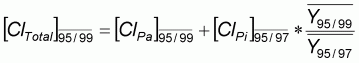![]()
![]()
![]()
Aboubacar Sidibé[15], Didier Gascuel[16] et François Domain[17]
Introduction
En Guinée, les espèces démersales côtières, économiquement importantes font l'objet d'une exploitation de plus en plus intense, à la fois par la pêche artisanale et par la pêche industrielle. Aujourd'hui, il est important d'assurer un meilleur suivi de ces ressources dans leur exploitation et dans leur gestion. Pour atteindre cet objectif, des évaluations de stocks sont réalisées avec des données disponibles pour établir des diagnostics pour les principaux stocks. C'est dans ce cadre que se situe le présent travail réalisé sur quatre stocks de poissons démersaux côtiers: Galeoides decadactylus, Pseudotolithus elongatus, P. typus et P. senegalensis. Les données et les méthodes utilisées sont d'abord présentées; les résultats issus de l'analyse des cohortes sont présentés en terme d'effectifs, de biomasses et de mortalités par pêche correspondantes. Enfin, des analyses prospectives en terme de rendement par recrue et de simulation de diagramme d'exploitation par pêcherie (pêche artisanale et pêche industrielle) sont faites et les résultats discutés dans une perspective de gestion et d'aménagement.
1. Données et méthodes utilisées
1.1 Données disponibles
Les données statistiques des pêcheries guinéennes proviennent du système d'enquête du CNSHB (Centre national des sciences halieutiques de Boussoura) et sont relativement récentes. Elles ne sont disponibles qu'à partir de l'année 1995 et concernent notamment les captures, les débarquements, les efforts de pêche, les fréquences de tailles et certaines caractéristiques de l'exploitation halieutique. Les données sur les espèces ou catégories d'espèces économiquement importantes dans la ZEE guinéenne, sont estimées par pêcheries et pour chacune des années 1995, 96, 97, 98 et 99.
Pour la pêche artisanale, l'estimation s'appuie sur un recensement annuel exhaustif des pirogues actives sur l'ensemble du littoral et sur des observations concernant des sorties de pêche et des débarquements, selon un plan d'échantillonnage stratifié par région administrative et par engin de pêche. Pour la pêche industrielle, les prises sont estimées à partir d'un suivi exhaustif des activités (jours de pêche) de l'ensemble des navires autorisés à pêcher dans les eaux guinéennes et à partir des données collectées par un réseau d'observateurs embarqués sur un échantillon de navires.
Au niveau de la flottille artisanale, dans la mesure où les rejets sont négligeables, les données mensuelles des structures de tailles des débarquements sont assimilées aux captures. Ces données sont estimées par zones géographiques et par engins. Les mesures de taille (environ 10 individus de chaque espèce par débarquement observé) sont dans un premier temps compilées par engin et par mois. Elles sont ensuite extrapolées aux captures totales par engin et par mois, sur la base du rapport poids des individus mesurés/poids des captures totales. Cette démarche présente l'inconvénient de ne pas tenir compte de l'effectif réel dans le débarquement observé et donne la même importance aux structures de taille provenant de petits ou de gros débarquements. Elle se justifie néanmoins si on considère les poissons mesurés comme étant issu d'un échantillonnage aléatoire simple, au sein de la population de l'ensemble des poissons capturables.
L'échantillonnage à bord des navires industriels pour l'estimation des histogrammes de taille au niveau des captures conservées est largement décrit par Lesnoff et al.., (1995). Une espèce choisie dans le trait échantillonné est mesurée et comptée. Pour chaque taille conservée, une extrapolation à l'ensemble du trait du jour est effectuée (tous les traits du jour n'étant pas échantillonnés). Les rejets échantillonnés par les observateurs à bord des navires n'incluent pas la capture totale correspondante, par conséquent il n'est pas possible de faire une extrapolation directe sur la base des statistiques disponibles. Pour corriger cette situation, une démarche consistant à estimer un taux de rejet moyen global par type de licence de pêche à partir des informations sur les traits échantillonnés a été adoptée (Sidibé, 1998). Les effectifs par classe de taille dans les rejets des traits observés ont donc été extrapolés aux rejets totaux estimés à partir des captures conservées des bulletins statistiques (Anonyme,1995-1999) et du coefficient du rejet estimé par Sidibé (1998).
L'utilisation de telle méthode d'estimation requiert une hypothèse contraignante. Celle de l'existence d'un diagramme de capture industriel standard, hypothèse manifestement simplificatrice puisque tous les bateaux industriels ne pratiquent pas les mêmes pêches, pas avec les mêmes types d'engins, pas sur les mêmes zones. De plus, leurs diagrammes de rejets seront différents selon que les quatre espèces soumises à l'étude ici, font partir des espèces cibles, des prises accessoires ou des rejets. Les céphalopodiers et les crevettiers par exemple, rejettent beaucoup d'individus de grande taille qui sont logiquement conservés par les poissonniers démersaux.
Les effectifs capturés annuellement sont donc estimés par classe de taille, pour la pêche artisanale et pour la pêche industrielle. Compte tenu du petit nombre d'années disponibles pour les quatre espèces, une première analyse est conduite sur une pseudo-cohorte. Les effectifs sont ainsi regroupés pour obtenir une distribution des captures totales par classe de taille, en valeur moyenne sur les cinq années (1995, 1996, 1997, 1998 et 1999) à partir de la relation suivante:
|
|
(1) |
|
où: |
ClTotal =Capture totale moyenne
1995/1999 |
C'est sur cette dernière distribution, que les calculs sont effectués, conduisant à estimer l'effectif du stock, les coefficients de mortalité par pêche et les rendements par recrue.
Il est important de rappeler ici que les quatre stocks soumis à l'analyse dans ce travail sont définis comme unités de gestion en référence à la zone guinéenne s'étendant de 9° Nord au 11° Nord. Ils doivent être considérés comme isolés spatialement et biologiquement par rapport aux stocks de la Guinée-Bissau au nord et de ceux de la Sierra Leone au sud; et que la pêche dans ces zones n'a aucun effet sensible sur leur évolution en Guinée.
1.2 Conversion des données de structure de taille en âge (décomposition polymodale)
Par ailleurs, les effectifs capturés annuellement sont également estimés par groupe d'âge pour chaque espèce étudiée. La conversion taille/âge est effectuée par décomposition polymodale, sous l'hypothèse d'une loi de croissance déterminée. Etant donné qu'aucune différence de croissance n'est observée entre les femelles et les mâles au niveau de ces espèces, les estimations sont effectuées sans distinction de sexe.
Les paramètres de croissance utilisés pour les quatre espèces sont donnés ci-dessous. On admet ici que le to est estimé pour un âge compté en âge absolu (compté depuis la naissance).
. Galeoides decadactylus: L¥ = 47,00 cm; K = 0.161 an-1; to = - 1,63 an (Samba, 1974);
. Pseudotolithus elongatus L¥ = 51,76 cm; K = 0,256 an-1; to = - 0,44 an (Le Guen, 1971);
. Pseudotolithus senegalensis: L¥ = 52,70 cm; K = 0,350 an-1; to = - 0,65 an (Troadec, 1971);
. Pseudotolithus typus: L¥ = 89,70 cm; K = 0,175 an-1; to = - 1,033 an (Poinsard, 1973).
Les captures aux âges on été estimées en utilisant le Programme Normsep (Tomlinson, 1970). En effet, le Normsep (Normal Distribution Separator Using Steepest Descend Methode) permet une conversion des données structurées en taille selon l'hypothèse d'une distribution normale des individus en fonction de l'âge pour une taille donnée. On utilise ici un ajustement sous contrainte: la taille moyenne aux âges et l'écart type de chaque mode sont préalablement fixés en fonction de la loi de croissance retenue (Gascuel, 1994). Pour chaque sortie Normsep, on obtient l'effectif de chaque groupe d'âge. Le nombre de groupes d'âge est établi pour chaque espèce et le dernier groupe d'âge est considéré comme un groupe plus.
1.3 Mortalités naturelles
La mortalité naturelle de chaque espèce est estimée par différentes méthodes (Taylor, 1960; Pauly, 1980b; Caddy, 1991). D'autres valeurs de mortalités naturelles issues des travaux antérieures réalisés dans la région sur les même espèces (Caverivière et Thiam, 1994; Le Guen, 1971) sont comparées à celles estimées dans le présent travail (tableau 1).
La mortalité naturelle est ici considérée constante avec l'âge et avec les années. Les mortalités naturelles estimées par la méthode de Caddy (1991) sont utilisées dans la suite des calculs. Les autres valeurs de M serviront à l'étude de sensibilités des résultats des analyses.
Tableau 1: Estimation des différentes mortalités naturelles.
|
|
M (Caddy) |
M (Pauly) |
M*/2 (Pauly) |
M (Taylor) |
Travaux antérieurs |
|
G. decadactylus |
0,25 |
0,45 |
0,23 |
0,18 |
0,25 (Caverivière et Thiam, 1994) |
|
P. elongatus |
0,28 |
0,60 |
0,30 |
0,27 |
0,31 (Le Guen, 1971) |
|
P.senegalensis |
0,21 |
0,73 |
0,36 |
0,38 |
0,30 (Caverivière et Thiam, 1994) |
|
P. typus |
0,28 |
0,40 |
0,20 |
0,19 |
|
M*/2 (Pauly): En considérant la remarque faite par Sparre et Venema (1996), les résultats issus de la méthode de Pauly ont été divisés par 2. En effet ces auteurs notent que dans le cas de certaines espèces le résultat de Pauly semble être le double de ce qu'il devait être. Ce qui pourrait être le cas de ces quatre espèces compte tenu de leur longévité relative (supérieur à 5-7 ans).
