![]()
![]()
![]()
Un primer resultado de este análisis, es la ausencia de un gradiente latitudinal de biomasa. La presencia de un marcado gradiente latitudinal es quizás la característica global más sobresaliente en el Sistema de la Corriente de California, con valores altos en las latitudes mayores. De haberse repetiodo este partón en el pacífico Sur Oriental, se esperarían altos valores en el área III, situación que no se observa.
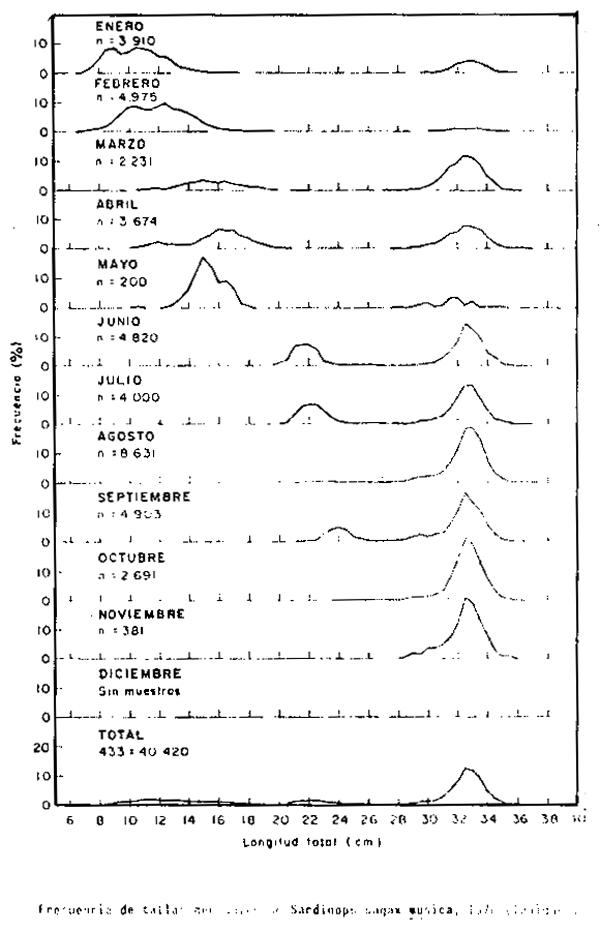
Los valores de densida promedio de huevos de achoveta E. ringens, reflejan bastante bien la localización de sus áreas de desova (Figura 5), con máximos en las áreas I y III. Los huevos de sardina, S. sagax, son notablemente menos abundantes que los E. ringens, refl ejando el dominio de E. ringens, en el ecosistema durante el período en estudio. En el área II, de 561 muestras analizada, sólo 7 contenían huevos de S. sagax, y en el área III de 170 muestras analizada, nunca se encontró un huevo de S. sagax al sur de 35°S entre 1964–73. Esto contrasta dramáticamente con la situación actual en 1982.
En la Figura 12 se mestra la distribución y abundancia de huevos de S. sagax para el período 1964–1973 (Fig. 12A) y para 1981 (Fig. 12B). Se ha escogido el mes de Agosto como representativo de la época de desove. De la comparación de los dos mapas resulta evidente la presencia en 1981 de un nuevo centro de desove alrededor de Talcahuano y el Golfo de Arauco (37°S), inexistente en la década anterior. Esta colonización hacia el sur de S. sagax, es posterior a la declinación del stock de E. ringens que siguió a El Niño de 1972 y refleja el reemplazo en el ecosistema del pez epipelágico filtrador dominante.
Santander y Castillo (1977) reportan las variaciones en la intensidad del desove de S. sagax en la costa peruana donde se observó una situación situación similar a la descrita para Chile. Al igual que en Chile, la estacionalidad del desove en Perú presenta también un máximo en Julio-Agosto. Durante el período 1966–1971, para los meses du Julio-Agosto y Septiembre, estos autores informan del análisis de 672 muestras, habiendo encontrado huevos de sardina sólo en 13 de ellas. El cuociente entre el esfuerzo de muestreo y la abundancia total de huevos puede ser considerado como un índice de la disponibilidad de huevos de anchoveta en la costa peruana. Durante este período este índice de disponibilidad fue de 2 huevos/unidad de esfuerzo. Geográficamente este desove se localizo principalmene en las áreas norte y central del Perú, desde Cabo Blanco (4°10'S) hasta Callao (12°05'S).
Durante 1972, Santander y Castillo (1977) observaron la expansión del desove a toda la costa perunana y su concentración por fuera de 35 millas de la costa, aparentemente evitanao el área costera de desove de anchoveta. El índice de disponibilidad durante 1972 alcanzó valores de 10 huevos/uinidad de esfuerzo. La abundancia promedio de huevos de sardina calculada sobre la base de las muestras positivas en los meses de Julio, Agosto y Septiembre, durante 1966–1971 fue de 103.4 huevos, en cambio durante 1972 fue de 260.2 huevos.
Estos antecedentes sugieren que el área de Arica-Mejillones sirvió durante 1964–1971 como una zona de refugio para las poblaciones de sardina, y que tanto hacia el norte, en Perú, como hacia el sur, en Chile, el desove de esta especie prácticamente no existió, a excepción de la región entre Cabo Blanco y Salaverry en el norte de Perú donde se observaron esporádicamente centros locales de desove.
Desde el punto de vista de la dinamicay genetica de pobaciones, una hipótesis de trabajo interesante es suponer que durante el predominio de la anchoveta en el ecosistema pelágico de Chile-Perú, las poblaciones de sardina se encontraron restringidad a dos zonas refugio separada: Cabo Blanco a Salaverry y Arica a Mejillones, habiendose limitado en forma efectiva el intercambio de información genética entre ambas subpoblaciones. El predominio de la anchoveta fue lo suficentemente largo y la condiciones ambientales lo suficientemente distintas en cada refugio, como para esperar cambios observables con técnica actuales (p.ej. DNA-mitocondriae) en la variablilidad genética de ambas subpoblaciones, aún después de la expansioón de las poblaciones posterior a E1 Niño ede 1972.
Al respecto es imposible dejar de comparar lo ocurrido en Chile-Perú durante la década del sesenta con lo ocurrido en California en la década del cincuenta. En California durancte el peredominio de la sardina, S. caerulea, las poblaciones de anchoveta, E. mordax, se concentraron en la zona de “Los Angeles Bight” (Seno del Sur de California), y en la región de Punta Eugenia en Baja California (Kramer y Ahlstrom, 1968). Con la progresiva expansión de las poblaciones de anchoveta, la sardina ocupo áreas cada vez mas restringidas, concentrándose en la zona del Seno del Sur de California (Kramer, 1970), área que ha servido como refugio para la especie.
En un intento de clasificación de los hábitats de las especies de peces peláicos de las corrientes de margen Oriental, Bakun y Parrish (1982) presentan climagramas construidos con los valores mensuales del cubo de la velocidad del viento superficial versus la magnitud del trnasporte de Ekman paralelo y perpendicular a la costa para distintas regiones geográficas de las costas de Norte y Sundamérica. El cubo de la velocidad de viento es una variable indicativa de la tasa de producción de energía de mezcla turbulenta por el viento en las capas superficiales del océano. En esta clasificación. En esta clasificacion, las regiones del Seno del Sur de California en Norteamérica y la zona de Arica, en la frontera chilenoperuana son las que más se asemejan entre sí, presentando a lo largo de todo el año valores muy bajos de las variables de clasificación. Esto indica que estas zonas son particularmente estables desde un punto de vista oceanográfico, establidad que al parecer está ligada a una mejor calidad de hábitat para las poblaciones de sardina y anchoveta, quienes las utilizan como refugio durante las épocas de predominio y expansión geográfica de su competidor.
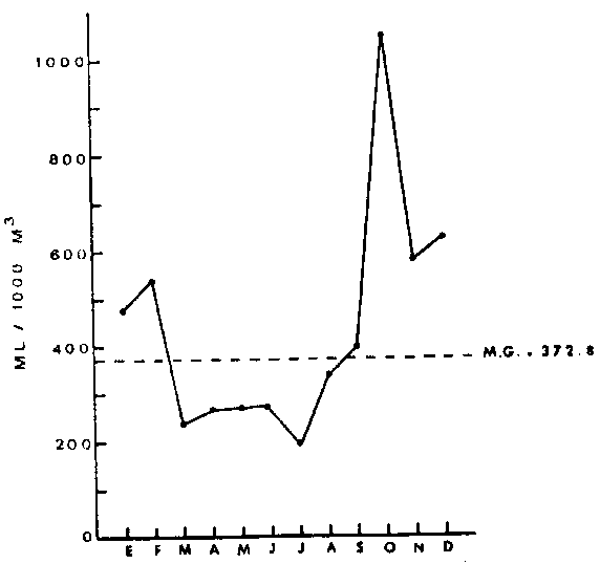 |
Fig. 13.-Variación estacional de la biomasa
de zooplaancton área I. Se grafican
los valores mensuales de las medias
geográficas de biomasa zooplanctónica. las unidades son ml zooplanction /1000 metors cúbicos de agua
filtrada. La mdia geométrica
total para el período 1964–1973 se
representa por un alínea de trazos. |
Las tres variables estudiadeas, presentan una clara componente estacional. La Figura 13 muestra los promedios mensuales de la biomasa zooplanctónica en el área I. Estos promedios están expresados como medias geométricas, producto de la transformación logarímica utilizada (la media gemométrica es el anti-logaritmo del promedio artimético calculado en base a datos log-transformados). El máximo de biomasa se produce en Octubre con un valor mayor a 1000 ml/1000 m3, equivalente a tres veces el promedio para los 10 años (372.8 ml/ 1000 m3). Se observan bajas biomasas durante los meses de otoño e invierno, y un segundo máximo en verano. Este patrón bi-modal coincide con la estacionalidad de las surgencias costeras, que alcanzan en esta zona su máxima expresión durante primavera-verano.
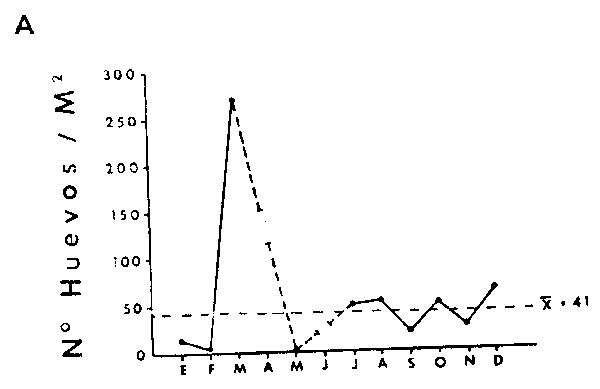
La Figura 14A muestr los promedios mensuales de abundancia de huevos y la Figura 14B los promedios de abundancias de larvas de anchoveta E. ringens. Los gráficos muestran dos máximos coincidentes para huevos y larvas. Wl máximo principal para los huevos ocurre en Diciembre y el secundario en Julio, en cambio lo inverso ocurre con las larvas, donde el máximo principal ocurre en Julio y el secundario en Diciembre. Este patrón estacional coincide con los resultados de Simpson y Gil (1967) basados en el análisis macroscópico de los estados de madurez de las gónadas del stock desovante. La bimodalidad en el ciclo estacional del desove, se también reflejada en el reclutamiento a la pesquería (Simpson y Buzeta, 1967).
La Figura 15A muestra los promedios mensuales de huevos y la Figura 15B los promedios mensuales de larvas de sardina, S. sagax. El ciclo estacional de la abundancia de huevos muestra un máximo en Marzo, que no tiene contraparte en el gráfico de abundancia de larvas. Una inspección de los valores originales, reveló que está sesgado por un solo valor extremo y por lo tanto su validez general es dudosa. Es interesante notar, sin embargo, que tanto el porcentaje de hembras maduras, como el valor del ídice gonádico para el período 1969–1977 (Serra et al., 1979), presenta un máximo en Febrero-Marzo. Esto sugiere que de haberse incoporado al presente análisis información posterior a 1973, este máximo pudiera haberse confirmado. De ser así, el desove de sardina estaría en la actualidad reproduciéndo el patrón de variación estacional del desove de anchoveta.
Esto subraya el hecho que lo ocurido en el ecosistema con p osterioridad a 1972, podría ser un verdadero reemplazo ecológico, al sustituir S. sagax a E. ringens como pez filtrador dominante capaz de explotar el ecosistema en forma similar. La información reportabda por Serra et al., (1979), confirma también la existencia del máximo de Septiembre, en base al muestreo del stock desovante.
En la Figura 16 se muestra la secuencia anual de gráficos mensuales de frecuencia de tallas para la pesquería de S. sagx de Iquiqe (20°S) durante 1978 (Rojas, 1980). El reclutamiento de los juveniles a la pesquería ocurre en Enero, caando, cuando se detectan máximos en la distribucion de tallas con modas en los 9.0 y 10.7 cm. De acuerdo a la información sobre edad y crecimiento disponible (Trautman y Aguayo, 1980), estos grupos modlea están en la vecindad de la longiutd ede peces de un año retrocalculado a partir de medición de otolitos. La longitud a la edad de un año ha sido estimada por dichos autores en 11.9 cm. En ausencia de datos experimentales sobre crecimiento temprano de S. sagax, esta información sugiere que el reclutamiento de Enero podría corresponder a los juveniles provenientes de los desoves tanto de Verano como de Invierno del año anterior.
Para investigar la variabilidad interanual, se seleccionó el área I, por ser la que permitía una major cobertura temporal del período en estudio. A las tres variables analizadas, biomasa zooplanctónica, huevos de E. ringens y huevos de S. sagax, se les eliminó el compondio estacional, recalculado cada estimador mensual como la diferencia entre el valor observado y su promedio mensual. Con estos estimaores residuales se confeccionaron series de tiempo con intervalos trimestrales de acumulación.
