![]()
![]()
![]()
T.T. KOZLOWSKI AND T.E. GREATHOUSEThis paper is based on research conducted in l 5369 while Professor Kozlowski was consultant in tree physiology with the FAO project in Malaysia for " Pilot plantations for quick-growing industrial tree species." At present he is with the Department of Forestry, University of Wisconsin, Madison, Wisconsin. T.E. Greathouse is with the project at the Forest Research Institute, Kepong, Selangor, Malaysia.
IN RECENT YEARS, especially in relation to the many United Nations Development Programme Special Fund (UNDP/SF) projects operated. by FAO, increased interest has been shown in the growth potential of pines introduced to the tropics. Pinus caribaea var. hondurensis is a case in point. In Malaysia, fast early height growth of up to 275 centimetres per year, volume production of about 14 cubic metres per hectare per year and absence of serious insect and fungus pests have been major factors which encouraged planting this species. This paper reports on characteristics and control of the shoot growth of pines introduced to tropical countries and cites some research areas in which physiologists can make important contributions.
Very little is known about the mechanisms controlling shoot growth except that they are similar in pines, such as Pinus taeda and P. elliottii, which grow in the temperate zone (southeastern United States) and in those which grow in the tropics. In both zones shoot growth often involves elongation of more than one terminal bud per shoot per year.
While the internodes of the first bud are expanding early in the growing period, the apical meristem becomes active and initiates primordia which become the leaves and axis of a new terminal bud. The new bud forms rapidly and commonly expands (flushes) to produce a second surge of shoot elongation the same year, often soon after the bud has formed. On any one shoot, additional buds may subsequently form and expand, all within the same year.
The number of stem elongations per year varies with individual trees, with species, and with climatic changes. As many as seven successive buds on a single terminal leader have been observed to elongate in one year (Wakeley and Marrero, 1958). The number of elongations is often positively correlated with tree vigour. Although the leader of an average P. taeda tree does not elongate more than two or three times annually, some pines can be caused. to elongate throughout the year by growing them in a favourable environment (Allen and McGregor, 1962).
Trees exhibiting multiple bud elongations per year may or may not show distinct periods of inactivity between elongations. Cessation of bud activity between elongations of two native New Jersey pine species (P. echinata and P. rigida) reported by Tepper, 1963), corroborated Kramer's (1957) observations on P. taeda grown in a near-optimal controlled environment. However, other studies led to the conclusion that some terminal shoots elongate continuously during much of the annual growing period, even in the temperate zone. Eggler (1961) reported that shoot growth of young P. taeda trees resulted from the sequential elongation of terminal buds. As soon as the year's first terminal bud stopped elongating, a second bud formed and began to expand rapidly. Allen and Scarbrough (1969) also noted rather continuous shoot elongation on ten-year-old P. palustris trees in Mississippi. Similar responses were observed on two Mediterranean pines (P. halepensis and P. laricio).
Apical dominance in the terminal buds of upper branches is reflected in the number of bud elongations per year. For example, Eggler (1961) reported from the United States that upper branches of southern pines elongated up to four times per year compared to zero or one elongation on lower branches. Fielding (1966) made a similar report for P. radiata growing in Australia.
Normal shoot growth of pines in the tropics is similar to normal growth in the temperate zone. The axis elongates by extension of a succession of buds formed on the terminal leader of the main stem. After a period of expansion, growth of the stem ceases briefly and a new terminal bud cluster forms. Shortly thereafter the recently formed buds expand to lengthen further the terminal leader and to produce a whorl of lateral branches. Generally from two to four such periods of growth occur annually, with the terminal leader elongating about 30 to 60 millimetres on the average during each period.
PHOTO: S. HO, FOREST RESEARCH INSTITUTE, KEPONG, MALAYSIA
In contrast, some pine trees develop abnormally (Figure 1) as a result of failure to set buds which would elongate to form lateral branches. Lloyd (1914) described such growth as " foxtailing " because the upper part of the abnormally elongating shoot had a conical or " foxtail " appearance (Figure 2).
Foxtailing, a striking form of apical dominance, often produces trees with up to 6 metres, and occasionally up to 12 metres, of branchless stem. This environmental response is a problem of variable degree wherever pines are grown in the tropics. Among pines reported to show foxtailing growth are P. canariensis, P. caribaea, P. cembroides, P. echinata, P. elliottii, P. kesiya (insularis), P. merkusii, P. oocarpa, P. palustris, P. radiata, P. taeda and P. tropicalis.