Enfin, il est important de rappeler ici que les estimations de ces différentes mortalités naturelles sont fondées sur les méthodes purement empiriques. La plupart d'entre elles, pour ne pas dire toutes, ne dépassent guère le niveau de la supposition. Ces valeurs ne peuvent donc, en aucun cas être prises comme des valeurs de références objectives, mais doivent rester des hypothèses dont il faudra déterminer l'impact sur les résultats de l'analyse de pseudo-cohorte.
1.4 Rappel du principe de l'analyse de cohorte ou V.P.A. (Virtual Population Analysis)
Au cours d'une analyse de cohorte ou de pseudo-cohorte, l'estimation des effectifs du stock à chaque taille ou à chaque âge ainsi que celle des coefficients instantanés de mortalité par pêche correspondants se fonde par la résolution d'un système à deux équations (équation de survie et équation de capture). Ces deux équations ont été largement utilisées (Beverton et Holt, 1957; Gulland, 1969; Jones, 1983; Mesnil, 1988). Dans le cas d'une analyse sur des données par classe de taille, elles se présentent sous la forme suivante:
Ni+1 = Nie-Zi (Dt) (2)
Ci = Ni(Fi/Zi)(1-e-Zi (Dt)) (3)
où i: indice de la classe de longueur; Ni: nombre d'individus dans le stock ayant atteint la longueur Li; Ci: nombre d'individus décédés suite à la pêche dont la taille est comprise entre Li et Li+1; (Dt): temps moyen qu'il faut à un poisson pour passer de la longueur Li à la longueur Li+1; Fi: coefficient instantané de mortalité par pêche de la classe i; Zi: coefficient instantané de mortalité totale; Zi = Fi + M; M est le coefficient de mortalité naturelle, supposé constant.
Pour n classes de taille, l'analyse s'appuie donc sur un système de 2n équations comportant 2n+1 inconnus (Les Ni, pour i = 1, n+1 et les Fi pour i = 1 à n).
Pour résoudre ce système, l'une des inconnues doit être fixée. Il s'agit généralement du coefficient de mortalité par pêche de la dernière classe de taille (FT: terminal). Les effectifs de la cohorte étant particulièrement difficile à évaluer directement, le coefficient de mortalité par pêche est le plus souvent estimé. C'est le processus d'initialisation de l'analyse de cohorte (Mesnil, 1988).
Dans la pratique cette initialisation s'effectue sur la classe de taille (ou d'âge) la plus grande de la cohorte et la résolution séquentielle du système se fait en remontant vers les classes de taille les plus petites. Jones (1983) a montré que si on procède de cette façon, l'erreur relative éventuellement commise en estimant le paramètre d'initialisation (FT terminal) tend à s'estomper au fur et mesure que l'on atteint les classes les plus petites; et que cette convergence est d'autant plus forte que le stock est fortement exploité.
Le choix de la valeur d'initialisation modifie donc très peu le diagnostic final par le fait de cette propriété de convergence des mortalités par pêche lors de l'analyse ascendante. Le choix du FT terminal dépend de critères logiques et de connaissances générales de la pêcherie.
La démarche adoptée ici pour l'estimation du nombre total moyen de poisson en mer, consiste à trouver l'effectif moyen dans chaque intervalle de classe et à le pondérer par le temps (Dt) passé dans cet intervalle. De ces effectifs du stock, issus de l'analyse des cohortes, on déduit une estimation des biomasses correspondantes. Sous hypothèse d'équilibre, la somme des biomasses par classe de taille ou groupe d'âge peut être considérée comme une estimation de la biomasse totale moyenne du stock, présente dans la ZEE guinéenne une année donnée. Ce calcul fait appel aux lois de croissance précédemment citées et la relation taille/poids (W = aLb). Les paramètres a et b de cette dernière relation pour les quatre espèces sont issus des travaux du CNSHB (cnshb, 1994):
a = 2,15.10-2 g/cm; b = 2,968 pour G. decadactylus
a = 0,55.10-2 g/cm; b = 3,140 pour P. elongatus
a = 0, 50.10-2 g/cm; b = 3,152 pour P. senegalensis
a = 0,67. 10-2 g/cm; b = 2,997 pour P. typus
L'estimation des effectifs du stock à chaque taille ou à chaque âge, ainsi que celle des coefficients instantanés de mortalité par pêche correspondants est donc conduite par les méthodes d'analyse des pseudo-cohortes en taille (Jones, 1983; De Verdelhan, 1987). Une analyse rectifiée des pseudo-cohortes en âges est également conduit (Laurec et Santarelli, 1986; logiciel Simuco, Mesnil, 1988). Cette méthode permet de s'affranchir des hypothèses de recrutement et d'effort de pêche stables (condition d'équilibre), qui sont mises en œuvre dans l'analyse sur fréquence de taille.
1.5 Correction des efforts de pêches
Pour s'affranchir de cette hypothèse d'équilibre mise en avant dans l'analyse sur structure de taille, on considère ici l'évolution des efforts de pêche théoriques estimés sur les mêmes stocks lors de l'analyse par l'approche globale (Sidibé et al., présent document). Pour minimiser d'éventuelles variations irrégulières et en tenant compte du nombre d'années disponibles (1995-1999), des moyennes mobiles d'ordre k = 5 sont calculées sur chaque série d'efforts théoriques et pour chaque espèce (tableau 2).
Tableau 2: Vecteurs des efforts de pêche théoriques issus du calcul de la moyenne mobile avec k = 5.
|
Années |
G. decadactylus |
P.elongatus |
P.typus |
P.senegalensis |
|
1994 |
3,0 |
7,9 |
6,7 |
6,4 |
|
1995 |
4,1 |
11,2 |
11,7 |
7,7 |
|
1996 |
5,0 |
13,1 |
13,6 |
7,9 |
|
1997 |
6,3 |
15,4 |
15,5 |
7,9 |
|
1998 |
7,2 |
16,8 |
17,9 |
8,4 |
|
1999 |
7,7 |
16,9 |
18,2 |
7,9 |
1.6 Diagnostic à long terme: interprétation et limites des rendements par recrue
Le principe de base d'un diagnostic de rendement par recrue est de permettre la quantification du potentiel de production d'un diagramme d'exploitation quelque soit le niveau du recrutement.
En d'autres termes, les rendements par recrue permettent de savoir si le régime d'exploitation permet de tirer un bon parti de chaque poisson entrant dans la pêcherie et si le recrutement est utilisé de façon satisfaisante.
En considérant une cohorte hypothétique entrant dans la pêcherie avec un recrutement quelconque R, on peut suivre, grâce à l'équation de survie, les effectifs de cette cohorte tout au long de sa vie. En connaissant en plus un vecteur de poids aux tailles (ou aux âges), l'équation de capture permet de calculer les captures pondérales et donc le poids total Y des captures que le diagramme d'exploitation aura permis de tirer du recrutement R. On peut alors calculer le rendement par recrue Y/R (en grammes ou kg par recrue) en divisant la production totale Y par R.
En faisant varier le vecteur de mortalité par pêche F (en utilisant par exemple des multiplicateurs mF du vecteur de référence), on peut tracer la courbe de rendement par recrue et prévoir quels effets sur la production par recrue aurait une modification du niveau global de l'effort de pêche. La courbe de rendement par recrue permet ainsi de définir le multiplicateur du vecteur de mortalité par pêche qui permettrait une maximisation de la production pour un diagramme d'exploitation donné.
Le calcul des rendements par recrue ne requiert aucune hypothèse contraignante autre que celles nécessaires à l'analyse des cohortes. Ni hypothèse de stabilité du recrutement, ni hypothèse d'équilibre de l'abondance du stock avec son milieu ne sont nécessaires. Ces hypothèses ont par contre un rôle évident dans l'interprétation des résultats obtenus.
Les calculs de biomasse, de biomasse féconde et de rendements par recrue sont ainsi menés à partir:
du vecteur F par classe de taille ou par groupe d'âge (valeur de référence 1995/1999),
du vecteur des poids à chaque taille ou à chaque âge, issu de la loi de croissance et de la relation taille/poids,
de la valeur de mortalité naturelle M précédemment citée,
de l'ogive de maturité des quatre espèces établie en Guinée par Domain et al. (2000a).
Il faut préciser ici que l'analyse fondée sur les distributions de fréquences de tailles a été privilégiée dans un premier temps car les données disponibles en Guinée se prêtaient le mieux à une telle analyse. L'hypothèse d'une mortalité par pêche constante ne peut cependant être admise pour ces espèces qui ont connu des efforts de pêche relativement croissants sur la période. L'analyse rectifiée des pseudo-cohortes en âge permet d'étudier l'impact de cette hypothèse d'équilibre sur les résultats du calcul des rendements par recrue.
2. Résultats
2.1 Structure démographique des captures
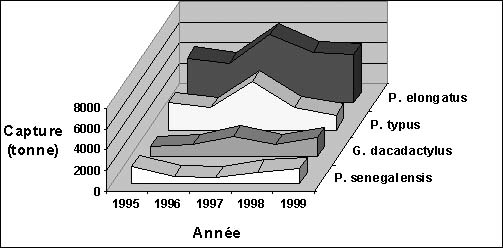
Les captures pondérales observées au niveau des quatre espèces varient énormément par année et par espèce. La séries de données étant relativement courte, il est difficile d'observer une quelconque tendance sur la période (figure 1). Si les captures de G. decadactylus et de P. senegalensis sont essentiellement réalisées par la pêcherie industrielle, le P. elongatus et le P. typus sont en revanche, plus ou moins exploitées par les deux pêcheries. Toutefois ces deux dernières espèces restent globalement plus capturées sur la période par la pêche artisanale que par la pêche industrielle (tableau 1, en annexe).
Figure 1: Evolution des captures des quatre espèces entre 1995 et 1999 (source: CNSHB).