 |
|
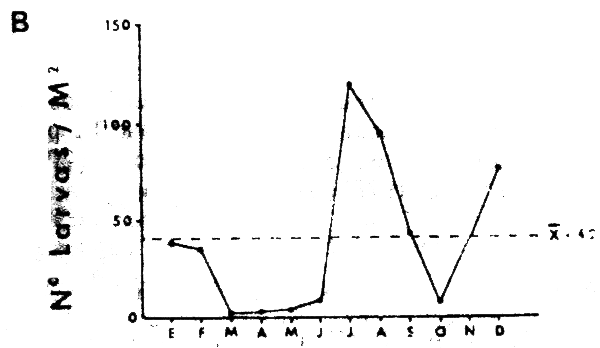 |
Fig. 14.- Variación estacional de huevos y larvas de anchoveta Engraulis ringens. A: promedias mensuales de huevos de anchoveta en aŕea I.B:promedios mensuales de larvas de anchoveta en área I. El promedio total de la serie de datos estáindicado por una línea de trazos.  |
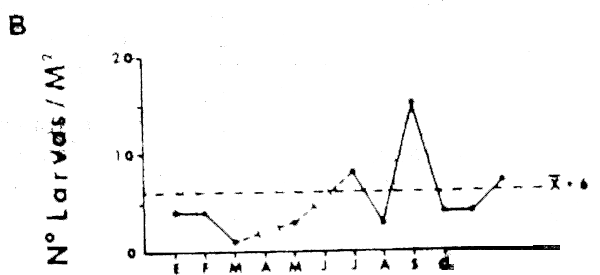 |
Fig. 15.- Variación estacional de huevos y larvas de sardina S. sagax. A: promedios mensuales de huevos de sardina en área I. B: promedios mensuales de larvas de sardina en área I. El promedio total de la serie de datos estáindicado por una línea de trazos. |
 |
|
Fig. 17.- Series de tiempo de biomasa zooplanctónica área I. A: Serie de estimadores trimestrales de biomasa zooplanctónica incluyendo variación estacional. B: Serie de estimadores trimestrales de biomasa zooplanctónica. Residuales representan el componente de variación interanual. Ambas series han sido estandarizadas con sus respectivas desciaciones estándar. Los períodos fríos y cálidos están indicados por las letras F y C respectivamente. |
Fig. 16.- Secuencia anual de distribuciones de frecuencia de tallas para la pesquería de S. sagax de Iquique (20°S), durante 1978 (de Rojas, 1980). 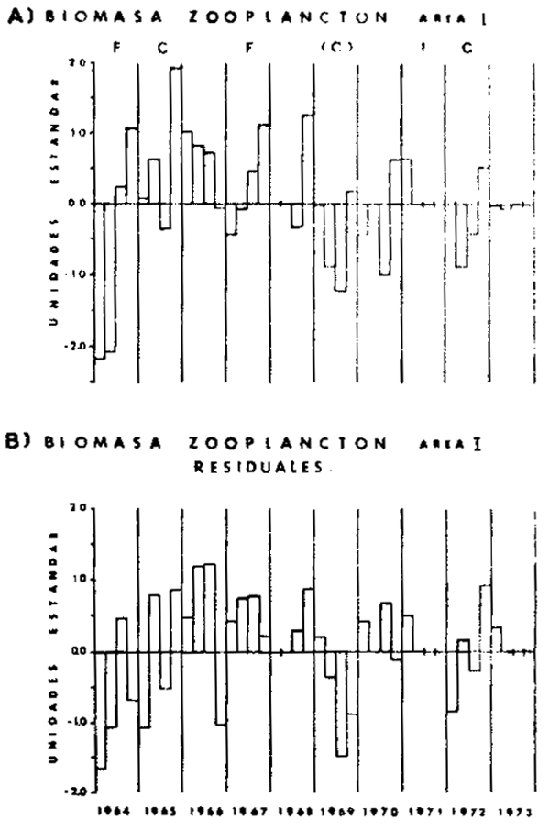 |
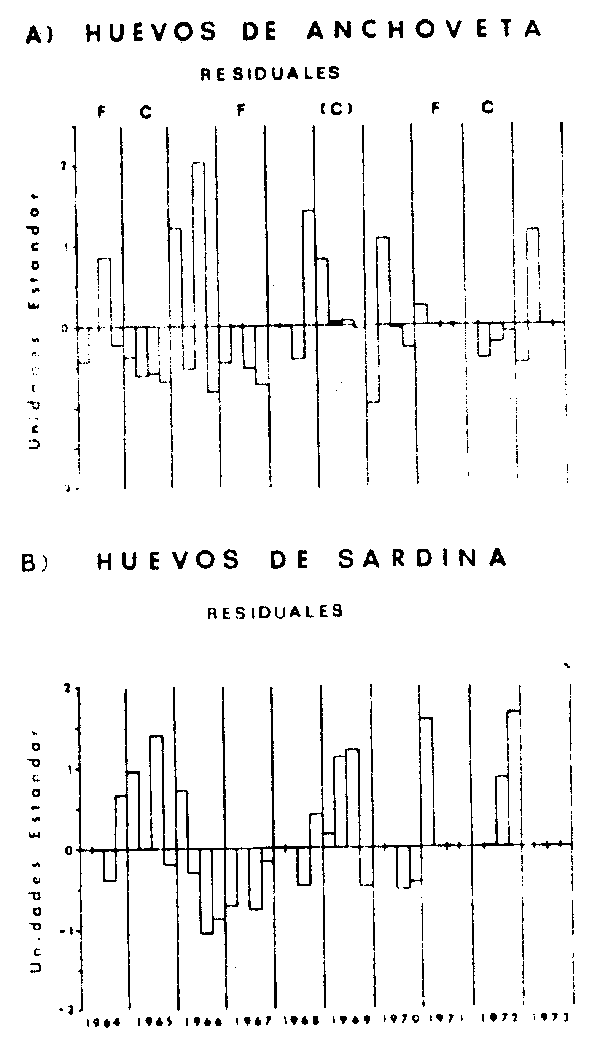 |
Fig. 18. - Series de tiempo de huevos de anchoveta
E. ringens y de sardina S. sagax área I.
A: Serie de estimadores trimestrales residuales de huevos de anchoveta representativos del componente de variacíon interanula.
B: Serie de estimadores trimestrales residuales de huevos de sardina representativos
del componenete de variación interanual.
Ambas series han sido estandar zizadas con
sus repecitvas desviaciones estádar.
Los períodos fríos cálidos estánindicados por las letras F y C repectivamente. |
La Tabla 3 compara las fracciones de la variabilidad total de las series de tiempo, asociadas al ciclo estacional y las series residuales. Estos porcentajes de varianza fueron calculados por diferencia enter las sumas de cuadrados de las series mensuales sin modificarlas (varianza total), menos la suma de cuadrados de las series mensuales de residuales (varianza internual). La diferencia de ambas sumas de cuadrados es precisamente el componente estacional.
| Estacional | Largo perfodo | N | |
|---|---|---|---|
| Biomasa cooplancton | 19.0% | 51.0 | 50 |
| Huevos Engraulis ringens | 29.7% | 70.3% | 54 |
| Huevos Sardinops sagax | 39.2% | 60.8% | 43 |
Los resultados de este análisis indican que la variabilidad total de la biomasa está distribuida en forma equitativa entre el componente estacional y el de largo período (o baja frecuencía). Este resultado contrasta notoriamente con la situacíon existente en la Corriente de California, donde en promedio sólo el 18% de la varianza total está asociada a la señal estacional. Debe tenerse en cuenta, como se comenta más adlante, que las distinatas metodologías utilizadas en ambos casos para la obtención de las muestras en terreno no son estrictamente comparables.
En las series de tiempo de abundancia de huevos, tanto de E. ringens como de S. sagax, sólo alredeor de un tercio de la varianza total está asociada al ciclo estacional y los dos tercios restantes a fluctuaciones de largo período interanuales. Esto permite sugerir que los stocks de peces pleágicos pudieran ser afectados en forma más directa que el zooplancton, por las alteraciones del régimen oceanográfico asociadas a los períodos fríos y cálidos.
Atendiendo a que de las tres series de tiempo una de ellas tiene unidades distintas, y a que entre las dos series de huevos, sus valores absolutos son muy diferentes, las series fueron estandarizadas respecto a sus propias varianzas. Con este fin, cada valor mensual fue escalado respecto a la desviación estándar mensual de la serie. Es así como estándar por sobre o bajo su promedio mensual.
La Figura 17A muestra la serie de tiempo de biomasa trimestrales y la Figura 17B la serie de residuales de biomasa. En 1964 se observan los valores más bajos de las series, coincidentes con un período anómalo frío (F en la Figura 17). Durante el período cálido de 1965 (El Niño), no se detecta una dismiñución de la biomasa en ambas series, por el contrario, estas presentan desviaciones positivas respecto al promedio. En general, se puede concluir que no existe una relación consistente con la respuesta esperada del ecosistema a las perturbaciones, es decir un aumento de la biomasa durante un período frío y una disminución durante un período cálido.
La información existenete presenta a lgunas limitaciones respecto a la representatividad de las muestras de zooplancton. La capa muestreada abarca sólo los 50m superficiales y por lo tanto, una importante fracción del zooplancton epipelágico ha quedado excluido del muestreo. La red de Hensen utilizada en arrastres verticales, aunque adecuada para censos de huevos del peces, no es particularmente apropiada para la captura de los miembros más móviles del zooplancton (por ejemplo, eufáusidos, quetognatos, e incluso algunos grupos de copépodos). Una diferencia important, entre los datos de California y los de Chile-Perú, es la extensión fuera de la costa del área muestreada. El estudio de la variabilidad biológica en California utilizó información colectada hasta 550 Km fuera de la costa, en cambio en Chile-Perú las áreas de acumulación abarcan sólo hasta 56 Km fuera de la costa. El carácter costero del muestreo en el Pacífico sur, lo hace poco adecuado para investigar fenómenos de gran escala, los que a juzgar por lo que se conoce de California, se originan en gran medida en respuesta a variaciones del flujo de todo el Sistema de Corrientes. Un muestreo biológico representativo con este fin, debiera por lo menos abarcar hasta el borde externo de la Corriente de Humboldt (ver Figura 3).
Sin embargo, estos sesgos de muestreo se mantuvieron constantes a lo largo de todo el período, y las conclusiones que se derivan son consistentes entre sí. Por lo tanto, cabe concluir que psoiblemente los cambios de abundancia de zooplancton, en la faja costera entre 18°y 24°S, no están estrechamente conectados a los efectos producidos por períodos anómalos fríos o cálidos.
La Figura 18 presenta las series de tiempo de los residuales de abundancia de huevos de anchoveta (18A) y sardina (18B). En ellas, el período cálido de 1965 muestra anomalías negativas en el caso de la anchoveta y positivas para la sardina. En 1969, otro período cálido, la anchoveta muestra sólo en el primer trimestre valores positivos significativos y el resto del año valores muy cercanos al promedio; en cambio la sardina vuelve a presentar anomalías positivas importantes. Durante 1972, el tercer período cálido de la serie, se vuelve a repetir el caso de 1965, con anomalías negativas de la anchoveta y positivas de la sardina. Es una observación relevante que esta relación inversa entre E. ringens y S. sagax se haya presentado en forma consistente en los tres períodos cálidos de la serie. Sin embargo, no existe una tendencia clara en la respuesta de cada especie, o de ambas, respecto a la secuencia de períodos fríos y cálidos. Por ejemplo 1967, un período frío contra lo esperado, presenta para ambas especies anomalías negativas.
La relación inversa en la intensidad del desove de ambas especies durante los períodos cálidos, es quizás el resultados más interesante de este análisis preliminar de los datos históricos de la región. Es evidente que esta respuesta inversa, no es suficiente por sí sólo para desencadenar una secuencia de reemplazo de una especie por otra, ya que de ser así, esto habría tenido lugar en 1965 o 1969. Al respecto debe recordarse que los residuales no representan abundancias absolutas, y que las abundancias de huevos de anchoveta fueron siempre mayores, durante el período. Un segundo aspecto a considerar, es que las series representan la variación del desove, es decir, solo la primera etapa del proceso que culmina en la génesis de una nueva clase anual en la población, y que en el mejor de los casos, el desove serviría para estimar el tamaño del stock desovante, pero en ningún caso es un buen índice del éxito de la clase anual siguiente. Precisamente, gran parte de los mecanismos tanto ambientales como biológicos, que determinan el éxito de una clase anual, actúan entre el desove y el reclutamiento de los juveniles al stock reproductor. Una comparación de estas series de tiempo con series de tiempo del reclutamiento en el mismo período es un objetivo altamente pertinente.
Es razonable esperar que los cambios en la dinámica y distribución de las masas de agua del Sistema en respuesta a condiciones anómalas, tengan consecuencias biológicas inmediatas respecto a las condiciones y estructura espacial del hábitat de las especies pelágicas.
Aunque hasta el momento no se conocen los mecanismos involucrados, es posible suponer que en el nécton tengan lugar desplazamientos poblacionales importantes, en respuesta a la redistribución de las masas de aguas. Por otra parte, las poblaciones planctónicas, debido a su menor capacidad de movimiento, pueden experimentar un deterioro de su hábitat localmente, con mortalidades naturales anómalamente elevadas. El desplazamiento de especies nectónicas, aunque pudiera significar el alivio de presiones ambientales negativas para las subpoblaciones adultas, puede significar el franco deterioro del ambiente para sus estadios larvales, en general mucho más sensibles a la calidad del ambíente. Salinidades, temperaturas, niveles de trubulencia y estratificación levemente distintos pueden ser letales para las larvas, ya sea en forma directa o alterando la disponibilidad (concentración) y calidad (composición y tamaño) del alimento larval. Elevadas mortalidades larvales, después de un desove exitoso, pueden ser igualmente efectivas para limitar el reclutamiento y éxito de la clase anual siguiente.
Agradecimientos
Deseamos expresar nuestros agradecimientos al Ingeniero Roberto Cabezas Bello, Director Ejecutivo y al Sr. Alfredo Sanhueza Seguel, Jefe de la División de Recursos de IFOP por las facilidades entregadas a los autores para la relización de este trabajo. La División de Investigación de la Sede Regional Talcahuano de la Pontificia Universidad Católica de Chile hizo posible el procesamiento de la información de biomasa y huevos y larvas de peces, otorgando el finaciamiento necesario. Agradecemos también a la Srta. Teresa Fierro, por su ayuda con las ilustraciones y a las Srtas. Liliana Lagos y Eliana Figueroa quienes dactilografiaron el manuscrito. La presentación de este trabajo en San José de Costa Rica ha side posible gracias al financiamiento generoso otorgado por la Comisión Oceanográfica Intergubernamental.
REFERENCIAS
Antezana-Jerez, T. 1978. Distribution of Euphausiids in the Chile-Perú Current with particular reference to the endemic Euphausia mucronata and the oxygen minima layer. Ph.D. Dissertation Scripps Institution of Oceanography, University of California San Diego.
Bakun, A. y R.H. Parrish. 1982. Turbulence, transport and pelagic fish in the California and Perú current system. CALCOFI Rep. (23):99–112.
Bernal, P.A. 1979. Large-scale biological events in teh California Current. CALCOFI Rep. (20):89– 101.
Bernal, P.A. 1980. Large-scale biological events in the California Current: the low-frequency response of the epipelagic ecosystem. Ph.D. Dissertation Scripps Institution of Oceanography, University of California San Diego.
Bernal, P.A. 1981. A review of the low-frequency response of the pelagic ecosystem in the California Current. CALCOFI Rep. (22):49–62.
Brockmann, C., E. Fahrbach, H. Huyer y R.L. Smith. 1980. The poleward undercurrent along the Perú coast: 5–15°. Deep-Sea Res. (27A):847–856. bl. Cañón, J.R.C. 1980. Prognosis del ambiente físcio de las pesquerías chilenas. Informe del Programa “Perspectivas de desarrollo de las pesquerías nacionales”, Corporación de Fomento de la Producción - Gerencia de Desarrollo. Instituto de Fomento Pesquero - Chile. Ref: AP 80– 3:152 pp.
Chelton, D.B. 1980. Low frequency sea level variability along the west coast of North America. Ph.D. Dissertation Scripps Institution of Oceanography, Univeristy of California San Diego.
Chelton, D.B. 1981. Interannual variability of the California Current: physical factors. CALCOFI Rep. (22):34–48.
Chelton, D.B., P.A. Bernal y J.A. McGowan. 1982. Large scale interannual physical and biological interaction in the California Current. J.Mar.Res. 40(4):1095–1125.
Enfield, D.B. 1981. Annual and nonseasonal variability of the monthly low-level wind fields over the Southeastern Tropical Pacific. Monthly Weather Rev. 109(10):2177–2190.
Enfield, D.B. y J.S. Allen. 1980. On the structure and dynamic of monthly mean sea level anomalies along the Pacific coast of North and South America. J.Physical Oceanogr. 10(4):557–578.
Johnson, D.R., T. Fonseca y H. Sievers. 1980. Upwelling in the Humboldt Coastal Current near Valparasío, Chile. J.Mar.Res. 38(1):–16.
Kramer, D. 1970. Distributional atlas of fish eggs and larvae in the California Current region: Pacific sardine, Sardinops caerulea (Girard), 1951 through 1966. CALCOFI Atlas No.12, vi pp, 277 charts.
Kramer, D. y E.H. Ahlstrom. 1968. Distributional atlas of fish larvae in the California Current region: northern anchovy. Engraulis mordax Girard, 1951 through 1965. CALCOFI Atlas No.9, xi pp, 269 charts.
McCreary, J. 1976. Eastern Tropical Ocean response to changing wind systems: with application to El Niño. J.Physical Oceanogr. 6:632–645.
McGowan, J.A. 1974. The nature of oceanic ecosystems. pp. 9–28. In The biology of the Oceanic Pacific, (C.B. Miller, ed.). Oregon State University Press, Corvallis.
