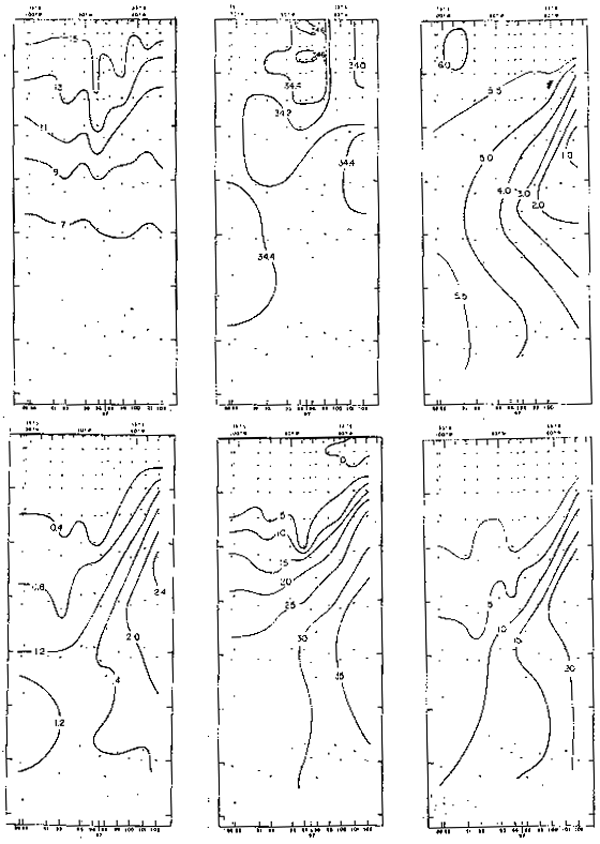
Fig. 10.- Secciones: a) temperatura, b) salinidad, c) oxígeno disuelto, d) fosfatos, e) nitratos y f) silicatos frente a Valparaíso, Chile, en noviembre de 1963 (de Forsbergh y Broenkow, 1965).
Fig. 11. - Nitratos y silicatos promedios a Om en verano e invierno.
Frente a las costas del Perú (Guillén, 1976) la elevada concentración de nutrientes (Figs. 11 y 12) se debe en parte al afloramiento costero y en parte al transporte del agua de mezcla. En la superficie del mar se encontraron valores promedios de: 0.2 a 3.20 ug-at/L de fosfatos, 00 a 24.0 ug-at/L de silicatos y 0.5 a 21.0 ug-at/L de nitratos, hallándose los valores más altos cerca de-la costa (>2.5 ug-at/L de fosfatos,> 15.0 ugat/L de silicatos y>15.0 ug-at/L de nitratos).
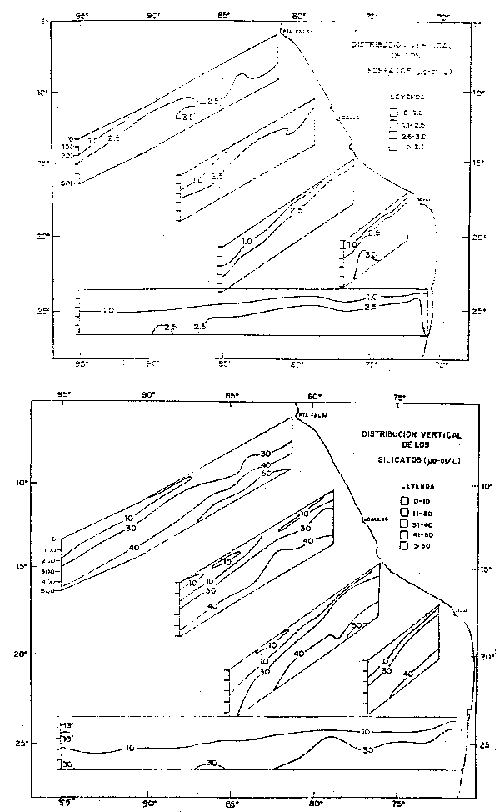
Fig. 12.- Secciones de fosfatos y silicatos en octubre-noviembre de 1960.
Las menores concentraciones (<1.0 ug-at/L de fosfatos, <5.0 ug-at/L de silicatos y <5.0 ug-at/L de nitratos) se observaron en las Aguas Subtropicales Superficiales y Aguas Ecuatoriales Superficiales.
Frente al Panamá Bight no se dispone de mucha información sobre nutrientes, motivo por el cual sólo nos referimos a los resultados de los cruceros realizados durante la Operación ACENTO. En la superficie del mar la concentración de los nutrientes fueron muy bajas. Los fosfatos en la superficie del mar se encontraron con valores menores de 0.2 ug-at/L en la mayor parte del área durante todos los cruceros con un rango de 0.44 a 0.02 ug-at/L a excepción del ACENTO 4, donde se hallaron valores mayores de 0.02 ug-at/L, con una concentración máxima de 0.44 ug-at/L. Thomas (1966) señala que los nitratos son los más importantes de los micronutrientes en el Pacífico Oriental Tropical. Los nitratos en la superficie del mar se hallaron con concentraciones menores de 1.0 ug-at/L. con un rango de 0.0 a 3.2 ug-at/L. Los silicatos presentaron valores menores de 2 ug-at/L en la mayor parte del area. Parra (1977) y Forsbergh y Broenkow (1965) hallaron en la superficie del mar (Fig. 13) un rango de 0.2 a 4.5, 1.0 a 5.0 y 3.0 a 30.3 ug-at/L de fosfatos, nitratos y silicatos, respectivamente, para los meses de abril-mayo y septiembre-octubre de 1976. Forsbergh (1969) y Parra (1977) hacen un estudio más detallado para el área. Frente al Ecuador la información sobre nutrientes es limitada. En la superficie del mar las concentraciones de fosfatos son relativamente bajas y se presentan con valores de 0.05 a 1.55 ug-at/L al oeste de las Islas Galápagos, correspondiendo las mayores concentraciones al este de los 88°W. Los silicatos en la superficie del mar se hallaron con concentraciones de 0.10 a 20.0 ug-at/L.
FLUCTUACIONES DE LAS CONDICIONES OCEANOGRAFICAS
Condiciones Oceanográficas Durante Años Fríos
En los últimos 20 años, según la distribución de las anomalías térmicas se tiene los siguientes años fríos: 1964, 1967–68, 1970–71 y 1974–75, con anomalías negativas mayores de 2°C. Como ejemplo se ha seleccionado el año 1967, cuyas principales caracteristicas son las siguientes.
La distribución de la temperatura y salinidad en la superficie del mar durante el verano e invierno son mostradas en las Figuras 3 y 4.
La Corriente Subsuperficial Ecuatorial (White, 1969) presenta dos ramales, uno al norte y otro al sur del Ecuador. Este último ingresa entre los 2° y 4°S y cerca de los 5°S el flujo dobla hacia el oeste hasta los 84°W donde se divide. Parte del flujo continúa hacia el oeste mientras que la mayor parte del flujo dobla hacia el sur. extendiéndose hasta cerca de los 10°S donde retorna a la costa. Allí el flujo dobla hacia el sur para formar la Corriente Subsuperficial Perú-Chile.
La Contracorriente Sur Ecuatorial (White, 1969) penetra a la región entre los 6° y 13° S, siendo su principal flujo entre los 6° y 8°S, asociado con una lengua de alto contenido de oxígeno mayor de 1.0 ml/L. Parte del flujo continúa hacia 84OW y cerca de los 5°S se junta con el flujo hacia el sur para formar la Corriente Subsuperficial Perú-Chile. Cochrane y Zuta (1969) mostraron un flujo hacia el este que tenía las misnias estructuras de masa de agua y dos cores de alta concentración de oxígeno cerca de 2°N y 2°S. Ambas ramas pasan en cada lado de las Islas Galápagos y se extienden hacia la costa de Sudamérica Ellos sugirieron que la Contracorriente Sur Ecuatorial abastece de agua a la Corriente Subsuperfical Perú-Chile.
La distribución del oxígeno disuelto en la superficie de 180 cl/t (Fig. 14) está de acuerdo con la aceleración potencial a 160 cl/t, observándose el ramal sur de la Corriente Subsueerficial Ecuatorial. Wooster y Cromwell (1958) hallaron dos cores de alto contenido de oxígeno al este de las Islas Galapagos, una a 1°S y la otra a 2°N. Por otra parte, Bennett (1963) halló alto contenido de oxígeno en ambos lados de las Islas Galápagos y sólo al este de las islas a 84°W encontro uil core de alto contenido de oxígeno sobre el Ecuador. Las dos lenguas de alto contenido de oxtgeno observadas una a 79° y otra a 87°W, está relacionada la primera con el flujo de la Corriente de Chile y la otra con el flujo hacia el oeste de la extension de la Corriente de Chile.
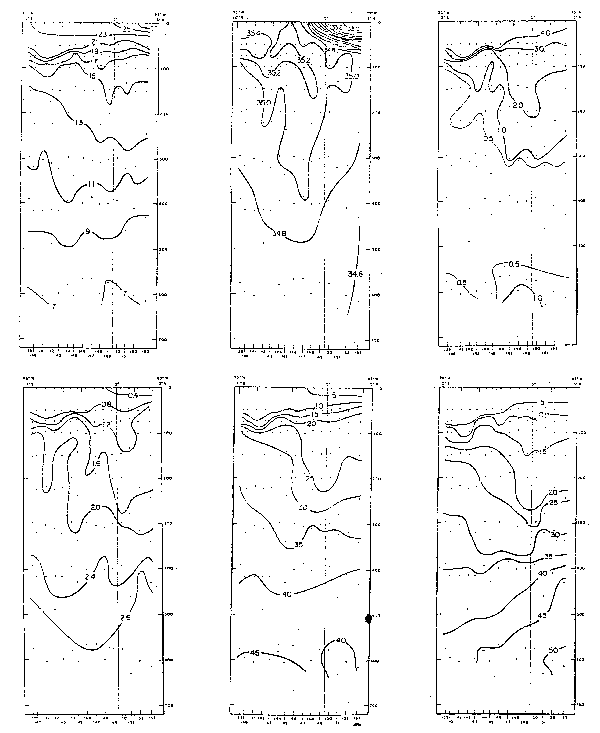
Fig. 13.- Secciones: a) temperatura, b) salinidad, c) oxígeno disuelto, d) fosfato, e) nitratos y f) silicatos frente a Colombia, Ecuador y Perú en diciembre de 1963 (de Forsbergh y Broenkow, 1965).
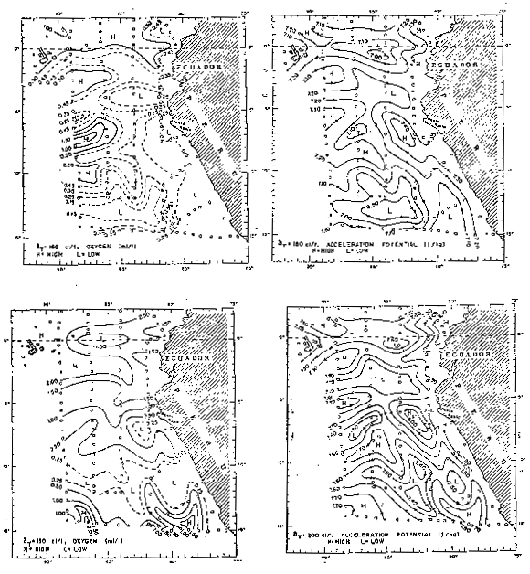
Fig. 14 - Oxígeno disuleto a la superficie donde δT = 180 cl/t y aceleración potencial a la superifcie donde δT = 200 cl/t en febrero — marzo de 1967 (de white, 1969).
La distribución de la acelaración potencial a 200 cl/t (Fig. 14) difiere un poco de la distribución a 160 cl/t (White, 1969) y muestra la rama sur de la Corriente Subsuperficial Ecuatorial, la que se extiende hacia la costa del Perú hasta cerca de 5°S, donde la mayor parte del flujo fluye alrededor del meandro de Punta Aguja, mientras que el resto del flujo fluye fuera de la región a lo largo de 5°S. Entre los 6° y 8°S ingresa a la región la parte norte de la Contracorriente Surecuatorial.
Una gran parte de este flujo continúa hacia el este para juntarse al flujo hacia el sur del meandro de punta Aguja. Dentro del meandro mismo se desarrolla un eddy anticclónico, con un definido flujo hacia el norte. Entre los 10° y 11°S ingresa a la región la parte sur de la contracorriente Surecuatorial y se extiende al este hasta 84°W.
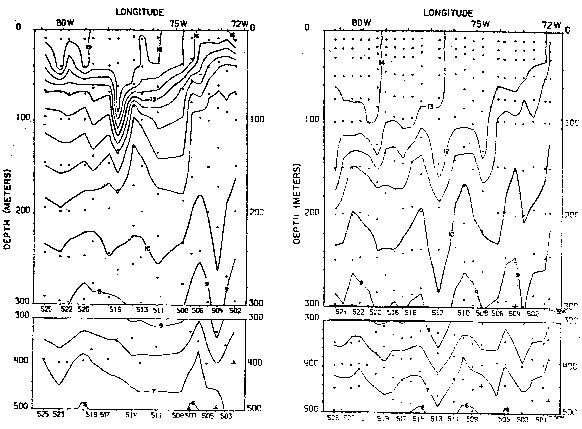
Fig. 15.- Secciones de temperatura frente a Valparaíso, Chile, en marzo y septiembre de 1967 (de Love, 1970–77).
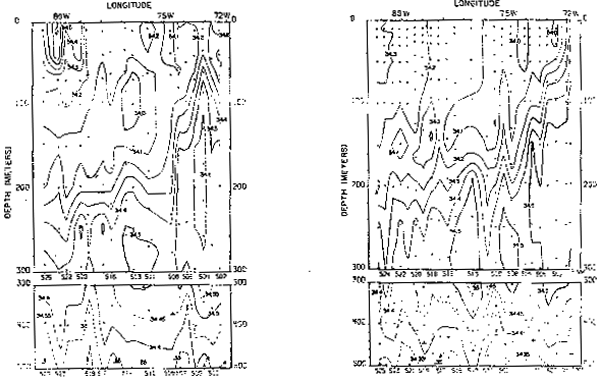
Fig. 16.- Secciones de salinidad frente a Valparaíso, Chile, en marzo y septiembre de 1967 (de Love, 1970–77).
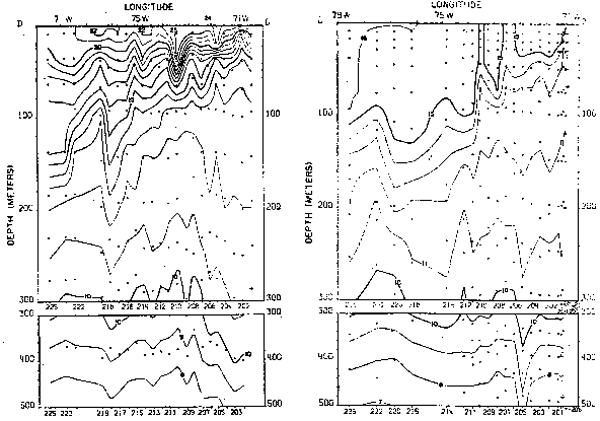
Fig. 17.- Secciones de temperatura frente a Iquique, Chile, en marzo y septiembre de 1967 (de Love, 1970–77).