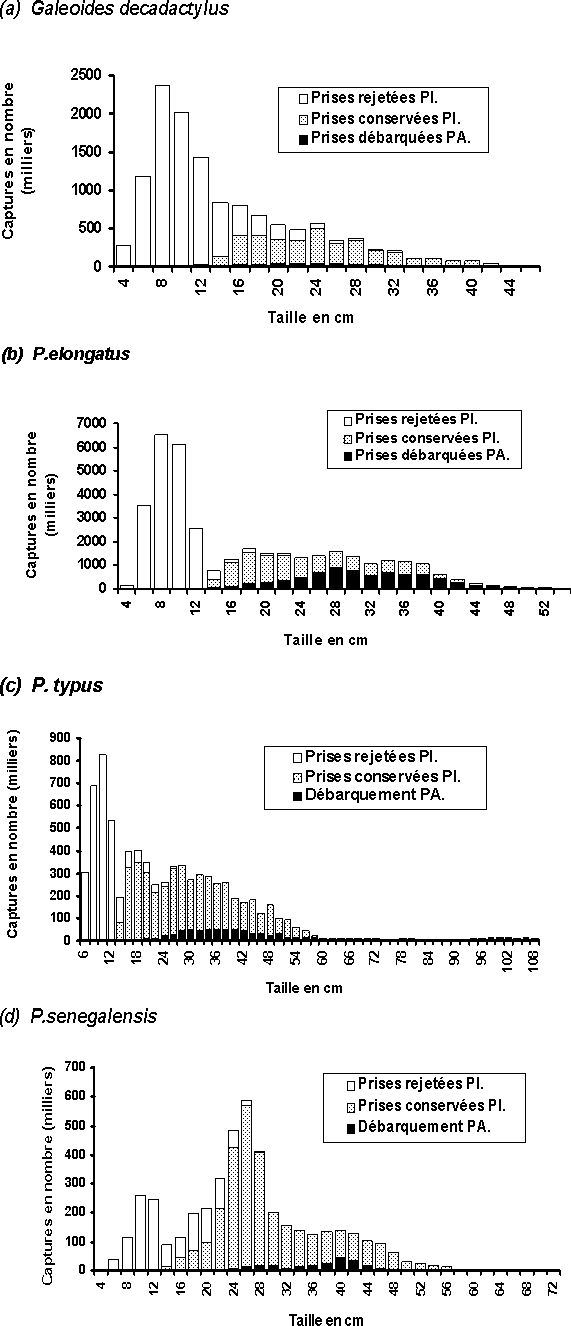
Différentes structures démographiques des captures sont également observées au niveau de chaque espèce exploitée (tableau 2, en annexe):
- Galeoides decadactylus (petit capitaine): La taille des petits capitaines capturés par la pêche artisanale et industrielle en Guinée est comprise entre 4 et 46 cm. L'essentiel des captures est constitué d'individus dont la longueur totale oscille entre 6 et 28 cm (figure 2a) et représente environ 91 pour cent du nombre total capturé. Plus de la moitié des captures, soit 71 pour cent, est rejetée à la mer et ceci, uniquement par les chalutiers industriels démersaux. Comparées à la pêche industrielle, les captures au niveau de la pêche artisanale sont très faibles, elles constituent seulement environ 10 pour cent des prises totales conservées.
- Pseudotolithus elongatus (bobo): La taille des bobos capturés par les deux types de pêche en Guinée est comprise entre 4 et 54 cm. La majorité des captures est constituée d'individus dont la longueur totale oscille entre 6 et 38 cm (figure 2b) et représente environ 95 pour cent du nombre total capturé. Un peu plus de la moitié, soit 53 pour cent des captures est rejetée à la mer par les chalutiers industriels.
Comparées à la production de la pêche artisanale, les captures de la pêche industrielle deviennent relativement moins importantes; en nombre, elles représentent aujourd'hui 26 pour cent des prises conservées, contre 29 pour cent de ces mêmes prises conservées en 1996 (Sidibé, 1998).
- Pseudotolithus typus (bar nanka): Les tailles du bar nanka capturés en Guinée varient de 6 à 108 cm. Les individus pêchés qui ont une longueur totale comprise entre 6 et 52 cm représentent 95 pour cent de l'effectif total capturé par les deux pêcheries (figure 2c). Une partie importante des prises totales, soit 35 pour cent est rejetée en mer toujours par la pêche industrielle. La pêche artisanale ne représente que 12 pour cent de l'effectif des prises conservées, mais les plus gros individus sont presque totalement capturées par cette pêcherie.
- Pseudotolithus senegalensis (bar sénégalais): Le bar sénégalais est pêché en Guinée à partir d'une taille de 4 cm, par la pêcherie industrielle uniquement. Les plus gros individus rencontrés dans les captures peuvent mesurer jusqu'à 100 cm de longueur totale. Toutefois, l'essentiel des captures est constitué d'individus dont la taille oscille entre 8 et 48 cm et représente environ 96 pour cent du nombre total capturé (figure 2d). Il faut noter qu'en moyenne 28 pour cent des captures totales sont rejetés en mer par la pêche industrielle. Le nombre d'individus capturés par la pêche artisanale est très faible, il ne représente que 6 pour cent des prises totales conservées.
Figure 2: Structure démographique moyenne 1995/1999 (en taille) des captures commerciales en Guinée du: (a) G. decadactylus; (b) P. elongatus; (c) P. typus; (d) P. senegalensis.
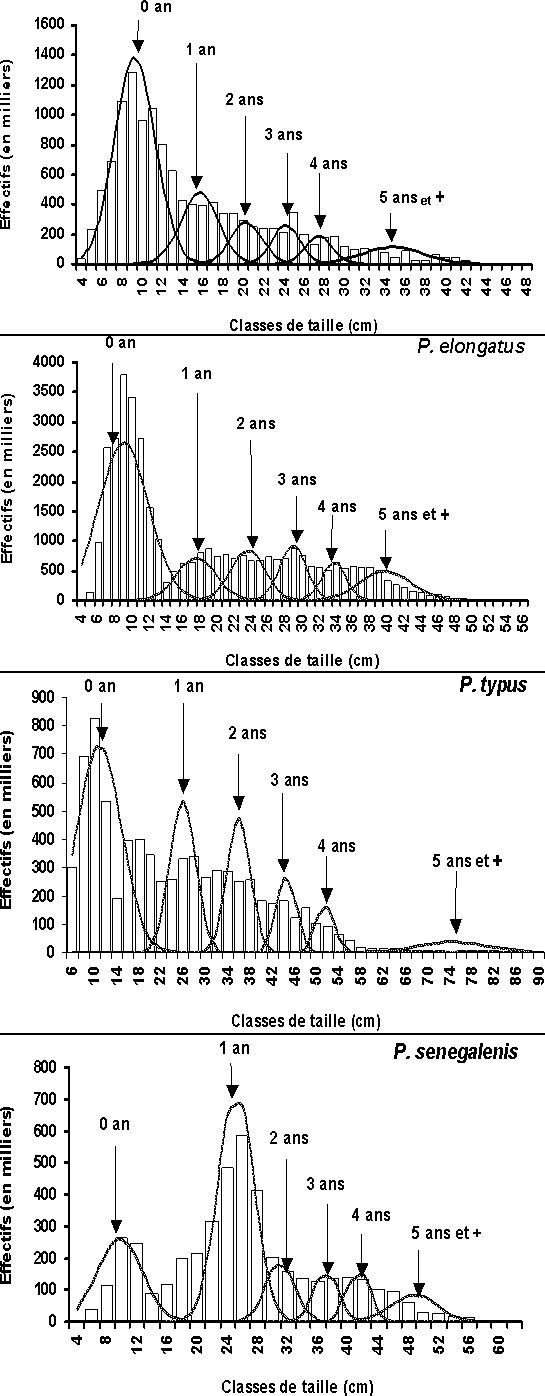
La conversion en âge des données structurées en taille est optimisée en cherchant à obtenir une fonction des écart-types aux âges linéaire et décroissante. Cette loi se justifie par des réalités plutôt biologiques et logiques que statistiques. En sortie de NORMSEP, on obtient pour chaque groupe d'âge et pour chaque espèce, une taille moyenne des individus et l'effectif total correspondant (nombre et pourcentage) de la pseudo-cohorte (cf. annexe).
Ainsi, avec les différentes lois de croissances citées ci-dessus, six groupes d'âges (de 0 à 5+) sont définis par décomposition polymodale de la moyenne (95/99) des effectifs capturés pour chacune des quatre espèces. Le groupe+ rassemble l'ensemble des individus d'âge égal et supérieur à celui de ce groupe+ (figure 3).
Figure 3: Décomposition polymodale des captures des quatre stocks en Guinée.
2.2 Effectifs et biomasse des stocks
L'analyse de pseudo-cohorte sur fréquence de taille conduit à une première estimation de l'effectif total moyen, de la biomasse totale moyenne et du recrutement moyen annuel pour les quatre stocks en Guinée. La taille de recrutement (Tr) pour chaque espèce est fixée ici par rapport à la taille moyenne du premier groupe d'âge issu de la décomposition polymodale (voir annexe); ce groupe étant considéré comme pleinement recruté.
Les résultats sont comparés à ceux issus de l'analyse rectifiée de pseudo-cohorte en âge (tableau 3). On observe que les nombres totaux de recrues sont peu différents dans les deux situations (analyse rectifiée et analyse non rectifiée). Par contre, l'effectif moyen et la biomasse totale moyenne sont beaucoup plus importants dans le cadre de l'analyse en taille (paramètres constants) que dans celui de l'analyse en âge. Ce qui pourrait certainement être lié à l'augmentation de l'effort de pêche considérée dans l'analyse rectifiée avec l'âge (effort de pêche variable) par rapport à la situation d'équilibre (effort et recrutement constants).
Il faut préciser que c'est la première fois qu'on procède à l'estimation de ces paramètres en Guinée par cette approche, les valeurs obtenues doivent donc être considérées comme fixant des ordres de grandeurs qu'il faut comparer et valider par des méthodes d'estimation différentes de celles utilisées dans le cadre du présent travail.