Miller, F.R. y R.M. Laurs. 1975. The El Niño of 1972–1973 in the Eastern Tropical Pacific Ocean, Bull.Inter-Amer.Trop.Tuna Comm. 16(5):403–408.
Parrish, R.H. y A.D. MacCall. 1978. Climatic variation and exploitation in the Pacific mackerel fishery. California Dept. Fish and Game, Fish Bull. 167:1–110.
Quinn, W.H. 1974a. Monitoring and predicting El Niño invasions. J.Applied Meteorology. 13(7):825– 830.
Quinn, W.H. 1974b. Outlook for El Niño-type conditions in 1975. School of Oceanography Oregon State University, Corvallis (mimeo).
Robles, F.L. 1979. Water masses and circulation in the South-East Pacific and the El Niño event. Ph.D. Dissertation, Department of Oceanography, University of Wales, U.K. 2 vols:175 and 156 pp.
Robles, F.L., E. Alarcó n y A. Ulloa. 1980. Water masses in the northern Chilean zone and their variations in the cold period (1967) and warm periods (1969, 1971–73). pp. 83–174. In Proceedings of the Workshop on the Phenomenon known as El Niño. UNESCO, Paris.
Rojas, O. 1980. Estructura poblacional de la sardina española (Sardinops sagax musica) en la zona norte de Chile. En Informe del programa: “Investigaciones del recurso sardina española (Sardinops sagax)”, Corporación de Fomento a la producción-Gerencia de desarrollo. Instituto de Fomento Pesquero - Chile. Ref:AP 80–5:1–18.
Ryther, J.H., D.W. Menzel, E.M. Hulburt, C.J. Lorenzen y N. Corwin. 1971. The production and utilization of organic matter in the Peru Coastal Current. Investigaciónes Pesqueras (Barcelona). 35:43–60.
Santander, H. y O.S. de Castillo. 1977. Variaciones en al intensidad del desova de la sardina Sardinops sagax sagax (J.) en la costa peruana en los años 1966–1973. Bol.Inst.Mar Perú. 3(3):73–94.
Serra, J.R., M. Aguayo, O. Rojas, J.R. Cañón y F. Inostroza. 1979. Anchoveta Engraulis ringens Jenyns, Teleostomi, Clupeiformes, Engraulidae. En “Estado actual de las principales pesquerías nacionales. Peces.” Corporación de Fomento de la Producción-Gerencia de desarrollo. Instituto de Fomento Pesquero - Chile. Ref:AP 79–78:52 pp.
Silva, N. y H. Sievers. 1981. Masas de agua y circulació n en la región de la rama costera de la Corriente de Humboldt, latitudes 18°S – 33°S (Operación Oceanográfica MARCHILE 10-ERFEN l). Ciencia y Tecnología Marinas, Comité Oceanográfico Nacional (Chile) 5:5–50.
Simpson, J.G. y R. Buzeta. 1967. El crecimiento y la edad de la anchoveta (Engraulis ringens, Jenyns) en Chile basado en estudios de frecuencia de longitudes. Bol.Cient.Inst.Form.Pesq., Santiago-Chile. 3:1–53.
Simpson, J.G. y E. Gil. 1967. Maduración y desove de la anchoveta (Engraulis ringens) en Chile. Bol.Cient.Inst.Fom.Pesq., Sanitago-Chile. 4:1–55.
SIO-WHOI-MIT. 1969. Physical and chemical data from the Scorpio Expedition in the South Pacific Ocean. USNS Eltanin, Cruises 28 and 29, 1967. Scripps Institution of Oceanography - Woods Hole Oceanographic Institution and Massachussetts Institute of Technology. SIO Reference 69– 15, WHOI Reference 69–56, 89 pp.
Trautman, S. y M. Aguyao. 1980. Edad y crecimiento de sardina española (Sardinops sagax musica). endash Informe del Programa: “Investigaciones del recurso sardina española (Sardinops sagax)”, Corporacion del Fomento a la Producción-Gerencia de desarrollo. Inst.Fom.Pesq. Chile. Ref: AP 80–5:1–24.
Wyrtki, K. 1967. Circulation and water masses in the Eastern Equatorial Pacific Ocean. International Journal Oceanology and Limnology. 1(12):117–147.
Zuta, S., D.B. Enfield, J. Validivia, P. Lagos y C. Blandin. 1980. Physical aspects of the 1972–73 El Niño phenomenon. pp. 11–62. In Proceedings of the Workshop on the Phenomenn known as El Niño. UNESCO, Paris.
Por
P.A. Bernal
BIOTECMAR
Pontificia Universidad Católica de Chile
Sede Regional Talcahuano
Casilla 127 - Talcahuano, Chile
y
D.B. Chelton
Jet Propulsion Laboratory
California Institute of Technology
Pasadena, California 91109
USA
Summary
The pelagic ecosystems of the California Current shows biological responses dominated by large scale, low frequency changes. These fluctuations are not associated with the coastal water movements. On the contrary, this study shows that they are strongly associated with inter—annual fluctuations of the transport of the California Current.
Zooplankton biomass time series were strongly correlated with the temperature field ( r= 0.77; p<0.05). This indicated that horizontal and vertical advection play primary roles in these changes. The biomass fluctuations are most highly correlated with current transport five months prior to the changes (r=0.77; p<0.05). At the same time, these changes are associated with fluctuations of thermal anomalies in the eastern tropical Pacific indicative of “El Niño” events (r=0.74; p<0.05), with a 4 month lag.
Density independent factors controlling populations in eastern boundary currents are discussed, emphasizing the importance of incorporating parameter estimations for the oceanographic fluctuations in fishery management models. The relation between anomalous changes in California Current transport and sea level observation time series from coastal tide—gauge measurements. For the northern hemisphere there is a highly significant relation with rising coastal sea level preceding the transport anomalies (r=0.83; p<0.05).
INTRODUCCION
Las corrientes de márgen oriental de los giros subtropicales, han side reconocidas desde temprano, como entre las más productivas de los océanos. Los niveles de producción primaria observados en ellas, son uno o dos órdenes de magnitud superiores a los observados en otros ecosistemas oceánicos. Por ejemplo, en Perú Ryther et al., (1971) reportan niveles de 11.7 gr—C m-2 d-1, en cambio para el ecosistema del giro central del Pacífico Norte, Venrick et al. (1973) citan valores típicos entre 0.1 a 0.2 gr—C m-2d-1. Esta elevada producción biológica se debe a las altas tasas de incorporación de nutrientes inorgánicos (fosfatos y nitratos) a las capas superficiales produciendo altos niveles de fitopláncton. Como el fitoplancton es la fuente alimenticia principal del zooplancton herbívoro, y este a su vez sirve de alimento para muchos peces pelágicos, existe un correspondiente dealta productividad en los niveles superiores de la cadena trófica.
En contraste con los ecosistemas oceánicos localizados en los giros centrales de los océanos, los ecosistemas de los sistemas de márgen oriental, además de estar abiertos a flujos de masa y energía canalizados directamente hacia la producción orgánica, están también abiertos a la entrada a gran escala de energía mecánica en la forma de advección horizontal y del forzamiento local de los vientos. Estos sistemas se caracterizan biológicamente, tanto por presentar una elevada heterogeneidad espacial de las poblaciones planctónicas residentes (Haury, 1976), como por exhibir una predictibilidad temporal reducida (Hayward y McGowan, 1979).
Para estos sistemas, se ha propuesto que a causa de los efectos disruptivos de las condiciones físicas en estas regiones, los mecanismos clásicos de interacción biológica (competencia y predacción), pueden no tener la oportunidad de jugar un rol dominante en el control de la composición de especies, ni en la modulación de sus cambios de abundancia (McGowan, 1974). Estudios de las fluctuaciones de biomasa planctónica en la Corriente de California entre 1955 y 1959, proveen evidencias en apoyo de esta hipótesis. Por ejemplo, la biomasa de copépodos, que en promedio corresponde a un tercio de la biomasa total, entre Octubre de 1955 y Octubre de 1958 disminuyó un orden de magnitud desde 44.8 gm / 1000 m3 a 4.1 gm / 1000 m3. Se ha calculado que este cambio es equivalente a un flujo de energia de 4 x 1012 watts en la Corriente de California. Colebrook (1977) utilizando análisis de componentes principales, demostró cambios de abundancia sorprendentemente coherentes tanto entre los 17 taxa analizados, como entre diferentes localidades. Estudios de sedimentos finamente estratificados provenientes de fosas anaeróbicas frente a las costas de California, indican que grandes fluctuaciones en las abundancias de peces y componentes del zooplancton, han tenido lugar en forma regular durante los últimos 200 años (Soutar e Isaacs, 1969 y 1974).
Aunque el aumento en número de algunos taxa a expensas de otros podría ser explicada en base a interacciones biológicas, dicha explicación se hace insostenible cuando la mayoría de los taxa tiende a aumentar (o disminuír) simultáneamente en gran parte del ecosistema. Considerando que las escalas temporales de los fenómenos demográficos relevantes para el fito y zooplancton son mucho más cortas que la escala de variación típica de los cambios observados en los sedimentos; y que Colebrook en su estudio documentó el cambio simultáneo en la abundancia de 17 taxa distintos; es razonablemente concluir que las condiciones ambientales de gran escala juegan un rol mucho más significativo que la competencia y predación en el control de la biomasa en la Corriente de California.
Para las corrientes de márgen oriental generalmente se acepta que el proceso principal responsable de la incorporación de nutrientes inorgánicos de origen no—regenerativo (y por lo tanto responsable de la “producción nueva”) es la surgencia costera. Si las surgencias costeras fueran responsables de la producción primaria, e indirectamente de la producción del zooplanton en estos sistemas, se debiera esperar que el máximo de biomasa observado se encontrará adyacente a la línea de costa, o por lo menos dentro de un radio de deformación de Rossby. Dos evidencias contradicen esta suposición. Primero, la zona enriquecida biológicamente en California es un orden de magnitud más ancha que un radio de deformación de Rossby. Segundo, el máximo de biomasa zooplanctónica observado se ubica entre 100–150 Km de la costa (figura 1) lo que se ve confirmado por la distribución de biomasa observada en cortes frente a San Francisco (figura 2a) y Los Angeles (figura 2b). Aunque las surgencias costeras indudablemente juegan un papel importante en forma local, es evidente que otros procesos deben ser responsables, a lo menos en parte, de las características biológicas de gran escala.
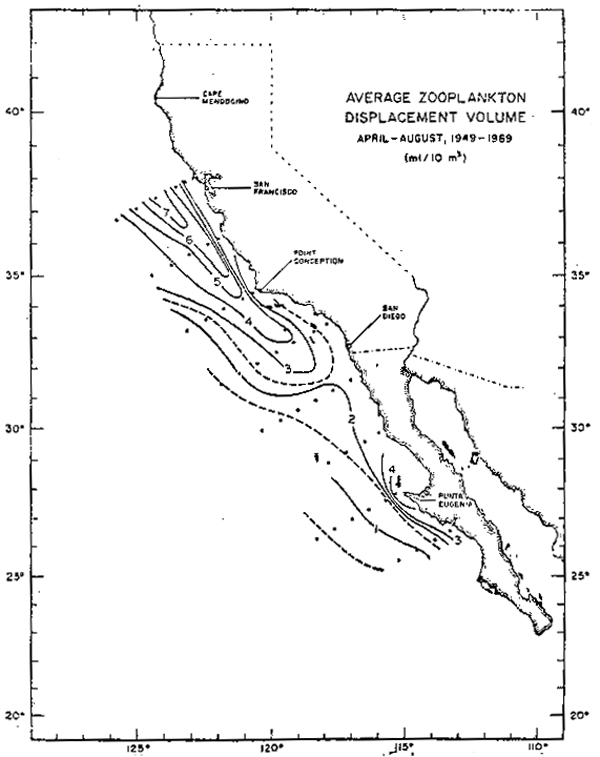
Fig. 1.- Distubución de biomasa zooplanctónica en la Corriente de California. Promedios 1979–1978.
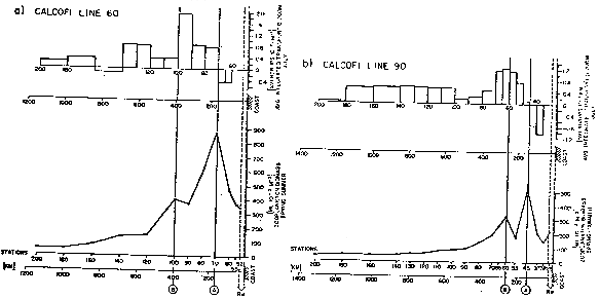
Fig. 2. (a) Transporte geostrófico y promedios de biomasa zooplanctónica. Línea CalCOFI 60
y (b) Transporte geostrófico y promedios de biomasa zooplanctónica. Línea CalCOFI 90.
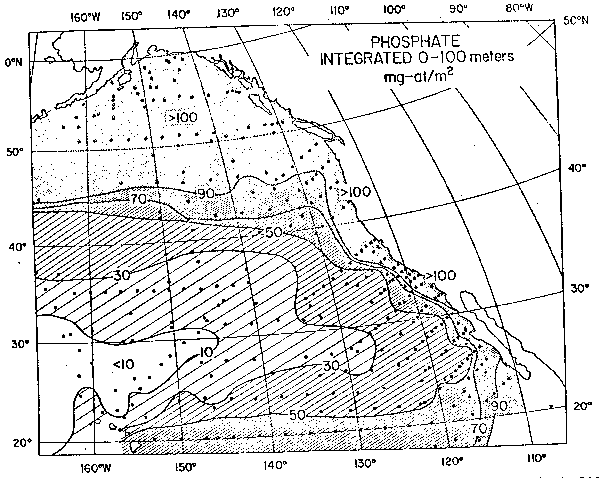
Fig. 3. - Distribución de fosfatos en el Pacífico Norte. Valores integrados desde 100 m hasta la superficie. Unidades mg—at/m2.
Tanto en el hemisferio Norte como en el Sur, los ecosistemas de márgen oriental se encuentran “corriente abajo” de sistemas oceánicos con altos niveles de nutrientes. La figura 3 muestra la distribución de fosfatos en el Pacífico Norte. La penetración hacia el sur de extensiones tanto en la distribución de biomasa como en la de fosfatos, enfatiza la importancia que tiene la advección horizontal de gran escala para la producción de estos sistemas. Reid (1962) propuso por primera vez la existencia de una relación entre advección y zooplanton en California. Wickett (1967) encontró una relación entre la biomasa de zooplancton en California y el forzamiento de los vientos superficiales en el giro de Alaska un año antes. Wickett sugirió que el zooplancton estaría respondiendo al mayor contenido de nutrientes de las aguas de la divergencia de Alaska que habrían sido transportadas por advección hacia el sur. Bernal (1980) y Bernal y McGowan (1981) presentaron evidencia relacionado el transporte geostrófico de origen subártico con la abundancia de zooplancton.
El próposito de esta presentación es dar cuenta de los resultados de diversas investigaciones que demuestran la existencia de fluctuaciones de baja frecuencia en la respuesta del ecosistema pelágico, en la Corriente de California, y examinar la relación que existe entre estas variaciones y las variaciones del medio ambiente físico.
CARACTERIZACION DE LA VARIABILIDAD BIOLOGICA INTERANUAL EN LA CORRIENTE DE CALIFORNIA
Con el fin de analizar la variabilidad biológica en la Corriente de California, a partir de la base de datos acumulados entre 1949 y 1969 por el programa CalCOFI (California Cooperative Oceanic Fisheries Investigations) se derivaron cuatro series de tiempo de la biomasa de macrozooplancton. Los datos originales provienen de arrastres oblícuos de plancton en los 140 m superficiales con una red de 0.78 m2 de boca equipada con una malla filtrante de 5 m de largo y 0.505mm de trama. La figura 4 muestra la distribución de las estaciones y los límites de las áreas geográficas de acumulación estadística. En cada área se utilizó como estimador mensual de la biomasa, el promedio de los volúmenes de plancton previamente loge transformados. En la figura 4 7nacute; representa el número típico de observaciones incluidas en los promedios mensuales.