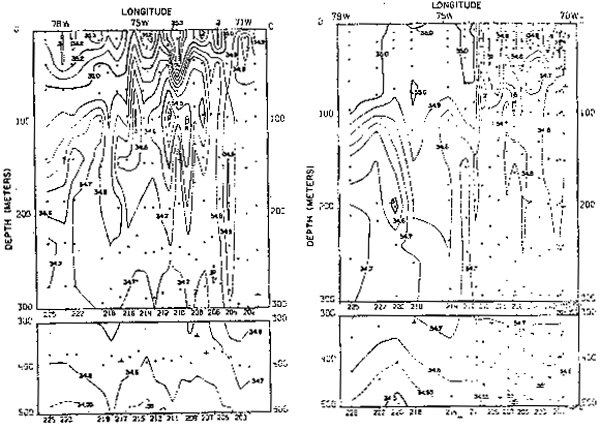
Fig. 18.- Secciones de salinidad frente a Iquique, Chile, en marzo y septiembre de 1967 (de Love, 1970–77).
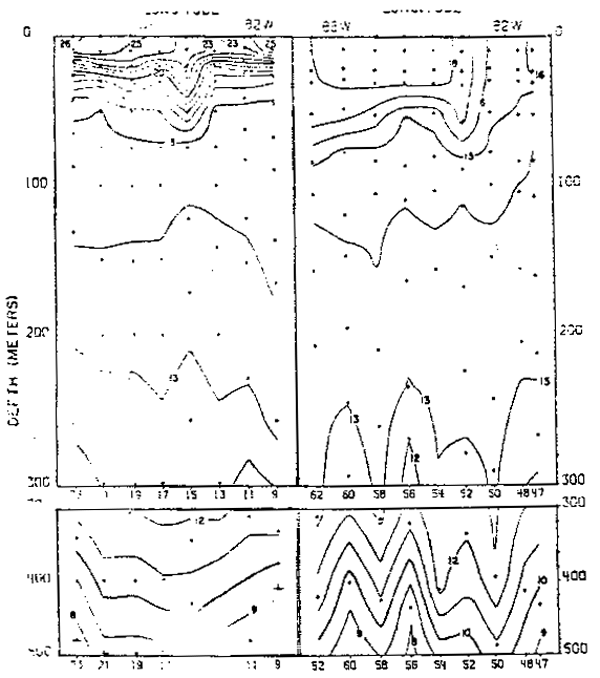
Fig. 19.- Secciones de temperatura frente a Cabo Blanco, Perú, en marzo y septiembre de 1967 (de Love, 1970–77).
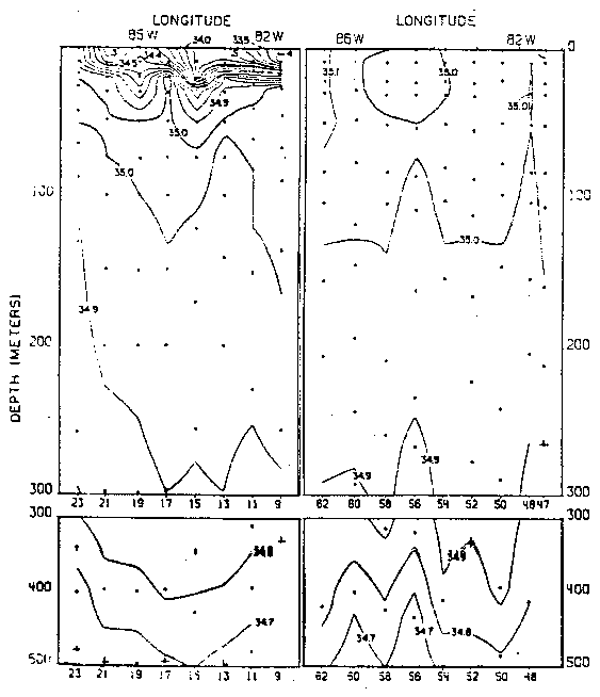
Fig. 20.- Secciones de salinidad frente a Cabo Blanco, Perú, en marzo y septiembre de 1967 (de Love, 1970–77).
Para la distribución vertical de la salinidad y temperature se han seleccionado las secciones frente a las latitudes 4°, 20° y 33°S (Figs. 15 a 20) en donde se destaca debajo de la capa superficial el flujo subsuperficial hacia el sur que tiene gran influencia en la circulación frente al Perú y Chile en la capa de 50 a 300 m de profundidad. La Corriente Subsuperficial Peruano-Chilena que se caracteriza por su salinidad relativamente alta, se hace más evidente al sur de los 15°S; así, frente a la latitud 20°S se presenta con salinidades mayores de 34.8 o/oo cerca de la costa y frente a la latitud 33°S con salinidades mayores de 34.5 o/oo en el verano (Fig. 16).
Otro flujo importante, principalmente frente a la costa de Chile, son las aguas subantárticas que son transportadas hacia el norte y que llegan en algunas oportunidades hasta los 12°S (Zuta y Guillén, 1970), con salinidades menores de 34.1 o/oo frente a la latitud 33°S y salinidades menores de 34.6 o/oo frente a la latitud 20°S, durante el verano.
Las variaciones del verano al invierno son mostradas en las figuras 15 a 20 en la que no sólo se observan cambios en la posición y características de las diferentes masas de aguas, sino también la intensidad dellos procesos de afloramiento-del que depende en gran parte el desarrollo de las pesquerías. Las aguas que afloran juegan un rol importante en la productividad de las aguas debido a que cada masa de agua tiene sus propias características físicas, quimicas y biológicas (Calienes y Guillén, 1981); así por ejemplo, frente a la latitud 4°S en el invierno, las aguas que afloran corresponden a las aguas de la extension de la Corriente de Cromwell, y las aguas que afloran frenteta la latitud 20° en el invierno, corresponden a las aguas subantárticas.
La rama sur de la Corriente Subsuperficial Ecuatorial (Figs. 21 a 23) es identificada no sólo por su temostato, sino también por su alto contenido de oxígeno y contenidos de nutrientes, los que presentan variaciones estacionales. La capa mínima de oxígeno disuelto de<0.25 también presenta variaciones no sólo en su extensión sino también en su espesor (Fig. 22). Los nutrientes también presentaron variaciones estacionales (Fig. 23), asi los nitratros se hallaron entre 20 y 25 ug-at/L en el verano, aumentando de 25 a 30 ug-at/L en el invierno.
Condiciones Oceanográficas Durante Años Calientes
El Fenómeno El Niño aparece irregulartnente frente a las costas de Sudamérica con grandes repercusiones en los niveles tróficos del ecosistema marino, afectando grandemente el rendimiento de las pesquerías. Los principales eventos ocurridos son: 1891, 1925–26, 1940–41, 1957–58, 1965, 1972–73, 1976 y 1982–83, los que tuvieron efectos limitativos en la producción biológics.
El Niño de 1965, se presentó frente a las costas del Perú (Guillén y Calienes, 1981) con mayor intensidad durante los meses de marzo y abril y fue causado por la invasión de Aguas Ecuatoriales Superficiales, caracterizada por temperatures de 24°–27°C, salinidades de 33.0 a 34.8 o/oo, asociado con bajas concentraciones de nutrientes, de clorofila "a" (0.18-1.0 ug/1) y de productividad (< 0.05 gC/m2/día).
Durante El Niño de 1972, se observó dos desplazamientos de agua hacia el sur transportando aguas del Golfo de Panamá por el este de los 85°W, siendo el de febrero-marzo el más alejado de la costa, mientras que el de diciembre fue más costero (Zuta et al., 1976).
Frente al Perú la primera fase tuvo lugar en febrero-marzo y fue caracterizado por la invasión de las aguas ecuatoriales hasta la latitud de 10°S, aproximadamente (Salinidad<34.5 o/oo) y bajas concentraciones de nutrientes (2.0–4.0 ug-at/L de silicates, 0.50–0.75 ug-at/L de fosfatos y 0.75-1.00 ug-at/L de nitratos). Su contenido de clorofila "a" fue menor que la encontrada en El Niño de 1965. La producción total fue la tercera parte del promedio. La segunda fase apareció en diciembre en donde las aguas procedentes de la región ecuatbrial se extendieron hasta los 10°S, aproximadamente, con temperatures de 23–25°C y salinidad de 34.0 ° 34.8 o/oo, asociado con bajas concentraciones de nutrientes y clorofila "a".
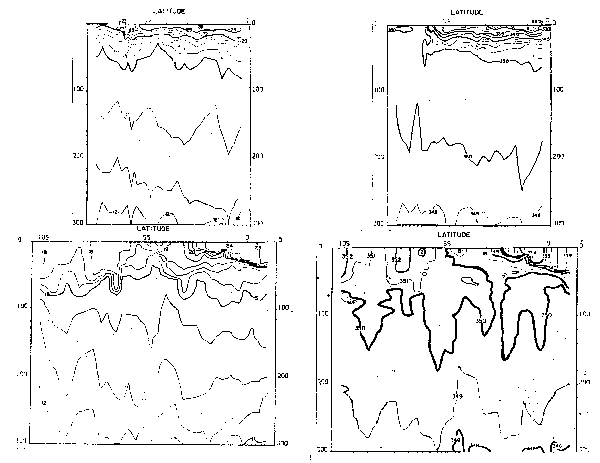
Fig. 21.- Secciones de temperatura a lo largo de 82°W en febrero y agosto 1967, 1970–77.
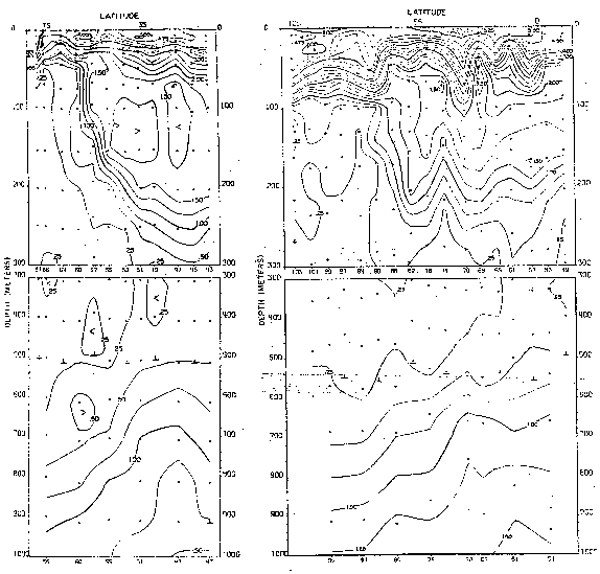
Fig. 22.- Secciones de oxigeno disuelto (m1/1) a lo largo de 82°W en febrero y agosto de 1967 (de Love, 1970–77).
Frente a Chile, durante el verano, la temperatura superficial del mar se halló ligeramente sobre el promedio (Caviedes, 1981). Sin embargo, al sur de los 28°S El Niño no tuvo influencia (Alarcón, 1975).
El invierno de 1972 se caracterizó por el predominio de las Aguas Subtropicales Superficiales (Salinidad>35.1 0/00) frente a la costa peruana al sur de los 5°S y al oeste de los 83°W frente al Ecuador. Las Aguas Tropicales Superficiales avanzaron en dos frentes, uno costero que se extendió hasta los 4°S y otro lejos de la costa frente a Esmeralda que se extendió en dirección SW hasta los 2°S.
Frente a Chile, las aguas se hallaron más calientes que lo normal, siendo más marcado en casi toda el área costera en donde predominaron las Aguas Subtropicales Superficiales con salinidades mayores de 35.0 0/00, cuyas salinidades son más altas que las encontradas en 1967 (Robles et al., 1976; y Sievers y Silva, 1976).
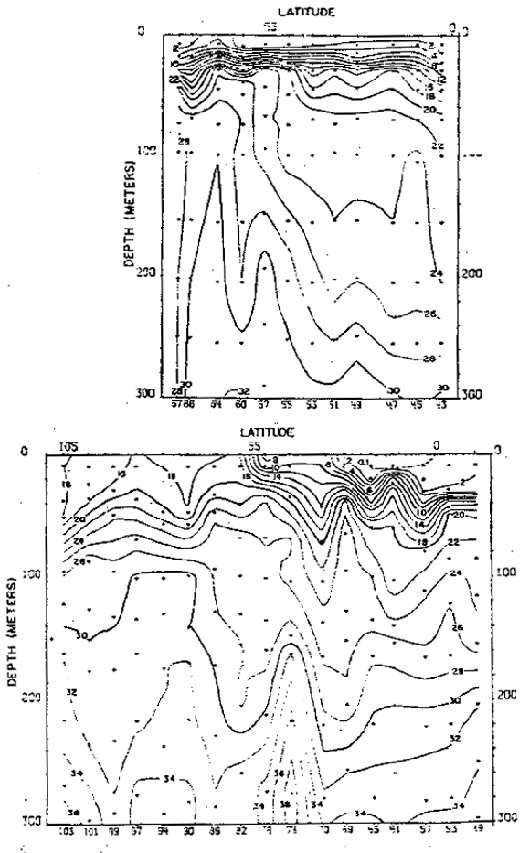
Fig. 23. - Secciones de nitratos (ug-at/1) a lo largo de 82° en febrero y agosto de 1967 (de Love, 1970–77).
El Niño de 1976 frente a las costas de Perú, en enero y befrero se observó un calentamiento de la temperatura superficial del mar al norte de los 15°S, debido a la presencia de las Aguas Ecuatoriales Superficiales (salinidad<34.8 o/oo) que se extendió hasta los 9°S. Sin embargo, el máximo calentamiento fue hallado en junio–agosto, asociado con Aguas Subtropicales Superficiales caracterizadas por su baja concentración en nutrientes y pro– ductividad. La distribución de la clorofila “a” fue muy diferente al promedio especial– mente en julio con valores 1.0 ug/L en casi toda la costa.
El Niño de 1982 frente al Perú (Zuta y Guillén, 1983), en el mes de septiembre se observó un calentamiento a lo largo de la costa con anomalíes mayores de 2°C que el pro– medio, originado por la aproximación a la costa de las Aguas Subtropicales Superficiales, caracterizadas por su baja concentración en nutrientes y productividad, cuyo calentamient continuó los proximos meses. En enero de 1983 (Fig. 24) se observó la presencia de aguas de la región ecuatorial (salinidad<34.8 o/oo) con temperaturas de 26°–29°C, que se extend ron hasta la latitud de 14°S aproximadamente, asociado con bajísimas concentraciones de nutrientes, (<6.0 ug-at/L de silicatos,<0.4 ug-at/L de nitratos, y <0.2 ug-at/L. de fosfatos). Las concentraciones de los fosfatos y nitratos fueron inferiores a los encontradas en El Niñ de 1972.
