![]()
![]()
![]()
In this report the tidal stratification parameter will be loosely referred to as “LOG10(h/u3)” where “u” is one of the definitions of tidal velocity given above.
A schematic representation of a front is given below
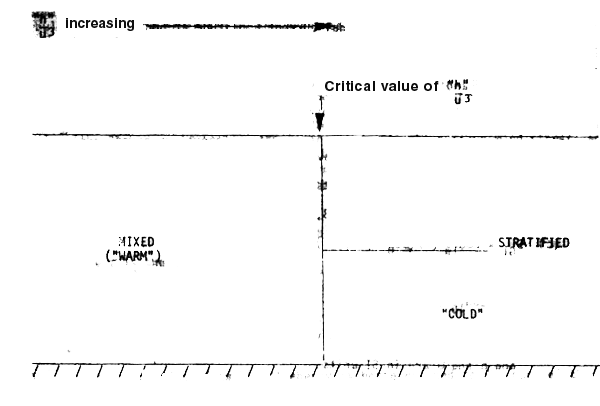
This simple picture of the position of a fron fixed in space may be modified by a number of factors.
(a) The mixing rate varies over the Spring-Neaps cycle (being a maximum at Springs), so we would expect the position of a fron to oscillate over this period, being displaced twoards the mixed water at Neaps and towards the stratified water at Springs
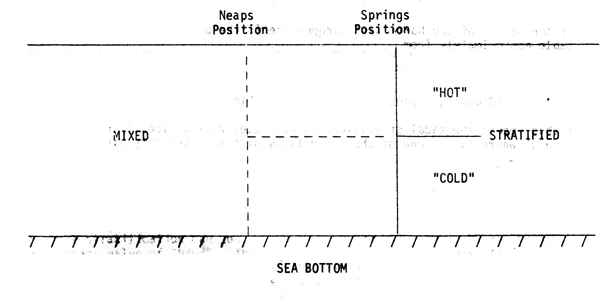
The expected movement of the front may be calculated from the variation of tidal velocity from Neaps to Springs(and any change in depth associated with the local bathymetry). However, observations have shown that the actual movement that can be attributed to this cause is considerably less than this (Simpson and Bowers, 1979) (the observed Springs/Neaps adjustment is typically 4 km). This discrepancy has apparently been resolved by modifying the simple model, and allowing the efficiency factor for mixing “ξ”, to vary with stratification (Simpson and Bowers, 1981) - it is known that vertical stratification inhibits mixing, and so it seems reasonable that “ξ” should decrease with stratification. Simpson and Bowers have shown that such a “variable efficiency” time-dependent model predicts considerably less frontal movement over the Neaps-Springs cycle. This modification to the model also leads to better agreement on frontal position over the summer season- the “constant efficiency” model predicts a marked advance of the front( towards the mixed water) in Spring, and a similar retreat in Autumn, not in accordance with observations, which show an almost constant frontal position from early June to late August (as predicted by the “variable efficiency” model). A further advantage of the variable efficiency model is that it predicts stron horizontal gradients in the degree of vertical stratification (i.e. a sharp front ) also in accord with observations.
(b) Frontal positions may also be modified by any water velocity normal to the front. There are several manifestations of this
(i) Tidal velocities, which advect the front back and forth. This is a major cause of the observed scatter of the positions of a given front in European waters - for a straight front, the displacement is simply given by the component of the tidal excursion normal to the line of the front.
(ii) Eddy-like features on the front. These have been observed in infra-red satellite images, and may draw their energy from the potential energy of the stratified fluid (baroclinic instability) or from the kinetic energy of the fluid (e.g., from tidal currents). These eddies may have scales of order tens of kilometres, but their dynamics are not well understood.
(iii) Steady flows. These may systematically distort the line of a front away from the position predicted by “h/u3” contours, as is probably the case with the front off Islay, to the North of the Irish Sea. In the case of an island, around which a fron would be predictble from “h/u3” values, a wake of mixed water may be formed in the downstream direction (e.g., the Scilly Isles) (See Simpson, 1981, for these examples).
Although the detailed position of a front may be changed from that predictable from a simple “h/u3” criterion, by the above effects, this basic rule still serves as a good indicator of the areas in which fronts would be expected to occur.
The requirements for the occurence of a front are mainly twofold
(a) There must be sufficient tidal (or wind) energy to form a mixed region, and conversely there must be adjacent regions where mixing is less and stratification can occur. As a rule of thumb, frontal structures seem to be found in areas of the world known for their high dissipation of tidal energy (e.g. the North West European Shelf, the Bay of Fundy and the Cook Strait).
(b) A condition for the formation of stratification is the supply of buoyancy to the fluid.This buoyancy is equivalent to a certain amount of negative potential energy that must be removed by supplying positive energy if vertical homogeneity is to occur. The relationship between buoyancy and potential energy is however dependent on where the buoyancy is put in - for instance, the potential energy of a column of water may be made more negative by the addition of positive buoyancy (light water) at the surface (the case we have considered up until now) or by the addition of negative buoyancy (heavy water) at the bottom. It is important to understand that the mixing processes described do not remove buoyancy - they simply increase the potential energy in a positive sense. If buoyancy (equivalent, for example, to heat or fresh water) is allowed to accumulate, then eventually some feedback process will“switch off” the buoyancy supply - for the case of heat, the sea will continually warm until surface losses balance the solar heating (as is the case for the daily- averaged conditions of a tropical lake). In such cases, any mixing processes will eventually mix the water vertically, removing any frontal structures originally present. There would appear to be two ways of getting round this problem
(i) If the buoyancy supply is periodic (e.g., seasonal), then during the buoyancy supply phase buoyancy can be stored in the water column, to be removed later during the buoyancy removal phase. This of course is what happens during the summer/winter cycle in non-tropical areas, and is the case for most of the examples of tidal fronts found in the liteature.
(ii) The buoyancy may be added at on epoint in the water column, and removed at another, thereby avoiding any accumulation of buoyancy. For instance, positive buoyancy could be added at the surface and removed at the bottom, thereby decreasing the potential energy of the water column through the formation of a light layer at the surface and a heavy layer at the bottom. This case coul be applied to tropical areas and only a simple modification (i.e., a positive and negative buoyancy supply) need be made to the basic model. Frontal positions would still be predictable from “h/u3” contours, so long as the buoyancy sources were reasonably constant horizontally. The mechanism for the removal of buoyancy from the bottom could be by lateral advection of mixing. A common example of this is an estuary, where positive buoyancy (fresh water) is added at the surface at the landward end and negative buoyancy (salty water) is added at the bottom at the seaward end. On average, no buoyancy accumulates in the estuary, and the negative potential energy input is balanced by tidal and wind mixing. In this example, the flows necessary to maintain the correct buoyancy balance are driven by the horizontal density gradients that arise from the buoyancy inputs - the system adjusts itself to yield a density structure that is roughly contant in time. In regions of an estuary where the mixing levels change in the horizontal, fronts may be formed. The above argument suggests that tropical areas, where the rate of heating does not vary much over the year and where the buoyancy input may be through heating, evaporation or the supply of fresh water, should not be discounted as regions for the possible formation of tidal fronts.
TIDAL MIXING IN SHELF SEAS
For a given rate of buoyancy input, the position of a front should be predictable from a fixed value of “h/u3”. If we consider the value for the North West European shelf, for stratification induced by solar heating, from equation (13) h/u32=100

Hence for a typical depth of water over a shelf of 100 metres, the frontal position should coincide with the 1 metre per second surface spring tidal velocity amplitude contour. As noted in section (2), this represents an area of high tidal dissipation, by global standards.
The tidal dissipation of shelf seas of the world was investigated by Miller (1966). He divided the shelf into 50 regions, and from computations of both the tidal energy crossing the boundaries with the ocean, and of the frictional dissipation at the sea bed, he obtained values of total dissipation for each region. These areas are shown in Figure 1 (from Miller's map), and unfortunately they are of very different sizes, and so do not give a good indication of the energy density available for mixing the water column.
Consider a shelf of length “L” (alongshore), width “W” (offshore) and depth “h”. Then from equation (4) the total dissipation of tidal energy over that shelf is given by
| E = WLρCD <|ut|3> | (15) |
where <|ut|3 > is the time average of the tidal velocity
(this may be a depth-meaned value, or a “bottom” value, so long as the drag coffecient “CD” is chosen appropriately.)
and ρ and CD are as defined for eqaution (3).
“E” is the parameter estimated by Miller, and it is related to “h/<|u|3”
by

Now, “ρ” and “CD” are approximately constant, and so we need to divide Miller's dissipation values (“E”) by the value of the shelf water “ hWL ”) to estimate the variation in “h/u3”. This would be a very tedious process, and so a further (rather gross) assumption is made that the depths (“h”) and widths (“W”) of all shelves are roughly the same. Miller's values have hence been divided by the shelf lenghts“L”) and used as estimators of the tidal dissipation levels in so far as they effect the “h/u3” parameter. Hence high levels of “E/L” indicate areas of low “n/u3” or areas where stratification may be removed by tidal mixing, giving rise to frontal formaton.
The computations were performed by measuring shelf lengths on Miller's map (similar to Figure 1), and applying the appropriate scale for that latitude to derive “L” .Some modifications of MILLER"s values was made using more recent data(Choi, 1980 (ryuku Islands0; flather, 1976 ( North West European Shelf0, Sunderman, 1977 (Bering Sea)): and data for the Cook Strait was added(Bowman, Kibblewhite and Ash, 1980). The results are shown in Figure 1, and TABLE 1. The values of “E/L” have been ranked in descending magnitude (the Okhotsk Sea has the largest value, and is ranked “o”). The numbers shown in the Figure 1, for each region, are the section number (as given in the table),'followed by the rank (in brackets).
There are several consistencies in Miller's paper, in which cases“best guesses” of the correct values have been made. Some sections of “zero” dissipation were present in Miller's map but not in his table - these have been denoted by “(-)” in Figure 1.
We know that “h/u3” frontal effects are just apparent in the Gulf of St. Lawrence, a relatively low dissipation area (pingree and Griffiths, 1980). We may hence regard this as roughly indicative of the lower limit of energy dissipation necessary for the pccirremce pf frpmta; strictires - hence all regions of rank less than about 23 may exhibit tidal fronts. Examination of Figure 1 indicates the following areas that fulfil this criterion:
(a) The North West European shelf(2), the gulf of St.Lawrence,(22), the Bering sea (20), the bay of Fundy and gulf of Maine(9) and the Cook Strait (4),where tidal fronts have been observed(rank shown in brackets).
(b) Most of the Pacific margin North of the Eqautor.
(c) The north and East margins of the Indian Ocean.
(d) Australia to he Lesser Sunda islands.
(e) The south-east and north-east coasts of South America.
(f) The near-Arctic and Arctic areas of the Hudson Strait, the Davis Strait and Norway to Svaldbard, all of which must fall into a special category.
It would appear rather ironic that of the shelf regions of the world that have received most attention form oceangraphers - the shelves of North West Europe, the U.S.A., Western South America and West Africa, only the former satisfies the above criterion.
An alternative way of looking at the importance of tidal dissipation is to compare the mixing effects of tidal currents and winds. It can be shown (Simpson and Bowers, 1981) that a tidal stream of surface amplitude“us” at Springs is equivalent to a root mean cubed wind of
| 9.49 us | (17) |
Hence for a typical wind of 5 metres per second, a tidal stream of surface Springs amplitude 0.53 metres per second exerts an equivalent mixing effect. Hence for a front to be dominated by tidal effects, e reqyire tidal currents of order 1 metre per second. This also agrees with the criterion derived form actual frontal values of the stratification parameter(equation 14).
THE BUOYANCY SUPPLY TO SHELF SEAS
(a) Surface Heating
The flux of heat through th surface of the sea is by no means determined simply by the rate of solar radiation and the meterological conditions, since th sea surface temperature plays an important role in the mechanisms involved in dissipating heat (back radiation, and “diffusive” and evaporative heat loss). An isolated lake, continually heated by the sun and atmosphere, will eventually reach an equilibrium temperature at which there is no total heat flux (expect for a very small geothermal heat flux upwards). If we know the sea surface temperature, then methods are available for estimating the various flux terms, in order to derive the total heat flux through the surface. An alternative approach is to look at the temperature history of a column of sea water and to thereby derive the total heat supplied to that column over a given period. This latter method takes account of the advection and diffusion of heat into the column from the surrounding sea water, but unfortunately does not tell us where in the water column the heat is introduced (as noted in Section 2, this must be known if we are able to computer the rate of input of potential energy to the system) and we are therefore forced forced to assume that the heat enters at the sea surface.
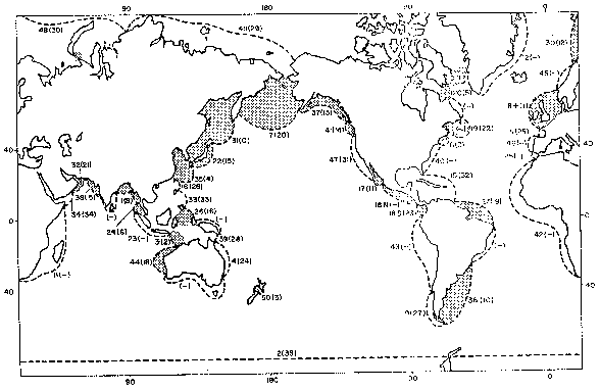
Fig. 1. Tidal dissipation of shelf areas (from Miller, 1966, and others).
