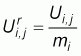![]()
![]()
![]()
E. De Oliveira[48], N. Bez[48] and G. Duhamel[49]
1. INTRODUCTION
The growing capacity of fishermen to repeat fishing operations at some known locations thanks to, for instance, precise maps of past activity and global positioning systems (GPS), is becoming more apparent as fine-scale information on fishing activities becomes available. Aggregated declarations at the level of statistical square levels hides the fine-scale information and the possibility to analyze small-scale fishing strategies. When available, information on fishing effort such as observers’ declarations or vessel monitoring system (VMS) reports may indicate that the tendency of specific-area repeat fishing is sometimes frequent (Rijnsdorp, van Mourik Broekman and Visser 2000). In this paper, we are concerned by the fact that small-scale fishing tactics may interfere in the relation between commercial catch per unit effort (CPUE) and indices of abundance and potentially reduce the relevancy of mean CPUE for resource analysis purposes. The relationship between CPUE and abundance can be biased (e.g. Hillborn and Walters 1987, Rahikainen and Kuikka 2002) as it is the product of several processes such as schooling behaviour of fish, variation in catchability, interactions between fishing vessels and spatial distribution of fishing effort.
In this paper, we consider the trawl fishery for Patagonian toothfish (Dissostichus eleginoides) off Kerguelen Islands in the Indian sector of the Southern Ocean. In spite of difficult conditions of exploitation, the fishery increased until the end of the 1990s. The fishery can be characterized in the context of this paper by the fact that (a) it is mono-specific - toothfish alone is caught, (b) fishermen do a lot of repetitive tows and (c), management rules changed a few years after the fishery started, switching from global to individual quotas. Making use of the spatio-temporal distribution of tows, Bez, De Oliveira and Duhamel (in press) showed that a mean depletion effect of 5 to 10 percent per tow exists under repetitive fishing for toothfish. This local depletion is stronger for icefish (Champsocephalus gunnari) and grey rockcod (Lepidonotothen squamifrons). However these local depletion effects are weak and the fluctuations observed around the decreasing trends can be attributed to changes in the fishing efficiencies from one tow to the next. The objective of this paper is to use the repetitive fishing to estimate fishing efficiency distributions and to interpret their changes with regards to the change in regulations.
Here, "local fishing efficiency" measures the capacity of a unit of effort to catch fish effectively accessible to fishing (Gascuel 1993). As with all components of catchability, the local fishing efficiency should be estimated with CPUEs based on the same underlying fish abundance, which is never possible. Robson (1966) circumvented this difficulty by assuming homogeneity of fish density over quadrats. Laurec (1977) used geostatistical tools to account for the differences between two CPUEs when using them. Here, we use the territorial behaviour of toothfish and the existence of a series of repetitive fishing to build ratios of consecutive CPUEs to estimate fishing efficiency probability distributions.
2. SURVEY AREAS
Two periods of the Patagonian toothfish trawl fishery are considered in this paper: (a) the beginning period of the exploitation (1986-87) under global quota, and (b), the full exploitation phase (1995-1997) under individual quotas. During the first period, 12 vessels were operating (mainly in the fishing zone No 1, Figure 1). During the second period, only two vessels were fishing switching between fishing zones 2 and 3 (Figure 1).
The Patagonian toothfish is a predacious demersal deepwater species (Duhamel 1981, 1992). Its range is the Southern Ocean and the tip of South America. It undertakes long distance migrations of more than 2000 km (Williams et al. 2002) during its life cycle. However, it may also be a territorial fish and we assume that it does not move during a small period of time, i.e. one or two days.
|
FIGURE 1 Crosses represent all the tows performed during the two studied fishing periods (1986-1987 and 1995-1997). Fishing zones are represented and numbered.
|
The presence of observers on board each boat allowed us to obtain accurate and precise information on tow coordinates, with a precision of 1 nm, tow durations and catches. CPUEs were then precisely georeferenced and expressed in hundreds of kg/hour (102 kg·h-1). Catches from tows shorter than 15 minutes have been removed from the dataset. Such short tow durations usually indicate a problem in the trawl behaviour (damaged gear, winches breakdowns, etc.).
