![]()
![]()
![]()
Sudd is a large area covered in aquatic plants and situated in Sudan, between 5° 30' and 9° 30' N in latitude and between 30° and 32° E in longitude, where it extends from Juba to Lake No. The word “sudd” is derived from the Arabic word “sadd”, meaning “block” (Gaudet and Falconer, 1983). The term is now being used widely for large solid floating vegetation islands and mats. The Sudd region in the Sudan has remained unnavigable for many years because large islands of papyrus have floated downstream of the Nile and lodged in channels, effectively blocking local river traffic. In 1980 the swamps of Sudd covered about 16 500 km2 (Hickley and Bailey, 1986). Typha dominguensis covers about three quarters of the total swamp, bordering Cyperus papyrus which forms a central core. Water hyacinth (Eichhornia crassipes) now forms an almost ubiquitous floating fringe to river channels and lakes in Sudd swamps.
Both before and after the Eichhornia invasion marginal vegetation in the Sudd has been shown to support a wealth of small and juvenile fish (Sandon, 1951; Hickley and Bailey, 1986) and rich and diverse macroinvertebrate faunas (Monakov, 1969). According to Bishai (1960/61) the best fishing grounds, giving a maximum annual yield of fish, were those not infested with water hyacinth. The main fishing grounds in the Sudd are in the main water passages, side channels and lagoons, and fishing camps are established there every year. In his review of the Sudd, Rzoska (1974) concluded that vegetation fringe in which Pistia stratiotes had a major role, formed a unique, and biologically the richest, habitat in the whole swamp system. Although Pistia has been largely replaced by water hyacinth, Bailey and Litterick (1993) have found that the fringing hyacinth in Lake Wutchung in the Sudd region has taken over the function of Pistia in harbouring large quantities of macroinvertebrates, with the highest concentrations present within the first 6 m of the fringe (Fig.34). While the outer fringe was dominated by coleopterans, odonatans, gastropods (including Bulinus forskalii, B. truncatus, Biomphalaria pfeifferi, B. sudanica), and the crustacean Caridina nilotica, further in the mat coleopterans, hydracarines and gastropods were prominent. In free-floating water hyacinth rafts covering up to 10 m2 there were over seven times fewer invertebrates, especially a notable reduction in crustaceans and ephemeropterans, suggesting that the macroinvertebrates there were subject to heavy preying by fish due to an easy access to the underside of such floating plant rafts. This agrees with the findings of low biomass of invertebrates on the isolated, off-shore Ceratophyllum, stranded on flooded trees of Volta Lake in Ghana (Petr, 1968), suggesting that apart from the increased exposure to predation, the floating plants have little chance of being recolonized by benthic-dwelling invertebrates.
Hickley and Bailey (1986) sampled fish in two major habitats, i.e. flowing channels and a lake situated within the sudd. The greatest diversity of fish (some 60 species) were caught in the lake's papyrus zone. Eichhornia crassipes, which forms a floating fringe of variable width in channels and lakes, were found to harbour rich periphyton and communities of aquatic invertebrates which the plant provides with shelter and food. Deeper inside of the Eichhornia mat, with a decrease in the concentration of dissolved oxygen reaching 15–20% of saturation level, the fish typically recorded were air-breathers, such as Polypterus senegalus and Ctenopoma spp (Hickley and Bailey, 1987). Many non-air breathing river and lake fish species were found to move through conduits in the permanent floodplain created by swamps, but the low dissolved oxygen content or hypoxic conditions, high concentrations of CO2 and ammonia must prevent their lateral dispersal into the swamp.
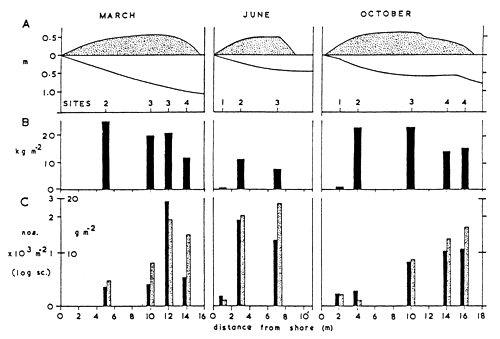
Fig. 34. Seasonal and spatial changes in the hyacinth fringe of the sudd in Lake Wutchung
(Sudan).
