![]()
![]()
![]()
Models in fisheries management
The fishery for the Brazilian sardine
The signs of human dominance and impact on the oceans have increasingly urged fisheries managers world-wide to account for the ecosystem impacts of fishing activities. The effect of fisheries on ecosystems are usually classified as direct and indirect impacts (Botsford et al., 1997; Goñi, 1998). Direct impacts include overfishing, by-catch and discard of non-target species, changes in genetic diversity of stocks, physical disturbances and habitat destruction by fishing gears. Recent assessments of the status of world marine capture fisheries revealed, for instance, that over 60% of the most important fish stocks are either overexploited or on the verge of becoming overexploited by current fishing intensity (Garcia and Newton, 1997), and that approximately 27 million tons of nontarget animals are discarded annually as “trash” fish (Alverson et al., 1994). Fisheries can potentially affect the genetic diversity of populations by selectively removing older and larger individuals (Goni, 1998), and by depleting small reproductive stocks of species such as salmon and herring (Policansky and Magnuson, 1998). Ultimately, the decrease in genetic diversity by fishing can cause loss of resilience of fish populations to both human and natural impacts. Fisheries are also responsible for physically damaging important marine habitats, specially with towed gears, i.e., otter trawlers, beam trawlers, and dredges (Hall, 1999). The effects of trawling on the sea bed vary from destruction of suitable habitats for the settlement of juvenile and adult phases of diverse marine organisms, changes in abundance and species composition of benthic communities, and concomitant changes in the fish species composition associated with the physical alteration of bottom structures (Goñi, 1998).
Indirect ecosystem impacts of fisheries are at least partly mediated through the food web, where the effects of fishing can ‘cascade’ to other components of the system. Marine ecosystems are structured in a complex array of predator-prey interactions, and by removing prey and/or predators from the system fisheries can affect the abundance of other species and impact the overall resilience of ecosystems. The ecosystem-wide effects of catching fish attracted little attention and probably had minor importance before the advent of large-scale industrialized fisheries. But, today, the scale of fisheries catches can considerably alter the structure of marine food webs. Heavy commercial fishing has often been associated with drastic changes in species composition in marine communities. Examples of major changes in marine ecosystems with intense fishing are present in almost every region of the world (Goñi, 1998; Hall, 1999), although in many cases it has proved difficult to separate the natural and anthropogenic causes of changes (Steele, 1998). Among the best documented examples are the cases of switches in dominance between sardines and anchovies in coastal upwelling (Lluch-Belda et al., 1989; Bakun, 1996). In the Bering Sea, human exploitation of whales and other top predators is thought to be responsible for cascading effects on other components of the ecosystem, such as declines in sea lions and seals, and the dominance of groundfish species such as pollock, Theragra chalcogramma, and large flatfishes (NRC, 1996; Trites et al., 1999). On Georges Bank, large scale disturbances caused by intense fishing and habitat destruction were also associated with apparent replacement of gadids and flounders by species of low commercial value, including dogfish sharks and skates (Fogarty and Murawski, 1998). Also, it is suspected that present exploitation patterns are resulting in a “fishing down marine food webs” phenomenon, by which heavy commercial fisheries are causing a progressive simplification of marine ecosystems in favor of smaller, high-turnover, lower-trophic-level fish and invertebrate species that are adapted to withstand disturbance and habitat degradation (Pauly et al., 1998; Pitcher and Pauly, 1998).
While evidence of ecosystem impacts of fisheries accumulates throughout the world, we see the development of fisheries management principles and guidelines which acknowledge the need to place fisheries in an ecosystem context (FAO, 1997; GESPE, 1997; Canadian Ocean Act; USA Magnuson-Stevens Act). FAO (1997) provided technical guidelines for the implementation of responsible fisheries, which make explicit the need to account for the ecosystem-scale effects of fishing:
“Responsible fisheries management should consider the impact of fisheries on the ecosystem as a whole, including its biodiversity, and should strive for sustainable use of whole ecosystems and biological communities.” “Harvesting any one species is almost certain to impact others,[...], the impact of ecological linkages (e.g. through the trophic chain) between species, may lead to changes in species dominance and affect the dynamic equilibria of the resource system, potentially affecting future options. These multi-species effects need to be considered in responsible fishing, which should aim to ensure that no species, whether targeted, by-catch or indirectly affected by fishing, is reduced to below sustainable levels.”The Magnuson-Stevens Fishery Conservation and Management Act (1996), which is the basis of fisheries management in U.S. federally managed waters, is one overarching example of mandated application of the principle of ‘ecosystem management’. The act maintains that fisheries are to be conserved and maintained to protect the marine ecosystem and to ensure the greatest overall benefit. Likewise, in Brazil, since the Earth Summit in 1992, an inter-institutional Fishing Sector Executive Group (GESPE) was created to implement a National Fishing and Fish Farming Policy plan aimed at fostering a better integration among institutions (public and private) for the reorganization of fisheries management in Brazil. The plan recognizes as the ultimate goal of fisheries management the need to ordinate and foster the sustainable use of fisheries resources, which is broadly defined as the maintenance of the ‘equilibrium’ of ecosystems and preservation of the species under exploitation; economic profitability of fishing activities; generation of jobs and a fair work compensation (GESPE, 1997).
