![]()
![]()
![]()
In the absence of a commercial or recorded subsistence fishery, it was not possible to estimate the standing stock or total population density of the small water impoundments. However valuable preliminary data discussed below are provided by standardized test fishing made in selected impoundments. From these, an index of abundance can be estimated by the catch per unit effort (CPUE).
The standardized method applied initially in Lesotho followed closely the Drottningholm method (Appendix 2) using benthic and pelagic multimesh monofilament nylon gillnets consisting of 14 randomly distributed panels of various mesh sizes (Table 6.1.) in a near exponential series. Each panel is 3 m long, and hence the total length of each net is 42 m. The benthic nets are 1.5 m while the pelagic nets are 6 m deep. The pelagic nets are marked at 3 m depth so that the catch can be separated into the upper and lower 3 m portions. The total area of the benthic nets is 63 m2 while that of the pelagic nets is 252 m2.
Nets were set overnight from sunset to sunrise. They were usually set between 16h00 and 18h00 and taken up the following morning, usually between 7h00 and 9h00. The nightly catch (approximately 15hrs) from each benthic net was considered as one effort.
Table 6.1 Specification of mono - and multifilament gillnets used by ALCOM in the Lesotho Lowlands.
| Order | Monofilament gillnet | Multifilament gillnet | ||
| Mesh size stretched (mm) | twine diameter (mm) | Mesh size stretched (mm) | twine tickness (denier) | |
| 1 | 10 | 0.10 | 35 | 210/4 |
| 2 | 16 | 0.10 | 45 | 210/4 |
| 3 | 32 | 0.15 | 57 | 210/6 |
| 4 | 150 | 0.25 | 73 | 210/6 |
| 5 | 76 | 0.18 | 93 | 210/9 |
| 6 | 50 | 0.15 | 118 | 210/9 |
| 7 | 25 | 0.12 | 150 | 210/9 |
| 8 | 66 | 0.18 | ||
| 9 | 100 | 0.20 | ||
| 10 | 44 | 0.15 | ||
| 11 | 86 | 0.20 | ||
| 12 | 60 | 0.15 | ||
| 13 | 120 | 0.25 | ||
| 14 | 20 | 0.12 | ||
Four benthic nets were used by depth zone and four pelagic nets in reservoirs over 6m deep. The pelagic nets are to be set over the deepest part of the lake and linked together as one unit.
Monofilament gillnet were set on 8 occasions from 1990 to 1992. However, since the fish species found in Lesotho reservoirs are large, the monofilament nets were badly damaged each time they were set. Underwater obstacles also frequently ruined the nets. Much time was spent in mending the nets and some mesh sizes were no longer the same.
A revised test fishing method using benthic and pelagic multimesh multifilament nylon gillnets (made in East-Asia and mounted locally) was therefore proposed. The gillnets consist of 7 panels of various mesh sizes in an exponential series (Table 6.1.). Each panel is 6 m long, and hence the total length of each net is 42 m. Otherwise the nets were similar to the Drottningholm nets.
Fishing effort similar to the Drottningholm method was also used for setting 4 benthic nets per depth zone and 4 pelagic nets over 6m depth. Benthic nets were not set in depth zones where the dissolved oxygen concentration was less than 3 mg/l, since it is unlikely that fish are present under these conditions.
Test fishing with multifilament gillnets were performed on 10 occasions from October '93 to April '94.
The fish caught in each net were sorted by species. Jubb (1967), supplemented by Bruton et al. (1982) were used to identify the species. Fish were measured individually to the nearest millimetre in both total and fork length and weighted to the nearest gram. Catch per unit effort (CPUE) was calculated as the sum of fish caught (weight or number) by one net in one night. Empty nets were included when calculating average catch per effort.
In conjunction with each test fishing exercise, selected water quality measurements were made. Temperature readings to 0.1°c were taken with a thermistor at one meter intervals. Dissolved oxygen was measured to 0.1 mg/l at 1 m intervals until less than 2 mg/l with an oxymeter. A secchi disk reading was taken and the pH measured.
Morphometric data on small water bodies and physio-characteristics of water in selected reservoirs have been exhaustively described in an earlier report (Tilquin & Maseatile, 1994). Background data are summarized for each reservoir selected in Appendix 1.