A: Fringe and shore profiles. Sites 1 = landward zone; Sites 2 = inner middle zone;
Sites 3 = outer middle zone; Sites 4 = outer zone.
B: Estimated fresh weight biomasses of hyacinth (kg m-2).
C: Estimated macroinvertebrate numbers (solid bars) and fresh-weight biomasses (stippled
bars, g m-2).
In the Sudd, submersed beds of Naias pectinata, Vallisneria aethiopica and Ceratophyllum demersum, with their rich periphyton, offer abundance of food to Heterotis, Distichodus and several tilapias.
Annual flooding of the permanent Sudd swamps inundates large areas of Oryza longistaminata grasslands, where a number of fish species enter, such as Clarias gariepinnus, Polypterus senegalus, Channa obscura, Oreochromis niloticus (Hickley and Bailey, 1987a).
In addition, large resident and migrant populations of birds are present in the Sudd. Of these about 20 species feed on submersed vegetation in shallow waters, over 100 species consume invertebrates, and some 40 species feed on fish or their fry (Hickley and Bailey, 1987).
Floating islands occur regularly on a number of water bodies in Africa, including major rivers such as the Nile and Congo. They cause a variety of problems, especially for boat transport, water intakes, but also for fishermen whose nets and other fishing gear become entangled. Floating islands made largely of papyrus have interfered with fishing in lakes such as George (Uganda), Jipe (Tanzania-Kenya), Naivasha (Kenya), Chad, Victoria, and Kyoga. Remote sensing has been used to follow the movement of floating islands on Lake Naivasha (Gaudet and Falconer, 1983), using the Landsat satellite data. This has allowed to determine their mobility, including the separation of the movements of papyrus and salvinia. The major movements take place in January-February, under the influence of the changing wind patterns. Prompt communication of the results to the fisheries community would assist in adapting their fishing strategies.
The word ‘sudd’ has been applied also to floating islands of aquatic macrophytes formed on some man-made lakes in Africa (Gaudet, 1979a). Within several years after the closure of the dam and filling of these large reservoirs sudd started forming. On Volta Lake (Ghana) mats became established in the shallows around drowned trees, with the major plants being Vossia cuspidata, Leersia hexandra and Oryza barthii (Photo 14). Some southern parts of the lake developed sudd through colonisation of Pistia stratiotes by Scirpus cubensis and Typha dominguensis, with Utricularia inflexa fringing the margins (Petr, 1986). Sudd has also developed on the shallow Kpong reservoir situated on the Volta River just downstream of the Volta Lake.

PHOTO 14: Flooded trees at the margins of Lake Volta, Ghana. Note the belt of Vossia cuspidata.
On Lake Kariba, after Salvinia molesta developed thick mats, it combined with Pistia stratiotes and Ludwigia spp to form sudd. For a number of years the sudd impeded passage and prevented fishing activities in some areas of this reservoir. Salvinia together with Ludwigia supported seven times more invertebrate biomass than did Ceratophyllum and Potamogeton pussillus. The benthos underneath Salvinia mats was suppressed (McLachlan, 1969).
On Lake Kainji, a reservoir on the Niger River in Nigeria, small amounts of Echinochloa sudd developed in the northern part (Gaudet, 1979a). Studies on the significance of sudd for fish stocks in African man-made lakes are needed.
Sudd formation is also reported for lakes and reservoirs in Asia and the Pacific. In Rawa Pening in Indonesia, an old reservoirs constructed prior to the second World War, up to 60% of the surface area is covered by water hyacinth. In Bung Borapet reservoir, Thailand, an area of several km2 is covered with vegetation (Coix aquaticus, Isachne globosa, Leersia hexandra, Phragmites karka and Scirpus grossus), of which the first three plants form thick floating mats which are very resistant to wind and wave action. Junk (1977) found very low concentrations of dissolved oxygen and the presence of hydrogen sulfide underneath the mats, and he suggested such water is useless for fish, as it would be virtually devoid of invertebrate fauna. Eichhornia crassipes and Salvinia cucullata, also very abundant, drift around the lake together with the floating islands. In Sirinumu reservoir, which flooded both tropical forest and rubber plantation, and now provides drinking water for Port Moresby, the capital city of Papua New Guinea, suddification took place among the flooded trees (Photos 15 and 16).