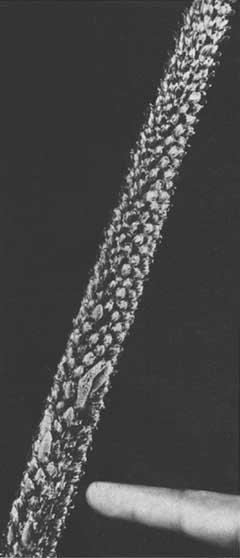
In a foxtail shoot the needles usually decrease in length as the shoot tip is approached. Near the tip the unexpanded needles are tightly packed and enclosed in unbroken sheaths. Below these the needles penetrate their sheaths and increase in length, giving the upper part of the continuously expanding shoot a conical appearance (Figure 2 ). New needle primordia are more or less constantly formed at the apex and are subsequently expanded, resulting in a progression of needle sizes and stages of development.
Needle retention appears to be increased during periods of foxtailing. As observed in Malaysia, the branchless stem portions retain needles during several growth periods (Figure 1), but lose these needles soon after the tree reverts to a normal growth pattern (Figures 3 and 4). A distinctive dimpled bark pattern is created by the long period of needle retention.
Various degrees of foxtailing were observed in large Malaysian plantations of P. caribaea var. hondurensis trees up to 15 years of age. For example: (a) in some trees continuous growth of the terminal leader occurred without pause after field planting, resulting in a single stem with no side branches (Figures 1; and 3); (b) foxtailing began at time of field planting and ceased after a few years. Thereafter a normal pattern of shoot growth occurred (Figure 4); (c) trees grew normally for a while and then foxtailed (Figures 5 and 6); (d) foxtailing began at the time of field planting, ceased after a few years, and subsequently resumed in the terminal leaders. Thus, conversions from continuous shoot elongation to a recurrently flushing pattern, and the reverse, were observed.
Although most reports of foxtailing refer to the phenomenon as a characteristic of the terminal leader (Lückhoff, 1964; Lanner, 1964, 1966), many examples of foxtailing of lateral shoots were observed in Malaysia (Figures 7 and 8). Either gradually or abruptly such laterals became vertically oriented. The excised apical parts of foxtailed branches were morphologically similar to those of foxtailed terminal leaders. Often the alternation of normal and foxtailed growth produced individuals of grotesque form (Figure 8).
In a personal communication A.F.A. Lamb cited the observations which caused him to conclude that foxtailing tends to be a juvenile response. In Malaysia many trees in the oldest plantations (15 years) were still exhibiting foxtail growth.
Most foresters consider foxtailing be an undesirable trait, as it is associated with wind breakage, reduction in diameter growth, absence of latewood: formation, presence of compression wood and restricted production. On the other hand, pending, investigations of other wood qualities, it may one day be desirable to breed foxtailed trees for knot-free wood.
REPRODUCTIVE GROWTH
Foxtailed trees differ particularly from norm al trees in reproductive growth characteristics. In Malaysia normal trees of P. caribaea var. hondurensis produced pistillate strobili in the top fourth of the live crown. On occasional trees pistillate strobili were produced on lower branches as well. Generally, more pistillate strobili per branch were observed on upper than on lower branches. Whereas four to five of these often were borne per upper branch, only one or two per :lower branch were noted. Pollen production in normal trees tended to be localized in the lower branches and in third- or fourth order side branches. Sometimes, however, apical shoots, as well as terminal shoots of primary branches produced pollen. In normal trees up to 15 years old there was a dearth of pollen production on any given day for receptive strobili. Many cones contained only one or two viable seeds.
With few exceptions, during periods of apical foxtail growth no strobili were found on trees up to 15 years old. Occasionally, however, the terminal shoot produced a large number (often several hundred) of staminate strobili which, at a given time, were in various stages of development (Figure 9). These pollen-producing strobili appeared in positions at which on normal trees, and on most foxtailed trees, needle fascicles developed. When a foxtailed tree reverted to a normal, recurrently flushing pattern of shoot growth and produced a whorl of lateral branches, pistillate strobili and/or staminate strobili generally appeared on these lateral branches, as on completely normal trees.
