![]()
![]()
![]()
Par Mr. O. RAILLAARD
et Mr. A. MENESGUEN
France
ABSTRACT
Oyster production of the Marennes-Oléron Bassin is simulated using a model integrating phytoplankton growth, oyster growth and horizontal transport of suspended matter. Firstly, simulations are compared to the measurements. At season scale, the phytoplankton and nitrates evolution are well predicted by the model. In compensation, the model does not simulate well the spring/neap tidal variability of the phytoplankton.
Secondly, the phytoplankton and oyster production are calculated as a function of the nitrates river inputs. Biotic capacity of the basin appears to be particularly influenced by these nutrients during summer. On the other hand, spatial variability of the impact of the nitrates variation is clearly shown. The primary productions of the north and north-west areas are poorly related to the nutrients from the river. In the north-east of the basin the high phytoplanktonic activity and the river proximity maintain to a quiet high level the sensivity production. In the oyster production area, the responses of the primary and oyster productions to nitrate variation are non-linear: the sensivity is more pronounced when decreasing nitrates than increasing. The trophics levels of the center part and the east part of this area are the most river-dependent.
The growth of the total primary and oyster productions predicted by the model for an increasing nitrate inputs, point put a limited fertilization of the bassin by the river. However, the uncertainty of the present model call into question this results quality.
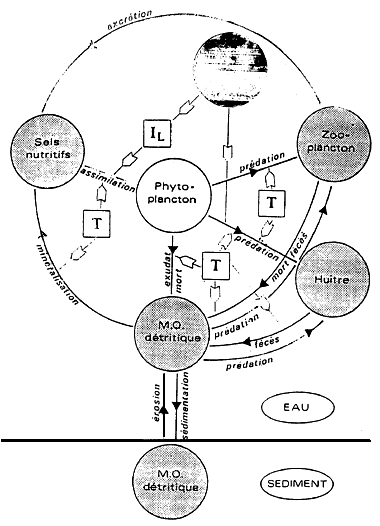
Figure 1 - Schéma conceptuel des transferts de matière et d'énergie au sein de l'écosystème conchylicole
IL : Intensité lumineuse disponible dans la colonne d'eau; T :Température de l'eau
INTRODUCTION
Depuis son implantation dans le Bassin de Marennes-Oléron (1970), l'huître C. gigas connaît une diminution graduelle de sa vitesse de croissance et un abaissement significatif de son taux de son taux de survie (Héral, 1986), entraînant ainsi une stagnation progressive de la production ostréicole. Cet affaiblissement général de l'état physiologique de l'animal est principalement du à l'augmentation croissante des stocks en élevage qui a abaissé les capacités trophiques du Bassin pendant cette période. L'objectif, à long terme, est de redonner à C. gigas des vitesses de croissance acceptables en gérant plus efficacement la production du bassin conchylicole. De par ses propriétés prédictives et explicatives, le modèle mathématique a semblé l'outil le mieux adapté à la réalisation de ce projet. Les travaux de Bacher (1989) ont ainsi abouti à la mise en place d'un premier modèle qui a permis d'étudier l'impact du stock d'huître et de sa répartition sur les performances de croissance de C. gigas.
La présente étude s'inscrit dans le cadre d'un aménagement de la production ostréicole du Bassin qui se concrétiserait par une régulation des débits de la Charente (mise en place d'un barrage de soutien d'étiage, contrôle de l'utilisation urbaine et agricole de l'eau du fleuve). De par ses apports en nutriments le fleuve Charente favorise la croissance phytoplanctonique au nord du Bassin conchylicole (Ravail et al., 1988). Les huîtres, consommatrices de phytoplancton et positionnées au sud de l'estuaire, sont donc suceptibles, compte tenu de la résiduelle nors-sud des courants de marée dans ce secteur du Bassin, d'être aussi affectées dans leur croissance par les variations de débit du fleuve.