Three species of fish were caught: Patagonian toothfish, icefish and grey rockcod. However the spatial distributions of each species was distinct (Figure 2). Each species is fished in a specific area and without overlap with the others. Five fishing areas have then been delimited (Figure 1), the first three being specific for toothfish. Catches located out of these five specific areas have been assigned to a supplementary area, noted as Area 6.
|
FIGURE 2 Fishing zone No6 corresponds to CPues of any of one of the five defined fishing zones (see Figure1). Black: mean CPue of all species, grey: mean CPue of Patagonian toothfish. White: mean CPue of icefish. White with black lines: mean CPue of grey rockcod.
|
Prospected areas for Patagonian toothfish have changed over time. Area1 was more exploited during the first fishing period. This area is dangerous for fishing gear, which resulted in the displacement of the fishery towards two new areas for Patagonian toothfish (Areas 2 and 3) in the late 1990s.
3. METHODS
3.1 Definition of trips
Each fishing season of each vessel has been divided into trips. These consist of sequences of CPUEs performed by a specific vessel in a given fishing area provided that these sequences last more than two days (Figure 3). For each trip, we searched for co-located tows, i.e. at the same location. For each trip, a percentage of co-located catches is then computed.
|
FIGURE 3 Areas of rectangles are proportional to the number of tows.
|
3.2 Definition of series of co-located CPUEs
A series of repetitive fishing operations have been defined so that: (a) each series comprises at least three non-zero CPUEs, (b) the maximum distance between all CPUEs of a series is one nautical miles and (c), the maximum time lag between two consecutive CPUE is two days. We denote by Ui,j the jth CPUE of the ith series, with th jÎ[1, Li] where Li is the length of the ith series (Table 1). Given the constraints used to define the series, Li ³3. All series have been standardized to their average making possible the comparison between them of:
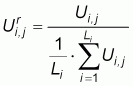
For each fishing period averages profiles of standardized series were then calculated as follows:
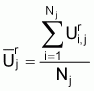
where Nj = the number of available observations at rank j.
We forced series to get at least three observations: N1 = N2 = N3 > Nj>3·
TABLE 1
Notations
|
Notation |
Meaning |
|
i |
Number of data series |
|
Li |
Lenght of the ith data series |
|
j |
Rank number in a serie; jÎ[1, Li] |
|
Ui.j |
jth CPUE of the ith series |
|
mi |
Average value of the CPUEs of the ith series |
|
|
jth CPUE of the ith series relative to the mean of the series |
|
Nj |
Number of available CPUEs at rank j |
|
|
Mean relative CPUE of rank j |
|
zi |
Fish density at the beginning of series i |
|
|
Local fishing efficiency for the series i and rank j |
|
|
Ratio between two successive CPUEs |
3.3 Local fishing efficiency
We considered the reference situation where swept area is constant within series and where no immigration and/or emigration of fish occur during a series. In this case, if we denote zi as the fish density at the beginning of each series and Fi,j as the fishing efficiencies of each tow, the first two CPUEs of a series are given by:
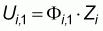
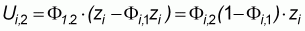
so that after j repetitions:

The ratio ratio
 between two successive
CPUEs is:
between two successive
CPUEs is:
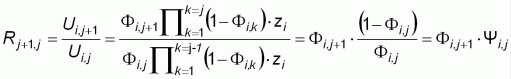
with
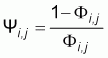
In the simple case where the fishing efficiency is constant,
the model is that of an exponential depletion. Departures from this simple model
imply that the fishing efficiency is variable and it is then modeled by a random
variable, .
.