Using estimates of the various heat flux terms at the sea surface, Hunter (1982) deduced a total flux of order 100 Joules per square metre per second for the northern part of the Arabian Gulf during the period of maximum heating. From estimates based on the temperature history of aa water column of the North West European Shelf, Simpson and Bowers, 1981, used a maximum heating rate of 100 Joules per square metre per second in their modelling works. Strokina, 1967, 1969, used this latter approach to produce seasonal maps of the heat flux to the upper layres of the world's oceans. While these are not really appropriate to the shelf areas, due to the strong dependence of the heat flux on sea surface temperature and local meterology, it is worth noting that for the southern hemisphere the positive heating flux is always less than about 160 Joules per square metre per second. It would hence appear that there i not a wide range in the values of maximum heating flux over the sea. For a factor of 2 change in buoyancy (or heat) flux, the critical stratification parameter (equations (7) to (12) changes by 0.3 For the North West European shelf, the stratification parameter changes by about l in 100 kilometres (Pingree and Griffiths, 1978), so a factor of 2 change in buoyancy input would be expected to move the fronts by only about 30 kilometres. Frontal position hence does no tdepend very strongly on the exact rate of supply of buoyancy.
In non-tropical areas, the heat flux to shelf seas generally shows a well defined annual periodicity. For the North West European shelf, it is positive from mid-March to mid-September (Simpson and Bowers, 1981), and significant stratification exists from April to November (Simpson, Hughes and Morris, 1977). However, the frontal positions are only approximately constant from early June to late August (Simpson, 1981). There would seem to be no reason why these timings should not apply approximately to the rest of the non-tropical northern hemisphere, and, with a shift of 6 months to the non-tropical southern hemisphere.
For tropical areas, where the seasoinal change in heating rate over shelf seas is small, it is necessary that there be some mechanisms for the removal of buoyancy if a steady stratification is to develop (as noted in Section 2). A possible system is shown schematicaly below
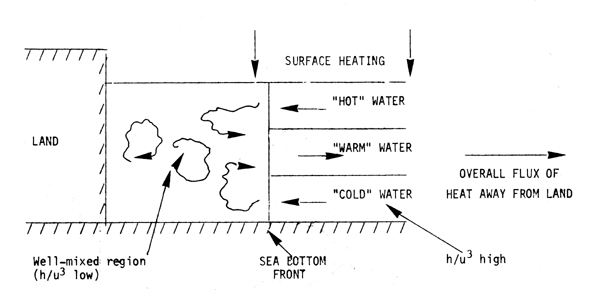
The horizontal density gradients induced would drive the circulation necessary to maintain a steady state. For larger systems, Coriolis accelerations would become important and the above two-dimensional circulation pattern would become three- dimensional, with a component of current in a longshore direction.
| Region | NO | Dissipation (ergs/s.×10-17) |
Dissipation Per Unit Length (ergs/cm./s./×10-9) |
Rank | *=Region of Potential Tidal Effects |
|---|---|---|---|---|---|
| Andaman Sea to 21° N | 1 | 6.5 | 5.5 | 8 | * |
| Antartica | 2 | 0.1 | 0.006 | 35 | |
| Australia to Lesser | |||||
Sunda Islands |
3 | 15 | 12.4 | 2 | * |
| Barrier Reef | 4 | 2.4 | 0.96 | 24 | |
| Bay of Biscay | 5 | 0.4 | 0.76 | 25 | |
Gulf of Maine |
6 | 2.3 | 4.9 | 9 | * |
| Bering Sea | 7 | 2.9 | 1.4 | 20 | * |
| N.W.European shelf | 8+ | 21.48 | 14.2 | 1 | * |
| Chile | 9 | 0.4 | 0.28 | 27 | |
| Davis Strait etc. | 10 | 2 | 1.9 | 17 | |
| East Africa | 11 | 0 | 0 | - | |
| Eastern Greenland | 12 | 0 | 0 | - | |
| Florida to Trinidad | 15 | 0.3 | 0.11 | 32 | |
| Formosa to Luzon | 16 | 0.5 | 0.65 | 26 | |
| Gulf of California | 17 | 4 | 3.2 | 11 | * |
| Gulf of Panama | 18N | 0 | 0 | - | |
| Gulf of Panama | 18S | 0.6 | 0.99 | 23 | * |
| Gulf of St. Lawrence | 19 | 0.8 | 1.0 | 22 | * |
| Hudson Strait | 20 | 12 | 10.7 | 5 | * |
| Japan Islands | 22 | 4.3 | 2.3 | 15 | * |
| Java to Sumatra | 23 | 0 | 0 | - | |
| Malacca Strait | 24 | 7 | 7.0 | 6 | * |
| Mediterranean | 25 | 0 | 0 | - | |
| Mindanao to N. Guinea | 26 | 2 | 2.0 | 16 | * |
| N.E. coast of S. America | 27 | 5 | 1.6 | 19 | * |
| North Bay of Bengal | 29 | 6 | 6.2 | 7 | * |
| Norway to Svaldbard | 30 | 3.2 | 3.0 | 12 | * |
| Okhotsk Sea | 31 | 21 | 14.6 | 0 | * |
| Oman-Persian Gulf | 32 | 1.6 | 1.2 | 21 | * |
| Philippies | 33 | 0.1 | 0.083 | 33 | |
| Red Sea | 34 | 0.2 | 0.080 | 34 | |
| Ryukyu Islands | 35 | 11.96 | 11.3 | 4 | * |
| S.E. coast of S. America | 36 | 13 | 3.9 | 10 | * |
| Southern Alsaka | 37 | 5 | 2.5 | 13 | * |
| Southern India | 38 | 4 | 2.3 | 15 | * |
| Torres Strait | 39 | 0.3 | 0.25 | 28 | |
| U.S. east coast | 40 | 0 | 0 | - | |
| Vancouver, Juan de Fuca | 41 | 1.6 | 2.4 | 14 | * |
| West Africa | 42 | 0 | 0 | ||
| West S. America | |||||
Equator to 42° S |
43 | 0 | 0 | ||
| Western Australia | 44 | 4.2 | 1.8 | 18 | * |
| Western Norway | 45 | 0 | 0 | ||
| Western Spain | 46 | 0 | 0 | ||
| Western U.S to Baja, CA | 47 | 0.4 | 0.13 | 31 | |
| Svaldbard to Taimyr | 48 | 0.3 | 0.16 | 30 | |
| Taimyr to Pt. Barrow | 49 | 0.6 | 0.19 | 29 | |
| Cook Strait | 50 | 2.73 | 11.4 | 3 | * |
(b) Supply or Removal of Fresh Water
In buoyancy terms, the addition of 1 joule of heat is roughly equivalent to 3×10-9 cubic metres of fresh water. Hence a surface heat input of 100 Joules per square metre per second has the same buoyancy effect as the addition of 3×10-9 cubic metres per square metre per second of fresh water. This is the equivalent of a river the size of the Mersey, England ,discharging 50 cubic metres per second over an area of 170 square kilometres(e.g., a square of side 13 kilometres), or a rainfall equivalent ot 3×10-7 metres per second or about 10 metres per year ( a large figure by global standards). Hence a river may input a significant amount of buoyancy(compared with surface heating) in the region around its mouth 9within a few tns of kilometres), but only over short periods could rainfall over the sea input a comparable amount of buoyancy.
During evaporation of water from the sea surface( which is especially important in tropical areas), latent heat is absorbed at a rate of about 2×109 joules per cubic metre of water. This cubic meter of water is equivalent to about 3×108 Joules of heat supply in buoyancy terms. Hence from the point of view of the buoyancy suply, the effect of evaporation is mainly to absorb latent heat, thereby cooling the water and adding the negative buoyancy(at a rate of 2×109 Joules per cubic metre). The loss of fresh water is only equivalent to about 15 per cent of this buoyancy flux (the equivalent of 3×108 Joules per cubic metre).
Over most of the world's oceans, the evaporation rate roughly cancels the precipitation rate(of order 1 metr per year) 9Budyko, 19740, so the previous comments concerning the general insignificance of rainfall apply also to evaporation. However, in tropical areas, evaporation may exceed precipitation and become a significant source of negative buoyancy - for example, the average evaoration rate over the Arabian Gulf of order 1.4 metres per year(Privett, 1959), compared with a rate of precipitation of 1.4 metres per year( Hydrographer of the Navy, 1967), the difference being equivalent in buoyancy terms to a heat loss of about 13 joules per square metre per second. Even though this evaporation rate is not particularly high, it is more likely than a rainfall to produce ocal areas of mixed and stratified waters for two reasons:
(i) Rainfall is independent of sea surface temperature and so does not have well-defined localized distributions over the sea. Evaporation, on the other hand is often high in shallow coastal regions due to lower relative humidity and (in the summer) to higher sea surface temperatures near the caost.
(ii) As noted previously, associated with evaporation is an absorption of latent heat equivalent, in buoyancy terms, to about 7 times the fresh water loss.
The dominant buoyancy sources for shelf seas are hence surfcae heating, river runoff and evaporation. It has been shown above the there are circumstances by which any of these can provide sufficient buoyancy for the formation of frontal structures.
CONCLUSIONS
Tidal shelf sea fronts are associated with regions of relatively high dissipation, areas which are indicated by rank in Figure 1.
Stratification may be induced by surface heating, river runoff or evaporation, and the majority of known tidal frontal structures are associated with seasonal changes of surface heating in non-tropical areas. However, we should not discount trropical areas as to possible sites for tidal fronts, generated by any of the three buoyancy sources, since in the estuaries of the world we have good examples of quasi0steady density stratification being maintained by the interactionof positive buoyancy (fresh water) and negative buoyancy (“ocean” water) under the control of tidal and wind-driven mixing.
For buoyancy input by seasonal heating in the non-tropical nothern hemisphere, stratification probably exists from April to November, with stable frontal positions from early June to late August. In the southern hemisphere one would expect these times to be shifted by 6 months.
The characteristic dimensions of fronts and eddies are quite diverse, ranging from short scales of meters to kilometers, corresponding to scales of relevance to larval fish, fish populations, and the aggregation of fishing activities based on “local” resource aggregations. Our knowledge of these events, processes and the dependence of fishery activities related to them needs to be expanded, certainly through more varied scale studies on the processes promoting them in order to better locate them in time and space. It follows that detailed studies of the biological milieu associated with the various features will yield insights into important processes, particularly related to fish reproduction, productivity and energy transfer, and fishery effectiveness measures.
REFERENCES
Allen, J.S. and R.L. Smith. 1981. On the dynamics of wind-driven shelf currents. Phil.Trans.R.Soc. Lond. A302:617–634.
Alvarino, A. 1980. The relation between the distribution of zooplankton predators and anchovy larvae. CALCOFI Reports XXI. (Also ICES, Early Life History Meeting, Woods Hole, April 1979, poster no.21).
Anderson, K.P. and E.Ursin. 1977. A multispecies extension of the Beverton and Holt theory of fishing, with accounts of phosphorus circulation and primary production. Meddr.Damm.Fisk-og Havunders, N.S. 7:319–435.
Anon. 1981. Circulation and fronts in Continental Shelf seas (conference proceedings). Phil.Trans.R. Soc.Lond. A302:515–689.
Bakun, A., J. Beyer, D. Pauly, J.G. Pope and G.D. Sharp. 1982. Ocean Sciences in relation to living resources. Can.J.Fish.Aquat.Sci. 39(7):1059–1070.
Bakun, A. and R.H. Parrish. 1981. Environmental inputs to fishery population models for eastern boundary current regions. pp. 67–104. In Workshop on the effects of environmental variation on the survivl of larval pelagic fisher (G.D. Sharp, ed.). IOC Workshop Rep. 28. IOC/UNESCO, Paris.
Baronov, F.I. 1918. On the questionof the biologicl baiss of fisheries. Nauch.issledov.iktiol, Inst.Izv.I. (1):81–128, Izv.ryb.i. nauch.isseldov. 1(1) Moscow.
Beardall, J., G.E. Fogg, P. Foster, I. Miller, C.P. Spencer and D. Voltolina. 1978. Biol-Contemp. 5:163–175.
Beverton, R.J.H. and S.J.Holt. 1957. On the dynamics of the exploited fish populations. Fish.Invest., Lond. Ser.2., 19:533 p.
Beyer, J. and P.Sparre. 1983. Modelling exploited fish stocks. In Application of Ecological Modelling in Environmental Management (S.E. Jorgensen, ed.). Elsevr Scientific Publishing Company, Amsterdam.
Bowman, M.J., A.C. Kibblewhite and D.E. Ash. 1980. M2 tidal effects in Greater Cook Strait, New Zealand, J.Geophys.Res. 85(C5):2778–2742.
Budyko, M.I. and D.H. Miller (eds). 1974. Climate and Life. Academic Press. New York. 508 p.
Chapman, W.M. 1972. The application of oceanography to the development and management of ocean fisheries. pp.13–21. In Progress in Fishery Science. Fifteenth Anniversary Celebration Symposium (R.W. Moore, ed.). University of Washington Press. Univ. of Washington Publications in Fisheries, New Series, Vol.5.
Choi, B.H. 1980. A tidal model of the Yellow Sea and the Eastern China Sea, Korean Ocean Research and Development Institute. Report 80–02.
Cushing, D.H. 1971. The dependence of recruitment on parent stock in different groups of fishes. J.Cons.int.Explor.Mer. 33:340–362.
Cushing, D.H., 1972. The production cycle and numbers of marine fish. Symp.Zool.Sco., London. 29:213–232.
Dooley, H.D. 1978. The role of axially varying vertical mixing along the path of a current in generating phytoplankton production. Phil.Trans.R.Soc.Lond. A302:649–661.
Fasham, M.J.R. 1978. The statistical and mathematical analysis of plankton patchiness. Oceanogr. Mar.Biol.Ann.Rev, 16:43–79.