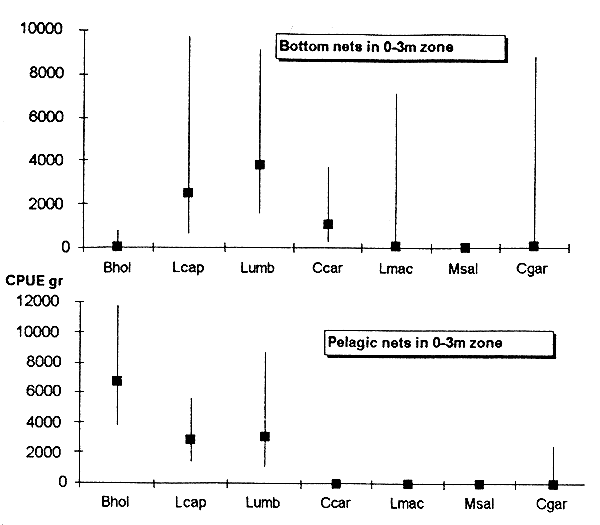
The Maqalika dam (64ha) was assessed on four occasions. Results of the three first test fishing exercises using monofilament gillnet were described in an earlier report (Mofolo et al. 1993b). Figure 6.1. shows that Labeo umbratus is the major species caught in the dam from 1990 to 1993 since Labeo capensis and Barbus aeneus individuals have got less abundant. The fish stock assessment of November 1992 is not taken into consideration as only bottom nets were used. As the catches from bottom and pelagic nets were recorded by depth zone, Figure 6.2. shows that most fish for all species except C. gariepinus were caught in the 0–3m zone.
Figure 6.1. CPUE from three test fishing undertaken in Maqalika dam, 1990–1993.
Baen=Barbus aeneus, Lcap=Labeo capensis, Lumb=Labeo umbratus, Ccar=Cyprinus carpio, Lmac=Lepomis macrochirus, Cgar=Clarias gariepinus.
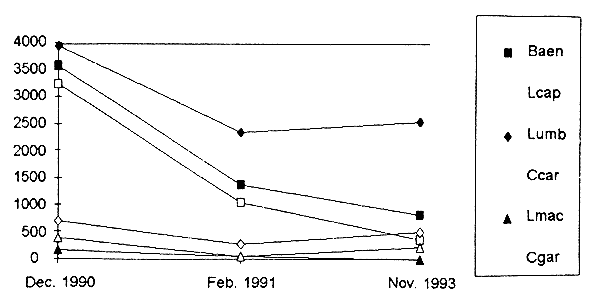
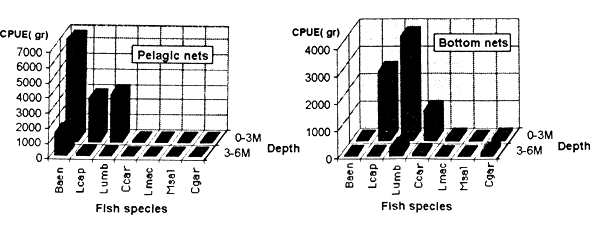
Figure 6.2. CPUE by net type and depth zone from test fishing in Maqalika Reservoir, 10 Dec. 1990.
Summary data on fish stock assessment in selected reservoirs
| Reservoirs greater than 10ha | |||||||||||||
| CPUE (g) | |||||||||||||
| Name | Date of Assessment | Surface area (ha) | Limit depth D.O. 3mg/l | Surface temperature (°C) | pH | Transparency Secchi disk (m) | Baen | Ccar | Cgar | Lmac | Leap | ||
| Maqalika dam | 16.11.93 | 64 | 6m | 25 | 7.8 | 0.25–50 | 817 | 512 | 221 | 0 | 352 | ||
| Sebaboleng dam | 7–9.12.90 | 18 | 3m | 23 | 8 | 0.25–0.30 | 0 | 389 | 368 | 57 | 0 | ||
| Thaba-Phatsoa | 3–5.12.90 | 25 | 6m | 21 | 7.6 | 1 | 5377 | 0 | 0 | 0 | 0 | ||
| Releting dam | 30.11.93 | 21 | 1.5m | 22 | 7.6 | 0 | 0 | 0 | 2134 | 139 | 0 | 0 | |
| Reservoirs smaller than 10ha | |||||||||||
| Name | Date of assessment | Surface area ha | Botom nets Number | Surface temperature | pH | D.O. 3mg/l | Transparency secchi disk | Fish Species | No. of fish caught | Total weight (g) | CPUE (g) |
| Luma lake | 29.11.93 | 7.5 | 5 | 19.7 | 7.7 | 3m (max. depth) | 0.00 | C. carpio | 3 | 696 | 139 |