The ratio of two consecutive CPUEs,
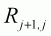 is only dependent on the
fishing efficiency. It is also considered as a random variable. Bez, De Oliveira
and Duhamel (in press) developed a method to estimate the probability density
function of the fishing efficiencies from the experimental histogram of these
ratios. The basics of this method consider that
is only dependent on the
fishing efficiency. It is also considered as a random variable. Bez, De Oliveira
and Duhamel (in press) developed a method to estimate the probability density
function of the fishing efficiencies from the experimental histogram of these
ratios. The basics of this method consider that
 follows a Beta distribution
with unknown parameters n and p. These are estimated by the best
fit of the p.d.f. of R to the histogram of the observations
follows a Beta distribution
with unknown parameters n and p. These are estimated by the best
fit of the p.d.f. of R to the histogram of the observations
 .
.
4. RESULTS
4.1 Quantitative importance of re-fishing
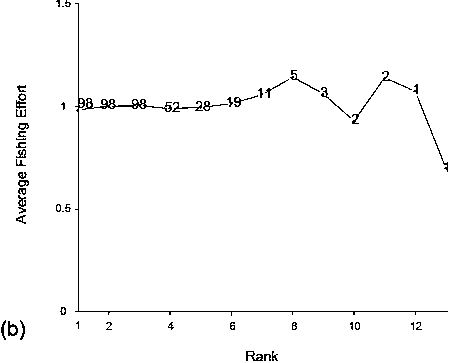
Trawlers operating in the Kerguelen fishery realize a high percentage of co-located catches when targeting Patagonian toothfish. On average, half of the catches performed during a trip are co-located. Thus, of 100tows, 50 will be done at a geographical location that has never been visited before and will not be fished again after that particular tow during that trip. The other 50 will be, at least, positional replicates (duplicates, triplicates, or more). The percentage of co-located tows ranges from 0 to 87 percent (Figure 4) and the majority of them are above 60 percent, which is a clear indication that re-fishing is an important fishing strategy in this deep-sea fishery and that looking at the statistical properties of the co-located CPUEs is a key for the understanding of the dynamic of this fishery. Series of co-located CPUEs include 15 percent and 24 percent of the CPUEs realized in the first and second fishing periods respectively. This indicates that the proportion of successive replicates, which was significant at the beginning of the fishery, increased over the fishery’s history.
|
FIGURE 4
|
4.2 Local depletion
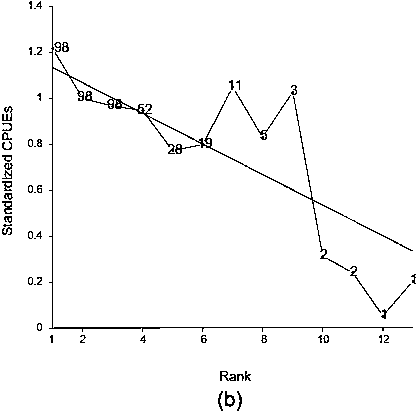
We have established 48 data series in the first fishing period and 98 in the second. The largest series had 13 observations. Amongst these 146 series, not all present showed a decreasing profile. Some do not exhibit any trend and others show an increase. However, relatively to their mean values, series of CPUEs exhibit a mean depletion effect over repetitive trawls (Figure 5). Average rates of depletion for Patagonian toothfish are 4 percent for the first period (Figure 5a) and 7 percent (Figure 5b) for the second period. Strong fluctuations exist around the trends. In particular, for the first period the fourth point of the mean profile departs clearly from the others, even though it is based on 20 observations (Figure 5a). The rate of depletion without this point is 10 percent.
|
FIGURE 5 The number of CPues available at each rank is indicated at each point location. representation of the linear regression line: (a) First fishing period,
(b) second fishing period.