Fearnhead, P.G. 1975. On the formation of fronts by tidal mixing around the British Isles, Deep-Sea Res. 22:311–321.
Flather, R.A. 1976. A tidal model of the Northwest European continental shelf. Mem.Soc.R.Sci.Liege, 6ee serie, X:141–164.
Foster, P., G. Savidge, G.M. Foster, D.T.E. Hunt and L.B. Pugh. 1976. Multivariate analysis of surface water characteristics in the summer regime of the Western Irish Sea. J.Exp.Mar.Biol.Ecol. 25(2):171–185.
Garcia, S. (in press). The stock-recruitemetn relationship in shrimps: reality or artefacts and misinterpretation. Oceanogr.Trop.
Garrett, C.J.R., J.R. Keeley and D.A. Greenberg. 1978. Tidal mixing versus thermal stratification in the Bay of Fundy and Gulf of Maine. Atmosphere-Ocean. 16(4):403–423.
Gieske, W.W.C., G.W. Kraay and M.A. Baars. 1979. Current 14C methods for measuring primary production: gross understimates in oceanic waters. Netherlands J. of Sea.Res. 13(1):58–78.
Glantz, M.H. and J.D. Thompson (eds.) 1981. Resource Management and Environmental Uncertainty, Lessons from coastal upwelling fisheries. John Wiley and Sons, New York, Chichester, Brisbane, Troronto. 491 p.
Graham, M. 1935. Modern theory of exploiting a fishery, and applications to North Sea trawling. J.Cons.int.Explor.Mer. 10:264–274.
Grainger, R.S.R. 1980. Herring abundance off the west of Ireland in relation to oceanographic variation. J.Cons.int.Explor.Mer. 38(2):180–188.
Grall, J.R., P. Le Corre, J. Le Fevre, Y. Marty and B. Fournier. 1980. Oceans. 6:235–274.
Gregg, M.C. 1980 Microstructure patches in the thermocline. J.Phys.Ocean. 10:915–943.
Harding, D., J.H. Nichlos and A.R. Folkard. 1978. Studies on planktonic fish eggs and larvae in relation to environmental conditions in the west central North Sea. Annls.Biol. 33:62–69.
Hardy, A.C. 1924. The herring in relation to its animate environment, Part 1. The food and feeding habits of herring. Fish.Innvest.Lond., Ser. II, 7, 53 p.
Hennemuth, R.C., J.E. Palmer, B.E. Brown. 1981. A statistical description of recruitment in eighteen selected fish stocks. J.Northw.Atl.fish.Sci. 1:101–111.
Hiort, j. 1914. Fluctuations in the year classes of important food fishes. lJ.du Conseil. (1):5–38.
holligan, P.M. 1981. Biological implications of fronts on the Northwest European continental shelf. Phil. trans.R.Soc,Lond. A302:547–562.
Houde, E.D. 1974. Effects of temperature and delayed feeding on growth and survival of larvae of three species of subtropical marime fishes. Mar.Biol. 26:271–285.
Houde, E.D. 1975. Effects of stocking density and food density on survival, growth and yield of laboratory-reared larvae of sea bream Archosargus rhomboidalis (L) (Sparidae). J.Fish.Biol. 7:115–127.
Houde, E.D. 1978. Critical food concentrations for larvae of three species of subtropical marine fishes. Bull-Mar.Sci. 28:395–411.
Houde, E.D. and R.C. Schekter. 1981. Functional and developmental responses of fish larvae to varying prey concentrations. Symposium on Early Life History of Fish, Woods Hole, Massachusetts, April 1979. Repp.P.-v.Réun.Cons.int.Explor.Mer. 178.
Hunter, J. 1982. Tidal and stratification/mixing models of Kuwait waters, Kuwati Bulletin of Marine Science (in press).
Hunter, J.R. 1982. The feeding ecology of marine fish larvae. In J.E. Bardach (ed.), The Physiological and Behavioural Manipulation of Food Fish as Production and Management Tools. International Center for Living Aquatic Resources Management, Manila.
Hydrographer of the Navy. 1967. Persian Gulf Pilot. 11th Edition. NP63, London.
Illes, T.D. and m.Sinclair. 1982. Atlantic herring: stock discretensess and abundance. Science. 215(4533):627–633.
James, I.D. 1981. Fronts and shelf-ciculation models. Phil.Trans.R.Soc.Lond. A302:597–604.
Jones, R. and W.B. Howell. 1974. Some observations on the population dynamics of the larval stage in the common gadoids. pp. 87–102. In the Early Life History of Fish (J.H.S. Blaxter, ed.). Springer-Verlag, New York.
Laevastu, T. and H.A.Larkins. 1981. Marine fisheries ecosystem: its quantitative evaluation and management. Fishing News Books Ltd., Farnham. 159 p.
Lasker, R. 1975. Field criteria for survival of anchovy larvae: the relation between inshore cholorophyll maximum layers and successful first feeding. Fish.Bull.U.S. 73:453–462.
Lasker, R. 1978. Ocean variability and its biological effects-regional review-northeast Pacific. Rapp.P.-v.Réun.Cons.int.Explor.Mer. 173:168–181.
Laurec, A. and J.-C. Le Guen. 1981. Dynamique des populations Marines Exploites: tome 1, concepts et modeles. Pub.du Centre National pour L'Exploitation des Oceans, CNEXO, Brest. 115 pp.
Laurence, G.C. and J.E. Beyer. 1981. A stochastic dynamic model of growth and mortality in winter flounder larve (Pseudopleuronectes americanus) reared in the laboratory under experimental conditions. Symposium on Early Life History of Fish, Woods Hole, Massachusetts, April 1979 Rapp.P.-v.Réun.Cons.int.Explor.Mer. 178.
MacCall, A.D. 1981. The consequence of cannibalism in the stock-recruitment relationship of planktivorous pelagic fishes such as Bngraulis. p. 201–217. IOC Workshop Report No. 28. (G.D. Sharp, ed.). IOC/UNESCO, Paris.
Mann, K.H. 1982. Ecology of coastal waters: a systems approach. Studies in Ecology, Volume 8 Blackwell Scientific Publications. Oxford, London, Edinburgh, Boston, Melbourne. 322 pp.
Miller, G.R. 1966. The flux of tidal energy out of the deep oceans. J.Geophys.Res. 71(10):2485– 2489.
Nihoul, J.C.J. (ed.). 1982. Hydrodynamics of semi-enclosed seas. Elsevier Scientific Publishing Company, Amsterdam, Oxford, New York. 555 p.
Owen, R.w. 1980. Eddies of the California current system: physical and ecological characteristics. In the california Islands (D.M. Power, ed.). Santa Barbara Mus.Nat.History (Calif.).
Owen, R.w. 1981. Patterning of flow and organisms in the larval anchovy enviornment. pp. 176– 200. In IOC Workshop Report No. 28. (G.D. Sharp, ed). UNESCO, Paris.
Owen, R.w. 1982. Microscale plankton patchiness in the larval anchovy enviornment. Rapp.P.- v.Réun.Cons.int.Explor.Mer. 178:364–368.
Parrish, R.H., C.S. nelson and A. Bakun, 1981. Transport mechanisms and reproductive success of fishes in the California Current. Biol.Oceanogr. 1(2):175–203.
Pauly, D. (Draft). Fish Production Dynamics in Tropical Waters: International Center for Living Aquatic Resources Management, Manila.
Petersen, C.G.J. 1984. On the biology of our flatfishes and on the decrease of our flatfish fisheries. Rep.Dan.biol.Stn. IV. 147 pp.
Pingree, R.D. and D.K. Griffiths. 1977. The bottom mixed layer on the continental shelf. Estuar.Coast.Mar.Sci. 5:399–413.
Pingree, R.D. and D.K. Griffiths. 1978. tidal fronts on the shelf seas arount the British Isles. J.Geophys.Res. 83(C9):4615–4622.
Pingree, R.D. and D.K. Griffiths. 1980. A numerical model of the M2 tide in the Gulf of St. Lawrence, Oceanologica Acta. 3(2):221–225.
Pingree, R.D., P.M. Holligan and G.T. Mardell. 1978. The effects of vertical stability on phytoplankton distributions in the summer on the northwest European continental shelf. Deep Sea Res. 25:1011–1028.
Pingree, R.D., P.M. Holligan and G.T. Mardell. 1979. Phytoplankton growth abd cyclonic eddies. Nature. 278:245–247.
Pingree, R.D., p.R. Pugh, P.M. Hollian and G.R. froster. 1975. Nature. 258:672–677.
Privett, D.W. 1959. Monthly charts of evaporation from the N. Indian Ocean (including the Red Sea and the Persain gulf). quart.J.Roy.Met.Soc.(Lond.). 85:424–428.
Reid, J.L., E. Brinton, A. Fleminger, E.L. Vernick and J.A. McGowan. 1978. Ocean circulation and marine life. pp. 65–130. In Advances in Oceanography. Plenum Press, New York.
Richards, F.A. (ed.0. 1981. Coastal Upwelling. American Geophysical Union, Washington, D.C. 529 pp.
Ricker, W.E. 1954. Stock and recruitment. J.Fish.Res.Bd.Canada. 11:559–623.
Ricker, W.E. 1958. handbook of computations for biological statitics of fish populations. Bull.Fish.Res.Bd.Canada. No. 119.
Rosenthal, H. 1970. Anfutterung und Wachstum der Larven und Jungfische des Hornhechts Belone belone. Helgolaender Wiss.Meeresuntersuch. 21:320–332.
Saetersdas, G. 1980. A review of past management of some pelagic stocks and its effectiveness. Rapp.P.-v.Réun.Cons.int.Explor.Mer. 177:505–512.
Savidge, G. 1976. A preliminary study of the distribution of chlorophyll A in the vicinity of fronts in the Celtic and Western Irish Seas. Estuar.Coast.Mar.Sci. 4:617–625.
Savidge, G. and P. Foster. 1978. Plankton biology of a front in the Celtic Sea. Nature, London. 271:155–156.
Schumacher, J.D., T.H. Kinder, D.J. Pashinski and R.L. Charnell. 1979. A structural front over the Continental Shelf of the Eastern Bering Sea. J.Phys.COceanogr. 9:79–87.
Sharp, G.D. 1981a. Report of the Workshop on the Effects of Environmental Variation on Survival of Larval Pelagic Fishes. pp. 1–47. In Intergovernmental Workshop Series, No. 28 (G.D. Sharp, ed.). UNESCO, Paris.
Sharp, G.D. 1981b. Colonization: modes of opportunism in the ocean. pp. 125–148. In Workshop on the Effects of Environmental Variation on the Survival of Larval Pelagic Fishes. 20 April-5 May 1980. Lima, Peru. (G.D. Sharp. ed.). IOC Workshop Report series No. 28. IOC/UNESCO, Paris.
Sharp, G.D. 1981c. Report of the expert consultation on the research status and potential of cod rearing in the North Atlantic. Organized in collaboration with the Svanøy Foundation, Svanøy, Norway, 2–9 August 1981. 12 pp.
Sharp, G.D. (ed.). 1981d. Intergovenmental Oceanographic Commission Workshop Report No. 28. Workshop on the Effects of Environmental Variation on the Survival of Larval Pelagic Fishes. 20 April-5 May 1980. Lima, Peru. UNESCO, Paris. 323 p.
Shepherd, J.G., J.G. Pope and R.D. Cousens. 1982. Variations in fish stocks and hypotheses concerning their links with climate. Mini Symposium ICES CM 1982/GEN:6. 22 pp. plus tables and figures.
Simpson, J.H. 1981. The shelf-sea fronts: implications of their existence and behaviour. Phil.Trans.R.Soc.Lond. A:302:531–542.
Simpson, J.H. and D. Bowers. 1979. Shelf-sea fronts' adjustments revealed by satellite IR imagery. Nature. 280(5724):648–651.
Simpson, J.H. and D. Bowers. 1981. Models of stratification and frontal movement in shelf seas. Deep-Sea Res. 28A(7):727–738.
Simpson, J.H. and J.R. Hunter. 1974. Fronts in the Irish Sea. Nature. 250:404–406.
Simpson, J.H., C.M. Allen and N.C.G. Morris. 1978. Fronts on the Continental Shelf. J.Geophys.Res. 83(C9):4607–4614.
Simpson, J.H., D.G. Hughes and N.C.G. Morris. 1977. The relation of seasonal stratification to tidal mixing on the Continental Shelf. A Voyage of Descovery. Pergamon, pp. 327–340 (Ed. M. Angle).
Simpson, J.H., D.J. Edelston, A. Edwards, N.C.G. Morris and P.B. Tett. 1979. The Islay front: physical structure and phytoplankton distribution, estuaine and coastal. Mar.Sci. 9:713–726.
Smith, P.E. and R. Lasker. 1978. Position of larval fish in an ecosystem. Rapp.P.-v.Réun..Cons.int. Explor.Mer. 173:77–84.
Steele, J.H. 1974. The structure of marine ecosystems. Blackwell Scientifc Publications. Oxford, London, Edinburgh, Melbourne. 128 pp.
Strokina, L.A. 1967. Determination of variation in the heat content of the oceans. Tr. Gl. Geofiz. Observ. 209:58–69.
Strokina, L.A. 1969. Variation in the heat content of the oceans of the Southern Hemisphere Tr.Gl.Geofiz.Observ. 249:58–64.
Sunderman, J. 1977. The semi-diurnal principal lunar tide M2 in the Bering Sea, Deutsche Hydrograph. Zeitschr. J. 30(3)93–93–101.
Sverdup, H.O. 1952. Some remarks on the place of hydrography in fisheries research. Rapp. P- v. Réun. Cons.int.Explor.Mer. CXXXI.p.7.