Tableau 3: Estimation de l'effectif total moyen annuel, de la biomasse totale moyenne et du recrutement moyen annuel des quatre stocks exploités en Guinée.
| |
V.P.A. (non rectifiée), en taille (analyse à paramètres constants) |
V.P.A. (rectifiée), en âge (analyse à paramètres variables) |
Taille critique moyenne du stock |
||||
|
Recrut. moyen (nbre en millions) |
Effectif total moyen (millions d'individus) |
Biomasse totale moyenne (en milliers de tonnes) |
Recrut. moyen (nbre en millions) |
Effectif total moyen (millions d'individus) |
Biomasse totale Moyenne (en milliers de tonnes) |
||
|
G. decadactylus |
18,8 |
179 |
15 |
19,7 |
45 |
7,8 |
24 cm |
|
P. elongatus |
73,7 |
549 |
34 |
75,2 |
180 |
21,2 |
30 cm |
|
P. senegalensis |
6,4 |
76 |
6,1 |
8,0 |
19,6 |
4,6 |
36 cm |
|
P. typus |
8,5 |
114 |
10,7 |
12,4 |
26 |
6,9 |
36 cm |
* La taille critique est la taille des poissons pour laquelle la biomasse de la cohorte est maximale avec le régime d'exploitation considéré.
2.3 Diagramme d'exploitation des différentes pêcheries
Les diagrammes d'exploitation par pêcheries sont estimés à partir des données de la pseudo-cohorte moyenne 1995-1999 pour chacune des espèces. Les quatre diagrammes mettent globalement en évidence une focalisation des pêcheurs industriels sur les individus d'âge jeune et moyen dans l'exploitation halieutique de ces quatre espèces en Guinée (figure 4).
La pêche industrielle se traduirait actuellement par des coefficients de mortalité par pêche (F) très importants (supérieur à 0,3) par rapport à ceux de la pêche artisanale (inférieur à 0,05) au niveau de G. decadactylus (figure 4a). La même situation est observée chez le P. senegalensis avec des mortalités par pêche industrielle supérieur à 0,8 sur les tailles jeunes (28 cm) et celles artisanales toujours inférieures à 0,2 (figure 4d). Au niveau des deux autres espèces (P. elongatus et P. typus), la pêche artisanale se traduit par des coefficients de mortalité par pêche (F) un peu plus élevés que ceux de la pêche industrielle pour les individus plus âgés; les plus jeunes individus sont, quant à eux, toujours soumis à des mortalités par pêche industrielle plus élevées surtout au niveau du P. elongatus. En revanche, les individus d'âge moyen des deux espèces subissent plus ou moins les mortalités par pêches des deux pêcheries (figures 4b et 4c).
Ainsi, en Guinée la pêche artisanale pourrait être qualifiée de pêcherie concomitante dans l'exploitation de G. decadactylus et de P. senegalensis. Les individus capturés au niveau de ces deux espèces par la pêche artisanale ont des tailles voisines, 24 à 28 cm pour le petit capitaine (figure 4a) et 38 à 44 cm pour bar nanka (figure 4d). Dans les autres cas de figures, on peut classer les deux pêcheries (industrielle et artisanale) comme étant partiellement séquentielles et partiellement concomitantes. La pêche industrielle capture toutes les tailles au niveau des quatre espèces et la pêche artisanale à son tour, capture toutes les tailles de P. elongatus et de P. typus (figures 4b et 4c).
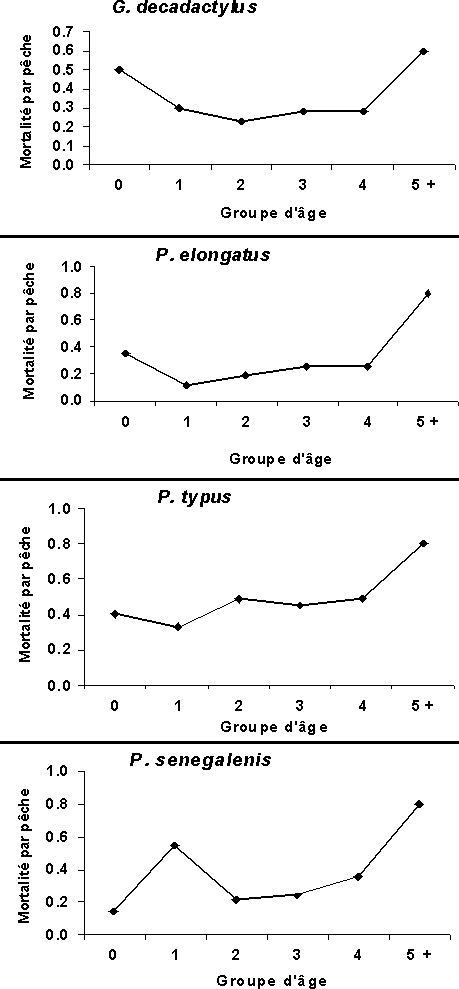
La mortalité par pêche par groupe d'âge montre des décès importants des jeunes individus (groupe 0) chez le G. decadactylus qui sont imputables à la seule pêche industrielle (figure 5).
Figure 4: Diagramme d'exploitation des deux pêcheries et du total des quatre stocks en Guinée: (a) G. decadactylus
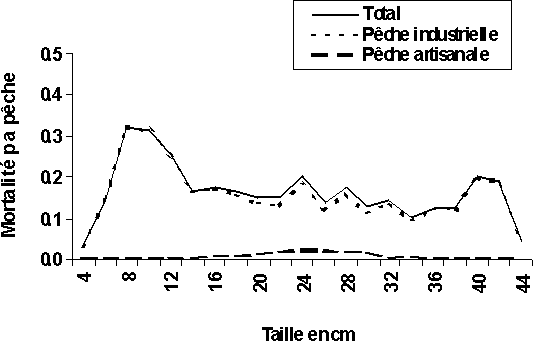
Figure 4: Diagramme d'exploitation des deux pêcheries et du total des quatre stocks en Guinée: (b) P. elongatus
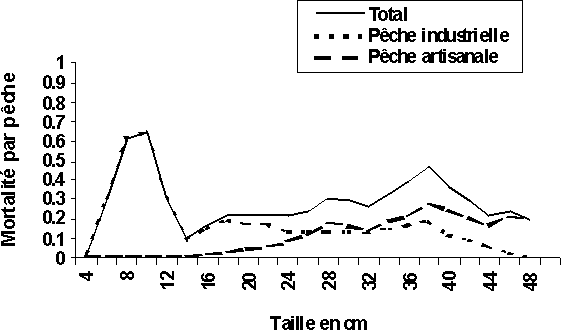
Figure 4: Diagramme d'exploitation des deux pêcheries et du total des quatre stocks en Guinée: (c) P. typus
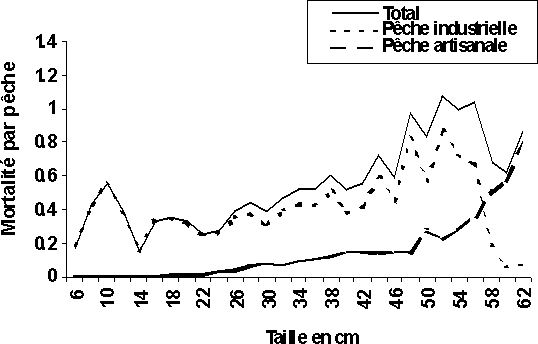
Figure 4: Diagramme d'exploitation des deux pêcheries et du total des quatre stocks en Guinée: (d) P. senegalensis.
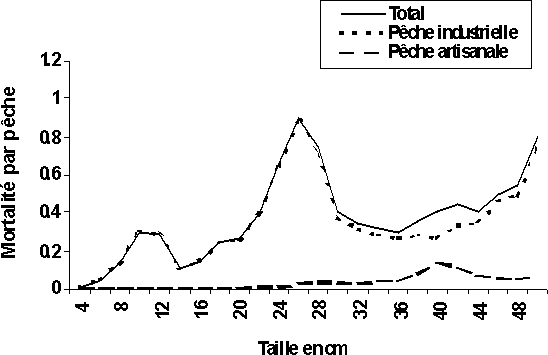
Figure 5: Evolution de mortalité par pêche en fonction des différents groupe d'âge dans l'analyse de pseudo-cohorte rectifiée avec variation de l'effort de pêche sur les quatre stocks exploités en Guinée.
2.4 Rendement par recrue des quatre stocks sur l'ensemble de la pêcherie à paramètres constants (conditions d'équilibre avec le milieu)
Les courbes de rendements par recrue sont établies pour des multiplicateurs du vecteur de mortalité par pêche variant de 0 à 2 et les captures totales par recrue sont comparées aux débarquements totaux par recrue. Rappelons que l'analyse en taille conduit à des diagnostics qui peuvent être trop optimistes, car on ne tiens pas compte de l'accroissement de l'effort.
L'analyse des différentes courbes de rendement par recrue indique que le régime d'exploitation de l'ensemble de la pêcherie guinéenne entraînerait actuellement des situations allant d'une pleine exploitation à une surexploitation plus ou moins marquée (figure 6).
Figure 6: Courbes des rendements par recrue de l'ensemble de la pêcherie en fonction d'un multiplicateur de mortalité par pêche totale sous hypothèse d'équilibre: (a) G. decadactylus, (b) P. elongatus, (c) P. typus et de (d) P. senegalensis.