|
4.3 Local fishing efficiency: Patagonian toothfish
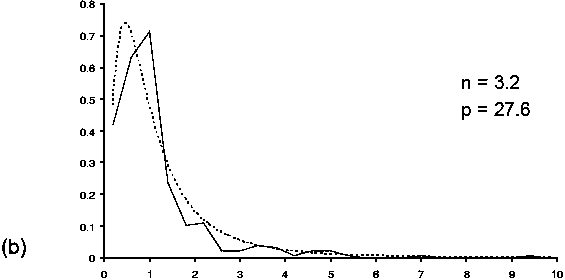
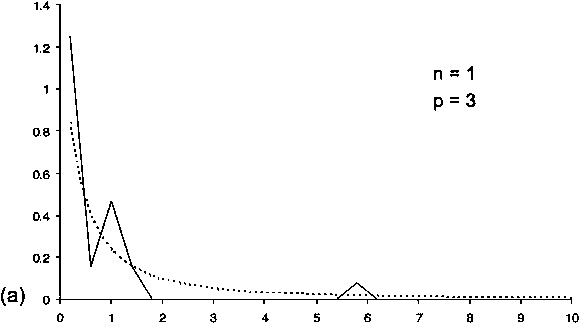
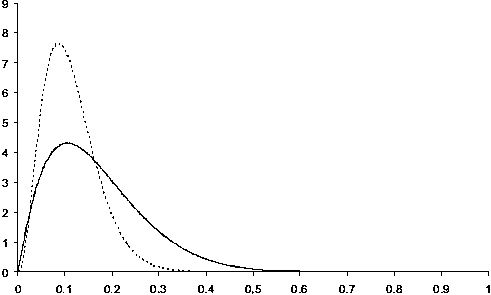
CPUE indices for Patagonian toothfish have been used to estimate the distribution of fishing efficiency for each of two trawling periods. Trawl durations were remarkably stable over repetitions. They do not show any trend, either increasing or decreasing (Figure 6) and we may assume that the area swept is constant within series. Experimental histograms of the ratio between two consecutive CPUEs present differences between the two periods (Figure 7) which induces differences in the estimated probability density functions of the fishing efficiencies for the corresponding periods (Figure 8). At the beginning of the fishery, fishing over efficiencies were spread over a large range of values (from 0 to 0.9) being, on average, equal to 0.28. After 10 years of exploitation, fishing efficiencies were uniformly smaller ranging from 0 to 0.4 and equal to 0.11 on average (Figure 8).
When data permit, the same procedure was applied to the results from individual vessels with the objective of distinguishing between more and less efficient boats. This was possible for vessels No. 19 (in the first period (Figures 9a and 10) and secondperiods (Figures9b and 10)), 29 (first period, Figures 11a and 12) and 54 (second period, Figures 11b and 12). Vessel No. 19 appears to be indicative of the mean behaviours in the fishery as its fishing efficiency resembles that of the set of fishing vessels, i.e. with a wide distribution of efficiencies during the first fishing period and a restricted distribution during the second period (Figure 10).
|
FIGURE 6 (a) first fishing period
(b) second fishing period.
|
|
FIGURE 7 Experimental histogram (the number of observations per class is indicated) and model (dotted line). Values for parameters n and p are indicated: (a) first fishing period,
(b) second fishing period
|
|
FIGURE 8 (continuous line) and for the second fishing period (dotted line)
|
5. DISCUSSION
5.1 Sensitivity of the results to assumptions and to working parameters
We observed that the depletion effect was less and less clear when extending the spatio-temporal window used to build data series. For instance, the mean depletion effect was no longer visible when including tows more than two days apart and/or five nautical miles apart, instead of the 2 days x 1 nm...nm window used in this study. Together with the fact that not all the series show a depletion effect and that some series even show increasing tendencies; this is an indication that the depletion effect is weak. But, to our knowledge, this is one of the few cases where this concept has been demonstrated with actual field data. However, it is often postulated a priori and included in bio-dynamic models (e.g. Maury and Gascuel 2001). Rijnsdorp et al. (2000) working on beam trawlers in the North Sea found a 10 percent decrease in CPUE over periods of 48 hours for flatfish fisheries. We failed to quantify the recover time partly because we could not control the sampling scheme (fishing grounds left out by a given vessel after a sequence of repetitive fishing are usually not left unfished long enough before fishing by other vessels start fishing).