Foxtailing of tropical pines appears to be a strongly inherited growth phenomenon, with its expression modified considerably by site and climatic factors. In Australia, an unselected crop of P. caribaea var. hondurensis had a much higher frequency of foxtails than unselected crops of var. caribaea or var. bahamensis. However, progeny from selected var. hondurensis parents had a lower incidence of foxtailing, with the amount of reduction depending on the degree of control of the parentage (Slee and Nikles, 1968). In Malaysia the amount of foxtailing of P. caribaea var. hondurensis differed markedly from site to site. It tended to be extensive (up to 40 percent) on some lowland sites where consistently high temperatures and heavy nonseasonal rainfall (approximately 254 centimetres per year) favoured continuous shoot growth. Foxtailing appeared to be less prevalent in the Cameron Highlands of Malaysia at elevations of about 1 520 metres than at lowland sites, probably due to slower growth at the high elevations.
TABLE 1. - VARIATIONS IN DISTRIBUTION OF PRECIPITATION IN NORTHWEST (LANGKAWI; KEDAH) AND CENTRAL MALAYSIA (KUALA LUMPUR SELANGOR)¹
|
|
Langkawi, Kedah ² |
Kuala Lumpur, Selangor ³ |
||
|
Precipitation |
Maximum in 24 hours |
Precipitation |
Maximum in 24 hours |
|
|
Millimetres |
Millimetres |
|||
|
January |
34 |
38 |
170 |
132 |
|
February |
46 |
26 |
166 |
126 |
|
March |
113 |
64 |
232 |
119 |
|
April |
213 |
71 |
268 |
134 |
|
May |
250 |
145 |
211 |
133 |
|
June |
282 |
143 |
122 |
100 |
|
July |
274 |
75 |
104 |
92 |
|
August |
286 |
148 |
160 |
112 |
|
September |
389 |
89 |
183 |
194 |
|
October |
401 |
86 |
281 |
221 |
|
November5 |
208 |
64 |
257 |
122 |
|
December |
32 |
37 |
239 |
108 |
|
Average annual |
2529 |
|
2394 |
|
¹ Rainfall records (1879-1958) compiled by Drainage and Irrigation Department, 1961. Government Press, Kuala Lumpur, Malaysia. - ² Station at Ulu Malaka, about 3.5 kilometres from seed orchard, data for 1946-58. - ³ Station at Tanglin Hospital, data for 1879-1958. - 4 In an average year, the first half to two thirds of March is also " dry " (i.e., c 30 millimetres). - 5 In the average year, the last third to half of No, ember is also dry.
FIGURE 3. - Five-year-old normal branched trees and branchless foxtail forms.
A site on Langkawi Island in northwest Malaysia with a distinct dry season (e.g., less than 61 millimetres of precipitation per month) lasting 3 to 4 months (Table 1) has been selected as a P. caribaea trial seed orchard site. Measurements will be taken monthly for at least 18 months to see if there is a correlation between height growth, foxtailing and rainfall patterns.
Evidence in support of marked climatic control of the foxtailing response comes from various sources. In South Africa Lückhoff (1964) found that foxtailing of P. caribaea was reduced at high elevations and latitudes with cool temperatures. For example, along the coast of Zululand (average altitude of 45 to 60 metres) foxtails of P. caribaea var. hondurensis averaged 43 percent; at Ntsubane (altitude approximately 460 metres) 26 percent; and at Dargal in the Natal Midlands (altitude approximately 1 200 metres) 13 percent. The percentage of crooked stems also was correlated with height above sea level (temperature regime). In Queensland, Australia, Slee and Nikles (1968) recorded a higher incidence of P. caribaea foxtails at Beerwah than at Bowenia, and attributed the difference to the more favourable environment at Beerwah. Further evidence of the marked effect of climatic control of the foxtailing trait comes from data showing little foxtailing of P. radiata in seasonal conditions of the Australian Capital Territory and abundant foxtailing in the less seasonal climate of Hawaii (Fielding, 1960; Lanner, 1966).
Intense competition can also inhibit shoot growth and suppress the foxtailing trait. For example, foxtailing does not occur in many native P. caribaea stands in British Honduras. On some soils, however, where weed competition is light the limitations on continuous growth are removed and foxtailing of pines does occur, emphasizing that climate is not the only environmental factor controlling foxtailing.
Effective reduction of foxtailing has been achieved in Australia by intensive phenotypic selection followed by progeny-test selection against the foxtailing trait (Nikles, personal communication). Foxtails present; in young stands often were removed in the thinning when trees were about 6 to 9 metres high. Early thinning of P. caribaea var. hondurensis stands in Queensland removed foxtailed trees and did not induce foxtailing in residual trees.