| Nyenye dam | 7.12.93 | 5.2 | Seine nets 3 batches | 22.0 | 8.8 | 2m (max. depth) | 0.05 | C.carpio | 33 | 5178 | NA |
| L. macrochirus | 12 | 890 | NA | ||||||||
| Mejametalana dam | 29.11.90 | 4.3 | 4(0–3m) | 22.4 | 7.7 | - | - | No fish caught | NA | NA | NA |
| 4(3–6m) | |||||||||||
| Borokhoaneng dam | 18.11.93 | 1.6 | 3 | 21.0 | 7.9 | 2.5m (max. depth) | 0.00 | C. carpio | 7 | 7079 | 2360 |
| C. gariepinus | 1 | 654 | 218 | ||||||||
| Thaba-Khupa No.1 dam | 6.12.93 | 3 | 4 | 20.5 | 7.65 | 1.4m | 0.00 | C. carpio | 7 | 6733 | 1683 |
| Leshoele No. 2 dam | 15–16.03.94 | 0.8 | 4 | 20.4 | 7.4 | 1.4m | 0.40 | No fish caught | NA | NA | NA |
| Rakolo No 1 dam | 23–24.03.94 | 1.8 | 4 & 2 floating nets | 24.0 | 8.8 | 3m | 0.05 | No fish caught | NA | NA | NA |
| Rakolo No.3 dam | 16–17.03.94 | 1.5 | 5 | 24.5 | 8.6 | 1.5m | 0.20 | C. carpio | 11 | 657 | 131 |
| JTC dam | 22–23.03.94 | 1 | 4 | 23.0 | 8.6 | 1.5m | 0.05 | C. carpio | 3 | 1711 | 428 |
| Agri College dam | 13–14.03.94 | 1 | 4 | 22.3 | 7.45 | 0.8m | 0.00 | C. carpio | 174 | 14268 | 3522 |
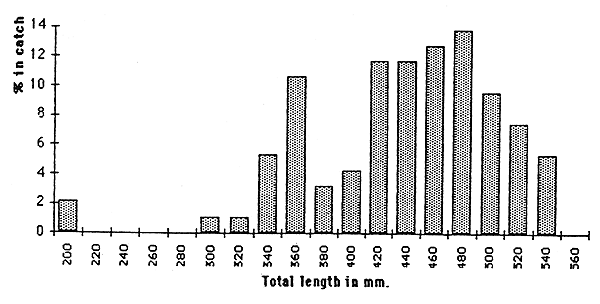
Thaba-Phatsoa dam, belonging to the foothills zone, has only two species, Barbus aeneus and Micropterus salmonides. Both species were caught in the benthic nets, with Barbus aeneus dominating; only Barbus aeneus was caught in the pelagic nets (Figure 6.3.). Length frequency distributions for Barbus aeneus are given on Figure 6.4.
Baen=Barbus aeneus, Lcap=Labeo capensis, Lumb=Labeo umbratus, Ccar=Cyprinus carpio, Lmac=Lepomis macrochirus, Cgar=Clarias gariepinus.
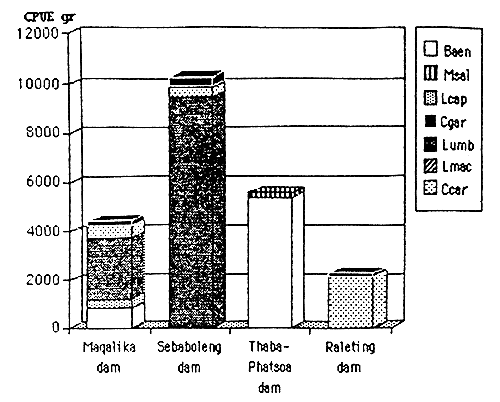
Figure 6.4. Length frequency distribution for Barbus aeneus in Thaba-Phatsoa dam
Raleting dam, belonging to the class Ib, is a large but silted and turbid impoundment where few C. carpio and C. gariepinus were caught.
Nine manmade reservoirs and one natural impoundment smaller than 10ha were assessed. Only individuals of three species were caught, C. carpio, C. gariepinus and Labeo macrochirus. The two latter species were caught only in Borokhoaneng and Nyenye dams respectively. (Table 6.2 and Figure 6.4).