|
FIGURE 9 Experimental histogram and model (dotted line). Values for the parameters n and p are indicated. (a) vessel No 19 - first fishing period
(b) vessel No 19 - second fishing period.
|
|
FIGURE 10 first fishing period (continuous line), second fishing period (dotted line)
|
The weakness of the depletion effect is also an indication that the use of tow-by-tow observers’ declarations need be compulsory. Aggregated data would not have allowed observation of this phenomenon. Relaxing the spatio-temporal window will enable including short spatio-temporal variability, which can be either large or small depending on the targeted species. The impact of this additional variability is as large as the studied signal is weak. The temporal variability is linked to the dynamics and to the mobility of the fish. The territorial behaviour of Patagonian toothfish is responsible for a high heterogeneous spatial distribution on a small scale.
|
FIGURE 11 Experimental histogram (the number of observations per class is indicated) and model (dotted line). Values for parameters n and p are indicated. (a) vessel No 29 - first fishing period and
(b) vessel No 54 - second fishing period.
|
Cells with zero-value CPUEs prevent the computation of a ratio. Ratios with a zero CPUE at the denominator have been grouped into a ‘maximum’ class, which was not used during the statistical analysis. However, only 7 percent of the CPUEs used in the series are null valued, which largely reduces the impact of this problem.
The use of successive CPUEs to analyze local depletion phenomenon is based on the assumption that there is no emigration and immigration of fish from, and into, the fishing area between two successive catches. This assumes that Patagonian toothfish do not move over a period of two days. Thus, all the fluctuations observed in the series of CPUEs around the exponentially decreasing reference model have been interpreted as owing only to variability of fishing efficiency. This simplistic assumption made it possible for the analysis to proceed further, but it means that an increase in CPUEs from one particular tow to the next has been interpreted as an increase of fishing efficiency rather than a movement of fish into the swept area. No biological field observations are available to support this assumption. However, Patagonian toothfish is known to be territorial and to make long migrations on a yearly basis (Williams et al. 2002).
The fishing efficiency has been modeled based on a random variable that can also be justified by the nature of what a fish capture is. Fish capture is dependent on several parameters, which are mainly uncontrolled or unknown. Their mixture at a macro scale results in apparent randomness in the fishing efficiency. The method used in this study makes it possible to estimate the p.d.f. of the fishing efficiency as opposed to an estimate of the fishing efficiency for any single tow. Based on the methods developed by Carle and Strub (1978), we could have used the series of CPUEs to estimate the fish density at the beginning of each series. However this would have required that individual series all had a consistent depletion effect, which was not the case.
|
FIGURE 12
|
A false idea of the sensitivity of the estimations of n and p can be found in Bez, De Oliveira and Duhamel (in press). It appears that slightly divergent fits of the histogram may induce large differences in the distribution of the fishing efficiency, i.e. in the parameters n and p. Still, the difference between the distributions of fishing efficiencies observed between the two fishing periods is large enough to indicate a real change.
5.2 Fishing efficiency, fishing regulation and fishing tactics
|
FIGURE 13
|
Generally speaking, the heterogeneous distribution of fishing effort is partly due to the skippers’ fishing strategy (e.g. cooperation or competition) and can bias the linear relation between abundance and CPUEs. In this study, the strong repeated fishing strategy that is evident is associated with a slight local depletion effect (5 or 10 percent decrease a tow on average). When this happens, co-located CPUE values are not representative of the general local abundance and depending on the intensity of local depletion and the frequency of repeated fishing, this effect could induce biomass underestimations. However, De Oliveira, Bez and Duhamel (2001) showed that in the case of the Patagonian toothfish fishery, this negative effect was compensated by the preferential location of repeated fishing in areas of high toothfish abundance.