Le présent modèle intègre la croissance phytoplanctonique, la croissance des huîtres et le transport particulaire. Dans un premier temps, les simulations obtenues dans des conditions nominales sont confrontées aux mesures effectuées en différents points du Bassin. On se propose ensuite d'étudier la sensibilité de la production primaire et ostréicole aux apports de nitrates du fleuve Charente.
I- MATÉRIEL ET MÉTHODE
A- Le modèle biologique
1- LES VARIABLES D'ÉTAT ET FORÇANTES (Fig. 1)
L'azote se révélant être, sous sa forme minérale dissoute, l'élément limitant de la production primaire (Héral et al., 1981), et sous sa forme organique particulaire (protéine) une source nutritive importante des mollusques (Héral et al., 1980) le présent modèle simulera le cycle de l'azote dans la masse d'eau et le sédiment. Les composantes pélagiques de la chaînte alimentaire sont: l'azote minéral, le phytoplancton, le zooplancton et l'azote organique détritique. Pour la composante benthique, ou distingue les huîtres, représentées par deux classe d'âge (1 an et 2 ans), la matière organique détritique. La forte variabilité spatiale et temporelle de la turbidité enregistrée à Marennes-Oléron et l'action déterminate de ce paramètre sur la poussée phytoplanctonique (régulation de l'énergie lumineuse dans la colonne d'eau) et sur le comportement nutritionnel et de croissance de C. gigas, justifient l'adjonction d'une variable d'état ((seston minéral)), qui constitue en effet le principal élément explicatif de cette turbidité. La température et le flux lumineux journalier moyen sont les deux variables forçantes du modèle.
2- LES LOIS DU MODÈLE
Hormis l'introduction du compartiment huître le modèle biologique est celui mis au point par Menesguen (1989).
L'effet de la température sur la vitesse des processus est considérée comme exponentielle sur la plage habituelle des températures de la mer à Marennes. La croissance du phytoplancton est déterminée par une loi de Michaelis-Menten pour l'effet des sels nutritifs et de Steele pour celui de la lumière dont le coefficient d'atténuation est une fonction linéaire de la concentration de seston minéral (Cloern, 1987; Chapelle, com. pers); sa mortalité naturelle est asservie au degré de limitation de croissance rencontrée par le phytoplancton. Le niveau d'activité nutritionnelle du zooplancton (broutage) est régulé par la concentration d'azote particulaire pélagique végétal d'après une lois d'Ivlev. L'efficacité d'assimilation (pourcentage assimilé de la nourriture consommée) est une fonction exponentielle inverse de la ration, le taux d'excrétion est calculé en prenant la valeur maximale entre une fraction constante de la nourriture assimilée et le taux d'excrétion basal croissant exponentiellement avec la température.
La prédation des mollusques sur le phytoplancton et la matière détritique est une fonction allométrique du poids, et régulée d'après une loi exponentielle décroissante de la charge sestonique pour des concentrations supérieures à un seuil donné, et ce, afin de simuler la baisse de l'activité de filtration provoquée par le colmatage des branchies (Widdows et al., 1979). Le taux de production des pseudo-fécès est une fonction exponentielle croissante de la consommation particulaire (inspirée de Bayne et al., 1976). L'ingestion de matière organique (azote particulaire) et minéral est calculée en refranchant les quantités émises sous formes de pseudo-fécès aux quantités consommées. L'efficacité d'assimilation est une fonction linéaire croissante de la température. L'azote assimilé par le mollusque est converti en calorie avec un coefficient supérieur pour le compartiment vivant (phytoplancton). L'énergie consacrée à la croissance somatique et gonadique est obtenue après retrait des dépenses métaboliques (respiration) de l'énergie ainsi assimilée. La mortalité des populations des deux classes d'âge (1 et 2 ans) est simulée par une loi du premier ordre (Bacher, 1989). L'effort de ponte est une fonction allométrique du poids (Deslous-Paoli et Héral, 1984). La date et la durée de la ponte ne sont pas simulées mais fixées d'après des mesures effectuées dans le milieu.