La Tabla l muestra una partición de la varianza total de las series en un componente estacional y uno residual de largo período. La estacionalidad da cuenta de un máximo de 30% de la variabilidad en el área I, y de un mínimo de 8.4% en el área III.
| ESTACIONAL | LARGO PERIOD | |
|---|---|---|
| AREA I | 30.0 | 70.0 |
| AREA II | 22.7 | 77.3 |
| AREA III | 8.4 | 91.6 |
| AREA IV | 13.6 | 86.4 |
Como el interés fundamental en este estudio son las variaciones de largo período, y atendiendo que el componente estacional representa en promedio alrededor del 18% de la varianza total, se procedió, a eliminar la estacionalidad de las series. Por esto, cada valor observado se redefinió como una desviación respecto a su promedio mensual calculado sobre todo el período de estudio. Las series presentan distintas varianzas totales en términos absolutos, por esto con el fin de facilitar las intercomparaciones, cada valor fue también escalado respecto a la desviación estándar del área respectiva. Este tratamiento de los datos hace que los elementos mensuales que componen las series representen anomalías de biomasa con promedio O y desviación estándar l. En la figura 5 se muestran las series para las cuatro áreas. Una simple inspección de los patrones de variación muestra que existe co-variación entre las áreas y que algunos eventos son comunes a dos o más de ellas.
La escala de tiempo característica de estas series es larga. La figura 6 muestra las funciones de autocorrelación calculadas para las cuatro áreas. La área I muestra la escala de tiempo más corta, 14 meses, y la área IV la más larga, 24 meses. Las áreas II y III muestran escalas de tiempo intermedias de 18 meses. En comparación es mucho más corta, alrededor característica del forzamiento de los vientos sobre la región es mucho más corta, alrededor de l mes.
Del cómputo del espectro de varianza para las cuatro series, resulta aparente que gran parte de la varianza total está asociada a fluctuaciones en la banda de baja frecuencia. La coherencia entre estos espectra es también significativa en las bajas frecuencias (Bernal, 1979). Los porcentajes de varianza asociados a distintas bandas espectrales indican que alrededor de un 50% de la varianza total está asociado a fluctuaciones de frecuencias menores a 0.05 ciclos mes-1, lo que equivale a decir que las fluctuaciones dominantes en las series presentan períodos característicos de 20 o más meses. Estos resultados contrastan notoriamente con las hipótesis propuestas por Walsh (1977) para el espectro de forzamiento de la respuesta biológica en ecosistemas de margen oriental. Este autor propone la existencia de un amplio máximo espectral entre 0.016 y 0.060 ciclos día-1 para el zooplancton, sin embargo, en California el zooplancton estaría respondiendo a un forzamiento con un máximo espectral en frecuencias menores a 0.0016 ciclos día-1, un orden de magnitud inferior a lo propuesto.
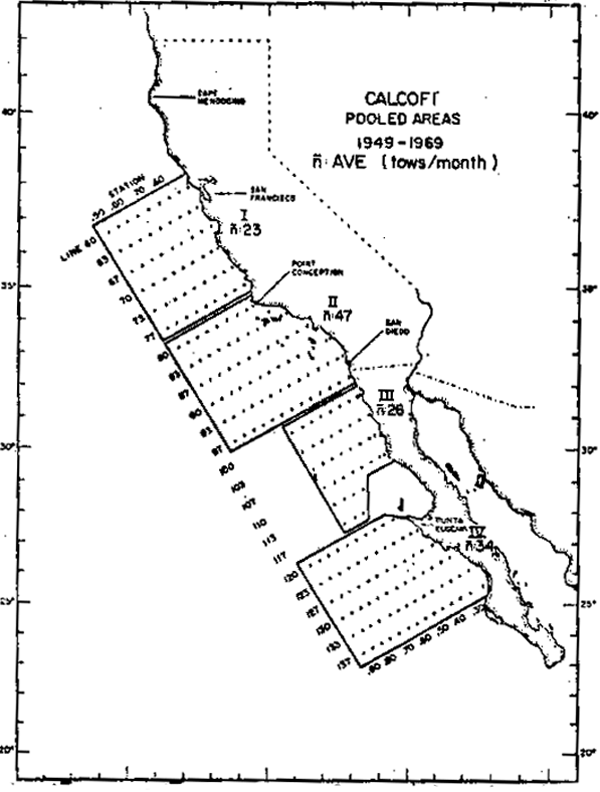
Fig. 4.- Areas de acumulación estadística para las series de tiempo de biomasa zooplanctónica “n” indica el número tipico de observaciones mensuales incluidas en un promedio.
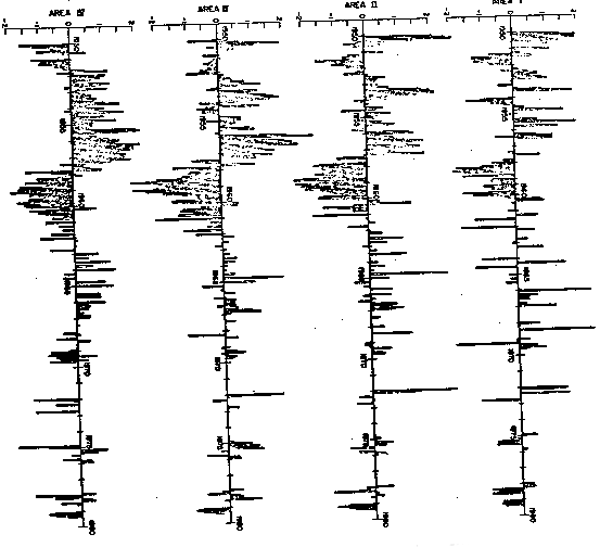
Fig. 5.- Series de tiempo de biomasa zooplanctónica para las cuatro áreas de acumulación. Cada valor es la desviación del promedio mensual respectivo escalado por su correspondiente desviación estándar. Cada serie tiene promedio cero y desviación estándar unitaria.
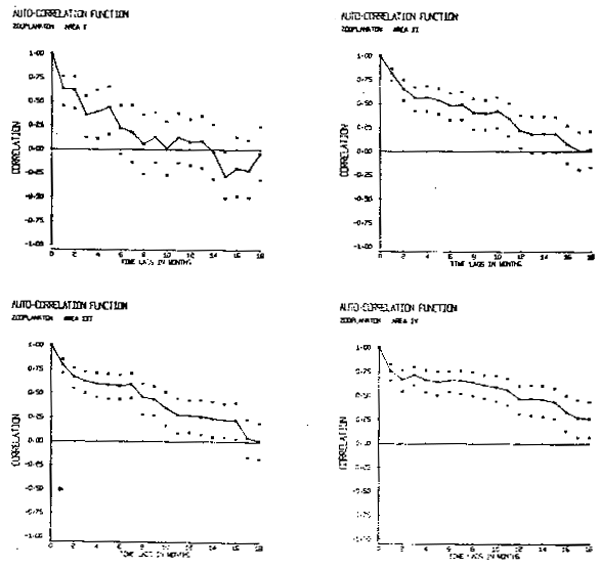 |
Fig. 6.- Funciones de autocorrelación de biomasa zooplanctónica. |
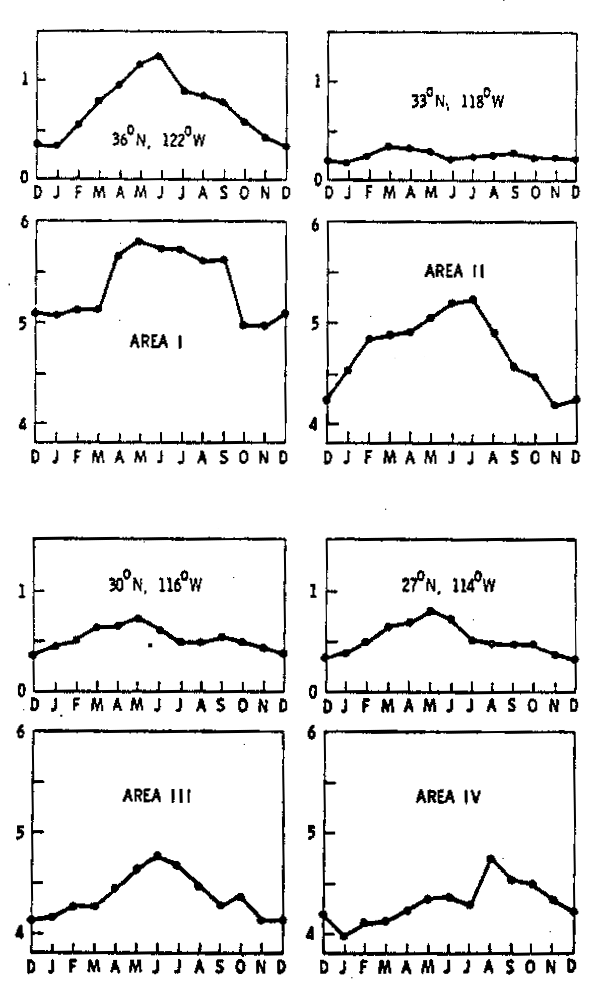 |
Fig. 7.- Ciclos estacionales de biomasa zooplanctónica (paneles inferiores) y del componente del stress del viento paralelo a la costa en dirección al ecuador en dinas cm-2 (paneles superiores), correspondientes a las cuatro áreas de acumulación. |
INTERACCION FISICA Y BIOLOGICA INTERANUAL DE GRAN ESCALA EN LA CORRIENTE DE CALIFORNIA
Antes de referirnos a la variabilidad interanual, discutiremos brevemente la variación estacional. La figura 7 muestra los promedios mensuales de biomasa (paneles inferiores) junto al ciclo estacional del componente paralelo a la costa del stress del viento en dirección al ecuador (Nelson 1977). Estos valores del stress del viento pueden ser considerados como un índice de surgencias costeras, con un aumento en su magnitud correspondiendo a un incremento de las surgencias. En los datos de biomasa existe un claro gradiente latitudinal de norte a sur, con valores más elevados en el norte. Por otra parte, como se mostró en la Tabla 1. la estacionalidad en las dos áreas norte es más pronunciada siendo responsable por 30 y 23% de la varianza total respectivamente.
Al inspeccionar la relación entre ambas variables, ocasionalmente se observa una buena correspondencia. Por ejemplo, un máximo del stress del viento durante Mayo en el área III, es seguido de un máximo de biomasa en Junio; y nuevamente un máximo del stress en Septiembre, es seguido un mes más trade por otro máximo en la biomasa. De aquí podría concluirse que existe una relación estrecha entre ambos procesos. Sin embargo, esta relación no se mantiene para las otras áreas. En todo caso, explorar relaciones causales entre ciclos estacionales es un ejercicio estadísticamente arriesgado por dos razones. En primer lugar el limitado número de grados de libertad (12), con variables generalmente fuertemente autocorrelacionadas, hace que el número efectivo disponible para las comparaciones sea aún menor. En segundo lugar, casi todas las series geofísicas y biológicas presentan estacionalidad, y permitiéndose cierta libertad con los desfasamientos entre variables, es siempre posible encontrar una correlación significativa. Un análisis de correlación entre el stress paralelo a la costa considerado como índice de surgencia y la biomasa de zooplancton, demostró que todos los valores de correlación en las cuatro áreas no son significativos para desfasamientos de 0 a 24 meses, obteniéndose valores menores a 0.25 en todos los casos. Es legítimo concluir entonces que las surgencias costeras no constituyen el proceso dominante que controla la variación interanual de la respuesta biológica en las corrientes de margen oriental.
Una interpretación conservadora de estos resultados, nos permite suponer la existencia de otros procesos que explique en parte la variabilidad biológica observada en las series. En el resto de este trabajo, discutiremos exclusivamente residuales no-estacionales de las variables.
La contraparte física de las series de biomasa en California, series de temperatura, salinidad y transporte, se obtuvieron en base a datos colectados en las estaciones que se muestran en la figura 8. Estas estaciones son aquellas que fueron ocupadas en mayor frecuencia temporal.
Atendiendo a la fuerte co-variación de las series de biomasa, y con el fin de generar una serie única representativa para toda la región, se calculó un promedio entre las series de las cuatro áreas. Este promedio toma en cuenta, por medio de un procedimiento estadístico objectivo, la presencia de vacíos en las series, (detalles del método se entregan en: Chelton, Bernal y McGowan, (1982). Las correlaciones entre esta serie única y las series por área son 0.73, 0.89, y 0.73 para las áreas I, II, III y IV respectivamente. Esta serie única se muestra en la figura 9a, junto a series de temperatura (9b), salinidad (9c) y transporte (9d). Las series de temperatura y salinidad representan valores mensuales, promediados sobre toda el área, de las observaciones a 10 m de profundidad. Chelton (1981) demostró que el campo de temperatura es altamente coherente sobre toda la región de la Corriente de California, y que el de salinidad, aunque más ruidoso, tiende a aumentar y disminuir en el mismo sentido sobre toda la región.
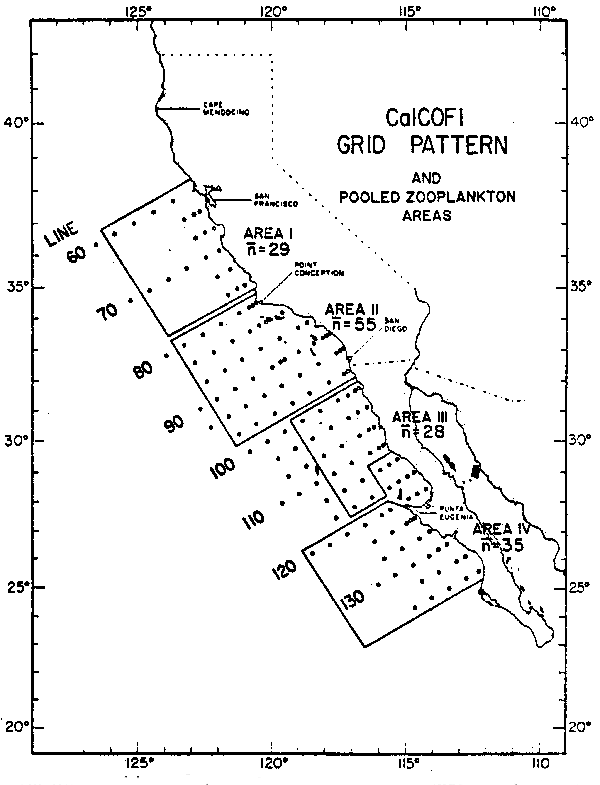
Fig. 8.- Red de estaciones hidrográficas frecuentemente ocupadas en el Programa CalCOFI: Los círculos abiertos representan estaciones sobre profundidades de menos de 500 m. Las áreas de acumulación para la biomasa zooplanctónica aparecen como cajas con línea contínua.
La información de transporte, fue tratada en forma diferente. Primero se calculó el campo de alturas dinámicas integradas sobre 500 dB y luego se llevó a cabo una descomposición ortogonal de la matriz de autocovarianza de este campo. Este procedimiento, conocido como Funciones Ortogonales Empíricas (F.O.E.), busca definir patrones espaciales invariantes que den cuenta de una alta proporción de la varianza temporal total del campo observada durante el período en estudio. Cada observavión mensual del campo (un mapa de alturas dinámicas) es descompuesto en un número limitado de componentes, (los patrones espaciales invariantes) ponderados por coeficientes temporales proporcionales a la importancia que cada patrón muestra en el mes en cuestión. La figura 10 muestra el patrón invariante asociado a la primera F.O.E. y en el panel d de la figura 9 se presenta la variación temporal del mísmo, considerado como transporte anómalo de gran escala. Superpuesta a la serie de transportes, se ha incluido la serie biológica representada por triángulos. Resulta evidente que la respuesta biológica del sistema está estrechamente asociada a estas variaciones del transporte de la corriente.