PHOTO 15: Stumps of rubber trees in Sirinumu reservoir, Papua New Guinea.

PHOTO 16: Sirinumu reservoir - suddification.
In the Amazon Basin in South America, non-attached sudd formations are called ‘floating meadows’ (see the following Section).
White-water rivers, mainly the Solimoes/Amazonas, harbour fish which are an important source of protein for the local population. Varzea floodplains contain a dense aquatic vegetation called ‘floating meadows’, composed of the grasses Paspalum repens and Echinochloa polystachya, interspersed with several other plant species. During the high water season lakes within the floodplain are connected by channels to the main rivers and the grasses form floating mats with submersed mass of rhizomes and roots which support a rich invertebrate fauna. These, together with detritus, provide an important food for small fish species and juveniles of important commercial species such as migratory characins. Characins spawn in varzea lakes at the beginning of the flood and their juveniles inhabit and feed among the floating plants. The three common fish of white waters near Manaus. Schizodon fasciatus, Rhytiodus microlepis and R. argenteofuscus, are herbivorous (Mendes dos Santos, 1981), alongside of a number of other herbivorous and predominantly vegetarian fish species.
The submersed parts of floating meadows are used by the aquatic invertebrates as a substrate and partly for nutrition. In varzea lakes the density of aquatic invertebrates reaches 780 000 individuals m-2 (11.6 g m-2 dry weight, or 62 g m-2 fresh weight) at the edge of the stands (Junk, 1973). Towards the inner parts population density and biomass decrease. In very dense stands anoxia prevails. However, population densities and composition can vary strongly in monthly intervals, in different parts of a lake, and between varzea lakes (Junk and Robertson, 1997). The invertebrate fauna of the floating vegetation of the varzea is considered to be the richest in the Amazon. Under the influence of wind and flow the meadows may break into pieces ranging in size from a small clump to a football field and pass down the lakes to the main river channels (Henderson and Hamilton, 1995).
In Lago Mariraua, an upper Amazonian floodplain lake of varzea type, for anchored meadows, total wet weight of fish was estimated in the month of December as 312 kg ha-1 and the number as 90 400 individuals ha-1. In drifting islands, fish biomass and density were estimated as 192 kg ha-1 and 12 700 individuals ha-1, respectively (Henderson and Hamilton, 1995). According to Lowe-McConnell (1987) the fish standing crop in tropical fresh waters ranges from 55.7 to 5 116 kg ha-1. In Lago Mariraua the biomass estimate was probably below the annual maximum because it was made when the water was rising and just begun to enter the flood forest. At this time of the year many pieces of floating meadows are breaking free. Within the canal outflowing from the lake there was a continuous passage of at least 30 islands per day, with a total surface area ranging from 100 to 1 000 m2. Fish families Cichlidae and Anostomidae favoured marginal, anchored meadows, but were frequently dispersed by drifting islands. The synbranchids and gymnotids (swamp eels and knifefish) are the groups most adapted for life within floating meadows. None of these were caught within the drifting islands. This suggests that these fish are able to detect changes associated with the break-up of the meadows and move into secure areas. The Siluriformes favoured drifting islands over anchored meadows. For this group the downstream drift may represent a seasonal migration to the main river channel. Juvenile fish avoided drifting islands, presumably to avoid being flushed from the lake. It is estimated, that within the drifting islands 14 to 140 cichlids, 9 to 90 anostomids, and 74 to 740 catfish were transported daily out of the lake.