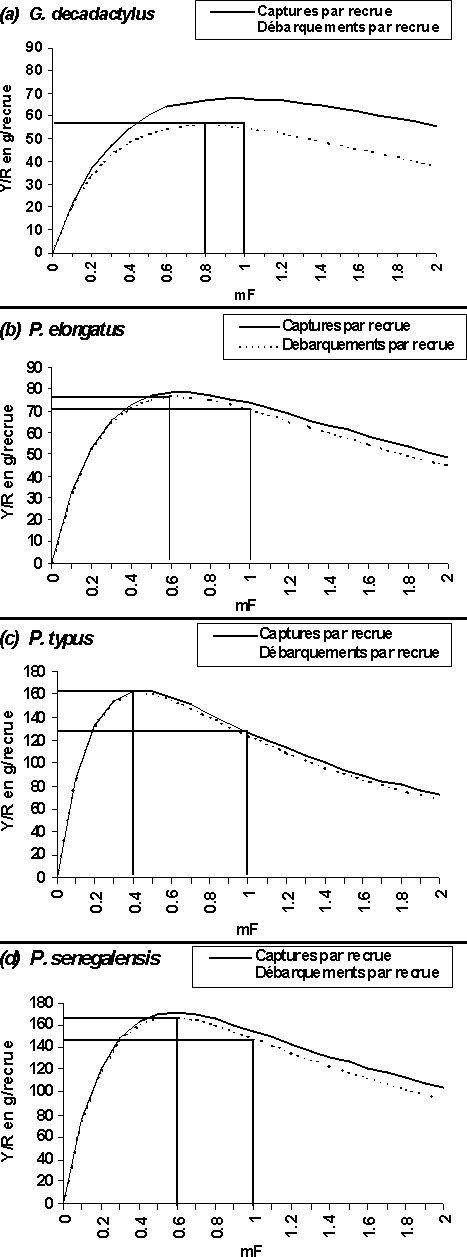
Le G. decadactylus est dans un état de pleine exploitation. La situation de maximisation des captures par recrue est en effet actuellement atteint. L'augmentation de l'effort de pêche ne permettra aucun gain supplémentaire de production par recrue, au contraire mettra le stock dans une situation de surexploitation. De plus, l'importance des rejets (la différence des débarquements et des captures) aux alentours de mFmax=1 est relativement grande et plus cet effort s'accroît plus les rejets sont importants dans les captures (figure 6a). La mortalité par pêche liée aux rejets au niveau du stock est le fait de la seule pêche industrielle et s'applique uniquement aux très jeunes individus. Elle ampute donc le potentiel de débarquement total quand elle augmente.
Pour les trois autres espèces (P. elongatus, P. typus et P. senegalensis), le régime d'exploitation de l'ensemble de la pêcherie entraîne une situation de surexploitation. Les efforts de pêche qui pourraient maximiser les captures par recrue au niveau des différents stocks sont aujourd'hui dépassés, puisque pour le bobo mFmax = 0,6 (figure 6b), pour le bar sénégalais mFmax = 0,5 (figure 6d) et pour le bar nanka mFmax = 0,4 (figure 6c). En d'autres termes, pour espérer retrouver les meilleurs rendements au niveau de ces trois espèces, il faudrait réduire de 40 pour cent l'effort de pêche global actuel subi par le P. elongatus, de 50 pour cent celui subi par le P. senegalensis et enfin de 60 pour cent celui exercé sur le P. typus.
Le rejet aux alentours de mF = 1 est actuellement très important chez le bobo (figure 6b) et pourrait être lié, uniquement à la mortalité par pêche industrielle des jeunes poissons. Plus cet effort est réduit plus les rejets sont faibles et plus le potentiel de débarquement est élevé. En revanche chez le P. typus et le P. senegalensis, il est remarquable qu'aux alentours de mFMSY, l'importance des rejets est faible (figures 6c et 6d) et quelle augmente quand l'effort s'accroît.
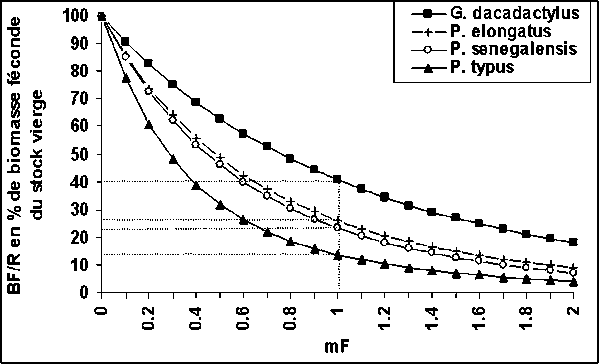
Par ailleurs, la biomasse féconde actuelle estimée au niveau du petit capitaine, du bobo et du bar sénégalais est supérieure à 20 pour cent de la biomasse féconde du stock vierge (figure 7), ce qui la situe très au-dessus du seuil empirique de surexploitation du recrutement généralement admis de 10 pour cent. En revanche, la biomasse féconde actuelle estimée au niveau du bar nanka est très proche des 10 pour cent de la biomasse féconde du stock vierge (figure 7). Ce qui pourrait traduire aujourd'hui un risque de surexploitation du recrutement de cette espèce en Guinée.
Figure 7: Biomasse féconde par recrue actuelle en pourcentage de la biomasse féconde vierge des quatre stocks en Guinée.
2.5 Diagnostic par pêcherie (pêche artisanale et pêche industrielle)
L'analyse des isoplètes de production par recrue totale (figure 8) permet de comprendre le comportement de chaque segment de la pêche mais aussi de confronter différents scenarii entre la pêche artisanale et la pêche industrielle dans la recherche de la production maximale pour chacune des quatre espèces.
Figure 8: Isoplètes de production par recrue totale en fonction de multiplicateurs d'efforts sur les deux pêcheries.

En terme de production totale par stock, toute modification de l'effort de pêche artisanal actuel aurait un impact quasiment nul sur la production du G. dacadactylus; par contre toute augmentation de l'effort de pêche industriel pourrait se traduire à court terme par une diminution des captures totales. D'une façon réaliste, le pêche industrielle en évitant de pêcher des jeunes poissons, aurait intérêt à maintenir son diagramme d'exploitation avec un mF compris entre 0,8 et 1,2 (figure 8a). Chez le P. elongatus et le P. typus, une augmentation de l'effort artisanal pourrait encore se traduire par une augmentation de la production totale, à condition bien sur que la pression de pêche industrielle soit baissée considérablement par rapport à son niveau actuel (figures 8b et 8c). En d'autre terme, la pêche industrielle doit rapprocher son diagramme d'exploitation de celui de la pêche artisanale, c'est à dire pêcher moins de poissons jeunes et plus de poissons de grandes tailles et plus âgés, dont le potentiel de production par recrue est nettement plus élevé.
Pour ces deux espèces l'interaction entre pêcheries est forte. Toute baisse de l'effort de la pêche industrielle se traduirait par un accroissement des captures de la pêche artisanale.
C'est à peu près la même situation qui est observée au niveau du P. senegalensis (figure 8d). Une légère diminution de l'effort de pêche industrielle actuel, se traduirait à court et/ou à moyen terme par une augmentation de la production totale par recrue.
2.6 Sensibilité des résultats aux incertitudes liées à la mortalité naturelle
L'étude de la sensibilité du rendement par recrue est réalisée en estimant le vecteur de mortalité par pêche avec différentes hypothèses sur les paramètres d'entrée de l'analyse de pseudo-cohorte. On teste donc ici la sensibilité du modèle de rendement par recrue à différentes valeur de mortalité naturelle (M).
Globalement les différentes hypothèses sur M ne modifient pas l'allure générale de la courbe du rendement par recrue mais plutôt son amplitude (figure 9). Il est en effet logique de constater que plus la mortalité naturelle est faible, plus le rendement par recrue maximal est élevé et que l'effort permettant d'atteindre ce maximum reste peu modifié.
Figure 9: Rendement de production totale par recrue pour les différentes hypothèses de mortalités naturelles.
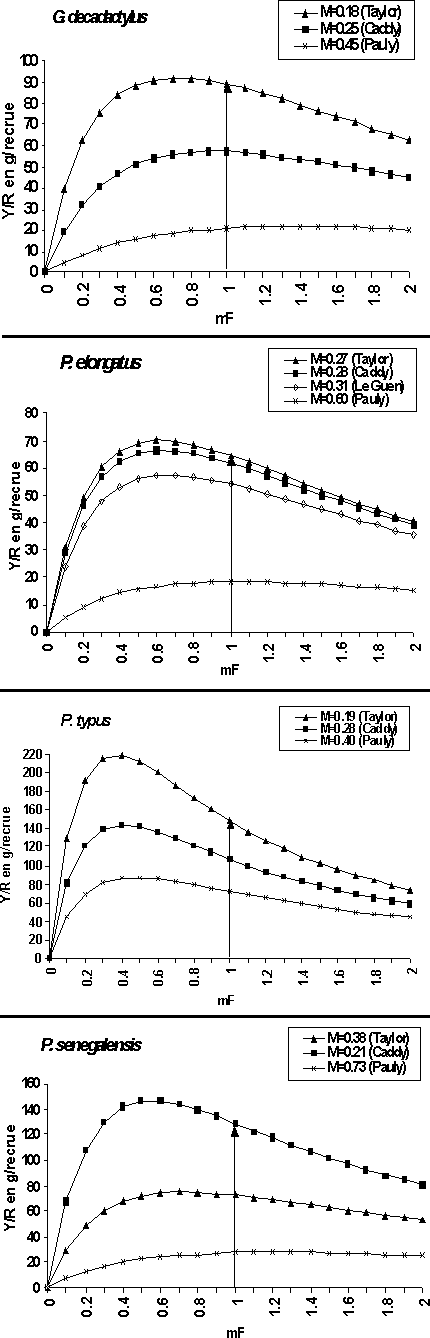
Les résultats de l'étude des rendements par recrue, notamment les valeurs maximales des Y/R sont donc sensibles aux différentes hypothèses sur M et sont certainement discutables d'un point de vue quantitatif. Cependant cette incertitude ne doit pas masquer la composante la plus fiable des diagnostics: dans la plupart des hypothèses réalistes testées, il semble aujourd'hui que le stock de G. decadactylus serait dans une situation de pleine exploitation; tandis que le P elongatus et le P. senegalensis se trouveraient dans une situation de surexploitation plus ou moins marquée et qu'enfin le stock du P. typus serait dans une situation de nette surexploitation (figure 9).