Scientific advisory work is therefore compelled to consider far as possible the more diverse aspects or consequences of fishing activities for marine populations and ecosystems. Mangel et al. (1996), for instance, proposed a set of principles for the conservation of wild living resources which explicitly recognize the need for scientific account of human impacts on ecosystems, e.g.
“The goal of conservation should be to secure present and future options by maintaining biological diversity at genetic, species, population, and ecosystem levels; as a general rule neither the resource nor other components of the ecosystem should be perturbed beyond natural boundaries of variation.” “Regulation of the use of living resources must be based on understanding the structure and dynamics of the ecosystem of which the resource is part and must take into account the ecological and sociological influences that directly and indirectly affect resource use.”The adoption of these guidelines and principles will require a critical re-examination of concepts, methods, tools, and regulatory measures commonly used in fisheries assessment and management. Although the need for ecosystem management has been widely recognized, scientific advice is still hampered by the lack of understanding of the complex dynamics of ecosystems, and the lack of consensus on which framework should be used to account for the ecosystem effects of fisheries. Despite the reality that fisheries are generally not restricted to affecting one species alone, the development of single species models and procedures for fisheries assessment and management has centered around that very assumption. And, due to our lack of ability to model complex systems, such methodology is still prevalent.
One of the major tasks of fisheries science is to estimate the tradeoffs of management choices (Hilborn and Walters, 1992). Also, as a scientific activity, fisheries research involves a process of learning about nature in which alternative ideas, or hypotheses, are contrasted against observations and experiences, so that a better understanding of the potential responses of systems being managed can be achieved through time. In these activities models are usually employed as tools to test our understanding of the mechanisms in the system, to provide the basis for both understanding of known patterns and prediction about situations not yet encountered, and to provide the means for evaluating the potential effects of various kinds of decisions (Hilborn and Mangel, 1997).
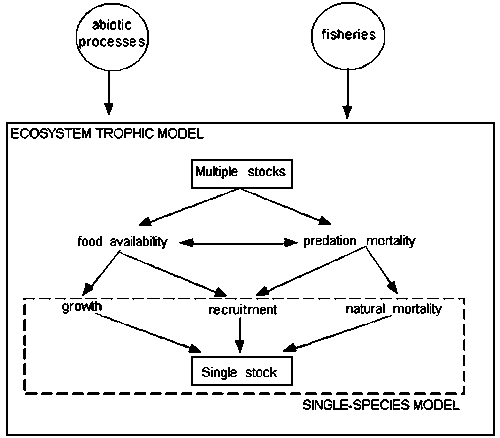
Fisheries assessment can be based on two types of modeling approaches: single-species and ecosystem or multi-species models (Figure 1). Single-species models are based solely on the analysis and representation of population processes such as growth, mortality and recruitment, and the impact of fishing on these, and have been the most widely applied approach to provide advice on the consequences of fishery regulatory tactics. Several processes can influence the productivity of an exploited fish population. Hilborn and Walters (1992) argued that some of environmental and multispecies effects on a stock are likely to be most important during the fish early life stages, when larvae and juveniles are more vulnerable to sub-optimal environmental conditions, predation and competition for food. For convenience, in a single species model the net effect of juvenile survival (i.e., recruitment) is usually represented by a stock-recruitment relationship. The focus on stock-recruitment relationships has also a practical reason, since it directly links a control variable (stock) and a rate of future stock production (recruitment).