Sverdup, H.O. 1955. The place of physical oceanography in oceanographic research. J. Mar Res. 14(4):287–294.
Szekielda, K.-H., D.J. Suszkowski and P.S. Tabor. 1977. Skylab investigation of the upwelling off the northwest coast of Africa. J. Cons. int. Explor. Mer. 37(3):205–213.
Theilacker, G. and K. T. Dorsey. 1981. Larval fish diversity, a summary of laboratory and field research. p. 105–142. In IOC Workshop Report No. 28 (G.D. Sharp, ed.). IOC/UNESCO, Paris.
Thomasson, E. M. 1981. Study of the Sea: the development of marine research under the auspices of the International Council for the Exploration of the Sea. Fishing News Books Ltd., Farnham. 256 p.
Voronina, N. M. 1978. Variability of ecosystems. pp. 221–244. In Advances in Oceanography (H. Charnok and G. Deacon, eds). Plenum Press, New York.
Por
Patricio A. Bernal*, Fernando L. Robles*(1) y Omar Rojas**
*BIOTECMAR
Pontificia Universidad Catolica de Chile
Sede Regional Talcahuano
Casilla 127 - Talcahuano, Chile
**Instituto de Fomento Pesquero-Chile
Casilla 1287
Santiago, Chile
(1) Direccion actual:
Comision Oceanografica Intergubernamental
Unesco. Place de Fontenoy 75700
Paris, Francia
Summary
In this study time series of physical oceanographic data (salinity, temperature and geostrophic flow) and biological data (zooplankton biomass, eggs and larvae of Engraulis ringens and Sardinops sagax) were examined for the ten year period of the 1970's for the southern region of the Chile-Peru Current system.
The principal water masses in the region are characterized and the variations in these experienced in response to the dominant oceanographic regions are also described. Special emphasis is given to the classification of the oceanographic conditions during the “El Niño” and “Anti-El Nino” periods, and their genesis is discussed in relation to the interactions of large scale ocean-atmosphere phenomena.
Estimations of seasonal variability and long-term residual biological variables are given, and the seasonal cycles are characterized. The seasonal zooplankton biomass component represents 50% of the total, in contrast with the California Current where this variable is about 18% on average. On the other hand, for the long-term analysis the residual variance of the eggs and larvae of E. ringens and S. sagax attain 70%. The inter-annual changes of the biological variables form no consistent pattern during sequential cold and warm periods. However, a reverse relation between spawning intensity is observed between E. rigners and S. sagax during warm periods. The extension of the distribution of S. sagax is documented for the period following the collapse of the anchoveta fishery after the “El Niño” of 1972.
INTRODUCCION
La presentación a esta Consulta de Expertos de una revisión sobre la variabilidad oceanografica del Sistema de Corrientes de Chile-Perú, requiere de una explicacion preliminar. En primar lugar, la mayor variabilidad de los stocks de peces pelagioc sque habitan los ecosistemas altamente productivos de los mergenes orientales de los oceanos, aparte de factores poblacionales intrinsecos, pareciera estar tambien asociada a la variabilidad oceanografica de gran escala observada en dichos sistemas. En segunde lugar, existe una abundate evidencia acumulada que demuestra el impacto que tienen cierto tipo de eventos oceanograficos semi-periodicos sobre las poblaciones de peces pelagicos en explotacio, por ejemplo las alteraciones sobre el reclutamiento de la anchoveta peruana, Engraulis ringens, causadas por el fenomenos de “El Nino”. La prediccion de estos eventos es un objetivo significativo en si mismo paral el manejo pesquero en la Region.
En tercer lugar, y quizás la razán de mayor importancia en el contexto de esta reunión, es el notable incremento de capacidad predictive que se adquiere respecto a las variaciones de los stocks de peces pelágicos, al incorporar estimaciones parametrizadas de la variabilidad oceanográfica en moelos pesqueros. Un ejemplo de esto último lo encontramos en el trabajo de Parrish y MacCall (1978), quienes analizaron la información contenida en series históricas de la captura de la caballa de California (Scomber japonicus). Estos autores, utilizando una variedad de relaciones stock-recluta tomadas de la literature, demostraron que estas, en el major de los casos, explicaban un 24% de la varianza total de la serie histórica de capturas para el periodo 1931-1968. En cambio, mediante la incorporacion de tárminus densidad-independientes modulados por variables oceanográficas, Parrish y MacCall mostraron que el nivel de varianza explicada por estos modelos mixtos llegaba al 75% de la varianza total. Las variables oceanográficas incorporadas son del tipo altura del nivel del mar, un índice de surgencias costeras, un índice de la divergencia oceánica de la capa superficial y la temperatura superficial del mar, variables todas ellas indi- cativas de procesos oceanográficos dominantes en los sistemas de márgen oriental.
En el presente trabajo, se presenta una caracterización física y boilógica de la Región Meridional del Sistema de Corrientes de Chile - Perú, se discute la variabilidad física de baja frecuencia en torno a el fenámeno de El Niño, y finalmente se analiza la variabilidad de baja frecuencia de series históricas de biomasa de zooplankton y de huevos y larvas de anchoveta, E. ringens, y sardina, sandrinaps sagarx.
Fuentes de Información
La información oceanográfica que se presenta y discute en este trabajo proviene de datos obtenidos en 80 cruceros realizados en la Region durante el periodo 1931 a 1973. Una listado exhaustivo de las fuentes se encuentra en Robles (1979). Una importante fracción de la información fue recolectada como resultado del esfuerzo institucional desarrollado por el Instituto de Fomento Pesquero de Chile (IFOP) en la decada 1964–1973. El banco de datos oceanográfico-pesqueros de IFOP, que incluye hasta 1980, 207 cruceros de investigación, 120 de los cuales de caracter oceanografico, ha sido descrito por Canón (1980), quien entrega la localizacion de las estaciones, estado de procesamiento y disponibilidad de la información.
La información biólogica sobre huevos larvas de peces y biomasa plantónica, provience en su totalidad de los archivos del laboratorio de Ictioplancton de IFOP, donde se llevó a cabo la separación, identificacion taxonómica, clasificación por estadios de desarrollo y cuantificación del material. El procesamiento automatico de dicha información para el período 1964-73 y su posterior analisis se realizó en el Departmento Biotecmar de la Pontificia Universidad Católica de Chile, Sede Regional Talcahuano.
La Oceanografía fisica del Pacifico sureste Medidional: caracterización y condiciones medias
En el Océano Pacífico al sur del Ecuador y con centro en las latitudes intermedias. se ubica el giro anticiclónico del Pacifico Sureste. El brazo oriental de este giro, frente a las costas de Sudamérica, lo forma el Sistema de Corrientes de Chile - Perú. La circulación oceánica y las masas de aguas presentes en la Región del Pacifico Sureste, están determinadas principalmente por la presencia de las aguas frías y de baja salinidad que fluyen de sur a norte como parte de este Sistema de Corrientes.
| Masas de agues presentes | Sigma - t | Profundidad m. |
Temperatura °C |
Salinidad %° |
Oxigeno ml/l |
|---|---|---|---|---|---|
| A)Area 1 : 75°-90°w; 18°-35°S | |||||
| Subtropical superficial (AST) | <25.30 | <35 | >18.5 | >34.8 | >5.0 |
| Subantártica (ASA) | 25.70–25.90 | 70–130 | 12.5–13.5/ | 34.2–34.4 | 4.1–5.6 |
| Ecuatorial subsuperficial (AESS) | 26.80–26.90 | 320–370 | 8.5 | 34.4–34.5 | 0.9–2.2 |
| Intermedia antártica (AIA) | 27.10–27.20 | 540–640 | 5.5–6.5 | 34.4–34.4 | 2.4–4.1 |
| B) Area 2: OOSTA-75°; 18°–28°S | |||||
| Subtropical superficla (AST) | <25.30 | <12 | >18.5 | >34.9 | >5.0 |
| Subantártica (ASA) | 25.90 | 25–40 | 14.5 | 34.7–34.9 | 0.6–1.0 |
| Arica-Mejillones | 26.40 | 100–110 | 12.5 | 34.7–34.9 | 0.6–1.0 |
| Ecuatorial subsuperficial (MESS) | 26.80 | 330–370 | 9.5–10.5 | 34.6–34.8 | 0.3–0.6 |
| Intermedia antártica (AIA) | 27.30 | 710–750 | 5.5 | 34.5 | 1.5–1.9 |
| C) Area 3: COSTA-75°W; 28°–39°S | |||||
| “Fiordos” | 24.70–25.60 | 10–17 | 13.5–17.5 | 33.9–34.2 | 5.3–6.0 |
| Subantártica (ASA) | 25.70–26.00 | 20–50 | 11.5–13.5 | 34.1–34.2 | 4.0–3.7 |
| Ecuatorial subsuperficial (ACSS) | 26.80–26.90 | 300–340 | 8.5 | 34.5–34.6 | 0.8–1.3 |
| Intermedia antártica (AIA) | 27.10–27.20 | 560–600 | 5.5 | 34.3–34.4 | 3.8–4.1 |
| D) Area 4: COSTA–77°W; 39°–48°S | |||||
| “Fiordos” | 24.60–25.10 | 0–40 | 11.5–13.5 | 32.7–33.4 | 5.5–6.6 |
| O2 minimo | 26.60–26.70 | 152–210 | 8.5–9.5 | 34.1–34.4 | 2.0–3.0 |
| Ecuatorial subsuperficial (AESS) | 26.90–27.00 | 290–390 | 6.5–7.5 | 34.3–34.4 | 2.5–4.3 |
| Intermedia Antártica (1) | 27.10 | 500–530 | 5.5 | 34.3 | 5.0–5.2 |
| (1) Sin datos en Invierno. | |||||
| E) Area 5(1) 76°–90°W; 35°–48°S | |||||
| Subantártica (ASA) | 25.20–25.30 | 10–30 | 13.5–14.5 | 33.7–34.0 | 5.9–6.2 |
| O2 mínimo | 26-70–26.90 | 220–270 | 6.5–8.5 | 34.1–34.4 | 3.3–5.0 |
| Intermedia antártica (AIA) | 27.10 | 450–470 | 5.5 | 34.3 | 5.5–5.8 |
| (1) Sin datos en Invierno. | |||||
Las principales masas de agua indentificables en el Sistema Chileno–Peruano, son: las Aguas Superficiales Sub-tropicales (ASST), las Aguas Sub–Antárticas (ASA), las Aguas tcuatoriales Sub-superficiales (AESS) y las Aguas Intermed Antárticas (AIA).
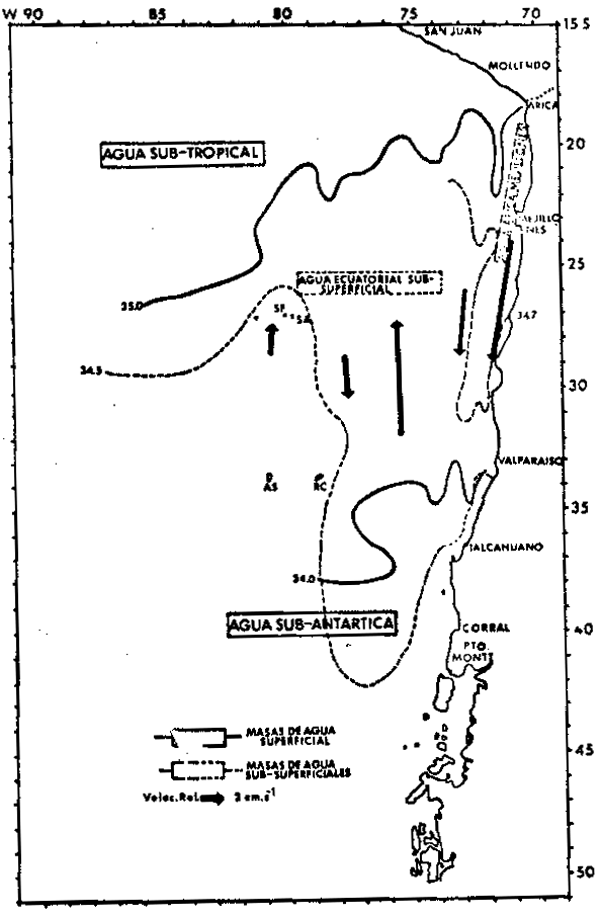
Las propiedades más relevantes de estas masas de agua se resumen en la Tabla 1. A continuación se ilustran los patrones promedio de distribución de estas masas de agua durante el verano (Figura 2). Durante el verano en la región oceánica (más allá de los 150 kms. de la costa), la presencia de las ASST se observa a ambos lados de una penetración hacia el norte de las aguas subantárticas (ASA) entre los 75° – 85°W. La ASST se caracteriza por un máximo de salinidad superficial con valores de 34.8 a 35.0 o/oo, por lo que su límite se muestra en la Figura 1 demarcado por la valores lina de 35 o/oo. Aguas pertenecientes a la ASST se observan generalmente sólo hasta los 30 m. de profundidad. Un amplio rango de temperaturas entre los 15° y los 24° se asocian al máximo salino. Centrada en los 72° 30' W. las ASST presentan una pentración costera hacia el sur que alcanza latitudes de 25° – 27°S en verano y de 23° – 25°s en invierno.
Las aguas sub-antárticas (ASA) son el elemento regional dominante en las capas super– ficiales. Estas son aguas frías, entre 10° y 18°C, y de baja salinidad, entre 32.0 0/00y 34.8 o/oo. La profundidad media del mínimo salino que tipifica a las ASA, decrece desde alrededor de 130 m. en el sector oceánico hasta 40 m. en el sector costero. El derrame hacia el norte de las ASA sigue dos cursos principales, uno oceánico entre 75° y 85°W. y otro más costero a lo largo de los 73° – 74°W.