On average, fishing efficiency has decreased by half from 1986-87 to 1995-97. Between the two periods, ten years of exploitation took place and induced an overall depletion of the resource (Anon. 1997). Many studies show that the ‘catchability’ is negatively correlated to stock abundance (MacCall 1976, Pitcher 1995, Mackinson, Sumaila and Pitcher 1997). Learning mechanisms may also have taken take place as toothfish is a long-living species. Meanwhile, selection of the fish most reactive against fishing gear is likely to have occured Overall biomass reduction and fish learning processes would favour a decrease of fishing efficiency (Figure 13).
Producing a contrary effect, fishermen’s knowledge about the area and fishing technology (new kinds of net, increase of the fishing gear size, etc.) would have improved between the two study periods. The development of GPS also increased the capacity of fishermen to return to a particular site where appropriate. These elements would favour an increase of fishing efficiency (Figure 13).
Evolutions, or changes, in the management of a fishery modify the fishing behaviour of fishermen. For the Kerguelen fishery, during the first period, the fleet was constrained by a global quota. Fishermen were then seeking the maximum catches to get the maximum possible proportion of the TAC. During the second period, individual quotas were implemented so that each fisherman was assigned an allowed catch quota.
This change in the fishing regulation induced a more relaxed exploitation of the fishing grounds: fishermen took fewer risks and did not search for large catches even when fishing on hot spots. In this latter case, they favoured small (i.e. less efficient) but repetitive tows in order to provide the crew with more regular catches while maintaining the same total catches. Vessel No. 19, which operated in both periods, is an instructive example in this regard. Its technical characteristics and its crew have not changed between the two periods. Yet still, its fishing efficiency became smaller and more uniform.
5.3 "Re-fishing"
Fishermen try to optimize their behaviour with regards to the constraints imposed by the resource and the management. When considering commercial CPUE measurements as samples of a fish resource, one must keep in mind that the sampling objective of commercial boats will differ from that of sampling to estimate a stock abundance. Generally speaking, commercial sampling networks do not allow the analysis of spatio-temporal variability (Laloë, Gaertner and Ménard 2002). The contrast is obvious when comparing the proportion of replicates, i.e. fishing in the same place which is usually zero in scientific surveys, but reached 80 percent in some of the commercial trips reported in this paper.
The consequences of a reduction of fishing efficiency and of repeated fishing on the mean CPUE can be easily indicated, at least qualitatively. If the change in fishing tactics consists of spreading the same total catches over several tows and if the fishing effort needed to get the same amount of fish is smaller when doing one tow instead of several, then the mean CPUE will decrease even though the biomass has not.
This study also makes it possible to revisit the criteria used to select a reference fishery when standardizing CPUEs. When data permit, i.e. when p.d.f. of fishing efficiencies can be computed for different methods or vessels, the reference group can be selected among those with a more stable fishing efficiency.
6. CONCLUSION
Identification and analysis of a series of co-located CPUE measures show, on average, a depletion effect on the Patagonian toothfish stock. From these data series, a model of depletion has been constructed and the p.d.f. of local fishing efficiency has been estimated for each period. On average, fishing efficiency has decreased by half from 1986-87 to 1995-97 and the p.d.f. is less dispersed during the second period. This change has been associated with the change in the fishery regulation, from a global quota to individual quotas.
Each fishery can be described by different parameters, which evolve with time, e.g. fishery regulation, stock abundance, resource behaviour, fishers behaviour or strategies and boat equipment. The improvement of fishermen’s knowledge of Patagonian toothfish and the improvement of fishing technology (e.g. GPS) are expected to increase fishing efficiency. To the contrary, a decrease in Patagonian toothfish biomass due to exploitation is expected to reduce fishing efficiency.
7. LITERATURE CITED
Anon. 1997. Rapport de la seizième réunion du comité scientifique. Comité Scientifique pour la conservation de la faune et la flore marine de l’Antarctique. SC-CCAMLR-XVI, 467 pp.