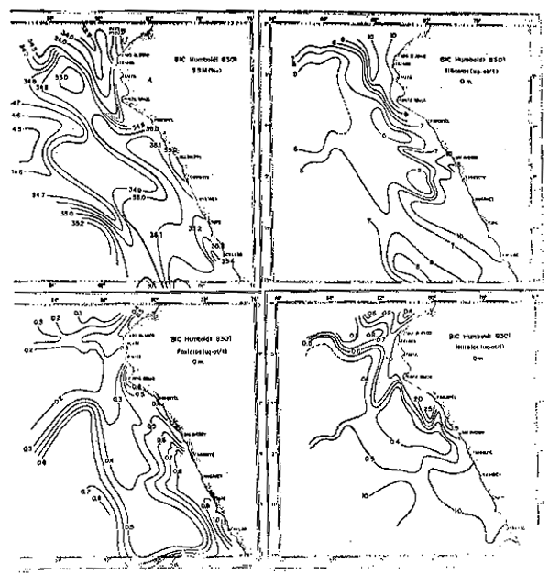
Fig. 24. - Salindad, fosfatos, silicatos y nitratos a Om en enero—febrero de 1982 (de Zuta y Guillén, 1983.
En 1975, en febrero–marzo (Wyrtki, 1976) hubo una transgreión de aguas (Fig.25) de la región ecuatorial hacia el sureste, sin llegar a la costa peruana con características de un Niño, y luego se replegó en los meses de abril–mayo y fue considerado como un aborto de El Niño. Las Aguas Ecuatoriales Superficiales se hallaron con temperaturas mayores de 27°C y salinidades menores de 34.5 o/oo, asociado con bajas concentraciones de nutrien– tes y producción primaria, inferiores al promedio. A pesar de su corta duración tuvo efectos en la productividad del área costera peruana y presentó cambios bien marcados no sólo en las propiedades físicas, sino también químicas y biológicas. Durante el otoño de 1975 frente a Chile, los afloramientos no estuvieron bien desarrollados (Basten, 1978) a pesar de que las temperaturas cerca de la costa fueron inferiores al promedio.
Durante el Fenómeno El Niño el afloramiento costero continúa, pero las aguas subsuper– ficiales que afloran son calientes y pobres en nutrientes, dando como resultado un decre– cimiento de la cantidad de fitoplancton y un efecto adverso en el zooplancton, peces, etc. Comparando las areas de afloramiento del Noroeste de Africa, Oregon y Perú, se tiene que de las 3 regiones solamente en el Perú ocurre El Niño, a pesar que los vientos costeros no disminuyen (Wyrtki, 1975) y los procesos del afloramiento costero continúan.
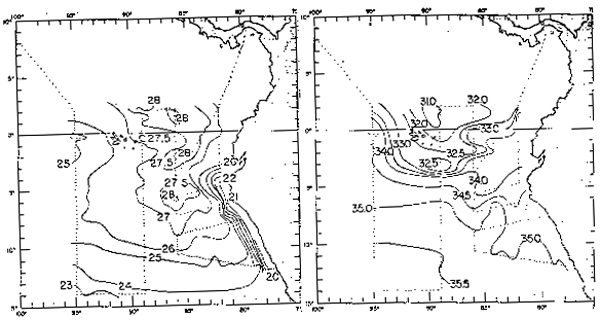
Fig. 25. - Temperatura y salinidad a Om en febrero–marzo y abril–mayo de 1975 (de Pattert et al., 1978).
Variaciones Estacionales de las Condiciones Oceanogáficas
Las variaciones estacionales tienen gran efecto en los diferentes niveles tróficos del ecosistema marino; y para mostrar estos cambios se han seleccionado secciones frente a Buenaventura, Galera, Islas Galápagos, Cabo Blanco y Arica, para el período 1964–1966 (Stevenson et al., 1970) en el que el año 1964 representa un año muy frío y el de 1965 un ano de El Niño.
En la sección frente a Buenaventura, una de las características del Panamá Bight (parte del Pacífico Oriental Tropical comprendida por el meridiano 81°W en el costado occidental, la entrada del Golfo de Panamá en el norte(71–72°N) y el costado sur a lo largo de 1°N) es la existencia de un domo termal que se puede observar por la curva ascendente de las isotermas que a veces llegan a la superficie. Dicho domo no señala necesariamente la presencia de afloramiento, ya que frecuentemente se encuentra debajo de la superficie.
La distribución estacional de la temperatura y salinidad superficial (Stevenson et al., 1970) sugiere que el domo se halla presente en la primavera del norte (febrero– marzo) y se encuentra centralizado cerca de los 5°N.
Las secciones verticales de temperatura (Fig. 26) muestran la evidencia del domo. Las figuras 26 y 27 muestran las variaciones trimestrales en la distribución de tempera– tura, salinidad y concentración de oxígeno disuelto.
En las secciones frente a Galera e Islas Galápagos, entre las aguas tropicales de alta temperature y baja salinidad del Bight y el agua fría y salina de la Corriente del Perú, se halla el límite frontal. El frente ecuatorial se extiende en dirección noroeste desde la frontera Ecuadore–Peru hasta pasar al noreste de las Islas Galápagos, el cual es más intenso cerca a la costa, debilitindose gradualmente hacia el oeste. La distri– bución de la temperature y salinidad en el frente ecuatorial varia estacionalmente y de un año a otro. Una característica del areá es la presencia de una célula (Stevenson et al., 1970) entre la costa y las Islas Galapagos cuyo tamano y ocurrencia varia estacio– nalmente. El frente ecuatorial de enero a marzo es mis meridional y débil, y la célula no está presente. En abril-junio cuando la velocidad de la Corriente del Perú aumenta puede formarse la célula. Las figuras 27 a 30 presentan las variaciones estacionales de la temperature, salinidad y oxigeno disuelto. La variacion de la Corriente Subsuperfi– cial Ecuatorial, caracterizada por su termostato y relativamente alto contenido de oxígeno disuelto, es observada durante El Niño de 1965 en el que se presentó en el verano e inten– sifico a principios del otoño. Stevenson y Taft (1971) muestran que las aguas de la Corriente Subsuperficial Ecuatorial ingresa cerca del Ecuador hacia el este con velocidades de hasta 37 cm/seg y luego dobla hacia sureste cerca de los 83°W y es hallado a 110 km de la costa del Ecuador y norte del Perú, caracterizada por su salinidad máxima de 35.13– 36.17 o/oo y temperatura de 17.2°C.
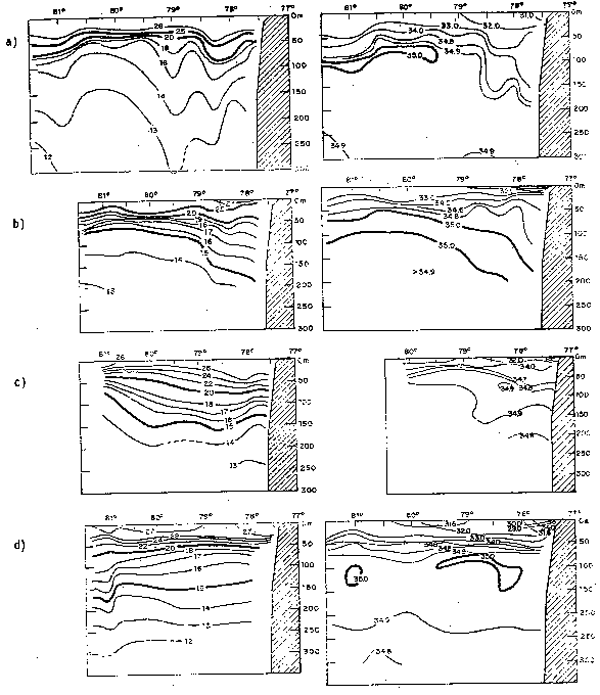
Fig. 26.- Secciones de tempeature y salinidad frente a Buenaventura: a) Cr. 64–4, b) Cr. 65–1, c) Cr. 65–2 y d) Cr. 65–3 (de Stevenson et al., 1970).
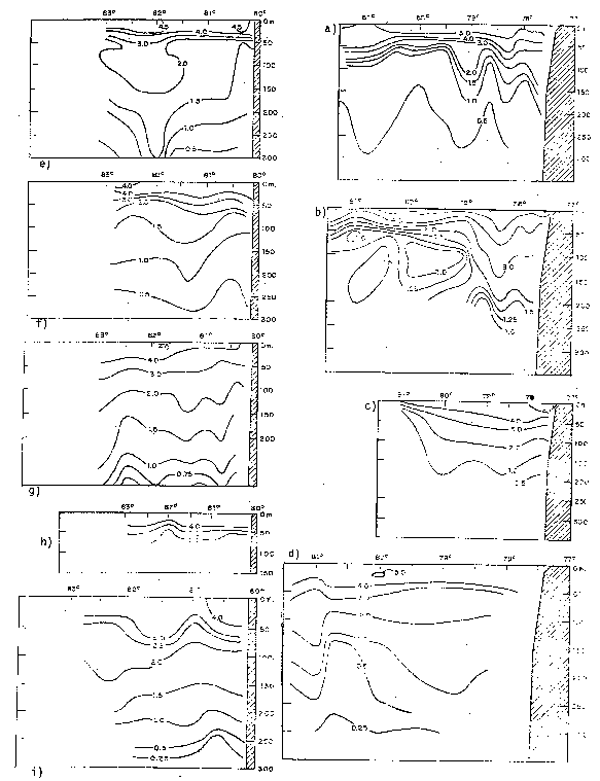
Fig. 27.- Secciones de oxígeno disuelto frente a Buenaventura y Galera: a) Cr. 64–4, b) Cr. 65–1, c) Cr. 65–2, d) Cr. 65–3, e) Cr. 64–4, g) Cr. 65–1, h) Cr.65–2, i) Cr. 65–3, (de Stevenson et al., 1970).
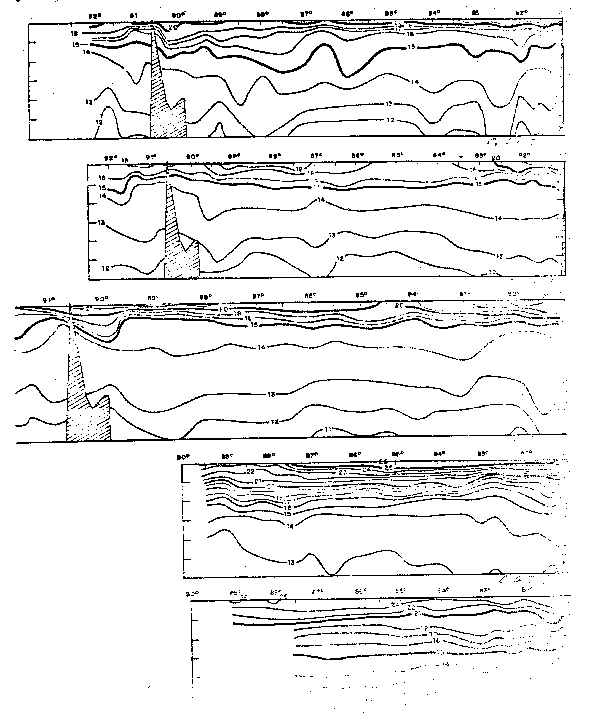
Fig. 28 - Secciones de temperatura frente a Galápagos:a) Cr. 64–2, b)Cr. 64–3, c) Cr. 64—4,d)Cr, 65–1 y e)Cr. 65–2 (de Stevenson et al., 1970).
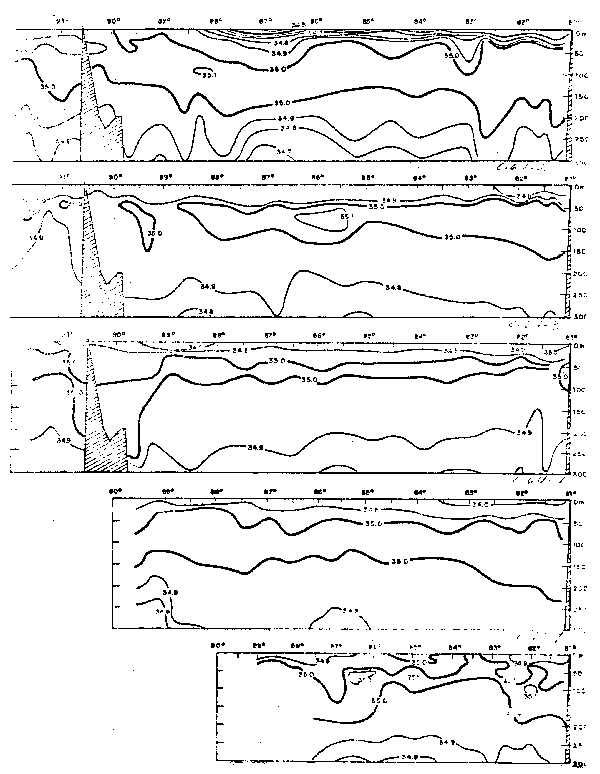
Fig. 29.- Secciones de salinidad frente a Galápagos: a) Cr. 68–2, b) Cr. 64–3, c) Cr. 64–4, d) Cr. 65–1 y e) Cr. 65–2 (de Stevenson et al., 1970).
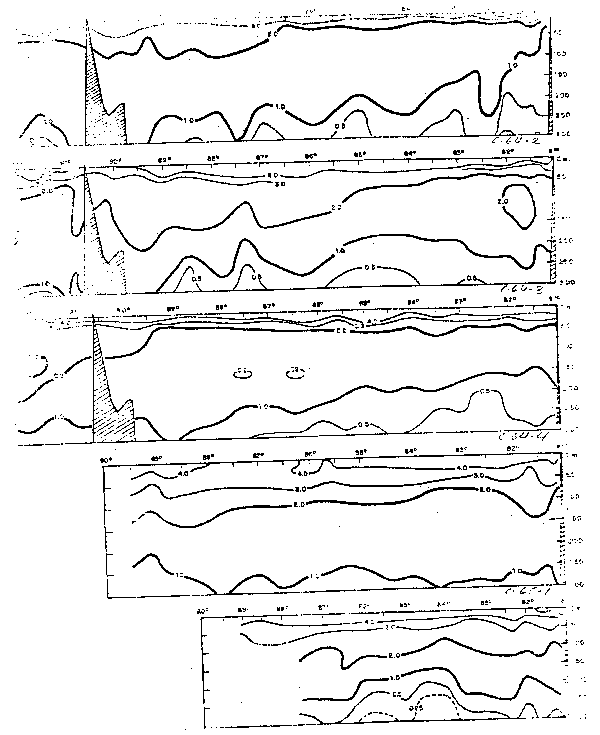
Fig. 30. - Secciones de oxígeno disuelto frente a Galápagos: a)Cr. 64–2, b) Cr. 64–3, c) Cr. 64–4, d)Cr. 65–1 y e) Cr. 65–2 (Stevenson et al., 1970).