Las aguas ecuatoriales sub-superficiales (AESS) se identifican por la presencia de un máximo salino sub–superficial con valores de salinidad entre 34.5 o/oo a 34.9 o/oo, asociado a un rango de temperaturas de 7° a 12°C. Las AESS se extienden hacia el sur a lo largo de la costa de Chile hasta cerca de la Isla de Chiloé (42° – 43°S). La presen– cia del núcleo del máximo salino sub–superficial característico de esta masa de agua, se mantiene a lo largo de la costa central de Chile alrededor de los 300m. de profundidad y en general coincide con la capa de mínimo de oxígeno y con la isopicna de 26.8 ot, lo que sugiere un desplazamiento de tipo isentrópico. La capa de mínimo de oxígeno cumple un rol importante en el ecosistema de la región, pues las concentraciones de oxígeno que la tipifican entre 1.5 a menos de 0.5 ml 02/lt, se encuentranentre las más bajas observadas en los oceános, presentándose ocasionalmente en esta capa condiciones de anoxia. Es improbable que la mayoría de las especies marinas, en particular los peces, puedan mantener en forma prolongada sus niveles normales de actividad metabólica al interior de esta capa y para muchas de ellas esta zona está efectivamente excluida de su habitat. Por estas razones se ha sugerido que para algunas especies pelágicas, esta capa podría actuar como refugio ante la presencia de predadores activos (Antezana, 1978).
El agua intermedia antártica (AIA) se identifica por la presencia de un mínimode salinidad profundo. De sur a norte en la región, los valores de este mínimo se elevan desde 34.2 o/oo, encontrándose asociados a un estrecho rango de temperaturas entre 4°y 7°C.
Un cuerpo de que cumple una importante función en la dinámica del sector costero del centro y norte de Chile y sur del Perú, es el agua de Arica-Mejillones (Robles, 1979). Esta masa de agua se caracterza por un máximo salino sub-superficial aún más elevado que el de las AESS, con valores entre 34.7 o/oo y en ocasiones hasta 35.2 o/oo. Asociadas a estas salinidades, se encuentra un rango de temperaturas entre 10° a 15°C. la superficie de 26.4 ∑t y se produce a una profundidad media de 100 m. A diferencia del AESS, el agua de Arica- Mejillones alcanza latitudes menores. En verano y en coincidencia con el desarrollo de aguas con altas salinidades tanto superficial como sub–superficial– mente en la zona noreste de la region, ellas alcanzan aproximadamente los 34°S. En el invierno se repliegan hasta aproximadamente los 31°S. al parecer en conexión con el avance estacional hacia el norte del agua de baja salinidad de de la corriente de los Fiordos.
Los patrones medios de circulación en la región pueden ser descritos apropiadamente con referencia a la Figura 3, que presenta una sección de velocidad geostróficas relativas a 1000 decibares, calculadas en base a una sección hidrográfica a perpendicular a la costa en los 28° 15' S., efectuada durante le Expedición Scorpio en el otono de 1967 (SIO–WHOI– MIT,1969).
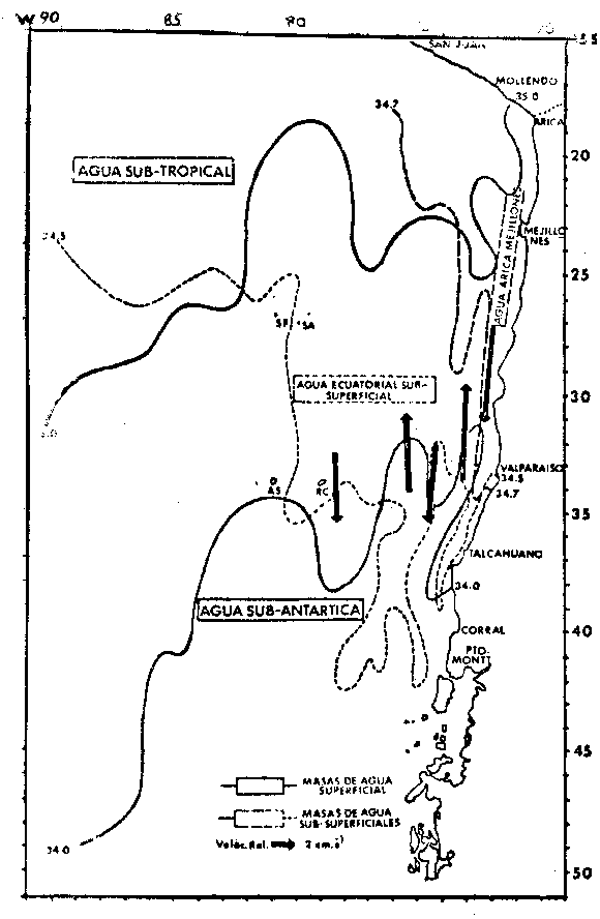 |
Fig. 1. Masas de agua y ciculación en el
Pacífico Sureste Meridional duante
Verano. Se representa esquemáticamente la extensión de las principales
masas de agua superficiales (línea
continua) y subsuperficiales Ilínea
de trazos). Los flujos principales
del Sistema de Corientes de Chile-
Perú están representados por vectores
que longitud es proporcional a la
velocidad promedio (modificado de
Robles, 1979). |
Fig. 2. Masas de agua y circulación en el
Pacífico Sureste Meridional durante
el Invierno. Se representa esqueáticamente la extensión de las principales masas de agua superficial
(línea continua) y subsuperficial
(línea de trazos). Los flujos
principales del Sistema de Corientes
de Chile-Perú están representados
por vectores que indican la dirección del flujo y cuyo longitud es
proporcional a la velocidad promedio
(modificado de Robles, 1979). |
 |
El sector oceánico oeste de la zona se ve dominado por un amplio y lento movimiento hacia el norte, con máximos en sus velocidades relativas de alrededor de 4 cm seg-1, ubi– cados a 1000 o más km de la costa. Este flujo corresponde a la penetración subantártica oceánica, coincidente con el borde oriental del giro anticiclónico del Pacífico Sureste y ha sido denominada la Corriente Oceánica Chileno-Peruana (Robles, Alarcón y Ulloa 1980).
Hacia la costa, se observa un flujo hacia el sur, que de acuerdo a Wyrtki (1967) , entra en la región alrededor de los 79°W, para seguir luego un curso paralelo a la costa a lo largo de los meridianos 76°–77°W, a 500 km. de la costa. Este flujo ha sido denomi– nadopor Wyrtki la Contracorriente Oceánica Peruana, y en la sección de la Figura 3 aparece representado con un máximo de velocidad de alrededor de 6 cm. seg-1 a 300 m de profundidad.
Entre 300 y 400 km. de la costa se aprecia el flujo comparativ amente más rápido de la Corriente de Humboldt, con un núcleo de máximas velocidades relativas de más de 18 cm. seg-1, alrededor de los 200 m de profundidad.
Entre la corriente de Humboldt y la costa, eventualmente pueden identificarse otros tres flujos, cuya presencia y constancia es más variable, particularmente en relacion al dominio de perídos fríos o cálidos en el Pacífico Sureste. El más permanente de estos tres flujos corresponde a una contracorriente, la llamada Contracorriente Costera de Chile (Robles, Alarcón y Ulloa, 1980), que en la Figura 3 aparece desplazándose hacia el sur a aproximadamente 100–200 km de la costa con máximas velocidades relativas en super– ficie de 10 cm seg-1.
El segundo flujo detectable en la zona costera, es una corriente muy superficial hacia el norte, cuya presencia es más marcada en asociación a los perídos cálidos. Esta corriente, que por corresponder 1967 a un período frío, no aparece en la Figura 3, ha sido denominada por Robles (1979) la Corriente de los Fiordos. Silva y Sievers (1981), la mencionan como la rama costera de la Corriente de Humboldt. Cuando este flujos superficial se encuentra desarrollado, se desplaza a unos 100 km de la costa, exhibiendo velocidades relativas entre 6 y 16 cm seg-1, pudiendo presentar flujos tipo “jets” con velocidades de hasta 40 cm seg-1. Johnson, Fonseca y Sievers (1980) detectron por medio de mediciones directas (correntometría) en los 33°S, un flujo costero superficial hacia el norte de unos 30m de profundidad, que puede corresponder a la Corriente de los Fiordos. Este flujo fue observado como distinto y superpuesto a una contracorriente subsuperficial relativa– mente acelerda (25 cm seg-1) núcleo se ubicaba a 100 m de profundidad.
El tercer flujo de la zona costera, es observable inmediatamente fuera de la costa y refleja en las regiones norte y central de Chile, el derrame hacia el sur de las aguas de Arica-Mejillones. Las velocidades geostróficas máximas de esta contracorriente general– mente fluctúan alrededor de los 20 cm seg-1, localizándose a 200 m de profundidad. En conjunto con la Corriente de los Fiordos, esta contracorriente participa y es afectada por los procesos de afloramiento costero, de manera que dependiendo de la latitud, estación del año, y variabilidad interanual, puede exhibir desde velocidades entre 4 – 6 cm seg-1 hasta desplazamientos superficiales y subsuperficiales fuertemente acelerados tipo “jet” En la zona norte de Chile, el núcleo de esta corriente yace unos 100 m de profundidad al igual que en la costa sur del Perú (Brockmann, Fahrbach, Huyer y Smith, 1980), este núcleo coincide o se ubica sobre la capa de mínimo contenido de oxígeno. En términos generales, este flujo hacia el sur es de bastante permanencia y parciera corrsponder más apropiadamente a la Corriente de Gunther o a la “Perú–Chile undercurrent” mencionadas por diversos autores en la literatura.
El Ecosistema pelágico del margen oriental del Pacífico Sur–este
El Pacífico Sureste y en particular el Sistema de Corrientes de Chile-Perú, es uno de los regímenes oceanográficos del mundo que presenta los más altos niveles de producción biológica. Frente a lascostas de Perú se han obtenido mediciones de producción primaria de 11.74 g–Cm2/día (Ryther et al.,1971). En contrastes a estas mismas latitudes, en el ecosistema del giro central del Pacifico es dable esperar valores de sólo 100 a 300 mg–cm2/ día, dos órdenes de magnitud menores.
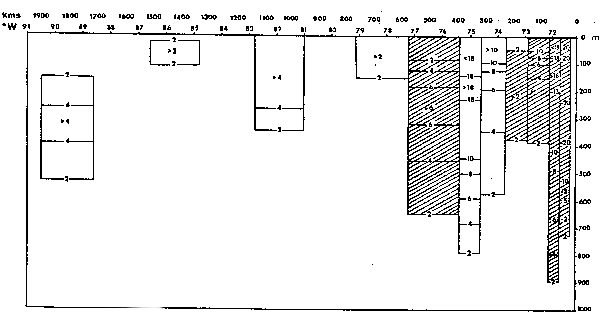
Fig. 3.- Sección de velocidades geostróficas en un corte perpendicular a la Costa en los 28°15'S., Otoño 1967. Las velocidades que se presentan en la figura, están referidas a un nivel de cero movimiento a 1000 dėcibares y se expresan en cm seg-1., La sección se extiende desde la costa hasta los 91°W (1900 Km de la costa). Los datos utilizados en la confección de esta figura provienen de la Expedición Scorpio (SIO-WHOI-MIT, 1969). Los segmentos achurados indican flujo en dirección al sur y los blancos hacia el norte.
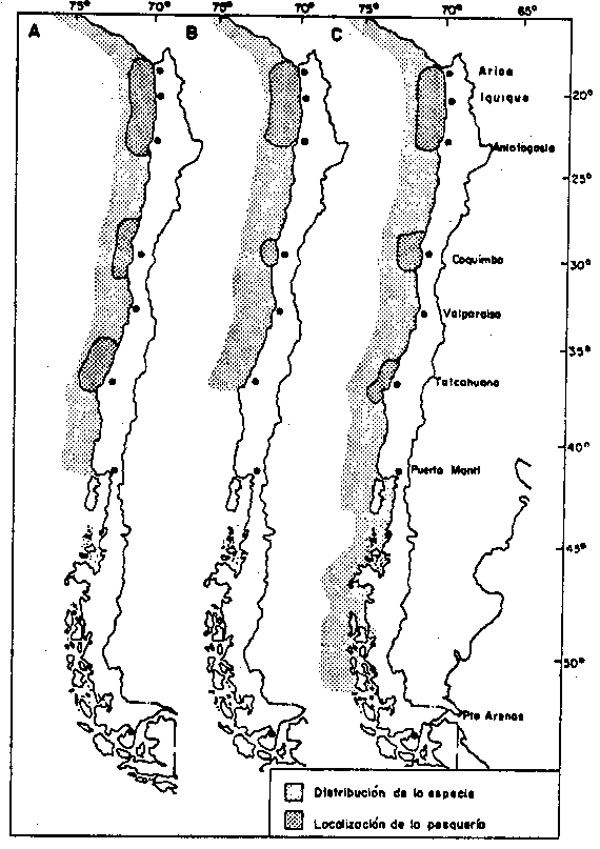
Fig. 4.- Areas de distribución de las principales especies de peces pelágicos y localización de sus pesquerías. A: Anchoveta Engraulis ingens; B: Sardina Sardinops sagar; C: Jurel Trachurus murphyi. |
 |
Esta mayor riqueza biológica tiene origen en dos procesos oceanográficos que actúan sinergísticamente. En primer lugar la presencia de surgencias costeras a lo largo del centro y sur del Perú y norte y centro de Chile, que incorporan nutrientes inorgánicos a las capas iluminadas de la superficie; y en segundo lugar el aporte de nutrientes que se incorporan al sistema desde las altas latitudes por advección en los flujos principales del Sistema de Corrientes de Chile-Perú.