Bez, N., E.E. De Oliveira & G. Duhamel..(in press). Repetitive fishing, local depletion and fishing efficiencies. ICES Journal of Marine Sciences.
Carle, F.L. & M.R. Strub 1978. A new method for estimating population size from removal data Biometrics 34, 621-630.
De Oliveira, E., N. Bez & G. Duhamel 2001. Etude de l’épuisement local d’un stock à l’aide de données commerciales de captures: cas de la pêcherie des îles Kerguelen. In: A. Biseau et al. (eds), Halieutique: complexité et décision. pp. 13-14.
Duhamel, G. 1981. Caractéristiques biologiques des principales espèces de poissons du plateau continental des îles Kerguelen. Cybium 1, 19-32.
Duhamel, G. 1992. Biologie et exploitation de Dissostichus eleginoides autour des îles Kerguelen (Sous-Division 58-5-1) in 1991 Selected Scientific Papers. Scientific Committee for the Conservation of Antarctic Marine Living Resources. SC-CAMLR-SSP/8: 85-106.
Gascuel, D. 1993. Efforts et puissances de pêche: Redéfinition des concepts et exemple d’application. In Gascuel D., J.L. Durand & A. Fonteneau (Eds.), Les recherches françaises en évaluation quantitative et modélisation des ressources et des systèmes halieutiques, pp. 159-181. ORSTOM Editions.
Hillborn, R. & C.J. Walters 1987. A general model for simulation of stock and fleet dynamics in spatially heterogeneous fisheries.Can. J. Fish. Aquat. Sci44,1366-1369.
Laloë, F., D. Gaertner. & F. Ménard 2002. Modèles mixtes de données halieutiques. Journal de la Société Française de Statistique tome 143, n0 1-2, 131,138.
Laurec, A. 1977. Analyse et estimation des puissances de pêche. J. Const. Int. Explor. Mer 37(2), 173-185
MacCall, G.R. 1976. Density dependence and catchability coefficient in the California sardine, Sardinops sagax caerula, purse seine fishery. California. Cooperative Oceanic Fisheries Investigations Reports, 18. pp. 136-148.
Mackinson, S., U.R. Sumaila & T. Pitcher 1997. Bioeconomics and catchability: fish and fishers behaviour during stock collapse. Fisheries Research (31): 11-17.
Maury O. & D. Gascuel 2002. Local overfishing and fishing tactics: Theoretical considerations and applied consequences in stock assessment studied with a numerical simulator of fisheries. Aquat. Living Resour., 203-210.
Pitcher, T. 1995. The impact of pelagic fish behaviour on fisheries. Scientia Marina 59:(3-4), 295- 306.
Rahikainen, M. & S. Kuikka 2002. Fleet dynamics of herring-change in gear size and implications for interpretations pf catch per unit effort. Can. J. Fish. Aquat. Sci 59, 531-541.
Rijnsdorp, A.D., P.L. van Mourik Broekman & E.G. Visser 2000. Competitive interactions among beam trawlers exploiting local patches of flatfish in the North Sea. ICES journal of Marine Science 57:894-902.
Robson, D.S. 1966. Estimation of the relative fishing power of individual ship. Int. Comm. Northw. Atlant. Fish. Res. Bull. 3, 5-14.
Williams R., G.N Tuck., A.J. Constable & T. Lamb 2002. Movement, growth and available abundance to the fishery of Dissostichus eleginoides Smitt, 1898 at Heard Island, derived from tagging experiments. CCAMLR Science 9: 33-48.
|
[48] Ecole des Mines de
Paris, CGES 35, Rue Saint Honoré, 77 305 Fontainebleau Cedex, France <[email protected]> <[email protected]> [49] Laboratoire d’ichtyologie générale et appliquée Muséum National d’Histoire Naturelle, Paris, France <[email protected]> |
![]()
![]()
![]()