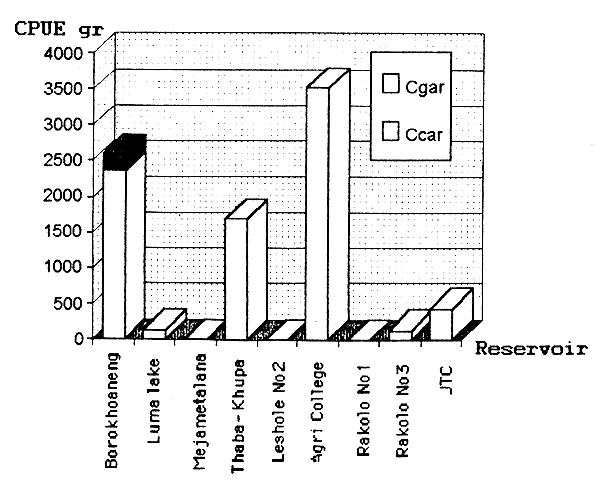
Breeding population seems to occur only in two dams, Nyenye and Agri College Maseru, where several fingerling were caught. In the other dams assessed, few large fish or no fish were encountered.
A rapid oxygen depletion is observed between 1 and 2m depth in most of the reservoirs. No fish could be found below 2m. A relatively high pH value above 8 is also recorded in many dams. All these small dams are very turbid. The secchi disk could not be read over 5cm depth except for Leshoele No. 2 and Rakolo No. 3 where the transparency was around 0.2–0.4m.
The lowland reservoirs sampled have a fish community of seven recorded species amongst the 11 found in Lesotho lowlands (Table 6.3.and Figure 6.6.).
Figure 6.6. The fish fauna in sampled lowland reservoirs (drawings from Skelton, 1994

a: Smallmouth yellow fish, Barbus aeneus
b: Sharptooth catfish, Clarias gariepinus
c: Orange River labeo, Labeo capensis
d: Mud mullet or Moggel, Labeo umbratus
e: Bluegill sunfish, Lepomis macrochirus
f: Common carp, Cyprinus carpio
g: Largemouth bass, Micropterus salmonides
Table 6.3 Indigenous and exotic fish species in the Lesotho Lowlands (from Chondoma, 1988).
| SPECIES | COMMON NAME | ORIGIN |
| Barbus anoplus | Chubby-head barb | Indigenous |
| Barbus aeneus | Smallmouth yellowfish | Indigenous |
| Barbus kimberleyensis | Largemouth yellowfish | Indigenous |
| Barbus paludinosus | Straightfin barb | Indigenous |
| Labeo capensis | Orange River labeo | Indigenous |
| Labeo umbratus | Mud mullet or moggel | Indigenous |
| Clarias gariepinus | Sharptooth catfish | Indigenous |
| Gephyroglanus | Rock catfish | Indigenous |
| Cyprinus carpio | Common carp | Exotic. Introduced from South Africa in 1896, 1915 |
| Micropterus salmonides | Largemouth bass | Exotic. Introduced from South Africa during the mid-1930s. |
| Exotic. Introduced from South Africa during the mid-1930s. | ||
| Lepomis macrochirus | Bluegill sunfish |
The most commonly occurring indigenous species is the African catfish (C. gariepinus). This is probably because of the ability of this fish to survive in little or no water for long hours. Three other indigenous species -- L. umbratus, L. capensis and B. aeneus -- were found but only in the largest reservoirs belonging to the Class IA (Table 6.4.).
It is noticed that three species have not been found in reservoirs so far: Barbus anoplus, Barbus kimberleyensis and Gephyroglanis sclateri. They are known to exist in lowland rivers (Table 6.3 and Figure 6.6).
Other species found are exotic. The common carp now existing in many reservoirs is a result of a stocking programme that was done immediately after the establishment of a national fish hatchery in 1964 (Chondoma, 1988). Records of this stocking are not well documented.
Largemouth bass (M. salmonides) was introduced in the mid 1930's into the lowland reservoirs for angling purposes (Shorth-Smith, 1962). Bluegill sunfish (L. macrochirus) was introduced at the same time to serve as forage fish for the latter species.