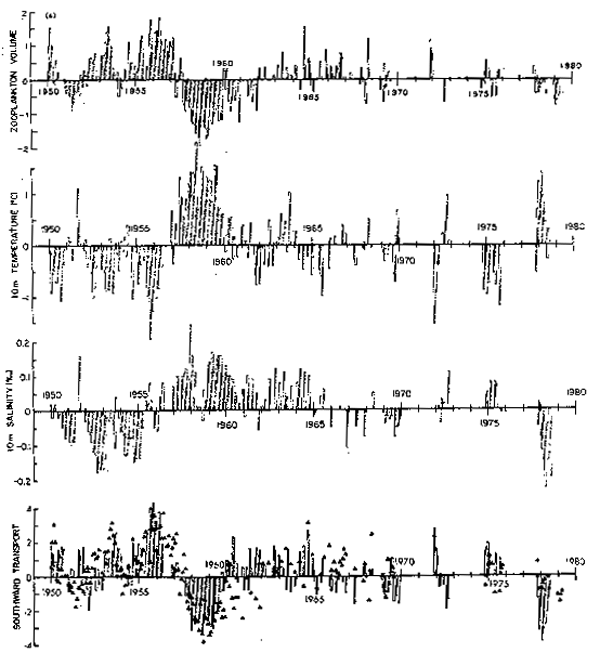
Fig. 9.- Series de tiempo de residuales no estacionales de cuatro parámetros en la Corriente de California: a) Serie única de residuales de biomasa de zooplancton (promedio de las cuatro áreas); b) temperatura promedio a 10 m sobre 150 estaciones hidrográficas; d) serie de tiempo de los coeficientes de amplitud de la primera Función Ortogonal Empírica de alturas dinámicas 0/500 decibars, los triángulos representan la serie de biomasa zooplanctónica 9a.
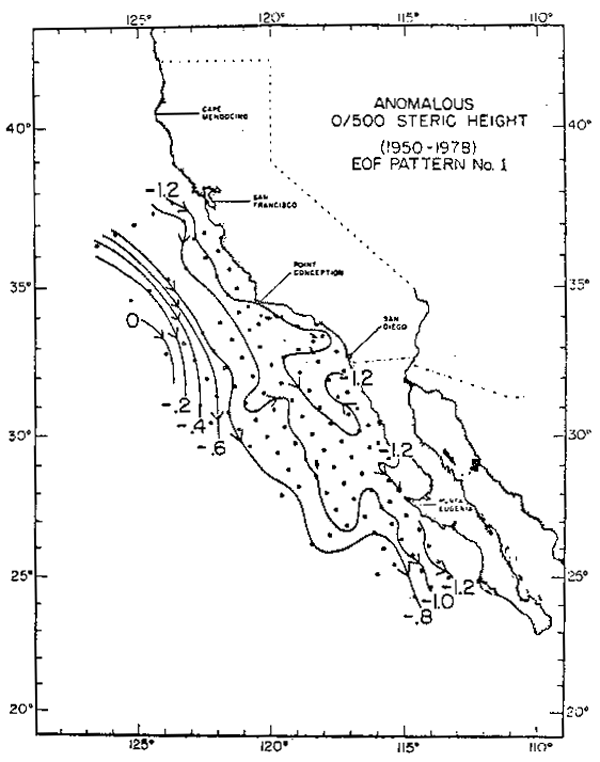
Fig. 10.- Patrón espacial invariante representativo de la primera función Ortogonal Empírica de alturas dinámicas. Las flechas indican la dirección del flujo cuando la serie de tiempo de los coeficientes de amplitud es positiva.
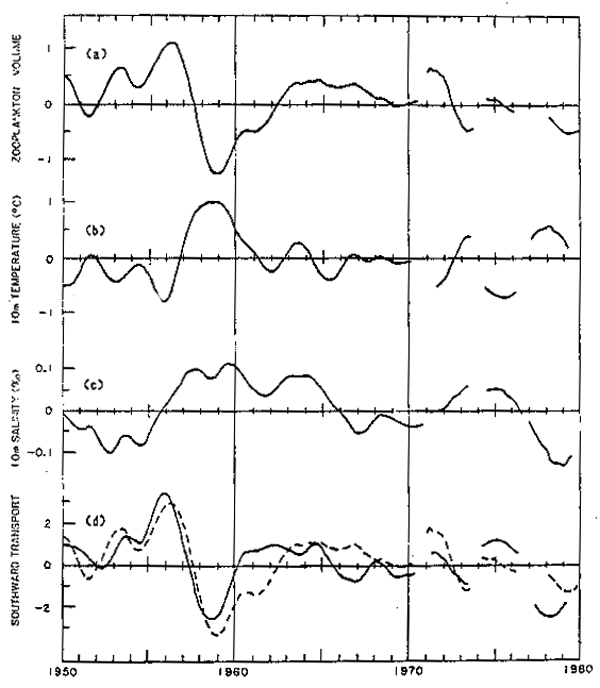
Fig. 11. - Componente de variación interanual de las cuatro series de tiempo de la Figura 9. La línea punteada en lld, representa la serie de biomasa zooplanctónica.
Una representación más compacta de la variabilidad de estas series, se muestra en la figura ll, donde se ha dejado solamente el componente interanual de la variabilidad al computarse el promedio móvil de 13 meses sobre los datos de la figura anterior. También en este caso se consideró objetivamente, la presencia de vacíos en las series.
De la comparación de estas series, resulta evidente que existe una relación inversa entre zooplancton y la temperatura. La correlación máxima es -0.77, con la temperatura precediendo al zooplancton por 6 meses. Valores por sobre o debajo de 0.33 son significativos con un límite de confianza del 95%. Esta asociación aunque importante en sí misma, no permite separar los dos procesos que pueden establecer un vínculo causal entre ambas series: la advección horizontal y la advección vertical. Ambos procesos físicos están estrechamente ligados, ya que un aumento en el flujo de la Corriente de California resulta en un incremento de la advección de aguas frías de origen subártico, pero también provoca un ajuste geostrófico del campo de densidad que resulta en un levantamiento de las isotermas y enfría la zona costera mediante advección vertical.
Estos procesos podrían ser distinguidos mediante las variaciones del campo de salinidad, ya que la distribución zonal y meridional de esta variable en la Corriente de California, hace que la advección horizontal disminuya la salinidad superficial y que la advección vertical la aumente. Como se mencionó, el campo de salinidad es más ruidoso y más heterogéneo en el espacio que el de temperatura. La variación interanual de la salinidad es muy pequeña (desviación estándar de 0.06 o/oo) y la relación entre temperatura y salinidad que resulta es débil. La máxima correlación observada es de 0.36, valor que es significativo sólo con un intervalo de confianza del 75%. De estos dos análisis es posible concluir que tanto la advección horizontal como la vertical juegan un rol en el control de las fluctuaciones interanuales del zooplancton. Debe recalcarse que la advección vertical a la que nos referimos, no se trata de surgencias costeras inducidas localmente por el vient sino que es producto del reajuste geostrófico del campo de densidad, es decir, por un cambio en la inclinación de las isopicnas en respuesta a cambios de gran escala y baja frecuencia en el transporte de la Corriente de California.
Son precisamente las fluctuaciones dominantes del flujo de gran escala las que estan siendo medidas por la primera función ortogonal empírica en la figura lld. Esta figura presta un apoyo contundente a la hipótesis que el aumento del transporte hacia el ecuador está asociado con un aumento de la biomasa zooplanctónica y que una disminución de este transporte produce el efecto inverso.
La función de correlación desfasada entre estas series es claramente asimétrica indicando que el zooplancton está mas fuertemente correlacionado con el flujo anómalo cuando este lo precede. Se observa una correlación máxima de 0.77 cuando las variaciones del transporte preceden al zooplancton por 5 meses. Para esta comparación, valores a 0.35 son significativos con un límite de confianza del 95%. Este es un resultado extraordinariamente bueno si se consideran los serios problemas del muestreo presentes en la base de datos originales
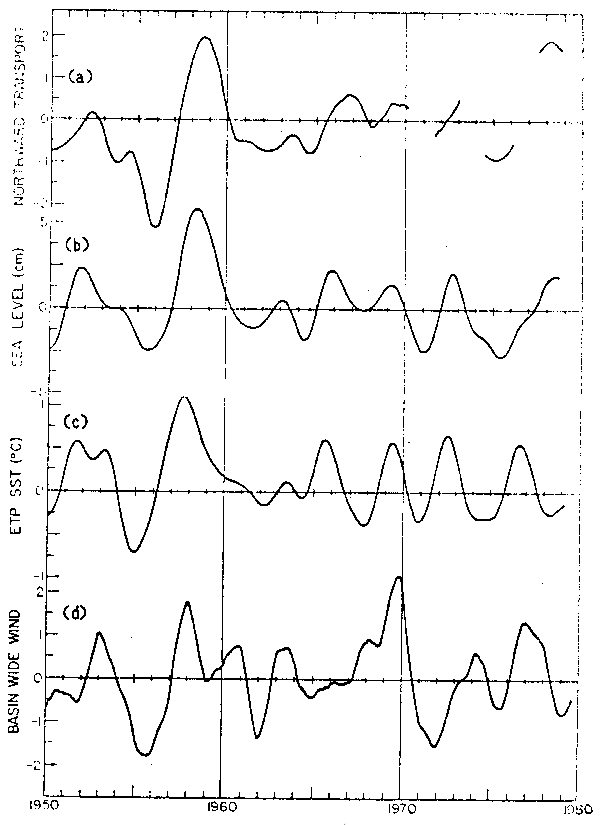
Fig. 12. - Series de tiempo de la variación de baja frecuencia, calculada por la doble aplicación de promedios móviles de 13 meses. a) índice de transporte hacia el norte; b) el nivel del mar promedio para California calculado en base a datos de San Francisco, Los Angeles y San Diego; c) anomalías de la temperatura superficial del mar en el Pacífico Ecuatorial Oriental indicativas de El Niño; d) Serie de los coeficientes de amplitud del patrón invariante dominante de los vientos sobre el Pacífico norte (Fig. 13).
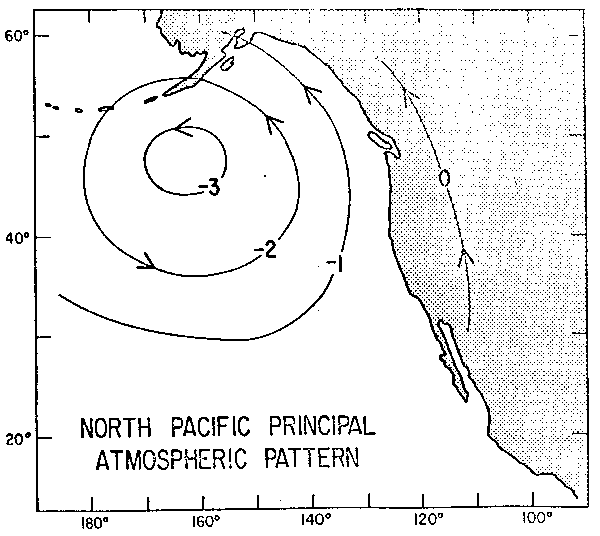
Fig. 13. - El patrón principal de variación de los vientos sobre el Pacífico Norte. Las flechas indican la dirección del flujo geostrófico cuando las amplitudes se consideran como positivas, (de: Davis, 1978).
Con el propósito de buscar un método que permita monitorear estas fluctuaciones de gran escala, se exploró la relación entre el flujo anómalo de la corriente con las alturas del nivel del mar registradas en mareógrafos costeros. La figura 12b presenta series de nivel del mar suavizadas con un doble promedio móvil de 13 meses para San Francisco, Los Angeles y San Diego. Los valores han sido corregidos respecto a los efectos barométricos inversos de la presión atmosférica. La figura 12a muestra las variaciones de flujo anómalo hacia el norte (la misma serie referida al patrón invariante con signo cambiado). Es aparente que existe una estrecha relación entre el flujo de la Corriente de California y el nivel del mar en la costa. Niveles del mar elevados se corresponden con flujo anómalo en dirección al norte y bajos niveles del mar indican flujos hacia el ecuador mayores que el promedio. La correlación máxima entre estas series es de 0.83, con valores mayores a 0.30 significativos al 95%. Esta máxima asociación se observa cuando la elevación costera del nivel del mar precede al transporte por 3 meses. Obviamente, de acuerdo a los resultados previos, las series de nivel del mar medido en la costa están también estrechamente asociadas a la variabilidad de gran escala del zooplancton, con una correlación máxima de -0.70 cuando el nivel del mar precede al zooplancton por 7 meses (significativo al 95%).
La causa de las variaciones de gran escala en el flujo de la corriente de California no está claramente establecida. Sin embargo, se observa una estrecha asociación entre estas fluctuaciones y los fenómenos de El Niño. En la figura 12c se muestran las anomalías térmicas en el Pacífico Ecuatorial Oriental (0° – 10°S; 80° – 100°W). La correlación máxima entre ambas series es de 0.74, con valores mayores a 0.39 significativos al 95% cuando anomalías positivas en el Pacífico Ecuatorial Oriental preceden las anomalías del flujo en California por 3–4 meses. Chelton y Davis (1982) demostraron que el intervalo de tiempo por el cual las variaciones térmicas del Pacífico Ecuatorial Oriental, preceden los cambios de nivel del mar a lo largo de la costa occidental de Norteamérica, aumentan monotónicamente a mayor distancia de los trópicos. Esta relación permite derivar una velocidad de propagación de fase para la perturbación de40cm, seg-1. Esta observación es consistente con modelos de El Niño que proponen que esta perturbación se propage en la forma de ondas costeras atrapadas o de Kelvin (McCreary, 1976), aunque la velocidad de propagación es sólo un tercio de lo esperado. En dicho estudio también se observó que la zona de flujo geostrófico oceánico asociado, es mucho más ancha que la predicción del modelo, lo que permite sugerir la presencia de ondas de Rossby propagándose hacia el oeste. Esta relación entre las anomalías térmicas del Pacífico Ecuatorial Oriental y el flujo anómalo de la Corriente de California, sin embargo, no da cuenta de algunos períodos anómalos, por ejemplo 1953, 1967–68, 1976 y 1978.
En la figura 12d se muestra la serie de tiempo de la variación de gran escala de los vientos sobre el Pacífico Norte. Estas son estimaciones cuasi-geostróficas de los vientos, a escala de toda la cuenca. El patrón espacial dominante de los vientos se muestra en la figura 13, donde los valores negativos indican circulación anticiclónica. Sorprendentemente se observa una correlación máxima de 0.43, cuando el nivel del mar precede a los vientos por tres meses.
Esta inesperada relación puede comenzar a entenderse al considerar la correlación entre las anomalías térmicas del Pacífico Tropical Oriental en relación a los vientos. En este caso la temperatura en el Pacífico Ecuatorial Oriental precede a los vientos por seis meses, mostrando una correlación de 0.48. Esta relación pone de relive la importancia de las teleconecciones que se dan en la atmósfera entre procesos que tienen lugar en la banda ecuatorial y en latitudes intermedias, es decir, los procesos transientes que ocurren en la atmósfera por la alternancia del dominio de las circulaciones de Walker y Hadley y que parecieran tener un rol dominante en la génesis de perturbaciones de El Niño.
REFERENCIAS
Bernal, P.A. 1979. Large-scale biological events in the California Current. Calif.Coop.Oceanic Fish.Inv.Rep. (20):89–101.
Bernal, P.A. 1980. Large-scale biological events in the California Current: the biological response of the epipelagic ecosystem. Ph.D. Dissertation, Scripps Institution Oceanography, University of California of San Diego. 184 pp.