By definition, ecosystem models consider not only the biological processes of the target population but also the ecological (e.g. predation, consumption, competition) and technological (e.g. by catch) interactions with other species in the ecosystem. Attention during the last decades has been given to the development of ecosystem models that describe patterns of trophic interactions in the food web, mainly represented by multispecies virtual population analysis, MSVPA (Sparre, 1991), and ecosystem mass-balance models such as Ecopath (Polovina, 1984; Christensen and Pauly, 1992). The implementation of the first type of modeling tool has been hampered in part by the need for extensive time series of catch-at-age data, difficult parameterization, the high degree of expertise required from the modeler, and the overall lack of transparency in the estimation procedure (Walters et al., 1997). Ecopath offers, on the other hand, a simpler approach for the reconstruction of trophic interactions in fished ecosystems, and has been widely applied to aquatic ecosystems (more than 100 Ecopath models have been published world-wide describing upwelling, shelf, lake, river, open ocean, and terrestrial farming systems; see the Ecopath homepage at http:\\www.ecopath.org). The approach has some advantages over other existent trophic models viz., it includes all trophic levels in the analysis (from primary producers to top predators) as opposed to, as in the case of MSVPA, focusing only on the commercially important fish species; the emphasis on ecological relationships makes it intuitively simple; it provides a means to incorporate and standardize large amounts of scattered information routinely collected by fisheries scientists and marine biologists (Pauly et al., 1998); and, more importantly, the widespread use of the approach creates opportunity for comparative studies of ecosystem’s response to fisheries impact. Moreover, further developments of the mass-balance model, which originally focused on describing systems at steady-state conditions, has resulted in a dynamic ecosystem model called Ecosim (Walters et al., 1997; Christensen and Walters, 2000) that is capable of answering “what-if” questions about policy and ecosystem changes that would cause shifts in the balance of trophic interactions.
Ecosystem models are in their infancy and have been little used to date in fisheries management. Ecosystem models have been proposed as tools for guiding the implementation of ecosystem principles in fisheries management, but exactly how the approach could be used, what are their advantages, limitations and complementary role to traditional single-species approaches is still unclear. Such comparisons have not been performed explicitly before. This work uses the fishery for the Brazilian sardine in Southeastern Brazil to analyze the use of a single-species and an ecosystem model in the assessment and management of a capture fishery. In this analysis models are used to assess and represent key ecological processes controlling the dynamics of the population and ecosystem, and to evaluate the likely effects of fisheries management decisions through model simulations.
Figure 1. Biological processes represented in fisheries assessment models. The diagram shows the increasing level of complexity when one goes from a single species-model (dashed box) to an ecosystem model. Ecosystem models attempt to represent all important ecological processes affecting production, such as food availability and predation mortality, while single-species models represent the net effect of these and other ecological processes on growth, recruitment and natural mortality. Both models represent the impacts of fisheries and abiotic variables on fish stock(s).
Sardine, Sardinella brasiliensis, inhabit coastal waters from Cabo Frio in the north to Cabo de Santa Marta Grande in the south of the Southeastern Brazilian Bight (Figure 2). The main environmental characteristic of the region is the seasonal presence of cold, nutrient-rich South Atlantic Central Water on the inner shelf, i.e. 10 to 50 m deep (Pires-Vanin and Matsuura, 1993). Sardine spawning occurs mainly during spring and summer, in the enriched environment formed downstream of the upwelling center of Cabo Frio (Bakun and Parrish, 1990).
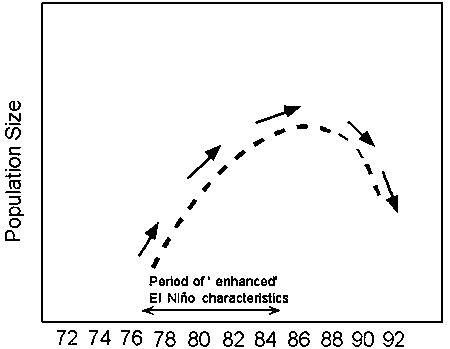
The Southeastern Brazilian Bight ecosystem is conditioned by cyclical oceanographic events that control changes in the patterns of energy flow and the productivity of biological communities (Pires-Vanin et al., 1993; Vasconcellos, 2000). The seasonal upwelling cycle is nested in a hierarchy of environmental cycles with different time scales, ranging from days to decades. On one hand, physical processes such as pulses of favorable wind stress, storms, and small scale eddies with low residence time, all influence biological processes at short time scales. Cycles in marine fish populations bear, on the other hand, close correspondence to the long term climatic-oceanographic regimes of the oceans (Steele, 1985; Lluch-Belda et al., 1989; Bakun, 1996). Bakun (1996), for instance, compiled examples of synchronous decadal oscillations in several fish populations that were geographically isolated from each other, but apparently driven by a common environmental property (periods of intensification of the El-Niño). These low-frequency population cycles varied among species, but were generally characterized by a period of rapid population growth in the decade from the mid-1970s to the mid-1980s followed by stock declines after the mid-1980s. According to Bakun the Brazilian sardine followed the pattern of “crashing” after the mid-1980s (Figure 3). The dynamic characteristics of the environment and resource have important consequences for fisheries management and indicate the need to develop robust strategies for fisheries viability under conditions of radically varying resources (Bakun, 1998).