Maqalika and Sebaboleng dams contain all the species that are known to be found in lowland reservoirs. The largemouth bass missing in fish stock assessment is occasionally caught with hook and line in Maqalika dam. However, Maqalika has never been stocked with any fish since its construction in 1983. Fish are assumed to escape from Sebaboleng dam which is immediately above it. Smallmouth yellowfish present in Maqalika but not reported in Sebaboleng are assumed to have come with the water regularly pumped from the Caledon River or as a result of upstream migration before the dam was built.
| SIZE CLASS | SPECIES COMPOSITION | FISH DENSITY | OBSERVATIONS |
| CLASS IA | Barbus aeneus | Medium | High density in Thaba-Phatsoa |
| Large reservoirs | Clarias gariepinus | low | |
| Cyprinus carpio | Medium to high | ||
| Low | Not found in Thaba-Phatsoa dam | ||
| Labeo capensis | High | ||
| Labeo umbratus | Low | ||
| Lepomis macrochirus | Low | ||
| CLASS IB | Micropterus salmonides | ||
| Large and silted reservoirs | Low | ||
| Clarias gariepinus | Low | ||
| Cyprinus carpio | |||
| CLASS IIA | Clarias gariepinus | Low | |
| Medium reservoirs | Cyprinus carpio | Low | High density in Nyenye dam |
| Lepomis macrochirus | Low | High density in Nyenye dam | |
| CLASS IIIA | Clarias gariepinus | Low | |
| Small reservoirs | Cyprinus carpio | Low | High density in Agri College Maseru dam |
| CLASS IV Ponds | Cyprinus carpio | Low | |
Most of the smaller lowlands reservoirs are built either in catchments that never had any recognizable stream or across minor rivers which are intermittent or ephemeral (Tilquin & Maseatile, 1994). Hence the fish fauna in classes II, III and IV are probably a result of stocking rather than natural occurrence.
Only three different fish species were observed in sampled reservoirs less than 10ha (classes IIA, IIIA, part of class IV). These are the common carp, sharptooth catfish and bluegill sunfish.
The class IB consisted of large and silted dams (e.g. Raleting dam, Tsa-Kholo dam) in which only common carp and catfish were found. These two latter species are likely to be the only fish which could be found in the small and very turbid reservoirs of the classes IIb, IIIb and IV.
Figures 6.7. and 6.4. show that numerous large individuals (above 300mm TL2) were caught for L. umbratus and B. aeneus, the two most abundant fish species respectively in Maqalika and Thaba-Phatsoa dams. Another important catch is observed in the length class 250–260mm TL and 200mm TL for L. umbratus and B. aeneus. Meanwhile no fingerling were caught from either species.
2 TL=total length, FL=fork length; SL=standard length.
Figure 6.7 Length frequency distribution for Labeo umbratus in Maqalika dam.
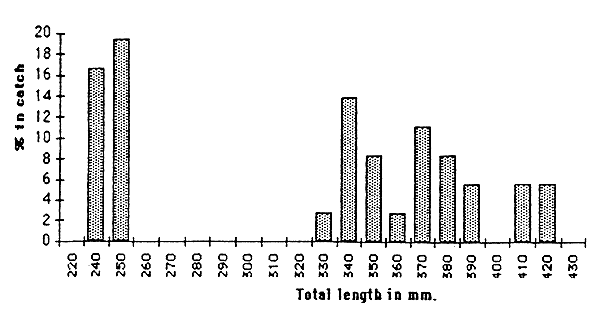
This erratic distribution should be confirmed through several fish stock evaluations. Breeding success may vary from one year to another for these two species. Meticulous catch monitoring, supported by a restocking programme, is probably needed to sustain gillnet fisheries in these reservoirs.
As mentioned earlier, lowland reservoirs have only seven recorded species. Why such a major river should have such poor fish fauna is not fully understood: the reasons probably lie partly in the geological history of the area and partly in the low winter temperatures which prevail over much of the system located in the highveld region of South Africa (Allanson & Jackon, 1983)
A second general point concerns breeding habits. Individual fish breed only once a year in Lesotho, generally when floods follow spring or summer rains. A spawning migration is often undertaken by adults to place the young initially in a favourable locality among inundated vegetation or gravel. This seasonal breeding habit has some extremely important implications for fishery management.
The fish growing season in Lesotho depends on water temperature but generally starts in October and lasts through April, roughly coinciding with surface temperatures above 20°C (Chondoma, 1988).
The paragraphs below briefly discuss important features of Barbus aeneus, Labeo umbratus and Labeo capensis, which represent the highest gillnet fishery potential in the Lesotho lowlands.