La variabilidad de la estructura térinica es mostrada en la Fig. 31 en la que el Agua Tropical Superficial a fines de 1971 se halló en la capa de 30–40 m. Inmediatamente debajo se observó una marcada termoclina que se extendió de 35–50 m. Debajo de la termo– clina la estratificación fue mínima y abarcó la capa de 50–250 m incluyendo isotemas de 14°C y 13°C. Este termostato parece ser debido al flujo de las aguas de la Corriente Sub– superficial Ecuatorial. Los cambios más notables fueron observados a partir de febrero– marzo de 1972, con la profundización de las isotermas de 13–20óC. En agosto–septiembre 1973, las condiciones fueron similares a noviembre–diciembre 1971. Las figuras 32 y 33 muestran los grarides cambios de temperatura y salinidad de noviembre–diciembre 1971 a febrero–marzo 1972.
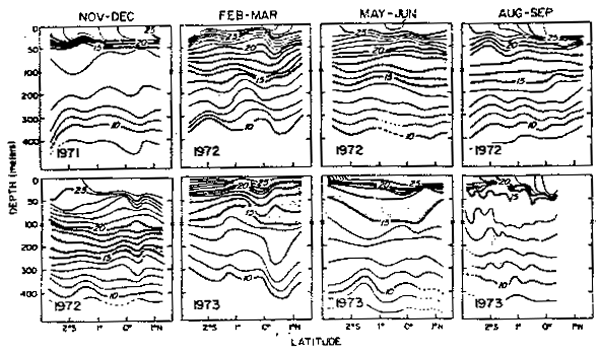
Fig.31.- Secciones de temperatura a lo largo de 82—83°W frente a Ecudor en noviembre 1971 - septiembre
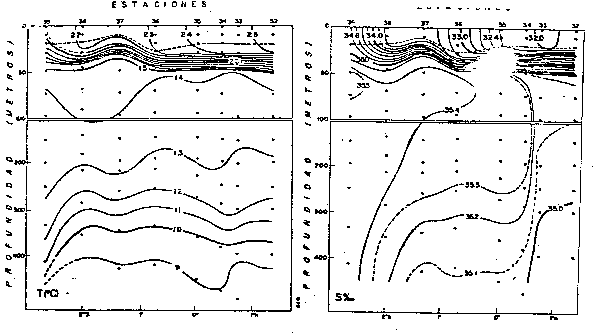
Fig.32.- Secciones de tempeatura y salinidad a lo largo de 82°30'W frente al Ecuador en noviembre-diciembre 1971 (de Enfield, 1976).
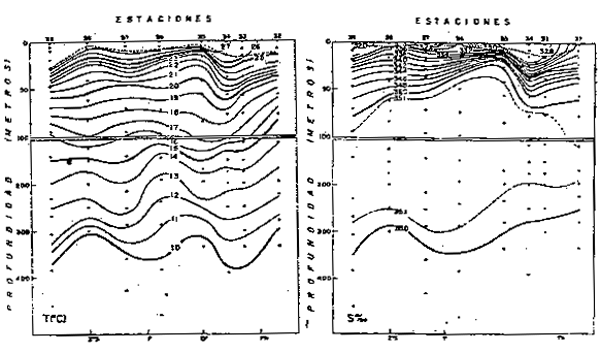
Fig.33.- Secciones de tempeatura y salinidad a lo largo de 82°30'W frente al Ecuador en febrero-marzo de 1972 (de Enfield, 1976).
En la sección frente a Cabo Blanco, la Corriente Peruana presenta a lo largo del Perú áreas de afloramiento, las cuales son identificadas por su bajatemperatura, bajo contenido de oxígeno disuelto, y alto contenido de nutrientes. Las aguas que afloran pertenecen a la Corriente Subsuperficial Ecuatorial y son procedentes de los 50 a 75 m de profundidad. La velocidad del afloramiento a lo largo de la costa del Perú ha sido estimada aproximadamente en 1.9 × 10-5 cm/seg (Posner, 1957) y de 5–30 × 10-5 cm/seg (Wyrtki, 1963).
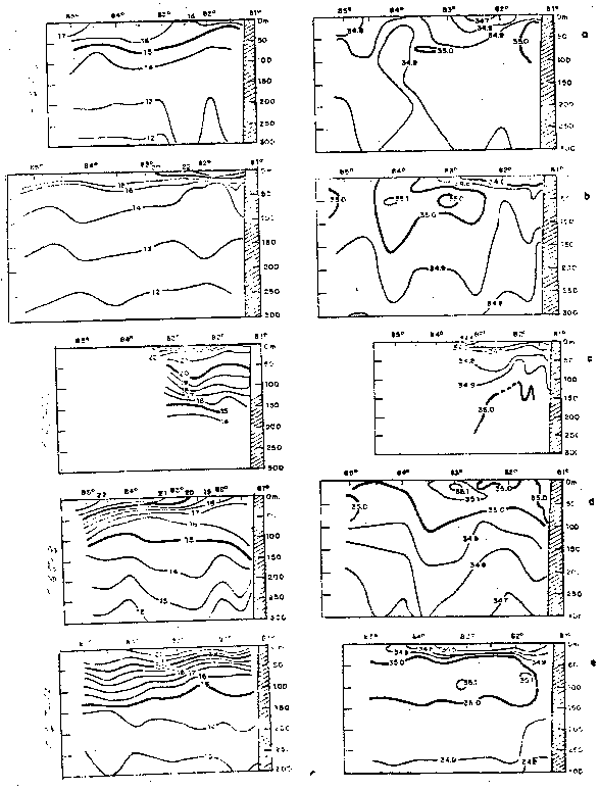
Fig. 34 - Secciones de temperatura y salinidad frente a Cabo Blanco: a) Cr. 64—3, b) Cr. 64—4, c) Cr. 65—2, d) Cr. 65—3 y e) Cr. 65—4 (de Stevenson et al., 1970).
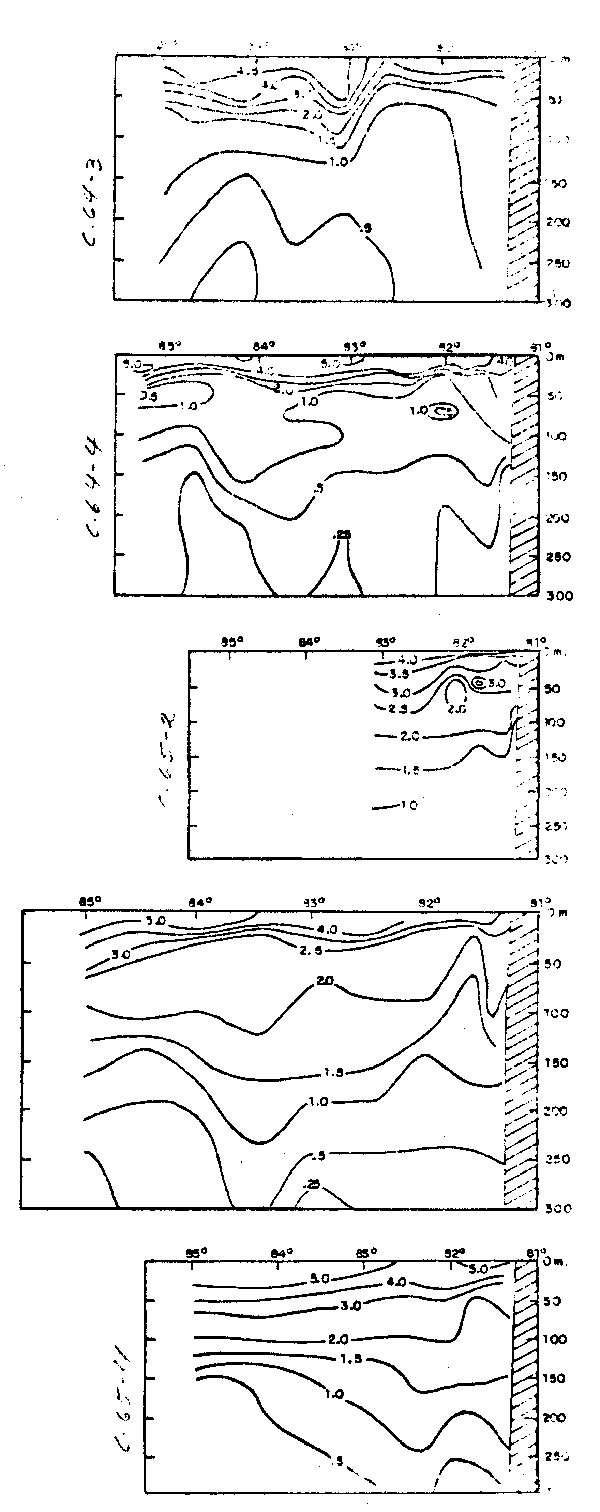
Fig.35.- Secciones de oxígeno disuelto frente a Cabo Blanco: a) Cr. 64–3, b) Cr. 64–4, c) Cr. 65–2, d)Cr.65–3 y e) Cr. 65–4 (de Stevenson et al., 1970).
La distribución durante El Niño 1965 (Figs. 34 y 35) es muy diferente a la del año frío y al promedio, tanto en la temperatura Como en la salinidad y oxígeno disuel to.
En la sección frente a Arica se observa una celda de agua cálida, salina (Figs. 36 y 37) a unas 100 y 200 millas al oeste de Arica, la que es poco protunda. En el verano dicha celda puede trasladarse hacia la costa cuando los vientos meridionales cesan. En la distribución vertical (Fig. 37) de la salinidad puede apreciarse la variabilidad de la Corriente Subsuperficial Peru-Chile que se caracteriza pos sus aguas más salinas y menor contenido de oxtgeno disuelto (Fig. 38) cerca de la costa, entre los 50 y 300 m de profundidad, la cual varía en extension y profundidad.
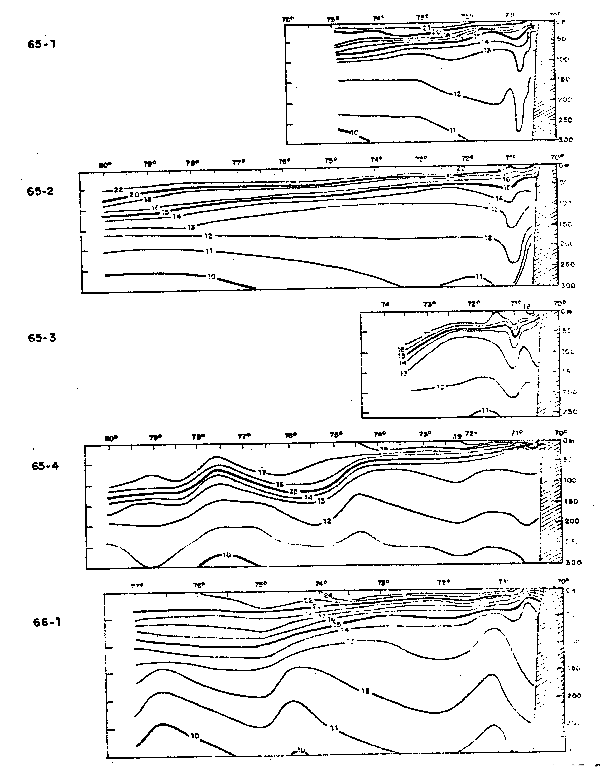
Fig. 36.- Secciones de temperatura frente a Arica: a) Cr. 65–1, b) 65–2, c) Cr.65–3, d) Cr. 65–4 y e) Cr. 66–1 (de Stevenson et al., 1970).
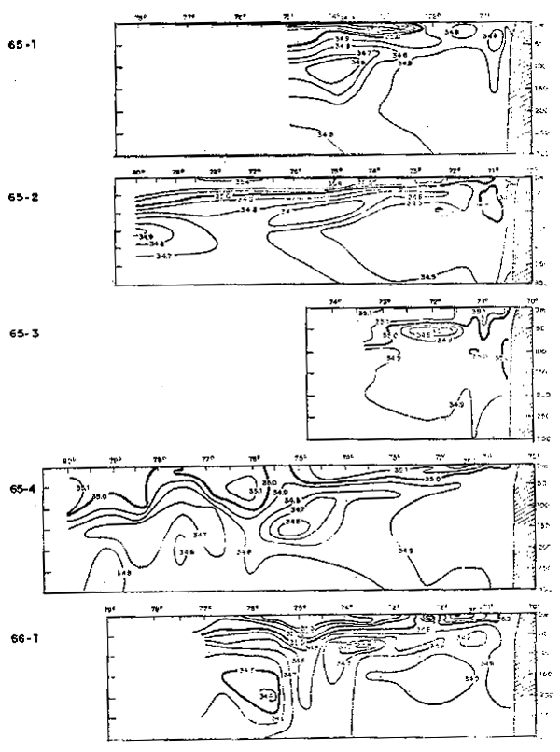
Fig. 37.- Secciones de salinidad frente a Arica: a) Cr. 65–1, b) Cr. 65–2, c) Cr. 65–3, d) Cr.65–4 y e) Cr. 66–1 (de Stevenson et al., 1970).
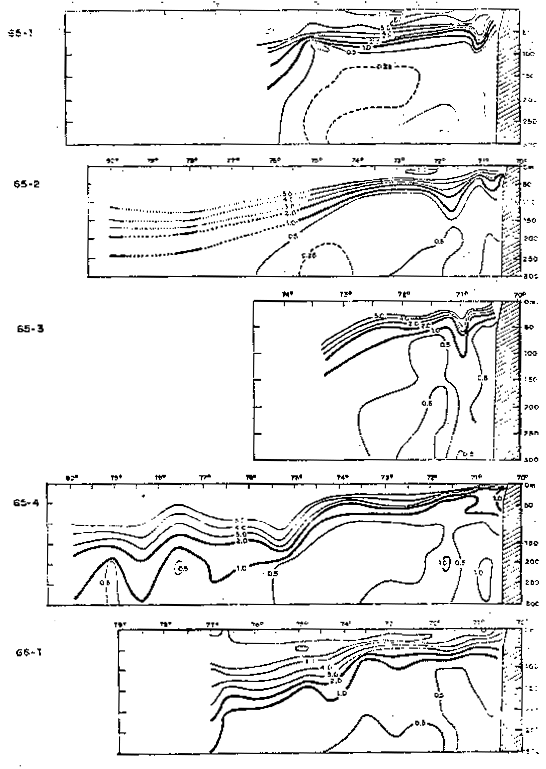
Fig. 38.- Secciones de oxígeno disuelto frente a Arica: a)65–1, b) 65–2, c) Cr. 65–3, d) Cr. 65–4 y e) Cr. 66–1 (de Stevenson et al., 1970).
En la superficie del mar igualmente se observaron cambios estacionales y anuales de las condiciones oceanográficas; así, comparando las condiciones promedio de Primavera y otoño con un año frío y años El Niño, se observan cambios en la distribución de la temperatura y salinidad, afectando la distribucion y desarrollo de los organismos vivos.