Bernal, P.A. y J.A. McGowan. 1981. Advection and upwelling in the California Current. pp. 381– 399. In Coastal upwelling (F. Richards, ed.). A.G.U. Washington, D.C.
Chelton, D.B. 1981. Interannual variability of the California current physical factors. Calif.Coop. Oceanic Fish.Inv.Rept. (22):34–48.
Chelton, D.B. y R.E. Davis. 1982. Monthly mean sea level variability along the west coast of North America. J.Phys.Oceanog. (12):757–784.
Chelton, D.B., P.A. Bernal y J.A. McGowan. 1982. Large-scale interannual physical and biological interacion in the California Current. J.Mar.Res. 40:1095–1125.
Colebrook, J.M. 1977. Annual fluctuations in biomass of taxonomic groups of zooplankton in the California Current, 1955–59. Fish.Bull.US. 75:357–368.
Haury, L.R. 1976. A comparison of zooplankton patterns in the California Current and the North Pacific central gyre. Mar.Biol. 28:37–49.
Hayward, T.L. y J.A. McGowan. 1979. Pattern and structure in an oceanic zooplankton community. Amer.Zool. 19:1045–1055.
McCreary, J.P. 1976. Eastern tropical response to changing wind systems: with application to El Niño. J.Phys.Oceanog. 6:632–645.
McGowan, J.A. 1974. The nature of oceanic ecosystems. pp. 9–28. In The biology of the oceanic Pacific (C.B. Miller, ed.). Oregon State University Press, Corvallis.
Nelson, C.S. 1977. Wind stress and wind stress curl over the California Current. NOAA Tech.Rep. NMFS-SSRF-714, August 1977.
Reid, J.L. 1962. On the circulation, phosphate-phosphorous content and zooplankton volumes in the upper part of the Pacific Ocean. Limnol.Oceanog. 7:287–306.
Ryther, J.H., D.W. Menzel, E.M. Hulburt, C.J. Lorenzen y N. Corwin. 1971. The production and utilization of organic matter in the Peru Coastal Current. Inv.Pesq. 35:43–60.
Soutar, A. y J.D. Isaacs. 1969. History of fish populations inferred from fish scales in anaerobic sediments off California. Calif.Coop.Oceanic.Fish.Inv.Rept. 13:63–70.
Soutar, A. y J.D. Isaacs. 1974. Abundance of pelagic fish during the 19th and 20th centuries as recorded in anaerobic sediments off the Californias. Fish.Bull.US. 72:257–273.
Venrick, E.L., J.A. McGowan y A.W. Mantyla. 1973. Deep chlorophyll maxima in the oceanic Pacific.Fish.Bull.US. 71:41–52.
Walsh, J.J. 1977. A biological sketchbook of an eastern boundary current. pp. 923–968. In The sea, Vol. 6(E.D. Goldberg, I.N. McCave, J.J. O'Brien and J.H. Steele, eds). Interscience, New York.
Wickett, W.P. 1967. Ekman transport and zooplankton concentration in the North Pacific Ocean. J.Fish.Res.Bd.Canada. 24:581–594
by
Richard H. Parrish, Andrew Bakun, David M. Husby and Craig S. Nelson
Pacific Environmental Group
Southwest Fisheries Center
National Marine Fisheries Service, NOAA
P.O. Box 831
Monterey, California 93942
USA
Resumen
Las principales regiones oceánicas del mundo con corrientes marinas en sus confines orientales (i.e., Sistemas de California, Perú, Canarias y Benguela) parecen comportar dinámicas ambientales similares y contienen congregaciones similares de especies de peces pelágicos. En la medida en que especies correspondientes a diferentes sistemas funcionen como análogos, los estudios comparativos interregionales pueden producir información relativa a los efectos del ambiente en el éxito de la reproducción. Lo que de otra forma sería difícil de obtener de un sólo sistema regional.
Los archivos históricos con informes del clima marino proporcionan una fuente de datos en formatos similares sobre la climatología de procesos océanicos en varias regiones. Estos forman la base para reconocer las diferencias y semejanzas en las diversas áreas de desove de especies correspondientes. La selección natural implica que los hábitos reproductivos en cada caso representan la adaptación a los factores ambientales más cruciales. Por lo tanto los patrones de las relaciones de la reproducción y el ambiente sugieren la existencia de enlances de causalidad importantes.
Esta forma de deducir los factores ambientales que son cruciales puede dar una base racional para limitar las alternativas en cuanto a las variables que pueden explicar estos procesos. Permitiéndo de esta forma el uso más efectivo de las series de datos en la formulación de modelos donde el reclutamiento es dependiente de las condiciones ambientales. Además, el concentrarse en los principales interacciones stock - ambiente puede proveer las bases para utilizar la experiencia que se gana en un sistema para predecir el resultado de acciones en otro sistema.
Normalmente los datos sobre las variables causales primarias (alimento, predación, mortalidad dependiente de la densidad, etc) no están disponibles. Por lo tanto nos concentramos en varios procesos que pueden ser abordados usando datos del mar. Se piensa que estos procesos pueden tener un impacto en los estadios tempranos de la vida de los peces al alterar las variables causales primarias, particularmente el alimento. Estos incluyen: (1) mezcla por turbulencia generada por el viento, que puede dispersar las concentraciones de pequeña escala de las partículas de alimento necesarios para que las larvas se alimenten por primera vez en forma éxitosa; (2) transporte mar afuera, que puede arrastrar los huevos y larvas planctónicos fuera de las áreas costeras que le son favorables; (3) afloramientos costeros, que pueden ser favorables si se presentan con un retraso apropiado pero son perjudiciales si coinciden en espacio y tiempo con el desove. La temperatura es una variable causativa primaria por sus efectos directos sobre procesos fisiológicos.
Los cuatro principales sistemas de corrientes de los confines orientales del mar tienen mucho en común. La ubicación y estacionalidad del máximo transporte Ekman mar afuera parece estar estrechamente relacionado con el desplazamiento latitudinal y estacional de los sistemas de presión atmósferica y con las relaciones entre estos sistemas y la topografía de los continentes. Sin embargo, como se puede ver de las características ambientales que se presentan, hay también grandes diferencias entre ellas. Por ejemplo, los cuatro sistemas tienen anomalías máximas de bajas temperaturas mar afuera mar adento, asociadas con el máximo transporte mar afuera de Ekman de varano. En estas regiones de máximos afloramientos de verano se encuentran stocks de anchoveta y/o sardina con áreas de desove desplazadas hacia el polo y hacia el ecuador. Los sistemas de California y de Benguela tienen, ambos, una sóla área con anomalías superficiales de baja temperatura. En el sistema de California esta área está ubicada en el área de máximo afloramiento de verano, mientras que el sistema de Benguela tiene un área de afloramiento máxima de verano en Luderitz y un área de afloramiento máximo de invierno en Cabo Frío. Los sistemas de Perú y las Canarias tienen cada uno dos áreas separadas de anomalías superficiales de baja temperatura. El sistema peruano tiene un área localizada en el área de máximo afloramiento de invierno (San Juan o Chimbote) y una segunda área en la zona de afloramiento frente a la parte central de Chile, que tiene un máximo de verano en Talcahuano. El sistema de Canarias tiene un afloramiento máximo de verano frente a Lisboa y un segundo afloramiento máximo de verano entre Cabo Sim y Cabo Blanc. Durante el invierno la anomalía de aguas frias frente al noroeste de Africa se desplaza al sur y el máximo del afloramiento se extiende de Cabo Blanc a Cabo Vert.
Los centros de afloramiento (caracterizados por un fuerte transporte mar afuera, intensa mezcla por turbulencia generada por el viento, y reducida estabilidad de la columna de agua) parecen ser evitados como parte de los hábitos de desove. Las áreas de desove de anchovetas y sardinas tienden a estar en las áreas costeras donde el transporte y turbulencia inducidos por el viento son reducidos y la plataforma continental tiende a ser más amplia. Las áreas de desove de las poblaciones más grandes tienden a estar corriente abajo (hacia el ecuador) con respecto a los centros de afloramiento. La población de anchoveta (que ha sido el más grande de todos los stocks actuales de sistemas de corrientes de los confines orientales del mar) desova en un área de fuerte afloramiento. Nuestra hipótesis es que en este caso el ambiente costero particularmente amplio, y la dependencia de la profundidad de la capa de Ekman de la latitud y la estratificación puede servir para reducir el efecto perjudicial del transporte superficial mar afuera de Ekman.
Los patrones para evitar la mezcla por turbulencia y el transporte mar afuera en los hábitos reproductivos sugieren que estos procesos han ejercido un importante control sobre el éxito de la reproducción. Aparentemente estas variables merecen ser consideradas al modelar las variaciones del reclutamiento. En regiones donde las temperaturas características se encuentran dentro de límites fisiológicos, la temperatura no parece ser un factor dominante al determinar el ambiente para el desove.
La capa mínima de oxígeno frente al Perú es muy superficial en comparación còn otras regiones. Se sugiere que esto puede conducir a menor competencia entre los stocks de peces pelágicos y los mesopelágicos.
INTRODUCTION
In this work we have developed climatological fields of oceanographic and meterological data for the four “classical” eastern boundary current systems (Wooster and Reid, 1963). We attempt, through pattern recognition, to define similarities and differences in the characteristic environmental conditions in the spawning grounds of the recognized stocks of pilchard (Sardina and Sardinops) and anchovy (Engraulis) in these systems. Our approach is similar to Reid's (1967) analysis of the oceanic environs of the genus Engraulis. He included the entire life cycle and described the range of surface temperature and salinity in which Engraulis occurs. We have limited our comparisons to the environments in which the early life history stages occur (i.e., the spawning grounds) and have expanded the list of environmental variables.
A major motivation for international cooperation in fisheries research is the assumption that research methods and/or results derived from studying one population can be transferred to another. For example, the recently proposed “International Recruitment Experiment” (IREX)2 is based on the hypothesis that the biological responses to environmental conditions are similar for separate stocks of closely related species. The IREX proposal identifies six variables which are thought to account for most of the biological variability observed in exploited fishes. These variables are temperature, turbulence, transport, food, predation, and population density. Population variations of stocks within a species group, or even in an individual stock, could be caused by any, or all, of the six variables. The hypothesis does not imply that different stocks of a given species group have the same environmental variable as their limiting factor. Rather it implies that the stocks have a similiar functional relationship to that environmental variable.
In this work we are addressing a narrow subset within the IREX construct. The species groups considered are anchovies and pilchards, and we are limiting the analyses to comparisons of the environments of their spawning grounds in the four major eastern boundary currents. The regions of interest consist of the California Current off western North America, the Peru Current off western South America, the Canary Current off the Iberian Peninsula and northwestern Africa, and the Benguela Current off southwestern Africa (Fig. 1). Data which could be used to compare the predation, food, and density variables are not available for the spawning grounds of most of the stocks in the California, Peru, Canary, and Benguela Currents. We are therefore basing our comparisons upon several processes which can be indicated by available maritime data. These processes are thought to have important impacts on the early life history of the two species groups by altering the primary causative variables, particularly food. From the point of view of the utility of the study, results based on routinely available data offer the possibility of being followed in real time for management purposes.
Eastern boundary currents are characterized by equatorward surface flow and coastal upwelling. One result of these factors is that the eastern sides of the oceans have a very extensive temperate zone. For example, the 20° and 10°C isotherms at 10 meters occur at lat. 27°N and 60°N in the eastern North Pacific, and in comparison they occur at lat. 40°N and 45°N in the western North Pacific (Barkley, 1968). Another feature of eastern boundary current regions is the tendency for narrow continental shelves. Consequently the fisheries of eastern boundary currents are dominated by a small number of temperate pelagic fishes which can achieve very large populations. The physical environments of the four major eastern boundary currents are quite similar and therefore it is not surprising that their fisheries are dominated by very closely related, geographically isolated stocks or species (Table 1). Pilchards, anchovies, jack mackerel (Trachurus), and hake (Merluccius) achieve the largest populations; mackerel (Scomber japonicus) and bonito (Sarda) also are important in the fisheries of the eastern boundary currents, although they don't attain the population size of the other four species groups.
2 Report of Scientific Committee on Ocean Research (SCOR) and Advisory Committee of Experts on Marine Resources Research (ACMRR), Working Group 67 (Oceanography, Marine Ecology and Living Resources) developed in response to the Ocean Sciences in Relation to Living Resources (OSLR) resolution of the Intergovernmental Oceanographic Commission (IOC), 1 July 1982.
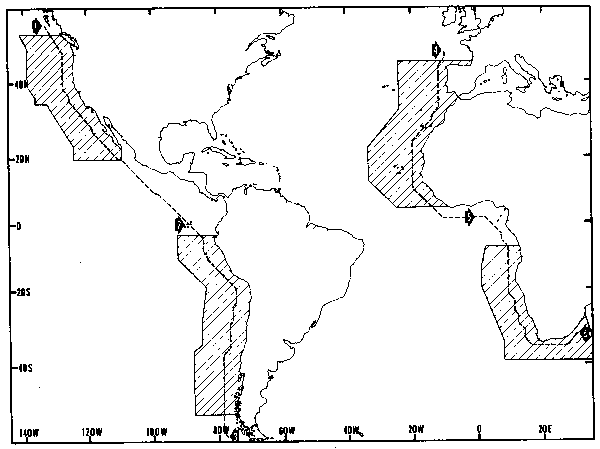
Figure 1. Geographical orientation of various summaries presented in this paper Appendix maps are assembled from summaries within 1–degree areal segments of the four eastern boundary current regions indicated by diagonal hatching. Alongshore vertical section (Figs. 3–6)are arranged along the dashed lines near the eastern ocean boundaries.
| CALIFORNIA CURRENT | PERU CURRENT | CANARY CURRENT | BENGUELA CURRENT |
|---|---|---|---|
| Engraulis mordax | Engraulis ringens | Engraulis encrasincholus | Engraulis capensis |
| Sardinops sagax | Sardinops sagax | Sardina pilchardus | Sardinops ocellatus |
| Trachurus symmetricus | Trachurus symmetricus | Trachurus trachurus | Trachurus trachurus |
| Merlucciue productus | Merluccius gayi | Merluccius merluccius | Merluccius capensis |
| Scomber japonicus | Scomber japonicus | Scomber japonicus | Scomber japonicus |
| Sarda chiliensis | Sarda chiliensis | Sarda sarda | Sarda sarda |
Fisheries for pilchards and anchovies are notorious for their extreme variability. All four current systems at one time or another have had fisheries dominated by pilchards (Fig. 2). The northern stock in the California Current system (Radovich, 1981) and the Namibian (Troadec, Clark, and Gulland, 1980) and South African (Cram, 1981) stocks in the Benquela Current system have collapsed under heavy exploitation. The Peru Current system (Serra et al., 1979) and the Canary Current system (FAO, 1978) have had recent increases in the size of pilchard stocks, and landings in these regions are presently dominated by pilchards. Conversely, anchovies presently dominate the landings in the Benguela and California Current systems and appear to be abundant, although lightly exploited in the Canary Current system. The collapse of the Peru anchoveta stock under heavy exploitation is well known (Tomczak, 1981).
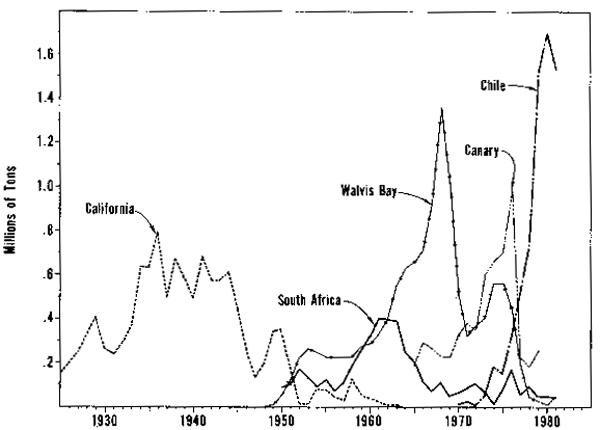
Figure 2. Yearly pilchard (sardine) landings from several eaśtern boundary current stock.