The smallmouth yellowfish (Barbus aeneus, Burchell, 1822):
Barbus aeneus, formerly called Barbus holubi (Steindachner, 1894) is the earliest correct name for this species (Skelton, 1993).
An exhaustive study carried out from 1978 to 1983 in the South African highveld Lake Leroux highlights the main patterns of the smallmouth yellowfish ecology (Allanson and Jackson, 1983). This fish has been shown to feed largely on invertebrates, including zooplankton, for the earlier part of its life history. Specimens of less than 6 cm occur near the shore and feed exclusively on bottom-dwelling insect larvae. At lengths above this the fish disperse and spend a greater proportion of time offshore and near the water surface, where it can take increasing quantities of zooplankton. Once they exceed 30 cm in length, the proportion of zooplankton (its major food up to this stage) in the diet decreases and is replaced by benthic algae and angiosperm material.
In the lake Leroux, smallmouth yellowfish does exploit zooplankton, but is seen selecting the largest individuals of the species present, virtually ignoring the smaller species which form the bulk of the zooplankton.
This fact of progression in dietary composition is not known for the other species found in lowland reservoirs. Since it was known to utilize zooplankton, the smallmouth yellowfish is expected to be an important component of the ecosystem in large lowland reservoirs, and to form an important link through which the planktonic production could be harvested.
On the other hand, because of the dependence of the smallmouth yellowfish on zooplankton, reduced availability of this food can led to eventual starvation and death of a large part of the population. Catastrophic mortality in 1978 and 1981 reported in the lake Leroux coincided with sharp increases of turbidity. This high suspensoid concentration reduced visibility, and the yellowfish became a less effective predator on the zooplankton community. This high turbidity probably also impairs the breeding success of this species which needs to migrate upstream to spawn over suitable gravel beds in clear running water.
Size at sexual maturity is variable and depends on growth rate. Population may stunt (i.e. ability to breed below the normal adult size) so that the size at sexual maturity is reduced. However, Skelton (1993) gives indicative figures of 200mm FL for males and 240mm FL for females. Fecundity increase from about 6,000 eggs at 300mm FL to about 60,000 eggs at 500mm FL.
Orange River labeo (Labeo capensis, A,Smith, 1841) and mud mullet or moggel (Labeo
umbratus, A. Smith, 1841):
The two Labeo species are bottom feeders, ingesting large quantity of algae and detritus. The
Orange River labeo (L. capensis) is adapted to scrapping algae off rocks while the Moggel (L.
umbratus) tends to be more associated with muddy bottoms.
The breeding habit of L. capensis is similar to that of Barbus aeneus, using shallow rocky rapid and gravel beds to breed. The sexual maturity size is about 220mm SL for males to 240mm FL for females (Skelton, 1993).
Orange River labeo are not subject to stunting. Starvation results in a failure to breed, while breeding itself depends on floods in tributaries and can fail completely during some years. These factors render this fish more vulnerable to meteorological events or overfishing.
Little is known about the biology and ecology of L. umbratus. It is recognized as a floodplain spawner with relatively high fecundity and a long life cycle. The fecundity and sexual maturity of L. umbratus differs from one locality to another. In Wuras dam situated in the South African Highveld, their fecundity is high (200.000 eggs at 400mm FL) and they are quite long when they attain sexual maturity (female, 360mm FL; male, 300 FL). Spawning apparently occurs in impoundments. Evidence also suggests that individuals may spawn more than once during a particular season (Gaigher, 1984).
Exercises undertaken in Maqalika dam with monofilament and multifilament gillnets show that the same number of species were caught with these two different materials, except for L. macrochirus. Multifilament material is more durable than monofilament material for fish stock assessment in Lesotho. However, monofilament gillnets were more efficient in the smaller mesh size. Multifilament nets should be complemented by a monofilament gillnet of stretched mesh size less than 50mm to assess small fish species or fingerling.
In reservoirs smaller than 10ha built across narrow and deep rivers or ravines, a depth over 3m is often reached. Shallow floating nets (1,5m deep) are advisable to assess the offshore area since anoxic zones are encountered below 3m or less where bottom nets are set.
The abundance of two fish species, common carp and sharptooth catfish, was probably underestimated. Due to their body shape, these species are difficult to sample with gillnet. Usage of seine nets is recommended to assess these two latter species where beaches with low slope and bottoms cleaned of submerged obstacles are identified (e.g. Nyenye dam). CPUE from seine netting is often not representative of the present fish composition of a dam since all different areas could not be properly assessed. Nevertheless it could give useful information on relative fish abundance, particularly on species difficult to catch with gillnets.