Las surgencias costeras tienen lugar en una estrecha banda estrictamente costera, equivalente a un radio de deformación de Rossby. En las latitudes del Pacífico Sureste Meridional, considerando la estratificación observada del campo de densidad en las capas superficiales, esta banda tiene un rango entre 4 a 35 km de ancho. La zona biológicamente enriquecida del Ecosistema por el contrario se extiende mucho más allá de los límites de esta banda, lo que pone de relieve la importancia para el ecosistema de los flujos advectivos de nutrientes
Por su caractér abierto y con fuerte influencia de la advección, gran parte de la biota presente enel Ecosistema, tiene afinidades con faunas cuyos centros de localización se encuentran fuera del mismo. Además de una fracciónmenor de componentes endémicos al Pacífico Sureste, en el ecosistema pelágico se encuentran especies de origen subantártico subtropicales, tropicales y cosmopolitas de aguas temperadas. En su borde exterior, aproximadamente a 1000 km de la costa, se ubica la frontera que lo separa del ecosistema del Giro Central del Pacífico Sureste, el que posee una biota propia definida y de mayor diversidad específica.
Los ecosistemas marinos y especialmente aquellos localizados en sistemas de corrientes son estructuras biológicas organizadas cuya fenomenología está dominada en gran medida por procesos físicos, lo que cualquier intento de comprender y explicar su diná– mica biológica deba cosiderar esta propiedad. Por estas razones los ecosistemas de margen oriental han sido considerados como ecotonos, ya que los procesos biológicos interactivos clásicos, como competencia y predación serían secundarios como mecanismos regulatorios al no tener oportunidad de jugar un rol dominante la modulación de los cambios de abundan– cia de las especies debido al efecto disruptivo de las cambiantes físicas (McGowan, 1974)
Los dos procesos físicos responsables de la entrada de nutrientes de origen no-regenerativo al sistema, surgencias y advección horizontal de gran escala presentan una diná mica con una marcada señal estacional. En la región meridional del Pacífico sureste, las surgencias costeras son máximas durante prima–verano, a diffencia de lo que ocurre en la costa peruana donde son máximas durante invierno. Esta estacionalidad en la costa chilena se origina como respuesta directa al predominio de vientos locales del sur y suroeste, en cambio en Perú debido a la distinta orientación de la línea de cost, se debe al pre– dominio de vientos del sur y sureste.
La estacionalidad en la circulación principal del sistema se acelera e intensifica durante el fin del verano y otoñ
La producción primaria en estas latitudes no es limitida por la disponibilidad de energía radiante, por lo que no es sorprendente que ella siga también un marcado ciclo estacional en respuesta a la disponibilidad de nutrientes inorgánicos en las capas super– ficiales, principalmente N–NO3.
Este Ecosistema ha sustentado la pesquería más grande de la historia: la pesquería peruano–chilena de anchoveta en la década del sesenta (objeto de otras contribciones en este volumen). La Figura 4 muestra las áreas de distribución geográfica y de localización de las pesquerías actuales de la anchoveta, E. ringens sardina española Sardinops sagax y Jurel Trachuras murphyi en la Región meridional del pacífico Sureste. En esta región también se explota la caballa Scomber japonicus. Esta pesquería, caracterizada por una captura diversificada, contrasta con la pesquería casi monoespecífica sobre E. ringens durante la década del sesenta. El ecosistema ha experimentado cambios significativos en cuanto a la composición de su fauna íctica pelágica con posterioridad a el fenómeno de El Nińo de 1972, que marcara el colapso de la pesquería de anchoveta peruana.
El más dramático de estos cambios es el aumento de las capturas de S. sagax y la extensión hacia el sur área de desove, cambio reminiscente del reemplazo inverso de la sardina por la anchoveta en el ecosistema californiano enla década del cincuenta. La captura de S. sagax 1979 alcanzó 1.6 millones de toneladas en Chile y en 1982 llegó cerca de los 2 millones de toneladas.
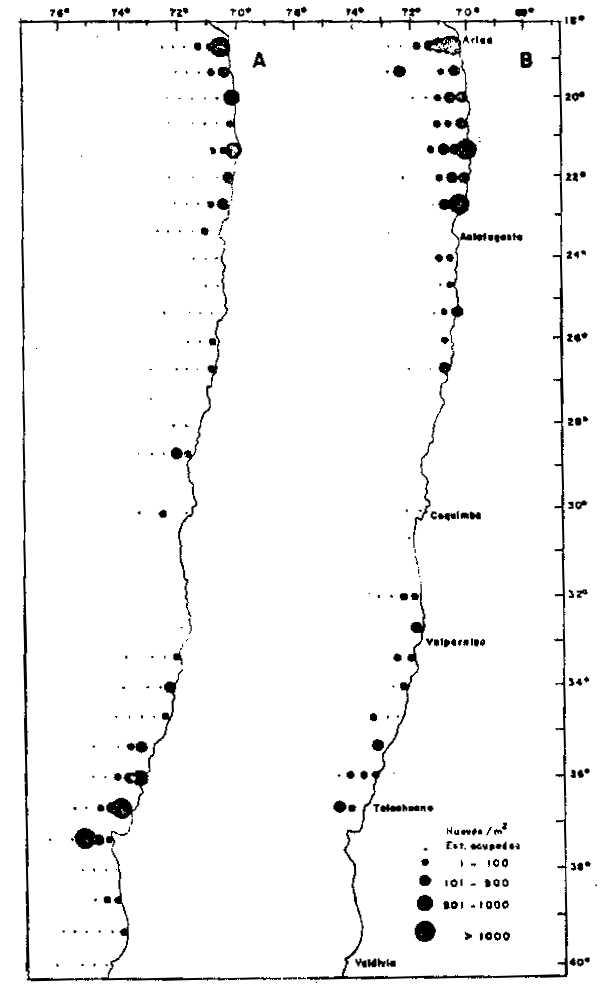
Fig. 5. - Distribución geográfica y abundancia relativa de huevos de anchoveta Engraulis ringens, 1963–1973. La abundancia relativa se expresa en huevos/m2 de superficie. A: desove de primavera–verano (Noviembre a Enero); B: desove de invierno (Julio a Sptiembre). El deove de primavera–verano se encuentra más restringdo a la costa, situación que coincide con el predominio de las surgencias costeras en la región.
La Figura 5 presenta la distribución geográfica del desove de E.ringens durante el período 1964–1973, basado en datos obtenidos en censos de huevos expresados en huevos/m2 de superficie. El desove de primavera–verano (Figura 5a) aparece más restringido a la costa que el desove de invirno (Fig. 5b). Primavera–verano es la época de máxima inten– sidad de surgencias costeras y en las que éstas se extienden más al sur, lo que coincide con la presencia de un centro de desove en las proximidades de Talcahuano y el Golfo de Arauco (aproximadmente 37°S). En ambos perídos los centros locales de desove con abundancias mayores a 1000 huevos/m2, se encuentran inmediatamente adyacentes a la costa y por lo tanto su extensión es conmensurable con el radio de deformación de Rossby. Esto sugire que la localización de estos centros está conectada a la presencia de centros de surgencia costera.
Variación Oceanográfica de baja frecuencia y el fenómeno de El Niño
Recientemente y como resultado de la disponibilidad de largas series de tiempo en el sistema de la Corriente de California, ha sido posible demostrar que en los sistemas de margen oriental aparte de la clara y estadísticamente definible estacionalidad, existen fluctuaciones de baja frecuencia o de largo período. Esta componente de la variabilidad total de estos sistemas ha sido observada tanto en la dinámica física (Chelton, 1980 y 1981), como en la dinámica biológica (Bernal, 1979 y 1980). Más aún, ha sido posible demostrar que éstas covarían en forma significativa indicando que existe algún grado de acoplamiento entre ambas dinámicas, en el dominio de bajas frecuencias (Bernal, 1981; Chelton y McGowan, 1982).
En el Sistema de Corrientes de Chile–Perú, el fenómeno de El Niñorepresenta la expresión más clara y de mayor importancia de este tipo de variabilidad (Robles, 1979). En téminos oceanográficos, El Niño es una invasión masiva de aguas cálidas al sistema de Chile-Perú, alterando la dinámica de sus flujos y la distribución de sus masas de agua. Los eventos de El Niño son la expresión de fenómenos de interacción océno–atmósfera a escala de toda la cuenca del Pacífico mediados a través de mecanismos que se desarrollan a nivel del Pacífico Ecuatorial.
La mecánica de esta interacción se puede conceptualizar en forma simple a partir del balance entre dos tipos de circulación atmosférica: la Circulación de Walker y la Circula– ción de Hadley. La Circulación de Walker es una celda convectiva de la atmósfera sobre el ecuador que responde a las gradientes térmicas superficiales del océano presentes a lo largo del ecuador. Cuando la temperatura superficial del mar es fría en el Pacífico Este, la convección (i.e. ascensión) de las masas de aire sobre ésta zona se debilita. En estas condiciones el aire desciende hacia la superficie del océano, fluye hacia el oeste para finalmente elevarse sobre las vastas regiones cálidas del Pacífico Oeste (Indonesia, Archipiélago Malayo). La Celda convectiva se cierra mediante el flujo de retorno hacia el este que se efectúa a través de la tropósfera superior. En los niveles bajos de la atmósfera, esta circulación está asociada a presiones atmosféricas altas en el Pacífico Oriental y bajas en el Pacífico Occidental.
La circulación de Hadley es una celda convectiva orientada meridionalmente, producida por la diferencia de temperatura entre el ecuador y las latitudes medias. Cuando en la zona ecuatorial las temperaturas superficiales del mar se presentan anómalamente cálidas, las masas de aire convergen hacia el ecuador, se levantan sobre la zona cálida y fluyen hacia el sureste y noreste por la tropósfera superior. Los vientos Alisios al norte y sur del ecuador se intensifican y por medio de teleconexiones con latitudes medias, aumentan la intensidad de los Vientos Bravos del Oeste, centrados a los 40°S. Debido a la convergencia, en bajas altitudes la presién atmosférica es anómalamente baja cerca del ecuador y alta en la zona subtropical.
En condiciones normales enel ecuador, el Pacífico Este (adyacente a las costas de Centro y Sudamérica), es comparativamente frío respecto al Pacífico Oeste (Indonesia, Archipiélago Malayo), por lo que la circulación de Walker se presenta bien desarrollada y los vientos de superficie asociados a ella ejercen un esfuerzo (stress) sobre la superficie del mar en dirección al este. En una primera aproximación este gradien– océan responden a esta fuerza de los vientos estableciendo un gradiente de presión de signo contrario, es decir en dirección al este. En una primera aproximación este gradiente puede visualizarse como una mayor elevación de la superficie del mar en las zonas adyacentes a Indonesia.
El Océno y la atmósfera, debido a su distinta capacidad calórica, se calientan(o enfrían) a distintas velocidades. Esta diferencia produce la aparición de extensas zonas de la superficie oceánica con anomalías térmicas positivas y negativas, que actúan como fuentes o captadores del calor atmosférico determinando el predominio de distintos regímenes meteorológicos.
Cuando aparece una anomalía positiva de la temperatura superficial del océano en el Pacífico Este, el gradiente térmico a lo largo del ecuador se debilita, debilitando a su vez la circulación de Walker. El equilibrio entre atmósfera y océano se rompe porque la fuerza (stress) ejercida por el viento no alcanza a balancear el gradiente de presión de las capas superficiales del océano. En las vecindades del Ecuador, donde los efectos de la rotación terrestre sobre las corrientes oceánicas son muy pequeños (el parámetro de Coriolis es idénticamente cero en el Ecuador), este desbalance produce underrame de aguas cálidas que fluye por la pendiente de presíon hacia el Pacífico Este. McCreary (1976) ha modelado este flujo como ondas planetarias de Rossby atrapadas en la singularidad ecuatorial, las que al llegar al continente Americano son atrapadas por el campo de densidad costero en la forma de ondas Kelvin, las que se propagan en forma simétrica hacia ambos hemisferios: norte y sur. Este derrame produce un hundimiento de la picnoclina (o termoclina) y al transportar por advección cantidades importantes de calor causa un aumento adicional de la anomalía térmica inicial. Este mecanismo actúa por lo tanto como un circuito de retroalimentación positiva, ya que provoca una disminución progresiva del gradiente térmico en el Ecuador, lo que torna aún más débil y eventualmente elimina la circulación de Walker.
La suma de estos procesos culmina con el evento de El Niño en el Pacífico Suroriental, donde entre otros efectos se observa el avance hacia el sur a lo largo de la costa norte del Perú, de aguas ecuatoriales superficiales de alta temperatura y baja salinidad, que se extienden considerablemente más al sur de su límite habitual. Otras consecuencias asociadas al fenómeno, son el debilitamiento o cesación del afloramiento costero, lo que permite el acercamiento a la costa de aguas oceánicas de altas temperaturas y altas salinidades. La reaparición de condiciones frías en el Pacífico Este invierte el proceso, reactivando y finalmente manteniendo la circulación de Walker.
Cabe hacer notar que el conocimiento actual sobre el fenómeno de El Niño apunta a su carácter bi–hemisférico. Bernal (1976) presenta evidencias que demuestran la presencia de El Niño de 1957–58 en las costas de California y Chelton, Bernal y McGowan (1982) han demostrado que el aparente nexo entre el flujo de la corriente de Calífornia con el forzamiento de los vientos a escala del Pacífico Norte, realmente refleja el acoplamiento de la Corriente de California con la aparición del fenómeno de El Niño en el Pacífico Oriental. Esto último resulta del hecho que a gran escala los vientos sobre el Pacífico norte están significativamente correlacionados con los eventos de El Niño.