INTERNAL CONTROL OF SHOOT GROWTH
Hereditary and environmental controls of seasonal shoot growth of pines are mediated through internal physiological processes (Kramer and Kozlowski, 1960; Kozlowski, 1969). To attempt to understand he nature of internal control of seasonal shoot growth it its important to recognize that in different climates, various degrees exist of internal control of shoot growth suspension. For example, recurrently flushing pines in the temperate zone (e.g., P. taeda of the southeastern United States) undergo several sequential periods of bud " quiescence " during the summer. Quiescence may be considered to be a simple type of reversible growth inactivity which occurs several times during the growing period and results in multiple flushes of growth per year. However, following the final annual flush of shoot growth in late autumn the buds gradually enter a very deep-seated state of dormancy which. cannot be broken by several of the environmental conditions which caused it. Buds in this condition may be predisposed to grow but will not do so even if placed in a favourable environment because internal physiological conditions inhibit bud growth. Such a state of true dormancy is entered into rather gradually. The tissues are in a state of deep dormancy only during a middle phase of the overall winter rest, period. True bud dormancy can be readily induced by low temperatures, short days, or interactions of both. In the temperate zone bud dormancy is commonly broken by extended periods of low temperatures. Complete removal of dormancy requires exposure of buds to a critical number of hours of a low threshold temperature. The length of exposure to cold required for breaking true bud dormancy varies greatly among species, varieties, and different buds on the same trees (Kramer and Kozlowski, 1960).
FIGURE: 6. - Five-year-old tree which grew normally at first and then grew in a f oxtail pattern.
Externally, the sequential phases leading to development of bud dormancy are characterized by the slowing down and finally the ending of internode expansion. Internally, however, the onset of dormancy involves accelerated meristematic activity. Eventually a group of primordia develop on a short axis in the dormant bud.
When a recurrently flushing pine of the temperate zone (e.g., P. taeda) is moved to a tropical climate it is not subjected to the severe environment that induces a very deep-seated state of bud dormancy. In the tropics the buds of such a tree are more likely to lapse into periods of quiescence rather than true bud dormancy.
From the foregoing considerations it should be clear that an overall theory of the mechanism of internal regulation of shoot expansion for pines of both tropical and temperate zones should account for three primary phases of bud development: (a) phasic development of dormancy leading to a truly dormant state; (b) breaking of dormancy leading to a nondormant state; and (c) growth initiation in the spring leading to a steady-state development (Smith and Kefford, 1964). Thus, bud inactivity can range from quiescence to true dormancy; but as transitional phases also occur, bud inactivity in pines can be characterized as a quantitative state, as has been done for some species of angiosperms (Kawase, 1961).
Considerable evidence shows that environmental factors influence bud dormancy by controlling the synthesis, translocation or destruction of varying amounts and kinds of hormonal growth regulators. For many years the regulation of bud dormancy was thought to be a single process rather than a series of transitional ones. Many early theories proposed that the overall process was limited by a single endogenous chemical regulator, and much emphasis was placed on control of Dormancy by auxin levels alone. Kozlowski (1964) reviewed the literature on these early theories.
FIGURE 7. - Marked apical dominance in lateral shoots of a five-year-old foxtail tree.
Available evidence now shows that bud dormancy is regulated by interactions among endogenous growth promoters and inhibitors. Various proportions Of growth promoters can produce different rations of cell division and expansion, thereby regulating growth and differentiation At least three major and several minor phases of overall bud development are recognized, with each controlled by growth-regulator patterns in the buds. Growth promoters, such as auxins, gibberellins, and cytokinins, as well as growth inhibitors appear to be variously involved. A key role of inhibitors (e.g., abscisic acid) in development of dormancy is indicated, whereas growth promoters, especially gibberellin-like substances, play a major role in dormancy release. Interactions among various growth promoters and inhibitors also are important in regulating the overall dormancy phenomenon. The primary evidence indicating that development of dormancy is largely regulated by endogenous inhibitor s may be summarized as follows:
(a) under short-day conditions the leaves of many woody plants inhibit growth of the shoot tip;(b) greater amounts of inhibitors are found in Cleavers and buds of many woody plants under short-day than under long-day conditions;
(c) when inhibitors are extracted from leaves of a dormant woody plant and reapplied to plants of the same species which were not dormant prior to the application of inhibitors, shoot elongation stops and sequential development toward a dormant state is initiated (Warring' 1965).