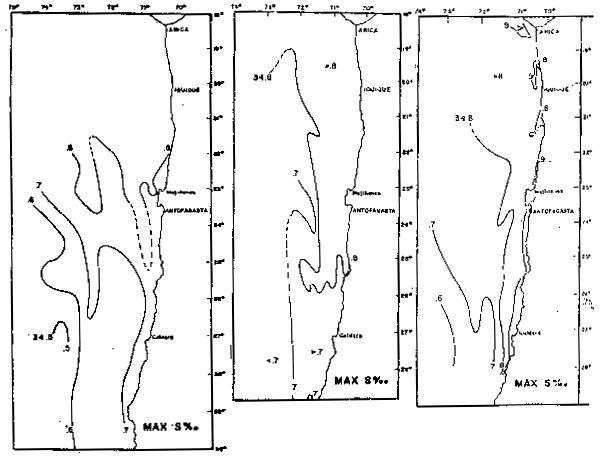
Fig. 39 - Distribucíon de la máxima de salinidad en el primavera de 1967 (a), 1971 (b) y 1972 (c) (de Robles et al., 1976).
Las variaciones anuales también pueden verse de la distribución de la máxima de salinidad (Fig. 39) de los años 1967, 1971 y 1972 (Robles et al., 1976).
VARIABILIDAD DEL MEDIO AMBIENTE Y SUS EFECTOS EN LA PRODUCTIVIDAD BIOLOGICA
Variabilidad del Medio Ambiente y sus Efectos en la Producción Primaria
La producción primaria depende de varios factores tales como: temperature, radiación, nutrientes, procesos de circulación y de mezcla, pastoreo del zooplancton, hundimientos del fitoplancton, etc. Uno de los mecanismos más impartantes para la fertilización de los océanos son los afloramientos, los cuales regulan la producción primaria.
Margalef (1978a, b) Vinogradov y Shuskina (1978), Walsh (1977) y Boie y Tomczak (1978) sostienen que los ecosistemas en los afloramientos son distintos de los ecosistemas marinos. Sin embargo, Cushing (1971 y 1978) señala que los ecosisternas de afloramiento costero no son ecosistemas distintos. Barber y Smith (1981) hacen una revisión completa de los ecosistemas de afloramiento costero y cornparan las areas de afloramiento de Oregon, Nbroeste de Africa y Perú.
Blasco et al., (1980) y Smith et al. (1980), frente al Perú en 1976, hallaron cambios día a día en la composicion del fitoplancton y zooplancton en una simple estacion, debido en gran parte al transporte advectivo.
Las mayores concentraciones de clorofila "a" y producción se hallaron en las áreas de divergencia, de mezcla vertical y en la capa de mezcla superficial. La más alta producción (Owen y Zeitzschel, 1970; y Forsbergh y Joseph, 1963) se halló a lo largo de la costa peruana, debido a] fuerte afloramiento (Fig. 40). La producción relativamente alta entre las Islas Galápagosy la costa fue debido al enriquecimiento de las aguas procedentes del afloramiento del norte del Peru.
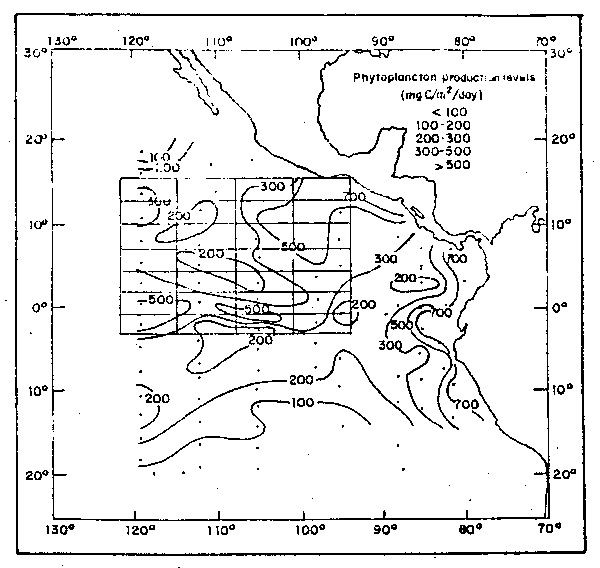
Fig. 40. - Producción primaria (mgC/m2día) en la capa eufótica de abril 1967 a marzo 1968 (de Owen y Zeitschel, 1970).
Frente a la costa de Chile el afloramiento costero es menor en comparación al Perú.
El afloramiento ecuatorial y la mezcla vertical explican la alta productividad cerca del Ecuador. Frente al Perú y Chile la producción varía estacionalmente de acuerdo a la calidad de las aguas afloradas y la intensidad y duración del afloramiento. La variabilidad del índice de productividad promedio frente a la costa peruana es mostrada en la Fig. 41, donde el mayor índice de productividad (>80 mg C/mg-Cl“a”/día) corresponds al área mayor de afloramiento. Frente al Perú afloran aguas de diferentes orígenes, dando por consiguiente, productividades diferentes; en algunos casos las aguas que afloran son procedentes de la Corriente Subsuperficial Ecuatorial que son mis productivas que cuando afloran las aguas subantárticas. La variación de la isopigna de 25.8 (Guillén y Calienes, 1981) muestra la divergencia en años fríos y años El Niño durante el verano e invierno.
Por otro lado, se ha observado que no sólo los años El Niño afectan a la produccion primaria sino también los años fríos (Guillén y Calienes, 1981), en el que a pesar del crecimiento intenso del fitoplancton, las aguas ricas en nutrientes contienen bajo contenido de células de fitoplancton debido a la inestabilidad de las aguas (Koblentz-Mishke, 1957), al rápido -hundimiento de los organismos o probablemente al más intenso pastoreo del zooplancton. De la distribución de anomalías térmicas, se observa los años en que ha ocurrido El Niño (1965, 1969, 1972, 1976 y 1982–1883) y los años fríos tles como: 1964, 1967–68, 1970–71 y 1974–75, lo que tiene, un marcado efecto en la producción primaria; así, en El Niño débil de 1975 (Cowles et al., 1977) se observo en la región ecuatorial (Figs. 42 y 43) una producción primaria menor de 0.2 gc/m2/dí, equivalents a la quinta parte del promedio para el área. Los efectos biológicos de 1975 fueron cortos y no afectaron grandemente a la costa peruana. En cambio, en 1972 la producción primaria frente a las costas del Perú fue de 0.39 gC/m2/día, un tercio de lo nomalmente hallado (Guilleñ y Calienes, 1981). Los efectos de El Niño y de los años fríos (La Niña) se puede observar en la distribución de la clorofila “a” (Fig. 44) en el que durante los años 1965, 1969, 1972 y 1976, la concentración fue bajísima y muy inferior al año promedio de 1966. Igualmente, los años fríos se caracterizaron por sus bajísimas concentraciones de clorofila “a”. Las figuras 45 y 46 presentan los cambios de producción primaria y clorofila “a”. frente a Cabo Blanco, de marzo a mayo de 1975.
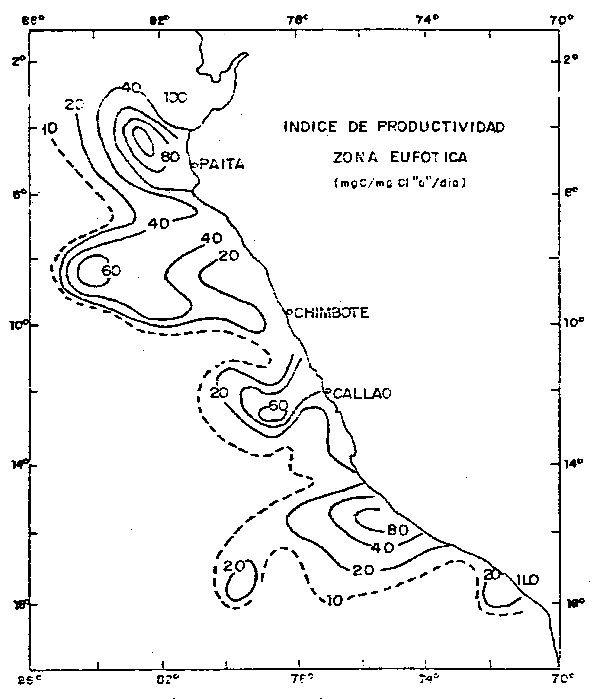
Fig. 41.- Indice de productividad promedio (mg/mg cl “a”/día) frente al Perú.
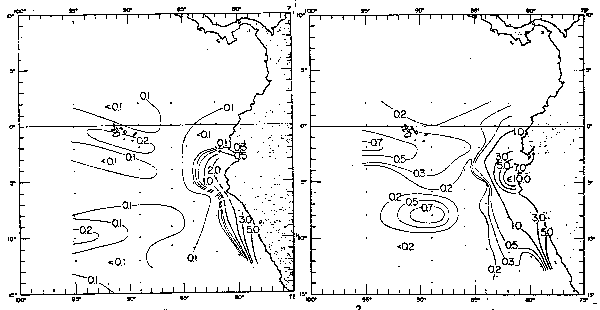
Fig. 42.- Distributión primaria (gC/m2/día) en la capa eufótica en febrero-marzo y abril-mayo de 1975 (de Patzer et al., 1978).
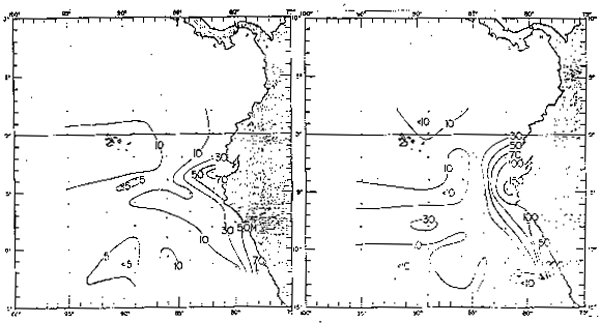
Fig. 43. - Distribución de la clorofila “a” (mg/m2) en la capa eufótica en febrero-marzo y abril-mayo de 1975 (de Patzert et al., 1978).
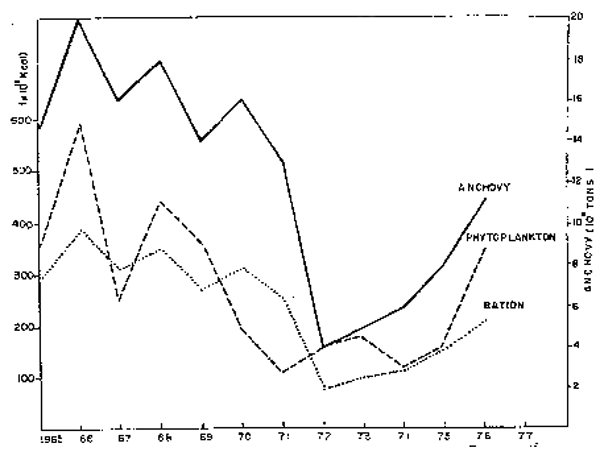
Fig. 44. - Biomasa de anchovesta (ton), fitoplancton (Kcal) y ración (Kcal) para el período 1965–77 (de Guillén y Calienes, 1981).
Fig. 45.- Secciones de clorofila “a” frente a Cabo Blanco en marzo y mayo de 1975 (de Patzert et al., 1978).
La productividad depende en gran parte de la composición química de las masas de aguas (Calienes y Guillén, 1981). Las aguas más productivas corresponden a las aquas frías de la Corriente Peruana con valores mayores de 800 mg C/m3/día en la superficie del mar, asociado con valores altos de nutrientes y clorofila "a" en la capa eufótica.
Fig. 46.- Seciiones de producción primaria frente a Cabo Blanco en marzo y mayo de 1975 (de Patzert et al., 1978).
Variabilidad del Medio Ambiente y Sus Efectos en la Pesquería
Las capturas mundiales de los últimos tiempos hanaumentado en forma espectacular de 19 millones de toneladas en 1950 a 61 millones de toneladas en 1980; y se espera un aument en los años futuros debido al aumento de la tasa de crecimiento demográfico mundial. Por otro lado, el desarrollo de modernos instrumentos de pesca, nuevas técnicas de localización y de captura, de barcos pesqueros y el equipo de procesamiento del pescado han aumentado las capturas en los océanos.
Las fluctuaciones naturales y distribución de las plantas y animales en los océanos son comunes. Algunas fluctuaciones son bien correlacionadas con los cambios observados en el ambiente físico. Otras fluctuaciones naturales tales como el florecimiento de una marea roja o la desaparicibn de una gran población, tal como la sardina del Pacifico, no son obviamente correlacionadas a los cambios en el ambiente físico, cuyo nicho biológico fue llenado por la anchoveta del norte, y su población desde entonces ha permanecido baja. La variabilidad natural del ecosistema y la sobrepesca han sido las causas principales para la desastrosa caída de la poblacion de la anchoveta peruana durante El Niño 1972.
La variabilidad del medio ambiente físico-químico no es el único factor que influye en la abundancia y la distribución de los recursos vivos, sino también los cambios de las actividades de pesca y las interacciones biológicas (Bakun et al., 1982).
Aún no se ha determinado cuáles son las variables ambientales más importantes en la variabilidad de las poblaciones de peces. Sin embargo, la distribución y abundancia de la mayoría de los recursos están relacionados directamente con la temperature, alimentación y oxígeno, principalmente.
Los pelágicos presentan dos ambientes bien definidos: la zona neritica que se caracteriza por sus aguas frías y su gran productividad, y la zona oceánica en la que predomina la Aguas Subtropicales Superficiales, que se caracterizan por sus aguas calientes y su baja productividad.
Alguna variabilidad puede ser también causada por cambios advectivos en la cantidad o clase de agua que es alimentada en la circulación costera por la Corriente Subsuperficial Perú-Chile, que fluye hacia el sur sobre la plataforma continental y en la parte superior del talud (Smith, 1978; Brockmann et al., 1980). Su core es bastante superficial sobre la plataforma continental y sus aguas aftoran en la mayor parte de los afloramientos a lo largo de la costa peruana (Smith et al., 1971). En forma paralela, la Corriente Subsuperficial Ecuatorial podría tener una importancia en la influencia advectiva en el ecosistema ecuatorial.
Durante El Niño, las aguas de afloramiento son desplazadas por aguas calientes, pobres en nutrientes y productividad, causando un efecto negativo sobre los organismos vivos. El efecto adverso sobre la anchoveta no es bien conocido, pero sí afecta el desove y a las anchovetas juveniles.
La revisión de la captura de las principales especies por países de la regíon nos permite seguir las fluctuaciones relatives a afios calientes y años fríos. Durante los eventos de El Niño, la captura de la anchoveta decrece mientras que otras especies tales como la sardina, jurel y cabana aumentaron notablemente en los siguientes años frente al Perú y Chile, debido a los grandes cambios oceanográficos. La calidad y cantidad del fitoplancton juegan también un papel muy importante como alimento para los siguientes niveles tróficos, ejemplo, la pesqueria de anchoveta en el Perú durante 1972 y 1976 (Fig. 44). Como resultado de los cambios bruscos del ambiente durante los años fríos y caiientes, se ha producido un cambio en la estructura del ecosistema marino, así como desplazamientos de ciertas especies, tal como lo ocurrido en el caso del desove de la sardina últimamente frente a las costas de Chile (Bernal, 1983), en el que se halló una nueva área de desove de s. sagax frente a Talcahuano y Golfo de Arauco. También se señala que los canibios interanuales de las variables biológicas no presentan una relación consistente con la ocurrencia de períodos frios y cálidos; sin embargo, se observó una relación inversa entre la irtensidad del desove de E.ringens y S. sagax y los períodos calidos. Frente a las costas del Perú, los cambios oceanográficos tienen un pronunciado efecto en el desove de la anchoveta (Fig. 47).