The basis for our comparison is, like the IREX proposal, dependent upon the hypothesis that closely related stocks or species have similar early life history requirements. This would suggest that the environments of the various spawning grounds are similar. That is, in the mean they should fall within the tolerance range of the species group. This does not imply that environmental conditions in the spawning grounds are optimal, only that stocks have recently maintained populations by spawning there.
How unrelated may the stocks be before this hypothesis becomes ridiculous? Most fisheries biologists would probably agree3 that the present California stock of pilchard (sardine) has the same physiological requirements as the stock had in the 1930's before it collapsed. However, the present remnant of the stock may have a different gene pool than the original. Most fisheries biologists would probably agree3 that, within a given species, physiology is nearly identical. Taxonomic distinctions at the species-generic level are quite plastic. The mackerel stocks of the eastern boundary currents are now considered to belong to the same species, Scomber japonicus. Just a few years ago they were considered to be different species and the genus was not Scomber but Pneumatophorus. Each eastern boundary current presently has a different species of Engraulis. These “species” are very similar and it takes considerable expertise to tell them apart. To quote McDowell (1962), “There are grounds for suspicion that this expertise is not possessed by the fish themselves….” We are assuming that in the case of geographically separated stocks of closely related species the differences in the physiological requirements of the early life history stages are negligible.
To provide a basis for our comparisons, historical subsurface oceanographic data and surface marine weather observations are used to examine the large-scale spatial and seasonal patterns of several environmental parameters which appear to have had important effects on the spawning habits of pelagic fishes in eastern boundary current systems. Our comparative studies are primarily focused on the atmosphere-ocean processes related to horizontal and vertical advection, vertical stability, and temperature. Bakun and Parrish (1980) discussed the rationale for selecting the particular suite of oceanic and atmospheric properties described in this study.
Characteristic annual mean distributions of oceanographic data and summer and winter climatologies based on surface marine weather observations are presented in the Appendix as charts of surface geostrophic flow, sea surface temperature and coastal temperature anomaly, surface wind- driven transport, an index of wind-generated turbulent mixing, and total cloud amount. The methods used to compile the climatological data are summarized in the Appendix. The format of placing the charts in an appendix, rather than in the text, was selected to facilitate simultaneous intra-and interregional geographic and seasonal comparisons of the large-scale oceanographic and atmospheric properties.
3 A hedge word is always necessary when discussing what fisheries biologists will agree upon.
REGIONAL ASPECTS
The surface flow field in eastern boundary current systems, at least on the time and space scales affecting reproductive success of fish stocks (Parrish, Nelson, and Bakun, 1981) can be considered to be a combination of the surface Ekman drift (i.e., the transport of surface waters under the direct action of the wind) and the underlying geostrophic flow field. Highly smoothed representations of the geostrophic flow patterns in the four regional systems (Appendix Chart 1) show major common features. Near the coast, large-scale flow is generally equatorward. At high latitudes the tendency is poleward. Within several hundred kilometers of the coast, a component of large-scale geostrophic flow toward the coast is a common feature. The far offshore regions are characterized by flow toward the coast in the higher-latitude west wind drifts, weak equatorward flow at mid-latitudes, and flow toward the ocean interiors in tropical regions.
Coastal temperature anomaly patterns (Appendix Charts 5 and 6) indicate a general tendency for cool coastal temperatures relative to those offshore. These patterns show local centers of intense cooling associated with coastal upwelling. There also are relatively warm areas within coastal indentations (responding to seasonal warming) and at the high latitude extremes of the regions (associated with warm advection in the poleward flows).
Surface Ekman transport (Appendix Charts 7 and 8) is generally offshore, but becomes onshore at higher latitudes during the winter seasons. Wooster and Reid (1963) have demonstrated that the major geographical and seasonal features of eastern boundary coastal cooling can be related to corresponding features in offshore Ekman transport. Major capes, marking substantial changes in large-scale coastline trend, tend to be regions of maximum offshore Ekman transport and of associated upwelling and coastal cooling.
GEOSTROPHIC CURRENT CHARACTERISTICS
The common large-scale features of the eastern boundary currents have been described by Wooster and Reid (1963). The surface flow in these large systems is predominantly in the sense of the prevailing winds, which is anticyclonic about the mid-ocean atmospheric high pressure cells. The weak (typically less than 50 cm sec-1) equatorward geostrophic flow in eastern boundary currents is due to the baroclinicity of a relatively shallow layer, generally confined to the upper 500 m (Wooster and Reid, 1963).
Another common feature of eastern boundary currents is a poleward flowing undercurrent, which is a subsurface current usually centered at 200–300 m near the continental shelf edge. The waters transported poleward by the undercurrents appear to be enhanced by coastal upwelling and may contribute water to the upwelling centers (Smith, 1978). A poleward countercurrent may occur seasonally at the surface during the winter and during periods when the equatorward wind stress weakens.
Coastal countercurrents are often part of semi-permanent eddy circulations, for example, the Southern California Eddy (Sverdrup and Fleming, 1941). A permanent cyclonic circulation over the continental slope and shelf between Cape Frio and south of Lüderitz in southwest Africa has been postulated by Nelson and Hutchings (MS). In the Canary Current region a cyclonic circulation has been observed in the shallow Banc d'Arguin region south of Cap Blanc (Mittelstaedt, 1974) and may be seen in the schematic diagrams of Rebert (1978).
In addition to the narrow coastal currents and the poleward undercurrents, the large-scale surface circulation in eastern boundary current systems is complicated by the effects of bottom topography and coastline orientation. The advent of satellite infrared imagery of the sea surface and intensive hydrographic surveys in the upwelling regions have identified the existence of many eddy-like features or intense frontal zones associated with upwelling sites or downstream of capes and headlands (Bernstein, Breaker, and Whritner, 1977).
California Current System
The major features of the California Current system have been described by Reid, Roden, and Wyllie (1958), Pavlova (1966), and Hickey (1979). The cool, low salinity waters of the west wind drift diverge at about lat. 45°N, one branch flowing north to feed the Alaskan Current and the other branch flowing southeastward to become the California Current. The equatorward flow between lat. 45°N and 20°N and within about 500 km of the west coast encompasses the bulk of the California Current. Maximum equatorward flow occurs approximately 200–400 km from the coast and is strongest in the summer and early fall. A portion of the California Current turns into the coast at about lat. 30°N and is associated with negative wind stress curl in this region (Nelson, 1977). There is a relatively permanent northward flow near the coast between lat. 30°N and Point Conception (lat. 35°N) which forms the eastern portion of the Southern California Eddy in the Southern California Bight.
The California Undercurrent is a poleward flow of warm, relatively high salinity water at depths of approximately 250–300 m which lies close to the continental slope (Wooster and Jones, 1970). This current is present along the coast from southern Baja California to Vancouver Island (Reed and Halpern, 1976). The northward flow at the surface occuring during the fall and winter along the coast north of Point Conception is commonly called the Davidson Current.
Peru Current System
The north-northwestward flow north of lat. 40°S has been labelled the Peru or Humboldt Current. Gunther (1936) first recognized two branches of this current: a Peru Coastal Current and a Peru Oceanic Current. The northward flow generally splits into the Coastal and Oceanic Currents at about lat. 25°S (Wyrtki, 1965). The annual mean geostrophic flow (Appendix Chart I) shows this bifurcation at lat. 25°S, long. 76°W and a component of the flow is directed onshore. Both the coastal and oceanic portions of the Peru Current originate in the sub-Antarctic region and transport cold, relatively low salinity water equatorward.
The Peru Countercurrent flows southward at about long. 80°W between the Peru Coastal Current and the Peru Oceanic Current. It is primarily a subsurface current with the strongest flow near 100 m.
The Peru Coastal Current coincides approximately with the upwelling region off Peru and Chile. Coastal bathymetry appears to have considerable influence on the current. Eddies are found wherever the coastal current is deflected strongly from the coast, especially near lat. 15°S (Cañon, 1978). The annual mean surface geostrophic flow is directed onshore north of lat. 15°S and flows strongly onshore in the coastal region between lat. 10°S and 12°S. Wooster and Sievers (1970) noted that the surface drift currents at the coast showed a minimum in equatorward flow between lat. 12°S and 14°S in all seasons, except summer.
During the austral winter the southeast trade winds reach maximum intensity. From April to September the Peru Current appears stronger than in other months and the countercurrent virtually disappears. At this time the two branches of the Peru Current coalesce and the contribution to the South Equatorial Current is the strongest.
The Peru-Chile Undercurrent is a narrow southward flowing countercurrent beneath the Peru Coastal Current. It is found offshore north of lat. 15°S, but south of lat. 15°S it follows close to the coast and can be identified as far as lat. 48°S (Silva and Neshyba, 1979). It is characterized by a low oxygen current.
Canary Current System
The large-scale geostrophic flow in the Canary Current region shows an equatorward component between about lat. 36°N and 20°N. The Canary Current leaves the coast in the vicinity of Cap Blanc (lat. 21°N), which is also the location of persistent upwelling. Cap Blanc is the site of a semi-permanent frontal zone between the North Atlantic Central Water advected southwest by the Canary Current and a low salinity South Atlantic Central Water mass which flows northward along the coast (Hughes and Barton, 1974).
During periods of weak northerly winds, which occur particularly in summer and auturnn between lat. 10°N and 20°N, a coastal northwest-flowing countercurrent may be observed (Mittelstaedt, 1974). This coastal countercurrent gives rise to a cyclonic circulation over the shallow Banc d'Arquin south of Cap Blanc. The coastal countercurrent may be a response to the large discharge of fresh water during the summer from the Gambia and Senegal rivers.
Hughes and Barton (1974) found evidence of a northward flow at a depth of about 250 m extending from Cap Vert to Cabo Bojador. This poleward undercurrent was indicative of a narrow stream of cool, low salinity water flowing parallel to the continental shelf edge.
Benguela Current System
The major large-scale geostrophic currents off southwest Africa are the equatorward flow between lat. 35°S and about lat. 20°S, which is called the Benguela Current, and the primarily westward flowing Agulhas Current, which flows along the coast of south Africa to approximately long. 20°E where it is deflected to the south and east by the confluence with the west wind drift. A portion of the flow continues toward the northwest around Cape Agulhas and feeds a semi-permanent coastal jet between Cape Agulhas and Cape Columbine (Bang and Andrews, 1974). The principal features of the Benguela Current are described by Hart and Currie (1960).
The Benguela Current consists of the cool upwelled water found within 150 km of the west coast between lat. 15°S and 34°S (Shannon, 1970). In general, it flows northward, roughly following the isobaths between lat. 34°S and 23°S, while north of lat. 23°S it tends to veer away from the coast. Closer inshore a southward moving countercurrent has been occasionally recorded. Hart and Currie (1960) deduced the presence of a southward undercurrent on the continental slope off the Namibian coast from the downwarping of the mid-depth isopycnal surfaces as they approached the continental slope.
The existence of a relatively permanent cyclonic circulation over the continental slope and shelf regions between Cape Frio and a point just south of Lüderitz has been noted by Nelson and Hutchings (MS). The cyclonic circulation is comprised of the northwesterly jet-like flow southwest of Lüderitz where the flow of the Benguela Current accelerates towards the gap in the Walvis Ridge, and a southward return flow near the coast between Cape Frio and Lüderitz. The southward flow near the coast below 30 m has been reported by many investigators. Surface dynamic topography patterns from a series of monthly hydrographic cruises in this area in 1959–61 showed many instances of cyclonic gyres in the region between lat. 25°S and 20°S and southward geostrophic flow north of Lüderitz during spring and summer (Stander, 1964).
DISSOLVED OXYGEN
Minimum oxygen layers are associated with eastern boundary currents. These minimum layers are shallowest and most pronounced in the lower latitudes and more extensive and pronounced in the Pacific than in the Atlantic (Figs. 3,4). For large-scale comparison of dissolved oxygen and its relationship to the upper mixed layer we developed annual dissolved oxygen and density (sigma-t) sections from data summarized by I-degree squares. The sections are situated several degrees offshore to allow continuous deep-water sections. This procedure introduces some bias in relation to smaller scale, nearshore features. For example, coastal upwelling or other circulation processes may result in intrusions of low oxygen water into shallower depths on the shelf, which are not indicated by our offshore sections. These intrusions are most likely to occur in the lower latitude areas which are closer to the low oxygen sources. Such intrusions have been reported in the Benguela (Stander and de Decker, 1969) and Peru Currents (Guillén, 1980).
In the California, Canary, and Benguela Currents the minimum oxygen level is altered significantly in the vicinity of the subtropical convergences. The low latitude boundaries of these regions are marked by sharp sigma-t gradients associated with the subtropical convergences (Figs. 5, 6). They occur at about lat. 3°S in the Peru Current, lat. 17°S in the Benguela Current, lat. 22°N in the Canary Current, and lat. 25°N in the California Current. The upwelling regions of the eastern boundary currents are also apparent in the density sections; there are few sigma-t surfaces intercepting the surface in the upwelling regions. Equatorward of the convergences there is a sharp dissolved oxygen gradient associated with the thermocline and low dissolved oxygen values occur very near the surface. In the Canary and Benguela systems the minimum oxygen layers deepen and disappear in the vicinity of the subtropical convergences (i.e., at about lat. 20°N and 20°S). In the California Current system the minimum oxygen layer deepens significantly near the subtropical convergence; however, an extensive minimum oxygen layer extends throughout the entire system. Throughout the California Current system low oxygen levels (i.e., less than 2 mL L-1) occur at about 300 m and the oxygen minimum layer occurs at 700–900 m.
The Peru Current is significantly different from the other three currents with respect to dissolved oxygen. The subtropical convergence occurs much closer to the equator and the low oxygen layer expands and shoals in the vicinity of the convergence. The oxygen minimum is at about 300 m throughout the Peru Current system. Off Chimbote, very low dissolved oxygen levels occur just beneath the upper mixed layer. However, at Coquimbo oxygen concentrations of 2 mL L-1 occur at about 300 m; this is similar to the California Current situation.
Low oxygen concentration could affect reproduction of pelagic fishes through reduction of growth rates of either larvae or adults (Pauly, 1981). It could also increase mortality rates by concentrating the adults or larvae to narrow layers near the surface where they might be more susceptible to predation. However, the “worst” dissolved oxygen conditions occur in the spawning areas of the Peruvian stocks which, at least in the case of Engraulis, lie in the most productive region. The other areas most likely to be influenced by low oxygen concentrations are the spawning areas of the Namibian stocks near Palgrave Point in the Benguela Current, and the Chilean stock, which has spawning grounds near Africa. These two areas are the next most productive regions after Peru.
We see two possible explanations for the association of low oxygen concentration and high pelagic fish productivity. Low oxygen concentration is associated with high nutrient concentrations. Low oxygen concentrations near the thermocline might therefore be associated with high primary production; more primary production, hence more fishery production. The other explanation concerns competition. Extensive egg and larval surveys have been made off Peru and California. Ahlstrom (1965) showed that mesopelagic fishes and hake accounted for about 22 percent and 17 percent, respectively, of the total larvae taken off California during the 1955–58 surveys. Both mesopelagic fishes and hake have their greatest abundance below the thermocline (Ahlstrom, 1959). Off Peru mesopelagic fishes and hake comprised only 4.1 and 0.37 percent of the larvae taken during 1966–68 (Santander and de Castillo, 1979). Anchovy larvae comprised 37 percent of the larvae in the California surveys and 92 percent in the Peruvian surveys. The very low oxygen levels just beneath the thermocline off Peru would be expected to severely limit the habitat space for hake and mesopelagic fishes. Habitat space for mesopelagic fishes in the California and Canary systems and most of the Benguela system is extensive and large stocks in these areas would be expected to be in competition with epipelagic fishes for food.