2.7 Rendement par recrue de pseudo-cohorte en âge: analyse rectifiée avec l'effort
L'analyse rectifiée des pseudo-cohortes est entreprise ici pour mesurer l'impact de l'hypothèse d'équilibre considérée dans l'analyse en taille. Tous les effectifs (Ni) et les mortalités par pêche (Fi) aux différents âges (âge arbitraire) sont obtenu sous le Programme Simuco (Mesnil, 1988).
Figure 10: Courbes de rendement par recrue de l'ensemble de la pêcherie relatives aux captures par âge (analyse rectifiée de pseudo-cohorte) et aux captures par taille (analyse de pseudo-cohorte sous hypothèse d'équilibre) en fonction d'un multiplicateur du vecteur de mortalité par pêche totale.
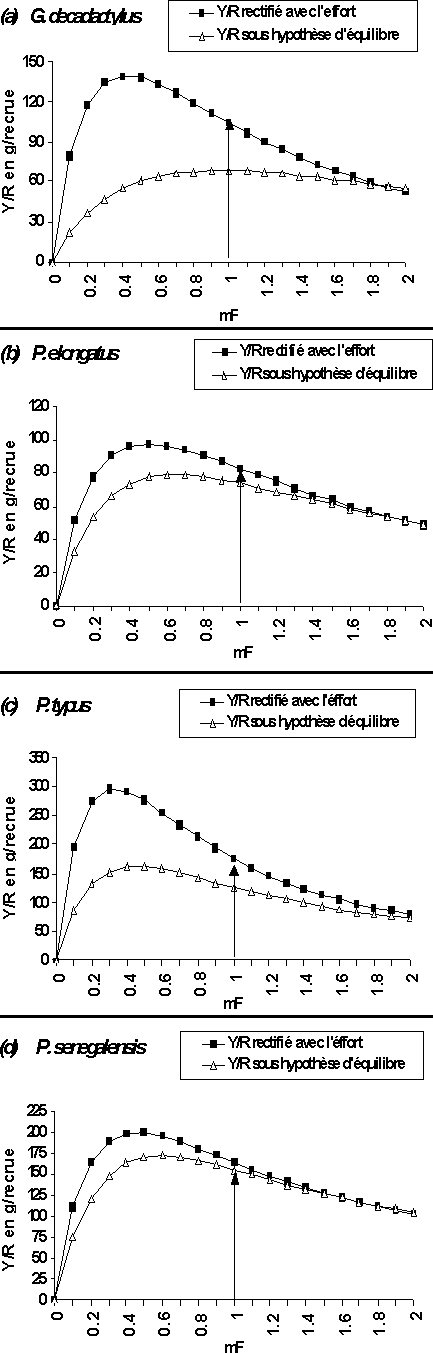
Les différentes courbes de rendement total par recrue issues de l'analyse rectifiée de pseudo-cohorte en âge sont comparées à celles issues de l'analyse sous hypothèse d'équilibre (figure 10). L'analyse rectifiée avec l'effort indique pour les quatre stocks une situation de surexploitation avec des niveaux maximums de production par recrue (point de vue quantitatif) beaucoup plus élevés et donc, une situation de surexploitation plus marquée.
Par ailleurs, à l'exception du cas de G. decadactylus où on observe une différence de diagnostics entre les deux analyses (figure 10a), les courbes de rendements par recrue des trois autres espèces présentent la même allure générale (figure 10b, 10c et 10d). Malgré la différence observée entre les niveaux maximums de production par recrue, les résultats issus des deux méthodes restent globalement le même. L'utilisation de l'une des deux structures, captures aux âges (analyse rectifiée avec l'effort) ou captures aux tailles (analyse sous hypothèse d'équilibre), conduit sensiblement au même diagnostic: celle d'une surexploitation de stock avec les mF de maximisation de production par recrue largement aujourd'hui dépassés.
Une synthèse des indicateurs halieutiques issus des deux méthodes d'analyse est présentée dans le tableau 4. Les niveaux de maximisation de production (mFMSY) dans les deux analyses sont aujourd'hui nettement dépassés par rapport à la situation actuelle (mF=1). Ce phénomène est particulièrement marqué chez le P. typus qui se trouve dans une situation de très nette surexploitation. D'autre indicateur comme le ratio: rendement par recrue actuel sur le rendement par recrue maximal (YmF=1/YMSY), permet de suivre le niveau de perte des captures dans les pêcheries. Le niveau 1 correspond à une exploitation maximal des stocks; plus cet indicateur est loin de 1 plus la pêcherie se porte mal. Enfin, la biomasse féconde et la biomasse totale sont aussi utilisées comme indicateurs de l'état des stocks exploités.
Tableau 4: Indicateurs halieutiques issus de l'analyse rectifiée avec l'effort et de l'analyse sous hypothèse d'équilibre (effort et recrutement constants).
| |
mFMSY |
YmF = 1/YMSY |
BF/R |
B/R |
||||
|
équilibre |
rectifiée |
équilibre |
rectifiée |
équilibre |
rectifiée |
équilibre |
rectifiée |
|
|
G. decadactylus |
0,9 |
0,4 |
0,999 |
0,748 |
39,2% |
30,6% |
40,6% |
31,4% |
|
P. elongatus |
0,7 |
0,5 |
0,940 |
0,850 |
27,6% |
38,7% |
34,1% |
43,6% |
|
P. typus |
0,4 |
0,3 |
0,784 |
0,595 |
15,1% |
18,7% |
21,5% |
22,8% |
|
P. senegalensis |
0,6 |
0,5 |
0,905 |
0,822 |
25,0% |
32,4% |
33,3% |
36,9% |
mFMSY = multiplicateur de l'effort de maximisation de production par recrue.
YmF=1/YMSY = indicateur de la perte de capture.
BF/R = pourcentage de la biomasse féconde vierge actuelle par rapport à la biomasse féconde du stock vierge.
B/R = pourcentage de la biomasse totale actuelle par rapport à la biomasse totale du stock vierge.
3. Discussion des résultats et conclusion
L'analyse de cohorte fondée sur les distribution de fréquences de taille (Jones, 1983) a été privilégiée dans ce travail. Parallèlement, une analyse rectifiée (variation de l'effort de pêche théorique) de peudo-cohorte en âge a été entreprise. Ce qui permettait de s'affranchir de l'hypothèse d'équilibre. Les résultats issus des deux démarches utilisées dans ce travail sont ici comparés et discutés.
L'analyse séquentielle de la population (ou VPA) nous a permis d'obtenir une première estimation de l'effectif moyen et de la biomasse totale moyenne de G. decadactylus, P. typus et de P. senegalensis en Guinée; et d'actualiser pour le P. elongatus des estimations issues des travaux antérieurs (Sidibé et al., 2000). Ces valeurs sont relativement différentes selon la méthodologie employée (tableau 3) et sont beaucoup liées à la qualité des données utilisées ici. Par conséquent, elle doivent être prises avec prudence et considérées à ce stade comme des ordres de grandeurs qui pourront être fiabilisées par d'autres analyses beaucoup plus fines avec des données plus complètes.
Les quatre stocks analysés dans ce travail sont tous exploités à la fois par la pêche artisanale et la pêche industrielle en Guinée. L'analyse des effectifs capturés a montré que c'est essentiellement l'effort industriel qui entraîne la mortalité des individus de petites tailles au niveau des quatre espèces; et que plus cet effort est important plus les captures dépendraient du recrutement annuel. Si on avait de mauvais recrutements au cours d'une année donnée, tout laisse à croire que les captures pourraient baisser l'année suivante. Plus généralement, la pêcherie industrielle a intérêt à rapprocher son diagramme d'exploitation de celui de la pêcherie artisanale qui épargne les jeunes individus avec une meilleure sélectivité (figure 4).
L'analyse des courbes de captures par recrue est très claire. A part le petit capitaine dont le diagnostic reste relativement optimiste (figure 6a), le régime d'exploitation de l'ensemble de la pêcherie, entraînerait aujourd'hui une situation de surexploitation des trois autres stocks en Guinée (figures 6b, 6c et 6d).
Le niveau des rejets dans les captures est aussi clairement mis en évidence autours de mF=1. Ils sont importants chez le G. decadactylus (figure 6a) et le P. elongatus (figure 6b). Et ce résultat confirme les observations faites sur les structures démographiques des captures au niveau de ces deux espèces (figure 2).
Par ailleurs, ces analyses n'ont mis en évidence aucune surexploitation du recrutement. Le niveau actuel de la biomasse féconde des quatre espèces est supérieur à 10 pour cent par rapport à la biomasse féconde du stock vierge (figure 7). On admet généralement que le risque d'une surexploitation de recrutement n'est réel que si la biomasse féconde, par l'action de la pêche est réduite jusqu'à 10 pour cent de son état vierge (Gascuel, 1993). Toutefois, il faut noter que la biomasse féconde du P. typus est aujourd'hui très proche de ce seuil critique en Guinée.
En terme de rendement par recrue par pêcherie, les analyses ont montré que les diagrammes d'exploitation artisanaux et industriels n'ont pas les mêmes potentialités chez les quatre espèces (figure 8). Globalement le potentiel de la production artisanale est fortement dépendant de l'exploitation industrielle chez le P elongatus (figure 8b) et le P. typus (figure 8c); et une diminution de l'intensité de pêche industrielle est susceptible d'entraîner une augmentation de la production par recrue artisanale.
Chez le G. decadactylus (figure 8a) et le P. senegalensis (figure 8d), le potentiel artisanal est peu dépendant de la pêche industriel. Toutefois, la pêche industrielle amputerait une part non négligeable au potentiel artisanal par la mortalité importante qu'elle provoque au niveau de très jeunes individus qui sont rejetés à la mer au niveau de ces deux espèces.