Both growth promoters and growth inhibitors are present during various transitional phases of dormancy. Whereas inhibitors appear to play a primary role in promoting dormancy, the breaking of dormancy appears to be associated largely with activity of hormonal growth promoters, especially gibberellin-like substances. Possibly other growth promotes s are involved and balances and interactions among them play a role in dormancy release. The observation that long days increase levels of growth promoters in buds is consistent with these conclusions. Inhibitors do not always decrease with release from dormancy (Wareing, 1965). Evidence is available of interactions of growth inhibitors (e.g., abscisic acid) and growth promoters; for example, gibberellic acid counteracts the effects of inhibitors (Thomas, Wareing and Robinson, :l965).
Seasonal variations in kinds and amounts of hormonal growth regulators present in trees are rather consistent with a theory of shoot growth control which involves changes in balance of growth promoters and inhibitors. Whereas dormant buds contain little growth promoter, swollen or opening buds contain large amounts. Both growth promoters and inhibitors have been found in pine buds (Allen, 1960; Ogasawara, 1961a, 1961b; Giertych, 1960; Giertych and Forward, 1966). The levels of growth promoters usually are higher in terminal buds than in lateral buds. Giertych and Forward (1966) found highest levels of growth promoters in the lower or midcrown of P. resinosa. But there also were higher levels of inhibitors in the lower or midcrown than in the upper crown, suggesting that the upper crown had a balance :in favour of growth promoters.
Considerable has been given in this paper to internal hormonal controls of shoot growth where true bud dormancy is involved. This was done in order to review the current state of knowledge and to emphasize the importance of changes in hormonal balances in controlling shoot growth. However, as bud dormancy may be considered an extreme kind of shoot growth inactivity and quiescence an intermediate state, much more information is needed on hormonal relations of tropical pines, which have quiescent rather than truly dormant buds during much of the year. Physiological and biochemical research on internal control of shoot growth of tropical pines has been very limited. Nevertheless, evidence from physiological studies of shoot growth of a number of recurrently flushing species indicates that hormonal controls are importantly involved. A reasonable working hypothesis is that the balance between hormonal growth promoters and inhibitors changes during the flushing-quiescence-flushing sequence of shoots of tropical pines. It is tempting to speculate that in a foxtailing shoot there is a continuous balance in favour of one or more growth promoters. However, more research is needed on several aspects of internal changes in hormonal growth regulators of tropical pines. Among the major questions which need to be answered are the following:
1. What differences occur in amounts and kinds of growth promoters and inhibitors during sequential flushes of shoot growth in tropical pines?2. What differences occur with time in amounts and kinds of growth promoters and inhibitors of foxtailing shoots?
3. What differences occur in amounts and kinds of growth promoters and inhibitors of a recurrently flushing species in the temperate zone during entrance to and exit from quiescent phases as against a truly dormant phase?
4. How does the annual cycle of growth regulator changes in such a species vary in the temperate zone from the changes in the same species when introduced into the tropics?
5. What are the differences in amounts and kinds of hormonal growth regulators of shoots in different crown locations of tropical pines and of recurrently flushing pines of the temperate zone?
6. What combinations of environmental factors will induce various internal hormonal balances which are associated with predictable shoot growth patterns ?
Excellent opportunities exist for answering some of these questions and thereby helping the understanding of internal growth control of tropical pines; and the answers may be very useful in growing tropical pines of predictable form by use of appropriate genetic materials and by environmental controls.
ALLEN R.M. 1960. Changes in acid growth substances in terminal buds of longleaf pine saplings during the breaking of winter dormancy. Physiologia Plantarum, 13: 555-558.
ALLEN, R. M. & MCGREGOR, W. D. H. 1962. Seedling growth of three southern pine species under long and short days. Silvae Genet., 11: 43-45.
ALLEN, R. M. & SCARBROUGH N. 1969. Development of a year's height growth in longleaf pine saplings. USDA. Forest Research Paper so-45.
BARRETT, W. H. G. & GOLFARI L. 1962. Descripción de dos nuevas variedades del "Pino del Caribe" (Pinus caribaea Mor.). Caribbean Forester, 23: 59-71.
CHALMERS, W. S. 1962. The breeding of pine (Pinus caribaea Mor.) and teak (Tectona grandis) in Trinidad - some early observations. Caribbean Forester, 23: 100-111.
EGGLER, W. A. 1961. Stem elongation and time of cone initiation in southern pines. For. Sci., 7: 149-158.
FIELDING, J. M. 1960. Branching and flowering characteristics of Monterey pine. Canberra, Forestry and Timber Bureau. Bulletin No 37.