Fig. 47.-Distribución y abundancia de huevos de anchoveta en agosto 1967 y 1972 (de Santander y de Castillo, 1979).
La distribución biogeográfica según ambientes ecológicos, son: 1) "la Provincia Panamena, que abarca desde Bahía California, Colombia, Ecuador, hasta el extremo norte del Peru; 2) la Provincia desde Punta Aguja (06°S) hasta la Isla Chiloé (42°S); y 3) la Provincia, Patagónica. También debe considerarse la Provincia Oceánica en el que pedominan las Aguas Superficiales Subtropicales.
Durante los eventos de El Nino se encuentra a menudo especies de a"ura frente a la costa peruana y norte de Chile, tales como: el barrilete negro (Auxie rochei), el barrilete (Katsuwonus pelamis), el dorado (Coryphaena hippurus), Sierra (Scomberomorus maculatus), peces volorides(Exocoetus volitans), el cangrejo nadador(Euphylax dovii), etc.
En Panamá, la más alta captura hasta 1977 correspondio a la anchoveta Pacifico Central, con 165 mil toneladas en 1977; luego comenzo a decrecer hasta lograr valores de 4 mil toneladas en 1979. Le sigue las capturas del arenque de hebra o pinchagua, que a partir de 1972 se observó un aumento alcanzando una captura de 116 mil toneladas en 1979. Luego le sigue el atún, el barrilete y los camarones Panaeus sp.
En Colombia la mayor captura correspondió a los characidos, luego le siguen los silúridos, anchoveta del Pacífico Central y róbalos.
En Ecuador la mayor captura correspondió a la pinchagua, que a parrir de 1972 aumentó su captura hasta lograr 575 mil toneladas en 1979; luego le siguen el atún y el barrilete. Sin embargo, la pesca tradicional y de importancia economics es la captura de camarones.
En el Perú, la captura de anchoveta ha declinado de 10.28 millones de toneladas en 1971 a 4.72 millones de toneladas en 1972, y luego ha continuado disminuyendo su captura. Las mayores capturas a partir de 1973 corresponden a la sardina, el jurel y la cabana con 1.17 millones, 68 y 42 mil toneladas, respectivamente, en 1981.
En Chile, la captura de la anchoveta en 1972 fue de 368 mil toneladas para luegg deccrecer, seguida por la sardina y el jurel con capturas de 1.5 y 866 mil toneladas en 1981. Las capturas de langostas, la cosecha de cholgas y algas en el sr sun de importancia comercial.
El Niño de 1972-73 (Caviedes, 1981): Durante los meses de verano "a temperature superficial del mar a lo largo de la costa de Chile aunientó ligeramente sobre el promedio. El año de 1972 (Caviedes, 1981) fue considerado como un ano anomalo por las condiciones oceanicas a lo largo de la costa durante el temprano invierno. En el invierno se observó un fuerte y ancho flujo de agua caliente en direccion sur entre las 100 y 150 millas de la costa, el cual causó un debilitaifliento del afloramiento costero y una disminucion del flujo costero hacia el norte.
En diciembre de 1972, la temperature de las aguas frente a Arica. Antofagasta y Caldera estuvieron 2°C arriba del promedio (Caviedes, 1981)
Las condiciones océanicas anómalas en las aguas del norte de Chile no influyeron en el rendimiento de la pesquería pelágica sino hasta el invierno en 1972 y luego fue más pronunciado solainente en el verano de 1973. En enero de 1972 se captueó 44% de toda la anchoveta capturada en 1972 (367 913 ton), mientras que en los tres meses de verano 1973, sólo se capturo anchoveta en un 30% de la captura correspondiente a 1973 (191 795 ton) La captura de las otras especies de aguas frias tales como la sardina disminuyó; en cambio, la captura de peces de aguas tropicales, aumentaron.
Las anomalíes oceánicas de 1975 no fueron tan pronunciadas como 1972-73 (Caviedes, 1981). En 1976 las condiciones oceariográficas al norte de Chile fuerce restablecidas dando conio resultado un alto rendimiento en la captura de pelagicos.
Rougiere y Chabanne (1983), comparando las variaciones de salinidad y el número o peso de la captura de la atún, hallaron que el período de beja salinidad correlaciona muy bien con las grandes capturas de atún, con un coeficiente de correlación para el período 1976–82 de -0.56. Sin embargo, considerando sólo los períodos cuando la salinidad es inferior a 35.5 o/oo y arriba de 36.0 o/oo, el coeficiente de correlación fue de -0.82, indicando que hay una fuerte correlación entre la salinidad de las aguas superficiales y la tasa de captura de atún.
REFERENCIAS
Alarcón, E. 1975. Oceanographic conditions in coastal waters off Coquimbo. International Symposium Coastal Upwelling Proceedings. Coquimbo, Chile, November 18-19. Fundación Chile: 149–161.
Alarcón, E. y J. Pineda. 1969. Descripción oceanogrifica estacional de las aguas frente a Valparaóso. Bol. Cient. IFOP, Chile. 11–31 p.
Austin, T.S. 1958. Variations with depth of oceanographic properties along the equator in the Pacific. Trans. Am. Geoph. Union. 39 (6):1055–1063.
Bakun, A., J. Beyer, D. Pauly, J.G. Pope y G.D. Sharp. 1982. Ocean sciences in relation to living resources. Can. J. Fish Aquat. Sci. 39: 1059–1070.
Basten, J. 1978. Oceanografla de las aguas costeras del norte de Chile. Resultados del Crucero de Otoño de 1975 (mayo). Rev. Com. Perm. Pacffico Sur. 9:83–102.
Bennett, E.B. 1963. An oceanographic atlas of the eastern tropical Pacific Ocean based on data from EASTROPIC EXPEDITION, October-December 1955. Bull. Inter-Ariier. Trop. Tuna Comm. 8(2): 33–165.
Bennett, E.B. 1965. Currents observed in Panama Bay during September-October 1958. Bull. inter- Amer. Trop. Tuna Comm. 10(7): 399–457.
Bennett, E.B. 1966. Monthly charts of surface salinity in the eastern tropical Pacific Ocean. Bull. Inter-Amer. Trop. Tuna Comm. 11(1):3–44.
Barber, R.T. y R. Smith. 1981. Coastal upwelling ecosystems. pp. 31–68. In Analysis of marine ecosystems (A.R. Longhurst, ed.). Academic Press, New York, San Francisco.
Bernal, P. 1983. (MS). Variabilidad biológica interanual en la Corriente de California y en la región meridional del Pactfico Sureste. Presentado a la Tercera Reunión del Grupo de Trabajo Conjunto COI/OMM/CPPS sobre Investigaciones de "El Niño". 47 p.
Blasco, D., M. Estrada y B. Jones. 1980. Phytoplankton distribution and composition in the Northwest Africa upwelling region near Cabo Corbeiro. Deep-sea Res.
Boje, R. y M. Tomczak. 1978. Ecosystem analysis and the definition of boundaries in upwelling regions. pp. 3–11. In Upwelling ecosystems (R. Boje and M. Tomczak, eds). Springer-Verlag, New York.
Brandhorst, W. 1963. Descripción de las condiciones oceanográficas en las aguas costeras entre Valparafso y el Golfo de Arauco, con especial referencia al contenido de oxígeno y su relación con la pesca. Min. Agri. Chile. 1–55.
Brandhorst, W. 1971. Condiciones oceanograficas estivales frente a la costa de Chile. Depto. de Oceanologfa, LJ. de Chile, Valparafso, Chile. Rev. Biol. Mar. 14(3):1–20.
Brandhorst, W. y R. Cañón. 1967. Resultados oceanográfico-pesqueros aéreos en el norte de Chile. Pub. IFOP, Chile. 29:44 p.
Brandhorst, W. y 0. Rojas. 1967. Investigaciones sobre los recursos de la anchoveta y sus relaciones con las condiciones oceanográficas en agosto-setiembre de 1963 y marzo-junia de 1964. Pub. IFOP, Chile. 31:17 p.
Brandhorst, W. 1968. Investigaciones sobre los recursos de la anchoveta y sus relaciones con las condiciones oceanográficas en agosto a octubre de 1964. Pub. IFOP, Chile. 36:17 p.
Brockrnann, C., E. Fahrabach, A. Huyer y R.L. Smith. 1980. The poleward undercurrent along the Peru coast, 5°15°S. Deep-Sea Res. 27A:847-856.
Calienes, R. y O. Guilltén. 1981. Masas de aglia y producción primaria en el Perú. Bol. Inst. Mar Perú-Callao. Volumen Extraordinario ICANE, 155-163.
Caviedes, C.N. 1981. The impact of El Niño on the development of the Chilean fisheries. pp. 351- 368. In Resource management and environmental uncertainty: lessons from coastal upwelling fisheries (M.H.Glantz and J.D. Thompson, eds). Wiley Interscience, New York.
Cochrane J.D. y S. Zuta. 1968. (MS). Equatorial currents east of the Galapagos Islands in February- March, 1967. Texas A & IA University, Departinerit of Uceanography.
Cowles, T.J., R.T. Barber y O. Guillén. 1977. Biological consequences of the 1975 El Niño. Science. 195:285-287.
Cushing, D.H. 1971. A comparison of production in temperate seas and upwelling areas. Trans. Roy. Soc. South Africa. 40:17–33.
Cushing, D.H. 1978. Upper trophic level in upwelling areas. pp. 101–110. In Upwelling ecosystems (R. Boje and M. Tomczak, eds). Springer-Veriag. New York.
Enfield, O. 1976. Oceanograffa de la región norte del frente ecuatorial aspectos ffsicos. Reinión de Trabajo sobre el fenómeno conocido como "El Niño". Guayaquil, Ecuador, 4–12 de diciembre de 1974. FAO Inf. Pesca. (185):299–334.
Forsbergh, E. 1963. Some relationship of meteorological hydrographic and biological variables in the Gulf of Panama. Buit. inter-Amer. Trop. Tuna Comm. 7(1):1-109.
Forsbergh, E. 1969. On the climatology, oceanography and fisheries of the Panama Bight. Bull. Inter-Amer. Trop. Tuna Comm. 14(2):49–365.
Forsbergh, E. y W.N. Broenkow. 1965. Observaciones oceanográficas del Océano Pacrfico Oriental recolectadas por el barco Stioyo Maru, octubre 1963 - marzo 1964. Bull.inter-Amer.Trop.Tuna Comm. 10(2):84–237.
Forsbergh, E. y J. Joseph. 1963. Phytoplankton production in the South-Eastern Pacific. Nature. 200(4901):87-88.
Guillén. 1967. Anomialies in the water off the Peruvian coast during March and April 1965. Stud. Trop. Oceanogr. Miami. 5:452–465.
Guillén. 1971. The "El Niño" phenomenon in 1965 and its relations with the productivity in coastl Peruvian waters. pp. 187–196. In Fertility of the sea (J. Costlow, ed.). Gordon and Breach, New York.
Guillén. 1973. Carbon/chlorophyll relationships in Peruvian coastal waters. pp. 373-385. In Oceanography of the South Pacific 1972 (compiled by R. Fraser). Wellington, New Zealand National Commission for UNESCO.
Guillén. 1976. El sisteina de la Corriente Peruana. Parte I: Aspectos frsicos. Reunión de Trabajo sobre el fenórneno conocido como "En Niño". Guayaquil, Ecuador, 4–12 de dicimbre de 1974. FAO Inf. Pesca. (185):243–284.
Guillén, O. y R. Calienes. 1981. Biological productivity and El Niño. pp. 255-282. In Resource management and environmental uncertainty: lessons from coastal upwelling fisheries (M.H. Glatz and J.D. Thoryipson, eds). Wiley Interscience, New York.
Gunther, E.R. 1936. A report on oceanographical investigation in the Peru coastal currents. Discovery Rep. 13:107–276.
Inostroza, H. 1972. Atlas oceanográfico de Chile. Instituto Hidrográfico de la Armada, Pub.3041, Valparaiso, Chile. 171 p.
Inostroza, H. 1973. Some oceanographic features of northern Chilean waters in July 1962. pp. 37- 46. In Oceanography of the South Pacific (compiled by R. Fraser). Wellington, New Zealand National Commission for UNESCO.
Jones, H.J. 1973. Vertical mixing in the Equatorial Undercurrent. J. Phys. Oceanogr. 3(3):286–296.
Knauss, J.A. 1960. Measurements of the Cromwell Current. Deep-Sea Res. 4:265-286.
Knauss, J.A. 1966. Further measurements and observationes on the Cromwell Current. J.Mar.Res. 24:205–240.
Koblentz-Miske, O.I 1957. On phytoplankton production in the western north Pacific Ocean in spring, 1955. Dokaldy Acad.Sci.USSR. 116:1029–1031.
Love, C.(ed.). 1970–77. Eastropic Atlas. Cire.NOAA/NMFS. (33) Vol.I al II.
Margalef, R. 1978a. Life—forms of phytoplankton as survival alternatives in an unstable enviornment. Oceanologica Acta 1:493–509.
Margalef, R. 1978b. What is an upwelling ecosystem? pp. 12–14. In Upwelling ecosystems (R. Boje and M. Tomczak, eds). Springer-Verlag, New York.
Mujica R. 1972. Oceanograffa del mar peruano en Historia Marftima del Peru. Talleres Gráficos, perú. 1:221–474.
Owen, R.W. y B. Zeitzchel. 1970. Phytoplanton production: seasonal change in the Oceanic Eastern Tropical Pacific. pp. 32–36. In Marine Biology International Journal on Life in Oceans and Coastal Waters. Springer-Verlag, Germany.
Ponser, G.S 1957. The Peru current. Bull.Bingham. Oceanongr. Collect. 16(2):106–155.
parra, R. 1977. Resultados oceanográficos del Pacffico colombiano durante el año de 1976. Boletin Cientffico Armada Necional Cartagena, Colomia. 1(1):41P.
Reid, J.L. 1959. Evidence of a Souch Equatorial Countercurrent in the Pucific Ocean. Nature 184(4681):209–210.
Reid, J.L. 1961. On the geostrophic flow at the surface of the Pacific Ocean with respect to the 1000 decibar surface. Tellus. 1394):489–502.
Reid, J.L. 1962. On circulation phosphte–phosphorus content, and zoplankton volumes in the upper part of the Pacific Ocean. Limnol.Oceanogr. 7:237–306.