L'analyse rectifiée de pseudo-cohorte en âge avec des séries d'efforts de pêche variables, a permis d'étudier l'impact de l'hypothèse d'équilibre sur les résultats. Les niveaux de rendement maximum par recrue sont beaucoup plus élevés dans l'analyse rectifiée de pseudo-cohorte en âge que ceux de l'analyse de pseudo-cohorte en taille à paramètres constants. Cette différence pourrait être mise au compte de l'augmentation de l'effort de pêche théorique considérée dans l'analyse rectifiée par rapport à celle non rectifiée. Toutefois, même si les niveaux maximums de production par recrue (point de vue quantitatif) diffèrent d'une analyse à l'autre, les courbes de rendements par recrue total obtenues sont très proches en terme de diagnostic global sur l'état des stocks (figure 10). Ainsi, nous pouvons dire que quelque soit la démarche adoptée (l'hypothèse d'équilibre ou de pseudo-cohorte rectifiée), le diagnostic final reste le même: celui d'une situation de surexploitation des stocks. L'étude de sensibilité des rendements par recrues aux différentes hypothèses de mortalité naturelle abouti également aux mêmes résultats (figure 9).
En conclusion, les résultats obtenus par les différentes méthodes d'analyse utilisées dans ce travail permettent d'affirmer globalement que les quatre stocks sont aujourd'hui dans une situation de surexploitation de croissance en Guinée et que l'effort de maximisation de capture pour chaque recrue est actuellement dépassé. Il existe peu d'éléments permettant d'avancer que l'intensité de l'exploitation sur ces espèces comme la plupart des autres espèces de la communauté à sciaenidés de Guinée a diminué aujourd'hui. Il est donc logique de penser que la situation ne s'est pas améliorée.
Bibliographie
Beverton R.J.H. et Holt S.J., 1957. On the dynamique of exploited fish populations. Fish. Invest. Lond., Ser. 2, 19: 1-533.
Caddy, J.F. 1991. Death rates and time intervals: Is there an alternative to the constant natural mortality axiom? Rev. Fish Biol. Fisheries, 1: 109-138.
Caverivière A. et Thiam M., 1994. Tailles moyennes et taux d'exploitation approchés des principales espèces de poissons débarquées par les chalutiers glaciers entre 1973 et 1989. In L'évaluation des ressources exploitables par la pêche artisanale sénégalaise, Tome2. Edité par M. Barry-Gerard, T. Diouf, A. Fonteneau. Colloques et Séminaire ORSTOM éds. Paris, pp 365-384.
CNSHB, 1994. Note de synthèse sur les travaux et conclusions du Groupe de travail d'analyse des campagnes de prospection du N.O. Nizery en Guinée, de 1986 à 1992. CNSHB, Conakry, Avril 1994, 8 p.
CNSHB, 1996. Bulletin statistique des pêches, année 1995, Sér. Bul. stat., 1, 34 p.+an.
CNSHB, 1997. Bulletin statistique des pêches, année 1996, Sér. Bul. stat., 2, 35 p.+an.
CNSHB, 1998. Bulletin statistique des pêches, année 1997, Sér. Bul. stat., 3, 44 p.+an.
CNSHB, 1999. Bulletin statistique des pêches, année 1998, Sér. Bul. stat., 3, 42 p.+an.
CNSHB, 2000. Bulletin statistique des pêches, année 1999, Sér. Bul. stat., 3, 41 p.+an.
De Verdelhan, 1987. Programmes VERDEJON: VPA et simulation sur fréquences de taille. IFREMER.
Domain F., Chavance P. et Bah A., 2000a.Notes sur la reproduction des espèces démersales du peuplement côtier guinéen. In DOMAIN F., CHAVANCE P. et DIALLO A. (éditeurs). La Pêche Côtière en Guinée - Ressources et Exploitation. Editions IRD/CNSHB, 393 p.
Gascuel D., 1993. Modélisation de la dynamique des stocks exploités par la pêche artisanale sénégalaise: intérêt, limites et contraintes de l'approche structurale. Symposium méthodologie d'évaluation des ressources exploitées par la pêche artisanale au Sénégal. Dakar, Février 1993, 17p.
Gascuel D., 1994. Une méthode simple d'ajustement des clés taille/âge: application aux captures d'albacores (Thunnus albacares) de l'Atlantique Est. Can. J. Fish. Aquat. Sci., 51 (3), p. 723-733.
Gulland J.M., 1969. Manuel des méthodes d'évaluation des stocks d'animaux aquatiques. Première partie: Analyse des population. Man. Sci. Aquat. FAO., 4: 160p.
Jones R., 1983. The use of length composition data in fish stocks assessments (with notes on VPA and cohort analysis). FOA Fish. Circ., 734: 60p.
Laurec A. et Santarelli-Chaurand L., 1986. Analyse rectifiée des pseudo-cohortes: Analyse des cohortes à partir d'une année de structure démographique des captures; correction des variations d'effort et/ou de recrutement. Doc. minéo. 19p.
Le Guen J.C, 1971. Dynamique des populations de Pseudotolithus (Fonticulus) elongatus (Bowd, 1825). Poisson, sciaenidae. Cah. ORSTOM, sér. Océanogr., 9(1): 3-84.
Lesnoff M., Traore S., Sidibe A. et Soumah M., 1995. Le système de suivi des statistiques de pêche industrielle. Archive du Centre National des Sciences Halieutiques de Boussoura, 23, 103p.
Mesnil B., 1988. Logiciels pour l'évaluation des stocks de poissons. ANACO: Logiciel d'analyse des données de captures par classe d'âge sur IBM PC et compatibles. FAO Doc. Tech. Pêche, 101, suppl. 3: 78p.
Pauly, D. 1980b. On the interrelationships between natural mortality, growth parameters, and mean environemental temperature in 175 fish stocks. J. Cons. CIEM, 39 (2): 175-192.
Poinsard, F. 1973. Croissance des Pseudotolithus typus dans la région de Pointe Noire. Doc. Scient. Centre Rech. Océanogr. Pointe Noire, N.S., 20, 11p.
Samba G., 1974. Contribution à l'étude de la biologie et de la dynamique d'un polynemidae ouest africain Galeoides decadactylus (BLOCH). Thèse Doct. 3e cycle, Univ. Bordeaux I, 114p.
Sidibe A., 1998. Le bobo (bossu), Pseudotolithus elongatus (Bowdich, 1825). Ressource et exploitation en Guinée. Evaluation et simulation d'aménagement. Mémoire de D.E.A. d'Océanologie Biologique et Environnement Marin. Ecole Nationale Supérieure Agronomique de Rennes (ENSAR), minéo, 32p. +annexe.
Sidibe A., Gascuel D., Domain F. et Chanvance P., 2000. Estimation d'abondance et changement de répartition spatiale: le cas du bobo (Pseudotolithus elogantus) en Guinée. In: Didier G., Chavance P., Bez N., et Biseau A. (éditeurs). - Les espaces de l'halieutique -. Colloques et Séminaire ORSTOM éds. Paris, pp 75-98
Taylor, C.C. 1960. Temperature, growth and mortality - the Pacific cockle. J. Cons. CIEM, 26: 117 -124
Tomlinson, P.K. 1970. Program NORMSEP. Normal distribution separator using steepest descent methode. 11 (1).2.4 to 11 (1).2.10.
Troadec, J.P. 1971. Biologie et dynamique d'un Sciaenidae ouest africain: Pseudotolithus senegalensis (V.). Doc. Scient. Centre Rech. Océanogr. Abidjan, 2(3): 225p.
ANNEXE
Tableau 1: Captures annuelles (en tonnes) des quatre espèces en Guinée - Source: Bulletin statistique CNSHB (1995-1999).
|
Année |
Espèces |
Pêche artisanale* |
Pêche industrielle |
Total |
| |
G. dacadactylus |
195 |
797 |
992 |
|
P. elongatus |
3 656 |
694 |
4 350 |
|
|
P. senegalensis |
233 |
1 408 |
1 641 |
|
|
P. typus |
2 458 |
112 |
2 570 |
|
|
1996 |
G. dacadactylus |
215 |
969 |
1 184 |
|
P. elongatus |
3 379 |
462 |
3 841 |
|
|
P. senegalensis |
84 |
637 |
721 |
|
|
P. typus |
1 683 |
462 |
2 145 |
|
|
1997 |
G. dacadactylus |
50 |
1 907 |
1 957 |
|
P. elongatus |
2 745 |
3 894 |
6 639 |
|
|
P. senegalensis |
100 |
469 |
569 |
|
|
P. typus |
1 277 |
3 376 |
4 653 |
|
|
1998 |
G. dacadactylus |
26 |
1 157 |
1 183 |
|
P. elongatus |
2 781 |
2 117 |
4 898 |
|
|
P. senegalensis |
148 |
1 005 |
1 153 |
|
|
P. typus |
1 383 |
786 |
2 169 |
|
|
1999 |
G. dacadactylus |
88 |
1 736 |
1 824 |
|
P. elongatus |
2 142 |
2 608 |
4 679 |
|
|
P. senegalensis |
104 |
1 378 |
1 430 |
|
|
P. typus |
745 |
1 078 |
1 451 |
* Les captures totales de la pêche artisanale pour l'année 1999 sont extrapolées par rapport au second semestre au cours duquel aucune enquête statistique n'a pu être effectuée.