Hay una estrecha conexíon entre la permanencia de la Circulación de Walker y las variaciones de la llamada “Oscilación del Sur”. La Oscilación del Sur implica un intercambio masivo de masas de aire entre el Pacífico Sureste y el Océano Indico Oriental (cerca de Indonesia). La Circulación de Walker es más activa en una situación de la Oscilación del Sur en que el Centro de Altas Presiones del Pacífico Sureste y el Centro de Bajas Presiones de Indonesia se encuentran ambos particularmente desarrollados. La situación inversa, Circulación Walker debilitada, se presenta cuando ambos centros están poco desarrollados. La variación interanual de la Oscilación del Sur se halla también relacionada a anomalías del nivel del mar que se observan a lo largo de una vasta región del margen oriental del Pacífico. De acuerdo a Enfield y Allen (1980), anomalías del nivel del mar registradas entre Crescent City (California) y Antofagasta (Chile), mostraron un alto grado de correlación con las variaciones de la Oscilación del Sur para el mismo período. (P.<0.0.1).
Quinn (1974a, b) ha elaborado un Indice de Oscilación del sur en base a las diferencias de presiones atmosféricas entre el centro de Altas Presiones del Pacífico Sureste, tomado en Isla de Pascua, y el Centro de Bajas Presiones de Indonesia, tomado en Darwin, Australia. La Figura 6 muestra para el período 1964–1973 este índice, junto a las desviaciones de temperatura superficial en Chicama, Perú y las desviaciones de la presión atmosférica en Arica, Chile. La concordancia que existe entre estas series de tiempo subraya la gran escala espacial de los procesos envueltos en el fenómeno de El Niño y en los períodos fríos inversos o anti–El Niño. En la Figura 6 se señalan como períodos fríos los años 1964, 1967, 1970–71 y 1973 y como períodos cálidos reconocidos como El Niño, a 1965, 1969 y 1972.
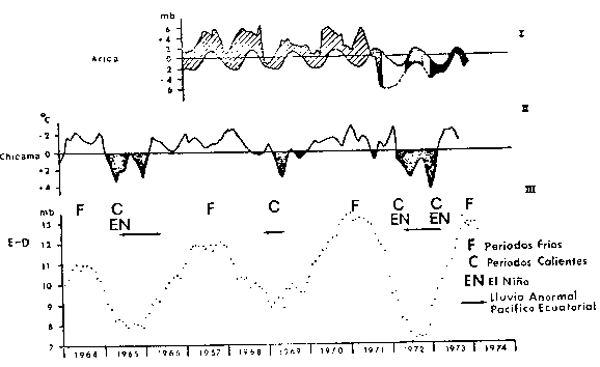
Fig. 6. - Series de tiempo indicativas del fenómeno de El Niño. I. Indice de presión atmosférica en Arica, Chile (de Robles et al., 1976). II. Anomalías de la temperatura superficial del mar en Puerto Chicama, Perú (de Miller y Laurs, 1975). III. Indice de la Oscilación del Sur: presión atmosférica en Isla de Pascua menos la presión atmosférica en Darwin, Australia (de Quinn, 1974a y 1974b). Sobre la serie del Indice de la Oscilación del Sur se indican los períodos fríos (F) y cálidos (C) que se presentaron durante 1964–1974. Las flechas horizontales indican presencia de lluvias anormales en el Pacífico Ecuatorial.
Los períodos fríos (F en la Figura 6) se ven asociados a la intensificación del torque atmosférico y de la circulación oceánica alrededor del centro de Altas Presiones del Pacífico Sureste. En la región meridional del Pacífico Sureste esto se traduce en una intensificación del flujo subantártico hacia el norte, con un dominio en las capas superficiales del océano de bajas temperaturas y bajas salinidades. Inversamente los períodos cálidos (C en la Figura 6) son concurrentes con períodos de relajación en la circulación atmosférica y oceánica del Pacífico Sureste y por lo tanto se suceden a la aparición de grandes diferencias en el Indice de la Oscilación del Sur (E–D en la Figura 6).
De acuerdo a Zuta et al. (1980) las manifestaciones físicas asociadas al fenómeno de El Niño en el Pacífico Suroriental son:
i) un derrame anormal hacia el sur, muy superficial, de aguas de altas temperaturas (23°–29°C) y bajas salinidades (32.5 o/oo – 34.5 o/oo). En la costa peruana estas aguas alcanzan hasta los 14°S;
ii) fuertes precipitaciones tropicales que se extienden hasta los 7°S y que superan entre 3 a 10 veces los promedios normales;
iii) un debilitamiento o cese de las surgencias costeras;
iv) aguas oceánicas de altas temperaturas y altas salinidades se aproximan a la costa como producto del debilitamiento de la divergencia costera.
Los dos últimos efectos son característicos a la región Meridional del Pacífico Suroriental.
Una observación importante y que no ha sido suficientemente enfatizada en estudios previos sobre El Niño, precisamente por ser propia a la Región Meridional del Sistema de Chile – Perú, es la drástica disminución de las salinidades en la región costera, en especial en la zona inmediatamente contigua a los fiordos del sur de Chile. Este es un resultado más del calentamiento de gran escala y de la relajación del torque atmosférico y deriva en forma directa del incremento de la pluviosidad en latitudes intermedias y de los aportes de los ríos en las regiones Central y Sur de Chile. Enfield (1981), citando a Miller y Laurs (1975), sugiere que los sistemas ciclónicos pudieran haber tenido más frecuentes desplazamientos hacia el norte durante El Niño de 1972. Esta hipótesis es consistente con la evidencia presentada por Enfield (1981) mostrando que tanto el núcleo de altas velocidades de los vientos como el centro del anticilón del Pacífico Sureste se localizaron más al norte de sus posiciones normales durante El Niño de 1976 – 77.
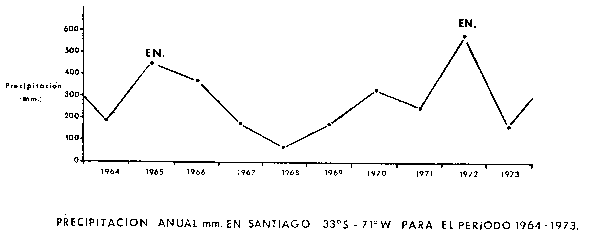
Fig. 7. - Precipitación anual en Santiago de Chile para el período 1964–1973. La precipitación se expresa en mm de agua caída. Máximos de pluviosidad anual que exceden los 420 mm han coincidido con el desarrollo de El Niño. Correlaciones similares se observan con datos de otras estaciones meteorológicas ubicadas en latitudes intermedias.
La Figura 7 muestra los promedios de precipitación anual en Santiago de Chile (33°S, 71°W.), para el período 1964–1973. Los promedios más elevados observados durante el período corresponden precisamente con los dos El Niño más importantes de la década. Si consideramos que el Invierno de 1982 ha sido uno de los más lluviosos en Santiago de Chile, habiéndose producido el desborde del Río Mapocho, un evento no observado en los últimos 50 años, no es demasiado arriesgado sugerir que un El Niño pudiera estarse gestando para la Primavera y Verano de 1982–83.*
* Partes del presente trabajo fueron presentadas al Segundo Seminario–taller Bases Biológicas para el Manejo de Recursos Renovables, realizado en Santiago del 23 al 27 de Agosto de 1982, fecha en la que los datos de precipitación para 1982 no existían. Forrest Miller, de la Comisión Interamericana del Atún Tropical sugirió en Agosto de 1982, en base a los cambios en la estructura térmica y los patrones de circulación de vientos en el Pacífico, el posible desarrollo de El Niño a fines de 1982, información no conocida por los autores a la presentación del trabajo. Entre Agosto de 1982 y la fecha de publicación de este volumen se ha desarrollado un El Niño en la región. En el número 91 del Boletín Informativo de la Comisión Permanente del Pacífico Sur se lee: “El Niño de 1982 se ha presentado a partir de octubre de 1982, con trastornos climáticos y oceanográficos en todo el Pacífico Sureste y, al parecer, asociado a similares trastornos en los más alejados lugares del planeta. Las lluvias muy intensas y persistentes, principalmente en Ecuador, con inundaciones y aluviones están produciendo daños incalculables en vidas humanas, agricultura, viviendas, transporte, subsistencia y salud. Por otro lado, la abrupta disminución de recursos pesqueros, propios de la zona costera, han ocasionado que la industria pesquera en Perú llegue a un estado de emergencia. El Niño de 1982 actualmente en desarrollo, al parecer será el más severo de los últimos 50 años y sólo comparáble con aquél de 1925”. Aparentemente anomalías positivas en las precipitaciones a latitudes intermedias, son un índice robusto para predecir la presencia de El Niño en Pacífico Sureste Meridional, y puede ser un índice útil a contrastar para la predicción de El Niño de alta intensidad y desarrollo completo en la región de Perú y Ecuador, y el Hemisferio Norte.
Condiciones oceanográficas en el Pacífico Sureste Meridional durante El Niños y Anti- El Niños.
Los cambios en la distribución de las masas de aguas en la zona meridional del Sistema de Corrientes de Chile–Perú durante períodos fríos y cálidos han sido discutidos en detalle por Robles (1979) utilizando el método de los núcleos modales. Esta metodología define los parámetros de una masa de agua que la identifican y permite por lo tanto trazar sus desplazamientos horizontales y verticales en respuesta a cambios en la dinámica del sistema. Estos núcleos corresponden a máximos y mínimos de salinidad que en un diagrama T–S (temperatura–salinidad) se encuentran asociados con un rango constante de temperaturas y por consiguiente con una misma densidad.
A modo de ilustración se han seleccionado tres inviernos, correspondientes a los años 1967, 1972 y 1973. Respectivamente ellos tipifican condiciones durante un año frío, un año de El Niño y condiciones posteriores al evento. En la Figura 8 se muestra la distribución, en la zona entre 17° y 25°S del máximo de salinidad superficial.
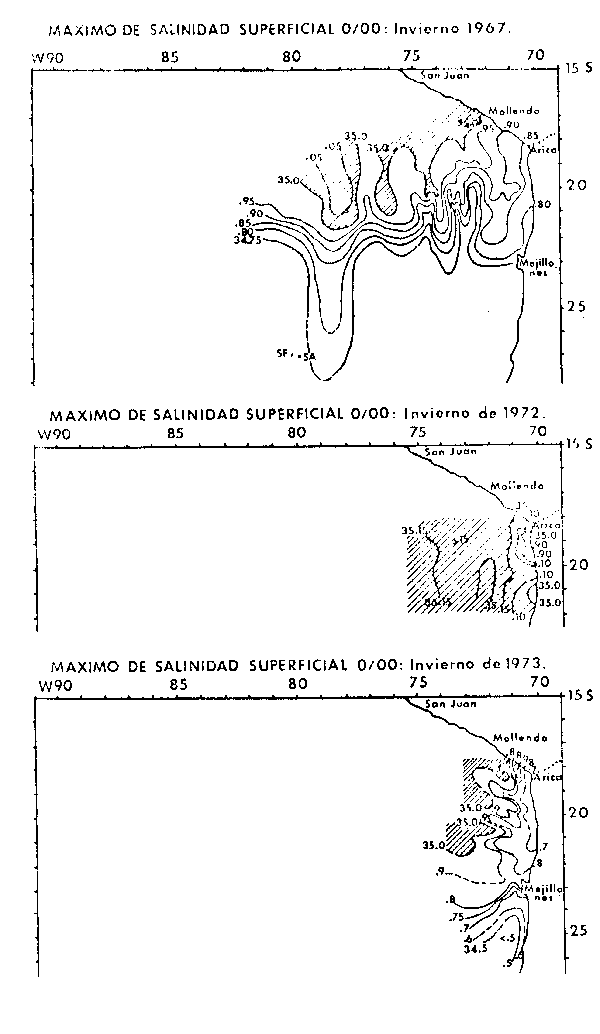 |
Fig. 8. - Extensión del máximo de
salinidad superficial
durante los inviernos de
1967, 1972 y 1973. Areas
con salinidades mayores
a 35 o/oo se muestran
achuradas. El invierno
de 1967 es representativo
de las condiciones dominantes durante un período
frío. En 1972, un año de
El Niño, la zona muestreada se encuentra cubierta
con aguas de salinidades
mayores a 35 o/oo, con
excepción de dos pequeños
lentes consteros. 1973
presenta condiciones de
transición posteriores a
El Niño. Ver discusión
en el texto. |
Durante 1967 las salinidades altas se ven desplazadas hacia el norte en comparación con las condiciones medias. Valores superiores a 35o/oo, sombreados en la Figura 8, se aprecian hacia el norte de la zona, siendo separados de las regiones costeras por salinidades bajas vinculadas a regiones ubicadas más al Sur. Pueden observarse tres intrusiones de altas salinidades hacia el sur: una a los 78°W, otra cerca de los 74°W y la tercera, más disminuída, ocupando la región de Arica-Mejillones. Durante el invierno de 1972, el máximo salino superficial aparece grandemente extendido en comparación con 1967. Las salinidades inferiores a 35.0 o/oo han sido desplazadas hacia el sur y hacia áreas costeras muy restringidas. El patrón predominante es de altas salinidades, homogéneas, que ocupan prácticamente la totalidad del rincón noreste de la región, con solo algunas indicaciones débiles de rasgos observados en 1967, por ejemplo una muy atenuada versión de la lengua de baja salinidad a los 72°–73°W. En 1973 los núcleos correspondientes al máximo salino superficial, muestran en comparación al invierno de 1972, un retiro generalizado hacia el oeste de las aguas subtropiclaes de alta salinidad. Consecuentemente, las áreas costeras se presentan re–ocupadas por aguas de salinidades relativamente bajas, siendo estos valores aún más bajos que los observados en 1967. Esto puede deberse a que la fuente de las aguas costeras para 1973, debió estar relacionada al mínimo salino costero, el que experimentó un reforzamiento durante el período cálido previo, a causa de la intensificación de la Corriente de los Fiordos. Las bajas salinidades en esta zona serían así aportadas por el re–establecimiento del afloramiento costero que se observó fuertemente debilitado en 1972.