Aquatic macrophytes occurring in whitewater floodplains in large quantities are of poor nutritional value in comparison with other food items (Junk et al., 1997). Stomach contents of 91 species of fish, analysed for food from varzea and igapo (black waters) did not show any fish to be feeding to a major extent on aquatic macrophytes. Wherever possible, the fish prefer high-protein algae with a C:N ratio of about 9.5 and fruit rather than low protein macrophytes with a mean C:N ratio of about 25. Aquatic macrophytes are preferred by juvenile fish not because they are a good food value, but because they offer shelter and a large variety of food items, such as detritus, periphyton, perizoon, and terrestrial invertebrates. As the juveniles become larger, they move to other habitats, such as the open water or the floodplain forest and change their diet. The indirect importance of macrophytes as a food resource for fish, for example, in the form of macrophyte-feeding terrestrial and aquatic invertebrates, detritus and associated bacteria and fungi as well as in the form of dissolved organic carbon from decaying macrophytes via the microbial loop, has not yet been evaluated (Junk et al., 1997).
From the two freshwater streams in Costa Rica (not to be classified as varzea system) investigated for fish, in the larger one fish were found to be closely associated with dense mats of floating aquatic plants Eichhornia, Hydrocotyl, Salvinia and Azolla. Three eleotrid fish species Dormitator maculatus, Eleotris pisonis and E. amblyopsis were almost always captured from the dense beds of floating macrophytes that lined the stream margins and sometimes completely covered the channel (Winemiller and Ponwith, 1998). The two Eleotris and to a lesser extent D. maculatus, were also occasionally captured from submersed stalks of immature water hyacinth. Juvenile Gobiomorus dormitor, the fourth eleotrid, were rarely captured in aquatic vegetation, although individuals of all sizes were sometimes captured near or underneath patches of floating plants. G. dormitor was the only eleotrid associated with more open habitats and was collected in the channel area. D. maculatus feeds mostly on coarse detritus and minor amounts of filamentous algae and diatoms, while both Eleotris species and G. dormitor were carnivorous, feeding largely on shrimp and small fish. There was a shift in the food as the fish grew bigger. Diet similarity among the three carnivorous eleotrids declined during the wet seasons, the period when their prey disperse at lower per-unit-area densities within the flooded forest that surround the streams.
Black and clear water regions of the Amazon Basin also contain floating meadows of grasses (Graminae), but these are less developed and diverse than those of white waters. This may be responsible for the better dissolved oxygen conditions in varzea lakes as compared to blackwater lakes (Fig. 35). Araujo-Lima et al. (1986) studied the fish composition underneath two floating plants communities, e.g. pure Oryza sp., and a mix of Oryza sp. and Paspalum repens, in a blackwater floodplain lake of the Rio Negro. The main difference between these two microhabitats was the complexity of the submersed root system. A higher concentration of fish was found under mixed vegetation. Younger fish of Metynnis cf. hypsauchen fed predominantly on Cladocera, Chironomidae and Trichoptera, switching over to filamentous algae and wild rice seeds. The food of Moenkhausia lepidura was similar, but this fish kept insects as food when it grew larger. The third species, Hemigrammus levis, fed mainly on Cladocera when young, changing more to chironomid larvae and other insects when larger.
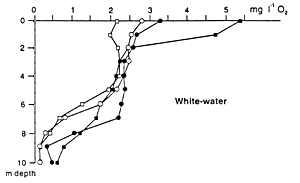
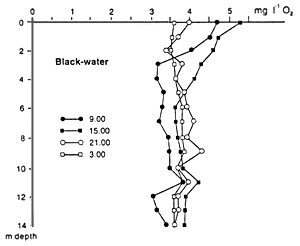
Fig. 35. Dissolved oxygen concentrations at different times of day in the white-water varzea lake Lago Inacio, and a black-water lake Lago do Prato in the Anavilhanas, Rio Negro, close to Manaus during high water level. (Junk et al., 1997).
Due to the lower biomass of aquatic plants and phytoplankton in the blackwaters as compared to whitewaters, fish in the Rio Negro depend much more on the flood forest than on the floodplain (Junk et al., 1997). Adult fish tend to overcome food shortages by their ability to feed on diverse items. Catch per unit of effort (gillnet) was on average 210 g m-2 day-1 in varzea as compared to 41 g m-2 day-1 in the igapo, and the catches are highest during the falling and low water levels (Fig. 36).