Robles, F. 1966. graphical description of oceanogrphic conditions off Tarapaca Province, based upon data from Marchil II Expedition. Instituto Hidrográfico de la Armada. Valparalso, Chile. 29p.
Robles, F. 1968 Descriptión gernral de las condiciones oceanográficas en aguas chileans. Sinópsis sobre algunos aspectos de las pesquerlas chilenas. Valparalso, Chile.
Robles, F., E. Alarcón y A. Ullao. 1976. Las mass de agua en la región norte de Chile y sus variaciones en un perfodo frfo (1967) y en perfodos cálidos (1969, 1971–1973). Reunión de Trabajo sobre el fenómeno consocido como “El Niño”. Guayaquil, ecuador, 4–12 de deliciembre de 1974. FAO Inf.Pesca. (185)94–196.
Rougerie, F. y J. Chabanne. 1983. Relationship btween tuna and salinity in Tahitian coastal waters. Tropical Ocean-atmosphere Newsletter. 17:12–13.
Santander, H y O.S. de Castillo. 1979. El ictioplancon de la costa peruana. Bol.Inst. Mar perú- Callao. 4(3):72–111.
Schaefer, M.,Y.M. Bishop y C.V. Howeard. 1958. Some aspects of upwelling on the Gulf of Panama. Bull.Inter-Amer.Trop.Tuna Comm. 3(2):79–131.
Sievers, H.A. y N. Silva. 1973 Variaciones estacionales de temperatura, salinidad y contenido de oxlgeno frente a la Bhfa de Valparalso (mayo de 1963-abril de 1969). Inv.Mar. 4(1);1–16.
Sievers, H.A. y N. Silva. 1976. Masas de aqua y circulation en el Oceanoéano Pacffico Sudoriental. Latitudes 18o5 33oS (Operación Oceeanografica Marchile VII). Reunión de Trabajo sobre el fenomeno conocido como “El Niñ”. Guayaguil, Ecuador, 4–12 diciembre de 1974. FAO Inf.Pesca. (185):197–231.
Silva, N. 1973. Variaciones estacionales de temperatura, slaindad y contenido de oxigeno en la zona costera de Valparaiso (setiembre de 1969 - agosto de 1970). Inv.Mar. 4(3):89–112.
Smith, R., D. Enfield, T.S. Hopkins y R.D. Pillsburg. 1971. La circulacíon en un sistema de afloramiento: Campana Pisco. Inv.Pesq. 35(1):9–24.
Smith, R.L. 1978. Poleward propagating perturbations in currents and sea levels along the Peru coast. J.Geophys.Res. 83:6083–6092.
Smith, S.L., K.H. Brink, H. Santander, T.J. Cowles y A. Huyer. 1980. The effect of advenction on variations in zooplankton at a single location near Cabo Nazca, Peru. pp. 400–410. In Coastal upwelling research, 1980 (F.A. Richards, ed.). American Geophysical Union, Washington, D.C.
Stevenson, N.M. 1970. Circulation in the Panama Bight. J.Geoph.Res. 75(3):659–672.
Stevenson, N., O. Guillén y J, Santoro. 1970. Marine atlas of the Pacific coastal waters of South America. Univ.Calif.Press. 23 pp. plus charts.
Stevenson, M. y B. Taft. 1971. new evidence of Equatorial Undercurrent, east of the Galopagos Islands. J.Mar.Res. 20(2):103–115.
Sverdrup, H.V., M.W. Johnson y R.H. Fleming. 1946. The oceans, their physics, chemistry and general biology. New York, Prentice-Hall. 1087 p.
Thomas, W.H. 1966. Surface nitrogenous nutrients and plytoplankton ib the northeastern tropical Pacific Ocean. Limnol.Oceanogr. 11:393-400.
Tsuchiya, M. 1968. Upper waters and circulation of the intertropical Pacific Ocean. John Hopkins Oceanogr.Stud. 4:50 p.
Vinogradov, M.E. y E.A. Shushkina. 1978. Some development patterns of plankton communities in the upwelling areas of the Pacific Ocean. Mar.Biol. 48:357–366.
Walsh. J.J. 1978. The biological consequences of interaction of the climate El Nino, and event scales of variabilty in the eastern tropical Pacific. Rapp.P,-v.Réun.Cons.Int.Explor.Mer. 173:182–192.
White, W.B. 1969. The Equatorail Undercurrent, the South Equatorial Countercurreny and their extension in the South Pacific Ocean, east of the Galapagos Islands during February-March, 1967. Texas A and M University Ref. 69-4-T, 74 p.
Wooster, W.S. 1952. Shellback Expedition 17 May to 27 August 1952. Scripps. Tech.Rep.Scripps Inst.Oceanogr. (SIO Ref. 52–63). 14 p.
Wooster, W.S. 1959. Oceanographic observations in the Panama Bight, Askoy Expedition, 1941. Amer.Mus.Nat.Hist.Bull. 118(3):117–151.
Wooster, W.S. 1960. “El Nino”. Rep.CalCOFI. (7):43–45.
Wooster, W.S. 1961. Further evidence of a Pacific South Equatorial Countercurrent. Deep-Sea Res. 8(3/4):294–297.
Wooster, W.S. 1969. Equatorial front between Peru and Galpagos. Deep-Sea Res. 16 Suppl:407– 419.
Wooster, W.S. 1970. Eastern boundary currents in the South Pacific. Scientific Exploration of the South Pacific. National Academy of Sciences, Washington D.C. 60–68.
Wooster, W.S., y T. Cronwell. 1958. An oceanograohic description of the eastern tropical Pacific. Bull.Scripps Inst.Oceanogr. 7(3):169–281.
Wooster, W.S., y M. Gilmartin. 1961. The Peru-Chile Undercurrent. J.Mar.Res. Vol. 1963, Bigham Oc. Lab. Yale University.
Wooster, W.S. y J.L. Reid. Eastern boundary currents. In The sea (M.N.Hill,ed.). New York, Vol. 2:253–280
Wooster, W.S., T.J.Chow y I.Barrett. 1965. Nitrite distribution in Peru water. J.Mar.Res. 23(3):210–221.
Wooster, W.S. y O.Guillán. 1974 Caracteristicas de “El Nino” en 1972. Bol.Inst.Mar perú–Callao. 3(2):44–71.
Wyrtki, K. 1963. The horizontal and vertical field of motion in the Peru Current. Bull.Scripps Ist.Oceanogr.8(4):312–346.
Wyrtki, K. 1964. The thermal structure of the easter Pacific Ocean. Dt.Hydrogr.Z., Ergan- zungsheft, Ab. 84p.
Wyrtki, K. 1965. Surface currents of the eastern tropical Pacific Ocean. Bull.Inter-Amer.Trop. Tuna Comm. 9;271–304.
Wyrtki, K. 1966. Oceanography of the eastern equatorial Pacific Ocean. Oceanogr.Mar.Biol. Ann.Rev, 4:33–68
Wyrtki, K. 1967. Circulation adn water masses in the eastern equatorial Pácific Ocean. Inst.J. Oceanol.and Limnol. 1(2):117–147.
Wyrtki, K., E. Stroup, W. Patzert, R. William y N. Qunn. 1976. Predicting and observing El Nino. Science. 191:343–346.
Zuta, S y O. Gulleén. 1970. Oceanograffa de las aguas costeras del perú. Bol.Inst.Mar Perú–Callao. 2(50;161–223.
Zuta, S. y W. Urquizo. 1972. Temperatura promedio de la superficie del mar frente a la costa peruana, perfodo 1928–1969. Bol.Inst.Mar peru-Callao. 2(8):462–519.
Zuta. S., D. Enfield, J. Valdivia, P.Lagos y C.Blandian. 1976. Aspection ffsicos del fenó meno “- Nino” 1972–73. Renunion de Trabajo sobre el fenomeno conocido como “El Nino”. Guayaquil, Ecuador, 4–12 de diciembre de 1974. FAO Inf.Pesca. 9185):3—61.
Zuta,S. y Q. Gullén. 1983 (MS). Informe del avance del Programa Oceanográfico del EREEN en el perú en 1981–82. Tercera Reunion de Trabajo del Comité Cientifico del CREEN. Cali Colombia, 12p.
by
J.R. Hunter1 and Gary D. Sharp2
1Southwest Fisheries Center La Jolla Laboratory
National Marine Fisheries service
NOAA, P.O. Box 271
La Jolla, CA 92038
USA
2Fishery Resources and Environment Division
Fisheries Department
FAO, Vai delle Terme di Caracalla
00100 Rome, Italy
Resumen
Entre los problemas de proyectar la producción de la pesca están la gran cantidad de variables complejas, dentro de las cuales la producción primaria y los procesos fisicos que promueven esta producción tienen gran importancia. Se revisa una vaz más la importanica de las escales de tiempo y espacio en relación a various procesos biologicos de importancia relacionados con la pesca, y se describe una clase especial de fenómeno fisico y sus caractreisticas.
Los frents marinos marinos en la plataforma derivan de numberosos procesos: frentes en la interrcupcion de la plataforma; frentes costeros; corripentes sobre la plataforma impusados par el viento; frentes causdos por las mares. Estos se describen en suficiente detalle como para que sus mecanismos pudan ser comparados, y se describen las caracteristices de los frentes marions de la plataforma. Estos son procesos determinados por las estaciones y la geografía.
Los requerimientos pora que se forme un frente son de dos tipos:
a) Debe haber sufuciente energia (del viento a las mareas) para formar un área de mezcla, con una región adyacente donde el grado de mezcla, cpm ima región adyacente donde el grado mezcla es menor y puedo ocurrir estratificación.
b) para que ocurra la estratificación dade haber una fuerte presión o empuje hacia arriba del fluido ya sea por el incremento de la densidead capas superiores, por ejemlo lluvia o pérdida de calor.
Si se tiene information sobre la observación de energia y los procesos de estratificación es posible hacer una serie de prediciones sobre dónde se pueden generar los frentes marinos en la platforma. Las Principales áread de plataforma continental de los océ del mundo se han ordenado para mostrar las regiones potenciales para la formación de frentes inducidos por mareas. Estos parecrian ser útiles para relcionar con áreas de producción de peces plagico-beriticos, tal como fuera sugerido por Illes y Sinclair (1982) pra Clupea harengus frente a la costa este de Canadá, y posiblemente merenca una mayor atención en relación en relació al potencial pesquero y sus variaciones.
INTRODUCTION
Since the advent of concern about the long-term productivity of the seas in the face of man's predation and perturbations, as reviewed in Thomasson (ed. 1981), there have been numerous general attempts to quantify system potentials, either via attempted simplification utilizing trophic concepts (Hardy 1924, Steele 1974, Cushing 1971), or through study of life table statistics arising from samples taken from fisheries and research ventures on many species (Beverton and Holt, 1957, Ricker 1958, Andersen and Ursin 1977). Numerous techniques have been applied toward obtaining estimates of the amounts, distributions and status of fishery resources. As yet no technique has been particularly successful at projecting fishery production, primarily due to the species and physical oceanographic variations arising from complex climatic and biophysical interactions.
As various management regimes have failed to stabilize fish populations, and there fore production for one reason or another (e.g. Saetersdal 1980), the recent generation of stock assessment specialists is once again opening their ranks and fora to discus sions of what variables beyond fishery statistics need to be considered in order to resolve or interpret the variations in population production.
There have been few revolutionary concepts introduced since the early insights of Johan Hjort (1914); Baronov (1918), Michael Graham (1935); or Peterson (1894). The variety of permutations and analytical approaches arising from these original observations, assertions, and interpretations are reasonably well documented in the recent surge of “how to” stock assessment manuals (Laurec and Le Guen 1981, Beyer and Sparre 1983, Pauly, M S). The latter reference even provides examples and programmes on magnetic cards for use with small desk top programmable calculators. These manuals cover virtually all conventional stock assessment techniques. This makes available the possibility to do the rather trivial mathematical operations in single species stock assessment to anyone capable of acquiring data and entering them into a keyboard, and leaves far more time to think about the more difficult problems of interpreting the results, and deciding what new or other parameters need to be incorporated in order to rationalize fishery management.
The more complex models is incorporating more than one species are not so readily available. Laevastu and Larkins (1981) have provided an introdution to the ecosystem model which evolved at the Northwest and Alaska Fisheries Center in Seattle, Washington. This model comprises many thousands of computer statements and as such cannot be rightly condensed so as to operate on a desk calculator. Laevastu has howevercreated a “teaching tool” version of the biological interactions segments of the ecosystem model which is available upon request.
Perhaps more important is the general trend in development of the conventional wisdoms to include, once again, ambient environmental (biotic and abiotic) parameters into stock assessment techniques. This has come about through the increasing awareness of the importance of early life history survival requirements, and causes of mortality associated with predation, starvation, and environmental perturbation, i.e. pollution or habitat destruction.
No attempt will be made here to review the so–called stock–recruitement problem, as this has been quite thoroughly done by Ricker (1954); Sharp and others in IOC Workshop Report No. 28 for oviparous fishes; Garcia (in press) for penaeid shrimp; and Hennemuth et al., (1981) for a variety of species. The further information needed in relation to the S/R problem is outlined in Bakun et al., (1982), and Sharp (1981a) and will be one subject of the Expert Consultation to Examine Changes in Abundance and Species Composition of Neritic Fish Stocks in Costa Rica from 18 to 29 April 1983.
What is becoming increasingly apparent is the relevance of smaller scale ocean processes in setting the limiting survival factors for each component of potential breeders in marine populations. This follows directly from the insights of the early fishery scienctists mentioned above, but has grown to be of major research concern only in the recent decades. This is not to say that there has not been substantial continu– ing effort since G.M. Dannevig released the first few hundred thousand Gadus morhua larvae hatched at the F1φdevigen Sea Fish Hatchery in 1886. But even at that time there were heated arguments over what caused the natural variations in cod populations. The eventual instigator of the “critical period” concept, Johan Hjort, was one of the leading critics at the onset of this project. Now, one hundred years later, Norwegian scientists are re–emphasizing the field of enhancement of natural populations through laboratory rearing, sea ranching and release programmes, with scaled feasibility studies as a basis for each stage in development (Sharp 1981c).
The major advocates of increasingly sophisticated studies of larval fish requirements have been Rosenthal (1970), Jones (1974), Lasker (1975, 1978), Laurence and Beyer (1981),Houde (1974, 1975, 1978 and 1980) and recently IMARPE staff at Callao, Peru. Theilacker and Dorsey (1981) reviewed relevantresearch results and have made several observations and recommendations regarding required standardization of techniques and reporting of results of studies of larval fish feeding, survival and growth, if such studies are ever to be compared in a meaningful fashion.
Cannibalism and predation on larval and juvenile fishes is recognized as the major mortality source resulting from other than larval fish feeding requirements (MacCall 1981; Smith and Lasker 1978; Andersen and Ursin 1977; Alvariño 1980).