Bottom longlines, described in Chapter 7, should also be tested as an alternative way of sampling catfish which is the only predator of importance in this river system. This technique has already shown promising results in Zambia (Fjälling & Fürst, 1991, Mattson & Mutale, 1992).
Reservoirs smaller than 10ha were found to have a low fish density. The low catch recorded in some nets created large variations in the data analysis (see Section 6.4.3). More nets should be used per depth zone to reduce the variation. The size of the fishing effort required to reach a certain precision of CPUE is related to how fish behave and disperse in the water volume (Fjälling and Fürst, 1991). It will be possible to tune the methods in respect of the specific behaviour of Lesotho fish species.
Anoxic conditions that prevail below the thermocline as found in this study appear to be typical for reservoirs in the region (unpublished findings from Zambia, Botswana). An anoxic hypolimnion imposes a natural and much more definite zonation of fish abundance than predetermined artificial strata (Mattson & Mutale, 1992). In most of the reservoir, low oxygen content (3mg/1) is observed below 3m and even 1,5m depth. It seems therefore logical to treat the depth where oxygen becomes critical as the maximum depth, and set the corresponding number of nets only.
The comparison of catch between depth zones shows that catches in the 3–6m zone are much lower and the relative abundance of the major species does not differ from the upper theoretical strata (Figure 6.2.). This situation can be explained by the rapid oxygen depletion noted above. It could be therefore proposed to increase the number of repetitions in the 0–3m zone to accurate CPUE of the different species.
Some formula exists (Degerman et al., 1988) to estimate the number of nets to set according to the variation encountered in the catches. But when catches are low and greatly different from one net to another, the effort estimation (hundreds of nets) is not realistic compared to the dam size. For instance, in reservoirs smaller than 3ha, will not be possible to set more than seven nets without risk of entangling them.
In further investigation, the depth-to-volume curves for the reservoirs could also be estimated and the distribution of fishing effort by depth modified (Fjälling & Fürst, 1991).
The most suitable time for test fishing is when the variation in activity for the different species is expected to be at a minimum. This is to ensure that slight changes in time will not affect results. In Lesotho lowlands, this means avoiding the beginning of the rainy season. The best test fishing period should be from January to April when the breeding period (catfish, carp) including upstream migration for some species (Orange River labeo, smallmouth yellow fish, mud mullet) is completed. The fish population should be stabilized in the reservoir during this period before the coldest period (May, June, July) when they considerably reduce their biological activity.
The distinct catchability between species is a strong bias in the result interpretation. Selectivity also varies with age, condition of fish, behaviour and size of fish. For instance, in fishes possessing spines, serrations or prominent teeth, large head, this selectivity is less marked since such fish are often tangled. In Lesotho lowlands reservoirs this usually occurs with catfish and carp. However, smallmooth yellowfish and Labeo spp., which formed the bulk of the catch and represented the greatest potential, are relatively less spiny. Being fusiform in shape, they are most often caught by their gills. Selectivity curves for these are therefore likely to be fairly accurate and are provided on Figure 6.8. for nets of 35, 45, 57, 73, 93, 118, 150 mm strechted mesh for Labeo capensis and Barbus aeneus. Selectivity curves for Labeo umbratus for 93mm mesh size are also presented in Figure 6.9.
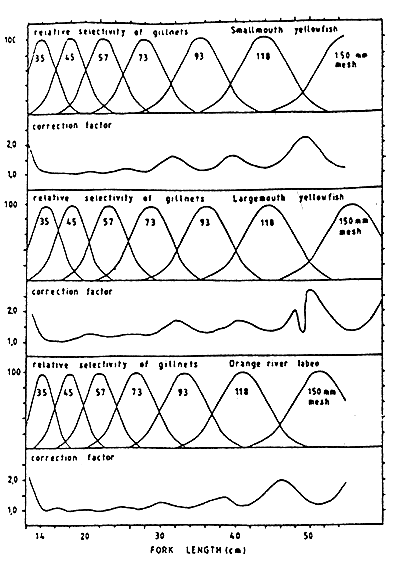
Selectivity curves show that the type of gillnet used is very selective. This property should be used to rapidly tune fisheries catches to the profile monitored of fish population.