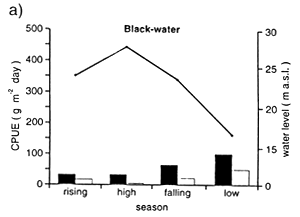
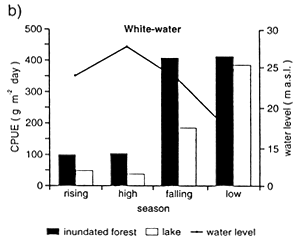
Fig. 36. Seasonal variations of catch per unit effort (CPUE) a) in the black-water lake Lago do Prato (Anavilhanas) and b) the white-water varzea lake Lago Inacio during 1990 and 1991 inside and outside a floodplain forest. (From Junk et al., 1997).
Due to natural diurnal fluctuations in dissolved oxygen content within and underneath macrophyte stands, most fish regularly leave the aquatic plants during the night and seek out open waters (Fig. 37). Most Amazonian fish are resistant to low oxygen concentrations, with 36 out of 43 species tested in experiments able to tolerate concentrations lower than 0.5 mg 1-1, at least for some hours (Junk et al., 1997). By about 8 a.m., when the dissolved oxygen content in macrophyte stands has risen as a result of the photosynthetic activity of phytoplankton and periphyton, the fish return (Fig. 38).
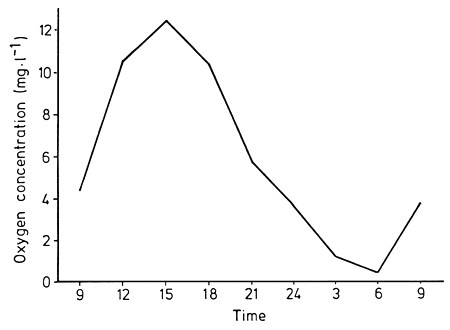
Fig. 37. Diurnal curve of dissolved oxygen concentrations underneath floating macrophytes in a varzea lake. (From Jedicke et al., 1989).
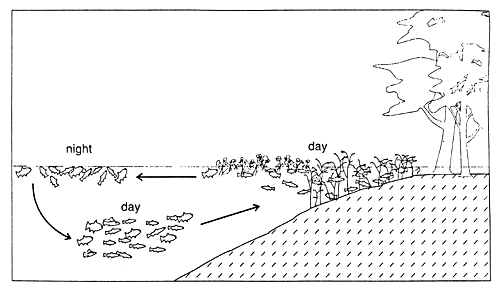
Fig. 38. Pattern of diurnal migrations of fish between macrophyte stands and open water in strongly hypoxic conditions. (From Junk et al., 1997).
The vegetation of the islands in igapo is characterized by flood forests. Most of the plant species display marked periods of leaf production and fall. The period of highest litter production is during the flood months, the maximum being in July. The expansion of the littoral zone from the main channel into the forests is accompanied by the expansion of the benthic fauna. During the flood period the highest numbers of shredders and macrocollectors are recorded from the igapo. Macroinvertebrate scrapers are limited to the rising and falling water periods (Fig. 39). In igapo, as on other types of floodplains, the main factor governing the aquatic productivity is the flood pulse (Nessimian et al., 1998). In this environment, the benthic fauna represents perhaps the most effective component in the energy flow between the aquatic and terrestrial compartments. Large river water level fluctuations force the fish to have flexible feeding and other habits. Characoid fish species, with their fast growth, early maturity, and high reproductive rates are well adapted to such changes and represent considerable part of fish biomass in varzea lakes. The blackwater river floodplain of the Rio Negro igapo system, in spite of having a higher fish species diversity, has a low fish biomass, which is only about 20% of that found in the whitewater floodplain of the Amazon River (Junk et al., 1997),
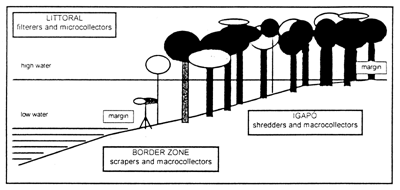
Fig. 39. Distribution of the major feeding categories of aquatic macroinvertebrates in igapo flood forest (Rio Negro floodplain). (From Nessimian et al., 1998).
![]()
![]()
![]()