Tableau 2: Effectif total moyen (milliers) des captures commerciales des quatre espèces en Guinée (1995-99).
|
Galeoides decadactylus |
||||
|
Taille (cm) |
Prises conservées P. industr. |
Prises rejetées P. industr. |
Prises débarquées P. artisanale |
Total en milliers |
|
4 |
0 |
269 |
0 |
269 |
|
6 |
0 |
1 179 |
0 |
1 179 |
|
8 |
0 |
2 376 |
1 |
2 377 |
|
10 |
5 |
2 008 |
0 |
2 012 |
|
12 |
21 |
1 408 |
4 |
1 432 |
|
14 |
121 |
699 |
11 |
831 |
|
16 |
372 |
409 |
26 |
806 |
|
18 |
373 |
273 |
35 |
680 |
|
20 |
308 |
183 |
50 |
541 |
|
22 |
292 |
140 |
50 |
482 |
|
24 |
450 |
65 |
52 |
567 |
|
26 |
262 |
27 |
45 |
334 |
|
28 |
309 |
21 |
35 |
365 |
|
30 |
187 |
5 |
29 |
221 |
|
32 |
189 |
6 |
7 |
202 |
|
34 |
105 |
5 |
10 |
120 |
|
36 |
111 |
2 |
4 |
117 |
|
38 |
84 |
0 |
3 |
86 |
|
40 |
86 |
0 |
2 |
88 |
|
42 |
40 |
0 |
1 |
41 |
|
44 |
4 |
0 |
0 |
4 |
|
46 |
2 |
0 |
1 |
3 |
|
Total = |
3 322 |
9 073 |
363 |
12 759 |
|
Pseudotolithus elongatus |
||||
|
Taille (cm) |
Prises conservées P. industr. |
Prises rejetées P. industr. |
Prises débarquées P. artisanale |
Total en milliers |
|
4 |
0 |
148 |
0 |
148 |
|
6 |
0 |
3 537 |
1 |
3 538 |
|
8 |
0 |
6 526 |
7 |
6 532 |
|
10 |
0 |
6 125 |
7 |
6 132 |
|
12 |
2 |
2 589 |
9 |
2 600 |
|
14 |
350 |
414 |
27 |
791 |
|
16 |
1 004 |
178 |
93 |
1 275 |
|
18 |
1 311 |
148 |
227 |
1 686 |
|
20 |
1 072 |
131 |
315 |
1 518 |
|
22 |
1 031 |
100 |
363 |
1 493 |
|
24 |
809 |
16 |
496 |
1 322 |
|
26 |
749 |
3 |
681 |
1 433 |
|
28 |
656 |
3 |
934 |
1 592 |
|
30 |
587 |
1 |
773 |
1 360 |
|
32 |
512 |
0 |
565 |
1 078 |
|
34 |
533 |
0 |
658 |
1 191 |
|
36 |
502 |
0 |
644 |
1 146 |
|
38 |
427 |
0 |
653 |
1 080 |
|
40 |
195 |
0 |
424 |
619 |
|
42 |
114 |
0 |
268 |
382 |
|
44 |
57 |
0 |
165 |
222 |
|
46 |
16 |
0 |
149 |
166 |
|
48 |
0 |
0 |
83 |
83 |
|
50 |
0 |
0 |
47 |
47 |
|
52 |
0 |
0 |
33 |
33 |
|
54 |
0 |
0 |
15 |
15 |
|
Total = |
9 927 |
19 917 |
7 637 |
37 482 |
|
Pseudotolithus typus |
||||
|
Taille (cm) |
Prises conservées P. industr. |
Prises rejetées P. industr. |
Prises débarquées P. artisanale |
Total en milliers |
|
6 |
0 |
304 |
0 |
304 |
|
8 |
0 |
692 |
0 |
692 |
|
10 |
0 |
825 |
1 |
826 |
|
12 |
5 |
529 |
1 |
534 |
|
14 |
83 |
108 |
1 |
192 |
|
16 |
325 |
70 |
1 |
396 |
|
18 |
341 |
55 |
5 |
400 |
|
20 |
296 |
40 |
10 |
346 |
|
22 |
203 |
36 |
11 |
251 |
|
24 |
223 |
15 |
21 |
258 |
|
26 |
289 |
11 |
29 |
329 |
|
28 |
294 |
0 |
44 |
339 |
|
30 |
216 |
0 |
52 |
268 |
|
32 |
248 |
0 |
45 |
293 |
|
34 |
239 |
0 |
50 |
289 |
|
36 |
202 |
1 |
49 |
253 |
|
38 |
210 |
0 |
48 |
258 |
|
40 |
134 |
0 |
51 |
185 |
|
42 |
128 |
0 |
43 |
171 |
|
44 |
150 |
0 |
34 |
184 |
|
46 |
93 |
0 |
30 |
124 |
|
48 |
135 |
0 |
24 |
160 |
|
50 |
68 |
0 |
33 |
102 |
|
52 |
77 |
0 |
19 |
96 |
|
54 |
45 |
0 |
17 |
62 |
|
56 |
28 |
0 |
15 |
43 |
|
58 |
5 |
0 |
15 |
20 |
|
60 |
1 |
0 |
12 |
14 |
|
62 |
1 |
0 |
12 |
13 |
|
64 |
4 |
0 |
10 |
14 |
|
66 |
1 |
0 |
8 |
9 |
|
68 |
0 |
0 |
11 |
11 |
|
70 |
0 |
0 |
12 |
12 |
|
72 |
0 |
0 |
10 |
11 |
|
74 |
1 |
0 |
7 |
7 |
|
76 |
0 |
0 |
8 |
8 |
|
78 |
2 |
0 |
9 |
11 |
|
80 |
1 |
0 |
9 |
9 |
|
82 |
1 |
0 |
7 |
8 |
|
84 |
1 |
0 |
5 |
6 |
|
86 |
0 |
0 |
7 |
7 |
|
88 |
1 |
0 |
6 |
6 |
|
90 |
0 |
0 |
7 |
7 |
|
92 |
0 |
0 |
6 |
7 |
|
94 |
1 |
0 |
8 |
8 |
|
96 |
0 |
0 |
11 |
12 |
|
98 |
1 |
0 |
15 |
16 |
|
100 |
0 |
0 |
17 |
18 |
|
102 |
0 |
0 |
19 |
19 |
|
104 |
0 |
0 |
10 |
10 |
|
106 |
0 |
0 |
14 |
14 |
|
108 |
0 |
0 |
13 |
13 |
|
Total = |
4 055 |
2 689 |
903 |
7 647 |
|
Pseudotolithus senegalensis |
||||
|
Taille (cm) |
Prises conservées P. industr. |
Prises rejetées P. industr. |
Prises débarquées P. artisanale |
Total en milliers |
|
4 |
0 |
5 |
0 |
5 |
|
6 |
0 |
41 |
0 |
41 |
|
8 |
0 |
116 |
0 |
116 |
|
10 |
0 |
262 |
0 |
262 |
|
12 |
0 |
246 |
0 |
246 |
|
14 |
13 |
77 |
0 |
90 |
|
16 |
42 |
75 |
1 |
118 |
|
18 |
70 |
128 |
0 |
199 |
|
20 |
98 |
116 |
1 |
216 |
|
22 |
211 |
103 |
4 |
318 |
|
24 |
421 |
57 |
7 |
485 |
|
26 |
558 |
14 |
14 |
586 |
|
28 |
391 |
6 |
16 |
413 |
|
30 |
185 |
1 |
16 |
202 |
|
32 |
149 |
0 |
9 |
158 |
|
34 |
123 |
0 |
15 |
138 |
|
36 |
111 |
0 |
16 |
127 |
|
38 |
108 |
0 |
28 |
136 |
|
40 |
93 |
0 |
47 |
139 |
|
42 |
98 |
0 |
34 |
132 |
|
44 |
85 |
0 |
17 |
102 |
|
46 |
86 |
0 |
9 |
95 |
|
48 |
58 |
0 |
5 |
63 |
|
50 |
29 |
0 |
3 |
31 |
|
52 |
23 |
0 |
2 |
26 |
|
54 |
15 |
0 |
3 |
17 |
|
56 |
10 |
0 |
1 |
11 |
|
58 |
3 |
0 |
1 |
4 |
|
60 |
2 |
0 |
2 |
4 |
|
62 |
1 |
0 |
1 |
2 |
|
64 |
0 |
0 |
1 |
1 |
|
66 |
0 |
0 |
1 |
1 |
|
68 |
0 |
0 |
2 |
2 |
|
70 |
1 |
0 |
2 |
4 |
|
72 |
0 |
0 |
2 |
2 |
|
74 |
0 |
0 |
0 |
0 |
|
76 |
0 |
0 |
0 |
0 |
|
78 |
0 |
0 |
0 |
0 |
|
80 |
0 |
0 |
2 |
2 |
|
82 |
0 |
0 |
0 |
0 |
|
84 |
0 |
0 |
0 |
0 |
|
86 |
0 |
0 |
0 |
0 |
|
88 |
0 |
0 |
1 |
1 |
|
90 |
0 |
0 |
0 |
1 |
|
92 |
1 |
0 |
0 |
1 |
|
94 |
1 |
0 |
0 |
1 |
|
96 |
0 |
0 |
0 |
0 |
|
98 |
0 |
0 |
0 |
0 |
|
100 |
1 |
0 |
0 |
1 |
|
Total= |
2 986 |
1 247 |
266 |
4 500 |
Tableau 3: Conversion taille-âge obtenue par décomposition polymodale sous contrainte des lois de croissance.
|
|
Classe d'âge |
Taille moyenne |
Ecart-type |
Pourcentage |
Nombre |
|
G. decadactylus |
0 |
9,80 |
2,00 |
55,1 |
7 032 |
|
P. elongatus |
0 |
9,5 |
2,90 |
51,9 |
19 458 |
|
P. typus |
0 |
11,80 |
3,89 |
47,3 |
3 618 |
|
P. senegalensis |
0 |
10,70 |
2,88 |
21,4 |
961 |
| [15] Centre national des
sciences halieutiques de Boussoura (CNSHB), BP.3738/39, Conakry, Guinée
E-mail: [email protected] [16] ENSAR, Laboratoire halieutique, 65, route de St Brieuc, 35042 Rennes Cedex, France [17] Centre national des sciences halieutiques de Boussoura (CNSHB), BP.3738/39, Conakry, Guinée E-mail: [email protected] |
![]()
![]()
![]()