Figure 3. Pattern of variation observed in many marine fish populations according to the “Dome-shaped” hypothesis (source Bakun, 1996). Populations following the ‘dome-shaped’ curve include sardine of Japan, Peru-Chile and California, anchovy of Beguela. Populations following an opposite phase include anchovy of Peru-Chile, California and Japan, sardine of Benguela. Populations that crashed after the mid-1980s include the Brazilian sardine, and Balistes sp. of west Africa.
Until the early 1900s sardine catches were mainly by artisanal fishers and used as food by coastal communities (Diegues, 1995). This artisanal fishery still exists in most states, catching sardines in bays and estuaries along the coast, using cast nets and seine nets. Today, most of the sardine catches come from an industrial fishery based on purse seiners. The first purse seiners appeared ca. 1910 and gradually diverged from the artisanal and small scale sector, mainly during the 1930’s with the introduction of power engines (Diegues, 1995). During 1967-1978 government tax incentives (Código de Pesca, Lei 221, 1967) attracted a considerable amount of capital to the fishing sector which expanded the number of industries for catching and processing fish products for export, and resulted in unprecedented changes in the fishery. Sardine became the main Brazilian fishery resource in terms of volume, with total annual catches increasing from ca. 38 thousand tons in 1964 to an historical peak of 228 thousands tons in 1973 (Figure 4). Average landings during 1983-1987 were 124 thousand tons.year-1, accounting for 31.8% of the total fish catches in the region and about 25% of the total Brazilian marine catch (IBAMA, 1995).
Figure 4. Sardine landings and the reconstructed stock biomass in the Southeastern Brazilian Bight
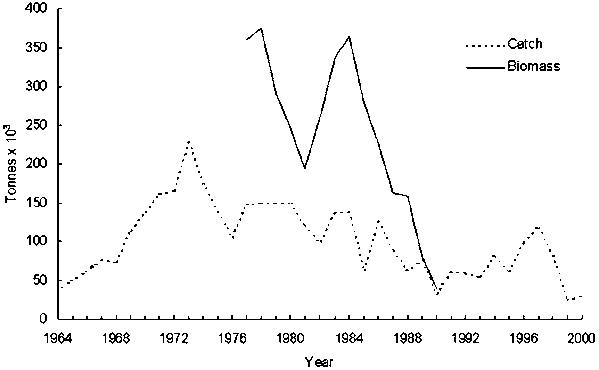
Another program of government incentives aimed at the modernization of the fleet lasted from 1983 to 1985. It resulted in an increase in fleet capacity (tonnage) of approximately 300% compared to the 1970’s, a doubling of the number of fishing vessels, and the introduction of technological innovations (e.g. sonar and power block) that considerably increased the fishing power of purse seiners (Valentini and Cardoso, 1991). The fishery then experienced a marked decrease in catches, mainly from 1987 to 1990, which culminated in the collapse of the stock and a crisis in the commercial/industrial sector (IBAMA, 1995). It is estimated that between 1977 and 1990 the stock biomass declined from ca. 350,000 tonnes to less than 80,000 tonnes, with an average fishing mortality rate of 0.8 year-1 (Figure 4; Vasconcellos, 2000). Since the collapse of the fishery there is no indication of a significant stock recovery (Habiaga, 2000). Catches increased from 1992 to 1997 reaching ca. 118,000 tonnes, but decreased again to less than 30,000 tonnes in 2000 (Cergole, 2000).
As has occurred in some other marine pelagic ecosystems subjected to intense fishing, the decline in sardine biomass was followed by a marked increase in the biomass of anchovy, a small pelagic species also abundant in the area (Castello et al., 1991). The decrease in sardine biomass was also accompanied by an increase in catches of triggerfish, Balistes capriscus, (Zavalla-Camin and Lemos, 1997), which is a semi-pelagic species as adult and pelagic planktonic feeders as juveniles. The understanding of the causes of stock collapse and ecosystem changes has been of particular concern for sardine stock assessment and management (Castello et al., 1991; Saccardo and Rossi-Wongtschowski, 1991; IBAMA, 1995; and Rossi-Wongtschowski et al., 1996).
The Southeastern Brazilian Bight also has important demersal fish stocks that are targeted by trawling fisheries. The main demersal stocks exploited in the region are the white croaker Micropogonias furnieri, king weakfish Macrodon ancylodon, weakfish Cynoscion jamaiscencis, triggerfish and the marine shrimps Xiphopenaeus kroyeri, Farfantepenaeus brasiliensis and F. paulensis. Offshore fisheries for tunas, sharks and rockfishes were developed during the last two decades following the depletion of many traditional stocks. Tunas and sharks are mainly caught with longlines and pole-and-line, while bottom longlines are employed in the rockfish fisheries (Gasalla and Tomás, 1998).
![]()
![]()
![]()
{kind=link}