Stretched mesh size of 93mm and above will not catch sub-adults (individuals inferior to 300mm FL). Retricting gillnet fishery to the upper mesh size limit of 120mm will leave some large individuals which are the favourite targets' of anglers'. This will depend on the management strategies people choose.
The “lake type” shape from the largest lowlands reservoirs offers few shelters for sub adults of Labeo and Barbus spp. to escape from an active gillnet fishery. Only a sustained restocking programme for those species could permit a reduction of the lower mesh size limit to 57mm stretched mesh size.
The results are biased by not being compensated for selectivity in the present study. However, it is also conceivable that the logarithmic mesh series of the nets used are mildly selective to fish size and that only minor corrections would have resulted from selectivity compensation (Mattson & Mutale, 1992).
The capturing efficiency of gillnets tends to decline with increasing catch. This effect has been demonstrated for a small fish species Barbus paludinosus in Botswana (Nermark and Mmopelwa, 1994) and Zambian reservoirs (Fjälling and Fürst, 1991; Mattson & Mutale, 1992).
Probable saturation effects also occur in high catches of L. umbratus or B. aeneus in lowland reservoirs.
Use of gillnets composed of panels of different mesh sizes or monomesh gillnet tied as a gang --Van der Mheen (1993) highlights the following complications. In this situation larger fish may “lead” along the small mesh size units until they come to and are captured by, a larger mesh unit. This will result in a biased sample with more large fish.
High CPUE recorded in the three larger dams, Maqalika, Sebaboleng and Thaba-Phatsoa, indicate that the present fishing pressure is low. The current use of hook and line alone is not enough to fish these dams to their maximum fisheries potential.
As already stressed above, zero catches were often observed in nets for species like largemouth bass (Thaba-Phatsoa dam), Orange River labeo (Maqalika dam) in large reservoirs or in reservoir under 10ha where small stocks were observed. High variance therefore follows.
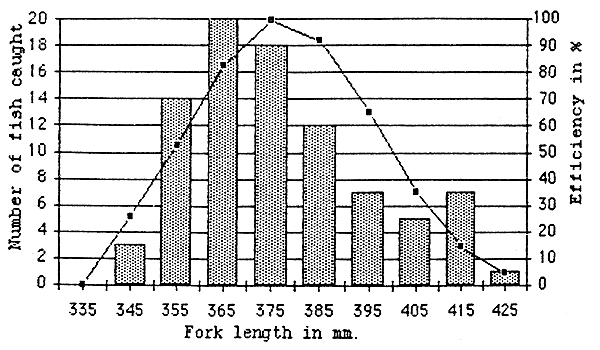
We only mention arithmetic calculations of CPUE in the present report. The priorities were to identity suitable equipment to sample lowlands reservoirs and propose a test fishing method adapted to the local situation. A further step would be to use a statistical method which could give a true statistical estimation of CPUE and related parameters (standard deviation, confidence intervals....). The In (CPUE+1) transformation which makes the variance independent of the mean CPUE is advocated by Degerman et al.(1988). Nevertheless this transformation has major impacts on the mean, especially in extreme cases where fish are caught in some of the nets while others are empty (Figure 6.10.). This effect has been also mentioned by Mattson & Mutale, 1992.
The present work in the light of ALCOM activities in Southern Africa could lead to a proposal for a preliminary standard method which could be made accurate by repeated exercises in selected reservoirs.
Several improvements from the Drottningholm method have been suggested for adaptation to the Lesotho situation. Firstly, two alterations of the test fishing gear are proposed. Multifilament materials are strong enough to catch the large species found in the Lowlands. They require much less time for net-mending than nets of monofilament gillnet. Further, the number of panels is reduced from 14 to 7; meanwhile the panel length is doubled.
Sampling repetitions in the 0–3m metre zone should also be increased to mildly reduce catch deviation. Lastly, the maximum depth should be considered as the depth where oxygen becomes critical (below 3mg/l). Only the corresponding number of nets should be set.
Henderson (1980) concluded that to assess the potential for development of a fishery, an order-of-magnitude estimate may be sufficient, until the fishery actually develops. In most cases, very crude sampling would be sufficient. When a fishery activity is present, the fishery itself may be used to refine the estimate.
The standardized test fishing method was seen to have some advantage; it provides fish in relative abundance, and the method can be repeated and compared between exercises for the same dam if conditions are the same.
![]()
![]()
![]()