FREEIZAILLAH BIN CHE YEOM. 1966. Some notes on Pinus caribaea Mor. grown in Malaya. Forestry Department of Malaya, Forest Research Institute. Research Pamphlet No 54.
GIERTYCH, M. M. 1964. Endogenous growth regulators in trees. Bot. Rev., 292-311.
GIERTYCH, M. M. & FORWARD, D. F. 1966. Growth regulator changes in relation to growth and development of Pinus resinosa Ait. Can. J. Bot., 44: 717-738.
KAWASE, M. 1961. Growth substances related to dormancy in Betula. Proc. Am. Soc. hort. Sci., 78: 532-544.
KOZLOWSKI T. T. 1964. Shoot growth in woody plants. Bot. Rev., 30: 335-392.
KOZLOWSKI T. T. 1969. Tree physiology and forest pests. J. For., 67: 118-123.
KRAMER, P. J. 1967. Some effect of various combinations of day and night temperatures and photoperiod on the height growth of loblolly pine seedlings. For. Sci., 3: 45-55.
KRAMER, P. J. & KOZLOWSKI, T. T. 1960. Physiology of trees. New York, McGraw-Hill.
KUMMEROW, J. 1962. Uber Wachstumsanomalien bei Pinus radiata unter Tropischen Bedingungen. Ber. Dtsch. Bot. Gesell., 75: 37-40.
LANGE, P. W. 1966. The occurrence and development of abnormal long leaders in Pinus radiata D. Don in the Cape Midlands. Forestry in South Africa, 7: 1-10.
LANNER, R. M. 1964. Modifications in the growth habits of exotic trees in Hawaii. Proc. Soc. Am. For., 36-37.
LANNER, R. M. 1966. The phenology and growth habit of pines in Hawaii. U.S. Forest Service Paper PSW-29.
LLOYD, F. E. 1914. Morphological instability, especially in Pinus radiata. Bot. Gaz., 57: 314-319.
LÜCKHOFF, H. A. 1964. The natural distribution, growth and botanical variation of Pinus caribaea Mor. and its cultivation in South Africa. Ann. Univ.. Stellenbosch, 39: 1-160.
MCWILLIAM, J. R. & RICHARDS, B. N. 1955. Caribbean pine in Queensland, some notes on its behavior and potentialities. Queensland Forest Service Note No 4.
OGASAWARA, R. 1961a . Studies on auxins and inhibitors in Pinus thunbergii. J. Jap. for Soc., 43: 50-54.
OGASAWARA, R. 1961b. Studies on auxins and inhibitors in the buds of Pinus strobus. J. Jap. for.. Soc., 43: 307-310.
ROMBERGER, J. A. 1963. Meristems, growth, and development in woody plants. U.S. Department of Agriculture, Forest Service. Technical Bulletin N° 1293.
SLEE, M. U. & NIKLES, D. G. 1968. Variability of Pinus caribaea (Mor.) in young Queensland plantations. Proc. ninth Commonwealth for Conf., 1-50.
SMITH, H. & KEFFORD, N. P. 1964. The chemical regulation of the dormancy phases of bud development. Am. J. Bot., 51: 1002-1012.
STREETS, R. J. 1962. Exotic forest trees in the British Commonwealth. Oxford, Clarendon Press.
TEPPER, H. B. 1963. Leader growth of young pitch and short leaf pines. For. Sci., 9: 344-353.
THOMAS, T. H., WAREING, P. F. & ROBINSON, P. M. 1965. Action of the sycamore "dormin" as a gibberellin antagonist. Nature, Lond., 205: 1270-1272.
WAKELEY, P. C. & MARRERO, J. 1958. Five-year intercept as site index in southern pine plantations. J. For., 56: 332-336.
WAREING, P. F. 1965. Dormancy in plants. Science Progress, 53: 529-537.
|
CANADIAN JOURNAL OF FOREST RESEARCH In 1971 the National Research Council of Canada will begin the publication of a new international journal, the Canadian Journal of Forest Research. It will publish in English or in French reports of original scientific research and discussions of them. All aspects of forestry science will be covered and interdisciplinary papers will be especially welcome. The journal will be issued quarterly beginning in March. Manuscripts should be sent to the editor, Dr. J.L. Farrar, Faculty of Forestry, University of Toronto, Toronto 5, Ontario. Annual subscription rates, in Canadian funds, are: multi-user, $10.00; personal, $5.00. Subscriptions should be sent to: Administration, National Research (council of Canada, Ottawa 7, Canada, with remittances made payable to the Receiver General of Canada. |
![]()
![]()
![]()
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}