Figure 3. Dissolved oxygen (mL L-1 vs. depth along the eastern boundary of the Pacific Ocean. The orientation of the section is shown in Figure 1. Initials at the top of the diagram indicate the latitudinal positions of locations discussed in detail in Section III of this paper.
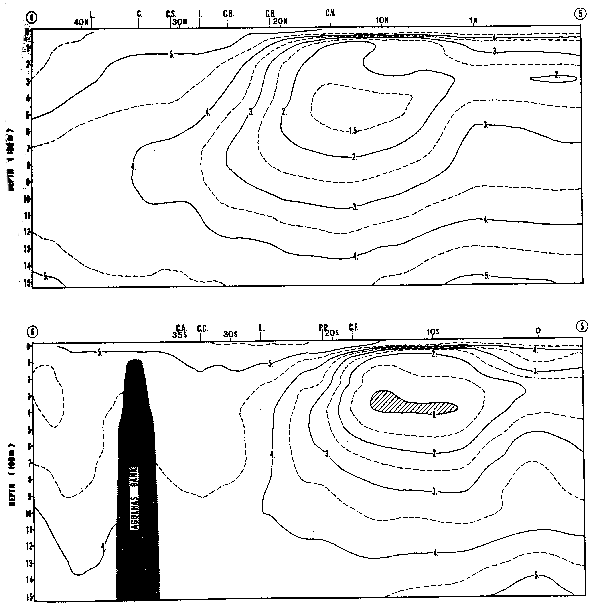
Figure 4. Dissolved oxygen (mL L-1) vs. depth along the eastern boundary of the Atlantic Ocean. The orientation of the section is shown in Figure 1. Initials at the top of the diagram indicate the latitudinal positions of locations discussed in detail in Section III of this paper.
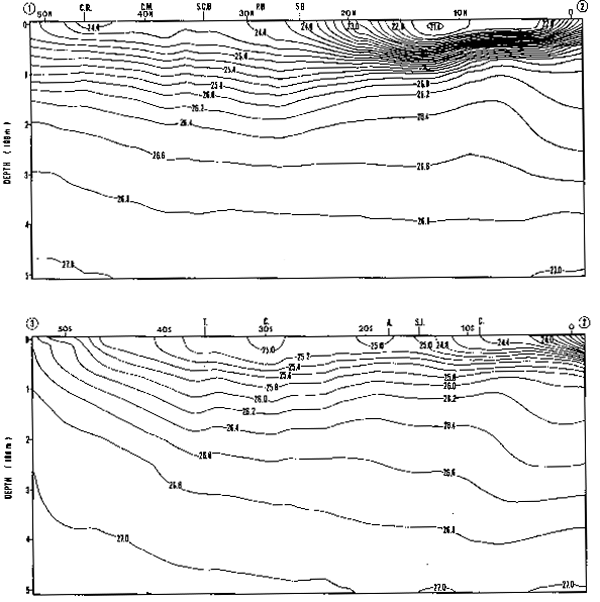
Figure 5. Sea water density (sigma-t) vs. depth along the eastern boundary of the Pacific ocean. The orientation of the section is shown in Figure 1. Initials at the top of the diagram indicate the latitudinal positions of locations discussed in detail in Section III of this paper.
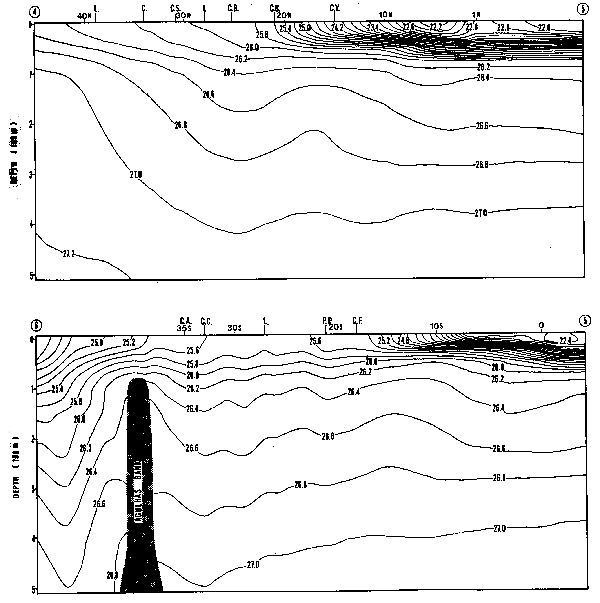
Figure 6. Sea water density (sigma-t) vs. depth along the eastern boundary of the Atlantic Ocean. The orientaion of the section is shown in Figure 1. Lines at the top of the diagram indicate the latitudinal positions of locations discussed in detail in Section III of this paper.
SUBREGIONAL ASPECTS
The charts presented in the appendix are a subset of a considerable volume of material produced and examined in this study. In order to make “cuts” of this material for presentation we have chosen several fixed locations in each eastern boundary region which appear to exemplify the range of environmental characteristics encountered in the near-coastal environments. These locations are labeled in terms of a recognizable feature of coastal geography (i.e., a coastline feature or a coastal city) and appear on the appendix maps and on Figure 7, and are listed in Tables 2 to 6.
Tables 2 and 3 briefly summarize information available on the geography and seasonality of sardine and anchovy spawning in the four eastern boundary current systems. The quality and quantity of information available varies widely among regions and within regions. Thus a fully consistent and complete synthesis is not available to us at this point. Tables 2 and 3 thus should not be considered to be “hard” data, and should not be cited as authority. They are presented as aids to recognition of patterns in the environmental summaries we have assembled (i.e., from the point of view of exploratory data analysis and hypothesis generation rather than of hypothesis testing). Our attempt at preliminary synthesis is intended to generate feedback from workshop participants and others with more specific knowledge of local situations. A discussion of the background and references to Tables 2 and 3 for the California and Peru systems are available in Bakun and Parrish (1982); major references for the Canary system are Bravo de Laguna, Fernandez, and Santana (1976), FAO (1978), and Belvèze and Bravo de Laguna (1980);major references for the Benguela system are King (1977), LeClus (1979), Badenhorst and Boyd (1980), Crawford (1980), and Crawford, Shelton, and Hutchings (1980).
Table 4 lists characteristic sea surface temperature values for 2-month segments of the annual cycle. These values were selected from bimonthly maps of long-term mean summaries which are described in the Appendix and from similar displays of the standard errors of the estimated mean values, in such a way as to characterize the shoreward 50 km of ocean or the locality over which spawning occurs. For example, in the Columbia River Plume, spawning occurs at distances of 100 km or more from the coast and so our presentations attempt to reflect conditions at that location, rather than directly adjacent to the coast.
Figure 7 displays the offshore component of surface Ekman transport plotted against the cube of the wind speed (indicating the rate of addition of turbulent mixing energy to the water column). Table 5 lists values of mixed layer depth obtained from bathythermograph data (Husby and Nelson, 1982). Sparsity of available data restricts this presentation to the California Current locations and the three northern locations in the Peru Current region. Profiles of stability structure for three Benguela locations based on hydrographic data, which lack the vertical resolution provided by bathy- thermograph traces, are shown in Figure 8. Table 6 contains the widths of the adjacent continental shelves.
Table 2. Spawning seasons of Pilchards (Relative size of dots is only meaningful at each location)

| JAN–FEB | MAR–APR | MAY–JUN | JUL–AUG | SEP–OCT | NOV–DEC | JAN–FEB | MAR–APR | MAY–JUN | JUL–AUG | SEP–OCT | NOV–DEC | ||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| COLUMBIA R. | 9.2 | 9.5 | 12.6 | 15.1 | 14.7 | 11.3 | LISBON | 14.0 | 14.0 | 16.2 | 18.0 | 18.0 | 15.2 |
| CAPE MENDOCINO | 11.0 | 10.6 | 11.1 | 12.1 | 12.9 | 12.1 | CASABLANCA | 16.4 | 16.5 | 18.8 | 22.3 | 21.6 | 18.1 |
| S.CALIF.BIGHT | 14.0 | 14.016.0 | 18.0 | 18.0 | 15.4 | CAP SIM | 16.6 | 16.9 | 17.9 | 18.0 | 18.7 | 17.4 | |
| PUNTA BAJA | 16.2 | 15.5 | 16.4 | 19.1 | 19.5 | 17.6 | IFNI | 17.1 | 17.6 | 18.2 | 19.8 | 20.0 | 18.4 |
| S.BAJA CALIF | 18.9 | 17.2 | 17.1 | 21.4 | 24.0 | 21.4 | C.BOJADOR | 18.1 | 17.6 | 19.0 | 20.5 | 20.9 | 20.2 |
| CAPE BLANC | 17.7 | 17.8 | 18.2 | 20.6 | 21.0 | 19.1 | |||||||
| CAP VERT | 19.6 | 19.5 | 23.0 | 27.2 | 27.8 | 24.5 | |||||||
| JUL–AUG | SEP–OCT | NOV–DEC | JAN–FEB | MAR–APR | MAY–JUN | JUL–AUG | SEP–OCT | NOV–DEC | JAN–FEB | MAR–APR | MAY–JUNE | ||
| CHIMBOTE | 17.6 | 16.9 | 18.5 | 21.3 | 21.1 | 18.6 | CAP FRIO | 14.8 | 14.9 | 16.5 | 18.7 | 17.3 | 15.7 |
| SAN JUAN | 15.6 | 15.1 | 17.3 | 19.6 | 18.1 | 16.7 | PALGRAVE PT. | 14.8 | 14.2 | 15.0 | 18.6 | 17.6 | 16.1 |
| ARICA | 16.8 | 17.6 | 20.3 | 22.9 | 21.8 | 18.9 | LUDERITZ | 13.0 | 13.0 | 14.0 | 15.0 | 14.0 | 14.5 |
| COQUIMBO | 13.6 | 13.5 | 15.3 | 17.1 | 16.5 | 14.5 | C. COLUMBINE | 14.0 | 14.7 | 16.0 | 16.0 | 15.0 | 15.0 |
| TALCAHUANO | 12.0 | 12.2 | 14.1 | 14.9 | 14.7 | 13.2 | C.AGULHAS | 15.2 | 15.8 | 19.0 | 20.0 | 18.4 | 16.6 |
| 46°S | 8.6 | 10.8 | 11.1 | 13.1 | 12.7 | 9.9 |
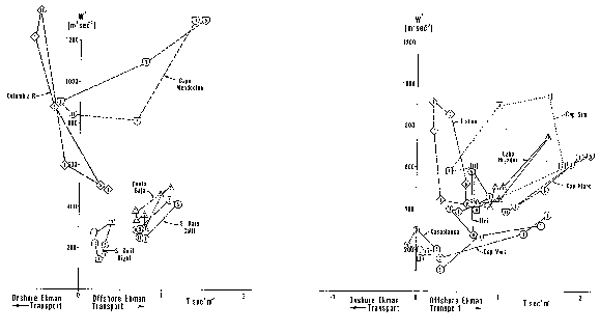
Figure 7. Characteristic seasonal relationship between W3 (cube of the wind speed) index of turbulent mixing energy producion and offshore Ekman transport at various locations in the California, Canary, Peru and Benguela Current systems. Each numbered symbol represents a 2-month of the two (e.g., 1 represents Jan.-Feb., 3 represents Mar.-Apr., etc.).
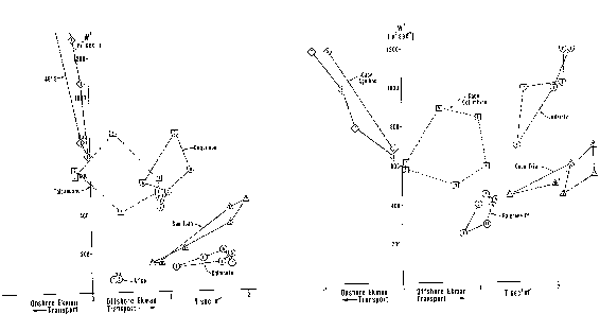
Figure 7. (continued)
| JAN–FEB | MAR–APR | MAY–JUN | JUL–AUG | SEP–OCT | NOV–DEC | |
| COLUMBIA R. | 103 | 118 | 10 | 10 | 16 | 43 |
| CAPE MENDOCINO | - | 91 | 42 | 28 | 54 | 27 |
| S.CALIF. BIGHT | 37 | 25 | 11 | 10 | 14 | 20 |
| PUNTA BAJA | 37 | 26 | 18 | 12 | 14 | 26 |
| S.BAJA CALIF. | 45 | 23 | 18 | 9 | 20 | 34 |
| JUL–AUG | SEP–OCT | NOV–DEC | JAN–FEB | MAR–APR | MAY–JUN | |
| CHIMBOTE | 40 | 23 | 12 | 7 | 12 | 29 |
| SAN JUAN | 89 | 87 | 19 | 29 | 20 | 73 |
| ARICA | 14 | 18 | 9 | 5 | 5 | 10 |
| COLUMBIA R. | 54 | LISBON | 43 |
| CAPE MENDOCINO | 21 | CASABLANCA | 56 |
| S.CALIF.BIGHT | 168 | CAP SIM | 38 |
| PUNTA BAJA | 29 | IFNI | 39 |
| S. BAJA CALIF. | 71 | C.BOJADOR | 40 |
| CAP BLANC | 62 | ||
| CAP VERT | 32 | ||
| CHIMBOTE | 132 | CAPE FRIO | 32 |
| SAN JUAN | 21 | PALGRAVE PT. | 73 |
| ARICA | 53 | LUDERITZ | 40 |
| COQUIMBO | 10 | C. COLUMBINE | 75 |
| TALCAHUANO | 44 | C. AGULHAS | 128 |
| 46°S | 93 | ||
*Distance from coast to 200-m contour (in the case of S. Calif. Bight, distanceis to the 200-m contour at the seaward edge of the continental borderland).
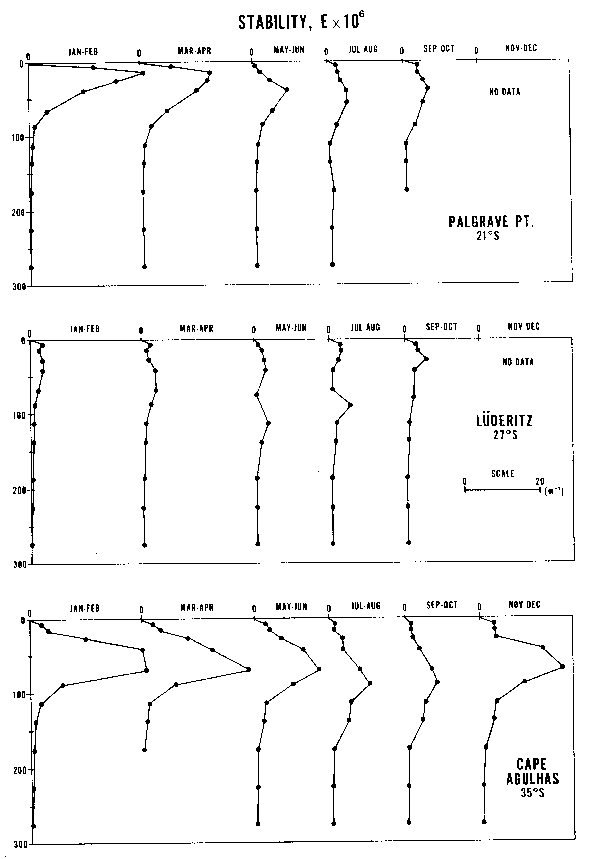
Figure 8. Static stability (m-1) vs. depth for 2-month segments (for which there are sufficient hydrographic data to assemble a representative profile) of the seasonal cycles at three locations in the Benguela Current region. Depths are in meters. A scale for magnitude of stability appears to the right on the middle panel.
![]()
![]()
![]()