La Figura 9 muestra los cambios en la distribución del núcleo del máximo de salinidad sub–superficial ecuatorial para 1967, 1972 y 1973. Durante 1967 se pueden identificar las siguientes características:
i) en estos niveles la franja costera se observa enteramente ocupada por aguas de altas salinidades con valores mayores de 34.8 o/oo. Estas se extienden hacia el sur desde el rincón noreste de la región, confinadas a un rango de profundidades restringido entre 100 y 150 m;
ii) en la región de Arica–Mejillones, se observa un lente de baja salinidad con valores superiores a 34.75 o/oo ubicado en los 100 m superficiales. Este lente aparece separando la extensión costera hacia el sur de las aguas de alta salinidad en dos ramas: una más litoral cuyo derrame coincide con profundidades entre 100 y 150 m. y otra rama que se proyecta al oeste del lente, a profundidades mayores de 250 m. La rama costera se asocia al agua de Arica–Mejillones. La rama exterior corresponde a la expresión costera del Agua Ecuatorial Sub–Superficial (AESS);
iii) el flujo central de bajas salinidades hacia el norte, limita también en este caso la extensión hacia el oeste de la expresión costera del AESS. El patrón exhibido por el flujo central de bajas salinidades, aparece reducido a una lengua que se extiende a lo largo de los 73°–74°W. Esta reducción permite un cierto desarrollo hacia el sur de la rama oceánica del AESS, cuyos núcleos alcanzan también profundidades de 250 m;
iv) las salinidades bajas que se observan en la Figura 9 en el sector oeste, corresponden al flujo subantártico oceánico y se ubican en profundidades menores de 200 m.
En 1972 las salinidades se presentan en esta zona, en general, más altas que en 1967, pero en la lente de baja salinidad son correspondientemente más bajas: menos de 34.7 o/oo en 1972 versus más de 34.75 o/oo en 1967. En la zona sur se aprecia la influencia del reforzamiento del sistema de bajas salinidades costeras característico de los años de El Niño, esto se evidencia en la disminución de los valores observados de salinidad en la extensión hacia el sur de la AESS. Esta disminución, comparada con 1967, fue en promedio de 0.2 o/oo. Es interesante recalcar que al margen de la diferencia en los valores de salinidad, el patrón de distribución de la AESS en ambos inviernos es similar. El rasgo más marcado en ambos casos lo constituye la extensión de estas aguas de alta salinidad, a ambos costados de un lente de salinidades relativamente bajas, que permanece en el rincón de la región, frente a la zona de Arica–Mejillones. Al mismo tiempo pareciera que la extensión de la AESS constituye desde la península de Mejillones al sur, una rama costera única.
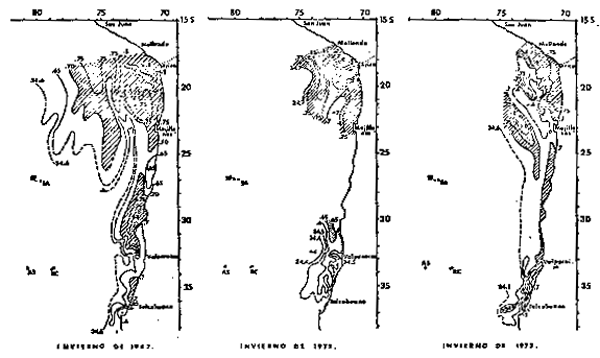
Fig. 9. - Extensió del máximo de salinidad ecuatorial subsuperficial (AESS) durante los invieros de 1967, 1972 y 1973. Areas con salinidades mayores a 34.74 0/00 se muestran achuradas. Los cambios en la distribución de este núcleo se discuten en el texto.
En 1973 la distribución observada parece representar un patrón de transición entre un período cálido y un período friío. El lente de bajas salinidades, con valores inferiores a 34.6 0/00, se present claramente defindo en el rincón noreste de la región. La extensión hacia el sur del núcleo ecuatorial subsuperficial alcanza latitudes mayores que en el invierno de 1972, lo cual es una indicción más del repliegue hacia el sur de la influencia costera de bajas salinidades. Sin embargo salinidades con valores relatviamente altos, superiores a 34.8 0/00, no alcanzan aún las latitudes observadas durante 1967.
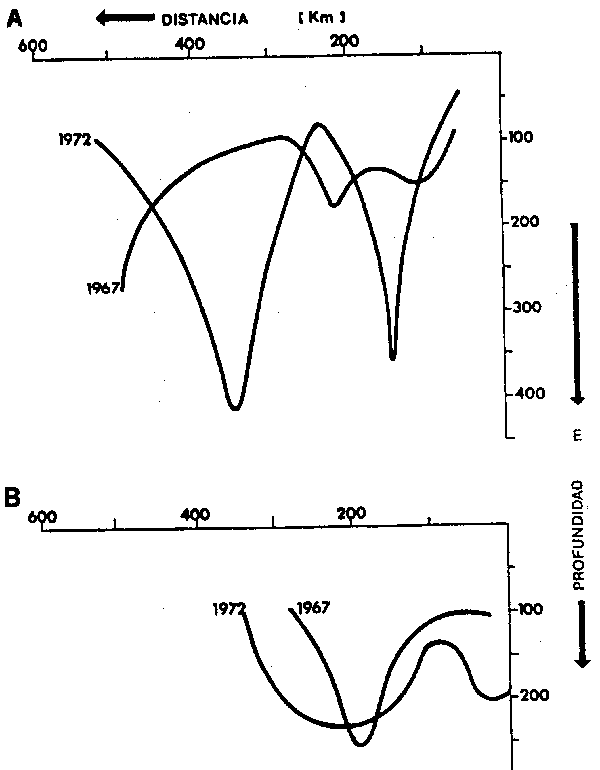 |
Fig. 10 - Cambios en la topografía del máximo de salinidad ecuatorial Sub-superficial durante E1 Niño de
1972. A: topografía del Máximo
de Salinidad Ecuatorial Sub-superficial en un corte a los 19°S.
durante los inviernos 1967 y 1972.
B: topografía del Máximo de Salinidad Ecuatorial Sub-superficial en
un corte a los 32°S durante los
inviernos de 1967 y 1972. |
Gran parte de los cambios que ocurren en la distribución de las masas de agua en respuesta a la influencia de períodos cáliodos yt fríos, son reflejados en su distribución vertical. Los núcleos discutidos en las Figuras 8 y 9 experimentan levantamientos y hundimientos considerables en respuesta a la dinámica anómala de estos períods. La Figura 10 sub-superficial en 1967 y 1972. En el panel superior (“A”) se muestra la topografía de este núcleo en un corte perpendicular a la costa a los 19°S, y en el panel inferior (“B”) uno a los 32°S. El contraste entre los períodos fríos y cálidos es más marcado en la zona norte donde se observa un pronunciado hundimiento de este uncleo durante E1 niño de 1972. En la zona sur las diferencias de niveles no son tan pronunciadas, excepto en la zona costera, en la que durante 1972 este núcleo se hace más profundo en respuesta a la intensificación del flujo superficial hacia el norte de la aguas menos densas y de bajas salinidades de la Corriente de los Firodos.
Variabilidad biológica en el sector meridional del Sistema de Corrientes de Chile-Perú, 1964–1973
La estrecha relación observada en el Sistema de la Corriente de California, entre las variaciones interauales de la biomasa zooplanctónia y eventos oceanográficos de gran escala asociados a períodos cálidos y fríos en el Pacífico Oriental, (Cheloton et al., 1982) permite suponer que una relación similar pudiera ser observble en el Pacífico Sur Oriental.
Como una primera aproximación a esta interrogante, se analizan a continuación series de tiempo (1964–1973) de biomasa del zooplancton, y de huevos de anchoveta, E. ringens, y sardina, S. sagax, en la región meridional el Sistema de Corrientes de Chile-Perú.
Las muestras de plancton de donde se deriva esta información, provienen de arrasteres verticales realizados en los 50 m superificiales con una red Hensen de 0.38 m2 de boca, equipada con una malla de 300 micras.
El tratamiento de los datos ha sido hecho en forma análoga al tratamiento de la información para la Corriente de California (Bernal, 1980). Los datos de biomasa, expresada como volumen de plancton en ml/1000 m3 de agua filtrada, han sido convertidos a log. Los estimadores mensuales de biomasa y de abundancia de huevos, representan el promeedio de todas las observaciones por mes dentro de un área geográfica de acumulación. La Figura 11 muestra las tres áreas de acumulación utilizadas en este estudio. El área I, entre los 18° y 24°S, se caracteriza por presentar una máxima estabilidad vertical a lo largo de todo el año y un mínimo de stress superficial del viento (Bakun y Parrish, 1982). El área II, entre los 24° y 34°S, abarca una zona donde el proceso de surgencia costera es dominante, con claras fluctuaciones estacionales, tanto en la intensidad del stress superficial del viento como en el transporte friccional de Ekman asociado (Bakun y Parrish, 1982). El área III entre los 35° – 40°S, es una zona dominada por características sublatárticas, donde tanto el streess superficial del viento como el transporte de Ekman asociado es muy variable a lo largo del año (Bakun y Parrish, 1982).
El límite oeste de estas áreas de acumulación, se estableció a medio grado de longitud fuera de la costa (56 km. aproximadamente), límite que dado el carácter restringido de la cobertura horizontal de los datos, incluye cerca del 90% de las observaciones disponibles.
 Fig. 12.- La distribución geográfica y abundancia de huevos de sardina Sardinops sagax. A: distribucióndel desove de sardina para el mes de agosto durante el período 1963–72. El desova se encuentra restringido a la zona entre 18° y 27°S. B: desove de sardina durante agosto de 1981. La distribucón del desove de la sardina con posterioridad a El Niño de 1972 muestr un nuevo centro de desove alrededor de Talcahuano y el golfo de Arauco (37°S). |
Fig. 11.- Areas giográficas de acumulación. Los estimadores mensuales de biomasa y de abundancia de huevos y larvas de peces representan el promedio de todas las observaciones disponibles para cada área de acumulación. Latitudinalmente las áreas están limitadas por secciones perpendiculares a la costa en los 18° 24°, 35°y 42°S. El línea mar afuera de cada área estáubicado a 0.5 grados de longitud de la línea de costa (aproximadamente 56 km de ancho), el que se muestra con línea de puntos. 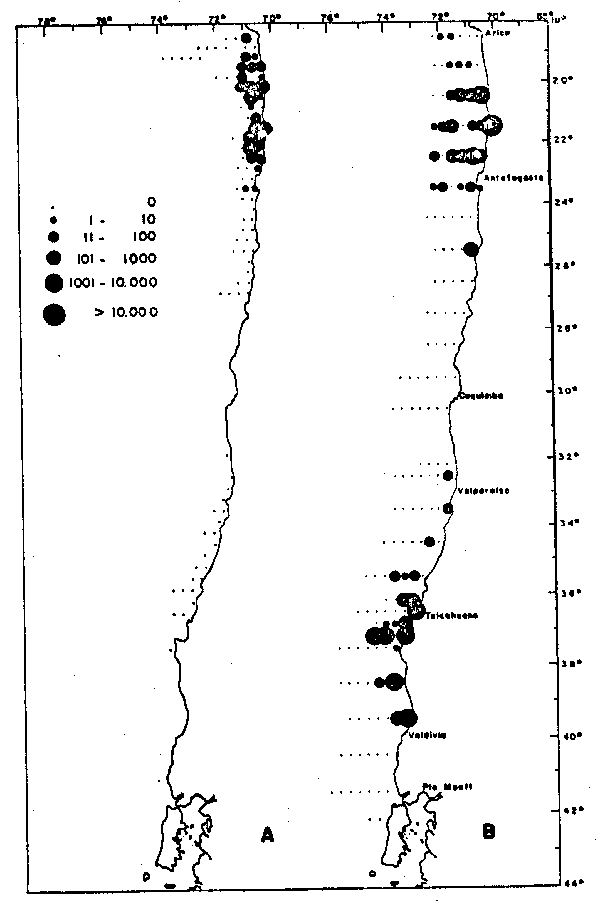 |
La Tabla 2 resume los valores promedios de las variables para las tres áreas. Los promedios de biomasa fueron Calculados sobre todad las observaciones, en cambio para los huevos, se incluyeron sólo los lances de plancton gue presentaron huevos en algún estadio de oesarrollo. En la Tabla 2, “N” representa el número de meses con estimadores mensuales, y “n” el número total de observaciones incluidas en el promedio respectivo.
TABLA 2.- RESUMEN DE DATOS POR AREAS DE ACUMULACION 1964–1973
| BD MASA | HUEVOS E. lingens | HUEVOS S. sagax | |||||||
|---|---|---|---|---|---|---|---|---|---|
| M.G. | N | n | 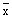 |
N | n | |
N | n | |
| AREA I | 372.8 | 50 | (2010) | 271.6 | 54 | (734) | 42.4 | 43 | (267) |
| AREA II | 256.5 | 25 | ( 561) | 33.9 | 22 | (110) | 2.3 | 4 | (7) |
| AREA III | 273.7 | 15 | ( 170) | 350.3 | 13 | ( 92) | - | - | - |
![]()
![]()
![]()