Other parameters which need more attention are those for physical processes which have been identified as important to the early life history stages and adults as well, namely gyres, frontal systems, horizontal microstructure and eddy systems in general (Bakun and Parrish 1981, Parrish et al., 1981, Owen 1981, Grainger 1980; and Illes and Sinclair 1982). These authors have discussed various neritic fish resources in respect to the physical transport phenomena associated with near shore , coastal fish populations.
Reid et al., (1978) and Voronina (1978), have described oceanic zoogeographic prov– inces in relation to ocean-scale circulation. The reviews of the hydrodynamics of semi– enclosed seas such as the Gulf of Mexico, the Mediterranean Sea and the Baltic (Nihoul, ed. 1982) indicate strong eddy and gyral phenomena dominate these areas, certainly having significant biological consequences.
What is needed is a catalogue, so to speak, of processes, habitat features and their geographic and temp oral distributions so that it might be possible to better understand the phenomena which promote or delimit neritic and oceanic fishery resource production potentials in their realistic, dynamic form.
Illes and Sinclair (1982) have stimulated this particular publication by pointing out the dependence of herring (Clupea harengus) populations, i.e., breeding components, upon gyral strengths, distributions, andpersistence. What is implied is that the discontinuity of occurrence of a specieson the short or long-term, indicates that there are specific, physically mediated requirements which occur within the same pattern of the species distribution. These are often physical processes which promote essential productivity of food, and, in the case of Atlantic herrings, simultaneously maintain larval coherence within the optimal survival conditions. The more mobile juvenile and adult stages are less “controlled” by the local physical processes, but respond in the case of drastic changes.
Similar phenomena and other transport processes are described by Parrish et al., (1981) which they use to explain the reproductive behavior, species success and general zoogeography of the California Current. Sharp (1981b) reviewed numerous interactive phenomena scaled in relation to species mobility ,larval requirements, food production and homing versus opportunistic species which can account for the vast behavioral array and species diversityobserved in most regions.
The concept of variable carrying capacities for either early life history stages or adults needs careful consideration. The implications for portrayals of species variations, monitoring and forecasting variations, or hindcasting variations in variously bahaving species are determined hierarchically by the magnitudes and variations in the important limiting physical processes, and whether these affect primarily early life history stages, or later stages. The near total dependence of early life history stages upon “local” conditions, in contrast to the numerous behavioral options available to more mobile stages, makes this an important consideration.
Now that the importance of early life history effects is better documented and various criteria for survival are being defined, it would be appropriate to step back from the fine scale processes, and look over the global and system scale properties of neritic zones as a first stage in focussing on the system dynamics relevant to fishery resources and their relative fragility.
Upwelling areas have been well documented as the sources of particularly high coastal nutrient and subsequent biological production (recent review in Richards 1981, Cushing 1972, Glantz and Thompson 1981, and Szekielda et al., 1977). Recent efforts to identify other sources of productivity (Gieske et al., 1979, Owen 1981, Pingree et al., 1978 and Gregg 1980) point to the thermocline - picnocline and their vertical counterparts, recognized as fronts of various types as widely occurring features harboring production in various forms and scales, from meters to kilometers in horizontal dimensions and centimeters to meters in vertical dimensions. Mann (1982) reviews production and energy transfer in coastal systems. Fasham (1978) reviewed the relative inadequacies of conventionally obtained plankton data in resolving these relevant scales of occurrence of ocean productivity. It appears that we are entering another age of discovery in this field , particularly as one takes into account the spontaneity of primary production processes and the responses of fish populations. The relative persistence, dynamics and interactions of fish populations and their invertebrate counterparts,are tied in various fashions to the same physical processes as promote, isolate ,and dis– perse the plankton populations supporting the food webs. As the fish resources usually start out in the plankton, graduating in size and relative independence with time , these physical features which promote production and /or aggregation need to be better defined, monitored and evaluated if we are ever to understand the true resource potential, their dynamics and their distributions. Sverdup was saying this 30 years ago (1952, 1955), Chapman 10 years ago (1972) and Bakun , Beyer, Pauly, Pope and Sharp said it again last year (1982) with perhaps a few more direct links between fish, plankton and physical processes from which to begin.
Shepherd, Pope and Cousens (ICES 1982) have summed up rather well the distinctions between various environmental processes which are likely to directly affect fish resources, from 1) direct physiological effects due to temperature, oxygen,salinity, pressure, etc; 2) disease; 3) feeding effects such as food abundance, food quality, time–space variations, localization of concentrations and competition; and 4) predation, the result of one individual being food for another, hence starting the cycle in 3) again. That these authors were also able to convince themselves that there are indeed statistically verifiable links between young fish survival and climate is satisfying, but not news.
In regard to production of food and sustenance offish populations, one source of primary production and aggregation phenomena which has only recently been given due consideration is the phenomenon of shelf sea fronts (Anon., 1981). Inthis collection of studies Holligan (1981) characterizes the primary and related secondary production emanating from northwest European continental shelffronts. He also classifies variously generated fronts,describing shelf-break fronts, coastal fronts, and the relatively important tidal fronts which have been observed in varying detail. Such observations have been made for the Ushant frontal system which extends around the coast of Brittany into western English Channel (Pingree et al., 1975, 1977, 1979, Grall et al., 1980), the Celtic Sea front between Ireland and Wales Savidge 1976; Savidge and Foster 1978, fronts in the Irish Sea (Savidge 1976;Foster et al., 1976; Beardall et al., 1978), the Isly front between northern Ireland and Scotland Pingree et al., 1978; Simpson et al., 1979) and the fronts around the Orkney and Shetland Isles (Pingree et al., Dooley 1978, and southern North Sea (Harding et al., 1978).
Allen and Smith (1981) describe wind driven shelf currents off Oregon, northwest Africa and Peru, each being uniquely controlled, some more by wind, others more by longshore currents and topography. The common denominator of these currents and shelf frontal systems is that they both promote eddy formations (Simpson 1981; James 1981), which through initial nutrient injection can set primary production going and zoo- plankters and fish aggregating into these systems may provide sufficient additional and recycled nutrients to sustain this production over a relatively longer period. The vortices or eddies may provide perfect larval fish development conditions (Illes and Sinclair 1982) and larger eddies, as for example off Southern California (Owen 1980, 1981, 1982) may provide adequate nutritional aggregation for sustaining fish reproduction, coherence of reproduction products through subsequent developmental processes, and even maintain the young fishes within a specific geographicsituation which might not sustain some pelagic populations otherwise (Parrish, Nelson and Bakun 1981).
In recognition of the relevance of shelf fronts to fish aggregation and repro– duction, particularly tidally generated shelf fronts, the following general des– cription of what they are all about is offered.
SHELF SEA FRONTS
Tidal shelf sea fronts are regions of strong horizontal variation in density, separating vertically stably stratified and well mixed regimes. Vertical density stratification is generated when there is some buoyant input to the sea, such as through surface heating, or runoff of fresh water (the majority of known tidal fronts are associated with the former mechanism). When water is stably stratified, it has lower potential energy than the same system would have after complete vertical mixing (since its center of gravity is lower). Hence vertical mixing of the water column requires the input of energy. This comes primarilyfrom tidal stirring and from the surface wind stress (on the North West European shelf, the former is more important (Simpson, Allen and Morris, 1978), while in areas such as the Arabian Gulf tides and wind may exert comparable influences (Hunter, 1982).
The balance between the input of buoyancy (and consequent lowering of the potential energy of the water column), and the addition of energy through mixing determines whether stable stratification will occur. If buoyancy input is high we may get stratification, while if mixing is high we may not. In regions where the buoyancy flux and/or the tidal mixing changes markedly in the horizontal, we may find achange from one condition to the other, generally associated with a strong horizontal gradient (a“front”).
The energy argument for the occurrence of tidal fronts was originally given by Simpson and Hunter (1974) who concluded that the presence of such fronts should be predictable from the parameter
where h is the water depth,
and u is the tidal stream velocity
(the reciprocal of this parameter is proportional to the tidal dissipation per unit volume ofsea water).
Since the tidal velocity occurs as a cubic power law in the above expression, this parameter covers a wide range in the shelf areas of the world - on the North West European shelf it varies over three orders of magnitude (Simpson, Hughes and Morris, 1977), which means that there is ample scope for the formation of tidal fronts in this region.
Shelf seafronts have generally been associated with buoyancy (i.e., less dense water) added at the surface (e.g., by heating, or freshwater inflow). However, vertical stratification can also be caused by the addition of negative buoyance by processes such as surface evaporation. This case is some what different from those considered already, in that surface addition of negative buoyancy (dense water) yields an unstable system denser water overlying light water) which will overturn, generating some mixing as it does so. However, the resultant water, even if vertically mixed, will still be denser than the water in an adjacent region where the evaporation may be less, and this horizontal density gradient can give rise to vertically shearedflows, the denser water passing underneath the adjacent lighter water. This phenomenon is common in tropical seas where evaporation is often a dominant buoyancy flux - dense water is formed in shallow areas where evaporation is highest (both due to the lower relative humidity near the coast, and (in summer) to the higher sea temperatures near the coast) and where a given rate of evaporation yields a greater density change, leading to an “inverted estuarine” circulation (seaward at the bottom, shoreward at the surface). A good example is the Arabian Gulf. It is quite feasible that horizontal fronts could be generated in such areas by lateral variation of tidal and wind mixing processes.
The physical and biological implications of sea fronts were recently reviewed at a conference organized by the Royal Society (Anon., 1981).
THEORY
A simplified derivation of the “h/u3” law will be given here. More detailed accounts have been given by Simpson and Hunter, 1974; Fearnhead, 1975; Simpson, Allen and Morris, 1978; and Simpson and Bowers, 1981.
Buoyancy may be described as a density anomaly in the sea water, and its “concentration” may be defined quantitatively by
-g(ρ - ρ0)
where g is the acceleration due to gravity,
ρ is the sea water density,
and ρ0 is a constant reference density
(note that ρ - ρ0 >> ρ0).
As this factor depends only on density ,it is an intrinsic property of the sea water, and as such is conserved in any system when due consideration is made of any sources and sinks of buoyancy. Buoyancy may be modified by the addition or removal of heat or salt, and it may be transported aroundthe sea like any other “contaminant” (i.e.,by deterministic flows (“advection”) and by turbulence (“dif– fusion”)). For small additions of heat or salt, the buoyancy change will be proportional to the quantities of heat or salt involved (in the case of evaporation, the removal of fresh water may be interpreted as an addition of salt, if desired).
The addition of buoyancy decreases the potential energy of the water column.
It can be shown that the rate of decrease in potential energy ,if all the buoyancy
is concentrated at the surface is given by

where B is the rate of addition of buoyancy per unit horizontal area of the sea,
and h is the depth of the water column.
(We here choose our zero of potential energy as that of the water column after complete mixing to a uniform density distribution. This formulation neglects any work done by atmospheric pressure if the volume of the water column changes during the mixing process. This assumption is reasonable for small density changes, over which it may be assumed that the density varies proportionally with temperature and salinity).
If no mixing were applied to the water column the potential energy would decrease continually with a larger and larger density anomaly at the surface. However, the presence of mixing increases the potential energy of the water column and may prevent the formation of stratification. Mixing in the sea is accomplished almost exclusively by turbulence, which is generated by the presence of large-scale shearing flows, predominantly associated with tidal and wind-driven currents. Turbulence removes energy from these currents, and most of this energy finally appears as heat. However a small proportion (of order 1 percent) may be made available for removing any stable stratification, thereby increasing the potential energy of the system. The rate of abstraction of energy from a sheared flow is proportional to the shear stress in the fluid, multiplied by the velocity shear. If we consider the whole water column, then turbulent energy is introduced at the boundaries due to slip in layers associated with these boundaries (the “boundary layers”) - the upper one due to the surface wind stress, and the lower one due to the tidal current . Each boundary layer introduces turbulent energy equivalent to the boundary stress multiplied by the “slip velocity” across the layer. Boundary stresses in the sea are generally proportional to the square of the slip velocity
| τ = ρCDν2 | (3) |
where τ is the stress,
ρ is the water density,
CD is a dimensionless drag coefficient,
and ν is the slip velocity.
The rate of generation of turbulent energy is hence related to the cube of the slip velocity
Rate of generation = τ ν per unit horizontal area
=ρCDν3 per unit horizontal area
For tidal currents, “V” is approximately the tidal stream velocity, while for wind generated turbulence, it is the surface wind drift (roughly 3 percent of the wind speed).
Simpson and Hunter, 1974, assumed that the quantity of energy available for mixing was related to (4) by a constant of proportionality, ξ
Rate of supply of energy for mixing =ξρCDν3 (5)
The onset of stratification of a water column should hence be determined by a balance of (2) and (5) - i.e., there is just enough buoyancy input to overcome the possible gain in potential energy due to mixing
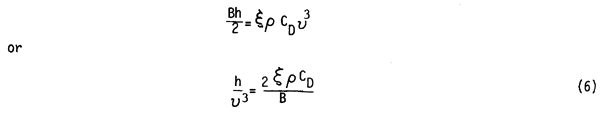
If, over a given sea area, the right hand side of (6) is constant, then the region where stratification can just occur (i.e. a “front”) should be associated with a constant value of “h/ν3”. This has been shown to be the case for tidal fronts on the North West European shelf (Simpson and hunter, 1974; Simpson, Hughes and Morris, 1977; Simpson, Allen and Morris, 1978; Pingree and Griffiths, 1978), in the Gulf of St. Lawrence (Pingree and Griffiths, 1980), in the Bering Sea (Schumacher, Kinder, Pashinski and Charnell, 1979), in the Bay of fundy and Gulf of Maine (Garrett, Keeley and Greenberg, 1978), and in the Cook Strait (Bowmna, Kibblewhite and Ash, 1980).
As the parameter “h/ν3” and vary widely, the logarithm of this quantity is generally used to derive a STRATIFICATION PARAMETER, several of which have been defined
(a) Ss = LOG 10(h/us3) |
(7) |
where us is the surface tidal current amplitude at Springs,
and h and us are in SI units.
(Simpson and Hunter, 1974, and other references associated with Simpson)
(b) Sp = LOG10 [h/Cd < |ut|3 >)] |
(8) |
where < ut3 > is the time average of the depth meaned tidal velocity cubed,
and h and ut are in GCS units.
(Pingree and Griffiths, 1978, 1980)
(c) SG1 = LOG10 [h/(ρCD < |ut|3 >)] |
(9) |
where h, ρ and ut are in SI units.
(SG2 = LOG10 (h/ν3) |
(10) |
where ν is the depth-meaned velocity amplitude for mean tides,
and h and ν are in SI units.
((9) and (10) due to Garrett, Keeley and Greenberg, 1978).
(d) SH = LOG10 (< |ut|3 >/h) |
(11) |
where h and ut are SI units.
(Hunter, 1982)
At a given place, these are related approximately by
| Ss = Sp + 0.57 = SG1 = SG2 - 0.45=-(SH + 0.82) | (12) |
For the waters of the North West Europen shelf, the position of a front is predictable approximately from
| Ss = 2 | (13) |
(Simpson, Hughes and Morris, 1977; Pingree and Griffiths, 1978)
![]()
![]()
![]()