Une loi du premier ordre gouverne la reminéralisation de la matière organique. Ce modèle biologique est une représentation très simplifiée des relations trophiques au sein de l'écosystème conchylicole. Toutefois, comme le précise Jorgensen (1988), les choix de structure concernant les modèles d'écosystème doivent avant tout être déterminés par le niveau des connaissances acquises sur l'écosystème et par les objectifs de la modélisation. La formalisation actuelle nous paraît répondre de manière satisfaisante à ces deux critères. Les modèles complexes sont des outils lourds, coûteux en temps calcul. De plus, si le fait d'accroître l'incertitude de ses résultats (Jorgensen, 1988). Un modèle simple, à même de montrer les tendances générales, est, pour cette raison, préférable. Les équations du modèle biologique sont présentés à l'annexe 1.
B- Le modèle de transport hydrodynamique
La partition géographique du Bassin de Marennes-Oléron est imposée par la variabilité spatiale des différentes variables simulées. Le transport des éléments particulaires et dissous est assuré par un modèle physique d'advection-dispersion dont les paramètres sont calculés d'après le modèle courantologique bidimensionnel à bancs découvrants du L.C.H.F. (Anon, 1979 b).
La faible profondeur du Bassin, associée au brassage intense de la colonne d'eau par les courants de marée et le vent, permet de supposer une répartition homogène des composantes particulaires et dissoutes de la masse d'eau selon l'axe vertical.
Les échanges avec de fond ne sont pas, à proprement parlé, simulés. La sédimentation est supposée constante et s'applique aux particules détritiques de la masse d'eau. La remise en suspension est exprimée d'après une fonction sinusoïdale, propre à reproduire l'engraissement du sédiment en été et son érosion en hiver.
1- EMPRISE DU MODÈLES, DISCRÉTISATION SPATIALE ET TEMPORELLE
L'emprise géographique du modèle doit être telle que l'on puisse supposer que les mécanismes simulés n'influencent pas les conditions aux limites. Le modèle est ainsi bordé, par une frontière Nord qui reçoit des apports de type océanique (pertuis d'Antioche), par une frontière Est où se déverse la Charente, et une frontière au sud du bassin qui aliment le modèle par des apports de nature océanique identiques à ceux de la limite Nord (fig. 2).
La simulation des processus biologiques considérés dans le ces présent ne nécessite pas, eu égard aux incertitudes quant à leur formulation, la précision apportée par les modèles hydrodynamiques (Bird et Hall, 1988). En conséquence, le modèle courantologique du L.C.H.F. est moyenné dans l'espace et le temps (modèle en boîtes) et ce, en accord avec les échelles de temps et d'espace des mécanismes biologiques. Les compartiments vivants de la chaîne alimentaire pélagique ainsi que les mollusques sont supposés intégrer les fluctuations dont la période est inférieure à celle de la marée (12h30). Cette échelle de temps est celle adopté par d'autres auteurs pour des modèles du même type associant physique et biolgie (Baretta et Ruardij, 1988; Klepper et Scholten, 1988).
Plusieurs critères décident de la partition géographique de la zone:
d'ordre courantologique, le tracé des boîtes aura soin de délimiter des zones hydrodynamiquement homogènes (en vitesse et en direction), défines approximativement à l'aide des grilles de courants résiduels eulériens du modèle physique du L.C.H.F. La taille de ces boîtes est de l'ordre de celle de l'excursion de marée responsable de l'homogenéisation des masses d'eau (quelques Km) (Bacher, 1989);
d'ordre biologique, l'homogénéité est de ce point de vue plus difficile à cerner, étant donné le nombre des variables à considérer. Les boîtes Sud doivent tenir compte des zones ostréicoles; l'estuaire de la Charente, qui présente d'importants gradients de salinité et de turbidité (Ravail et al., 1987), est scindé en deux boîtes (Fig. 2).
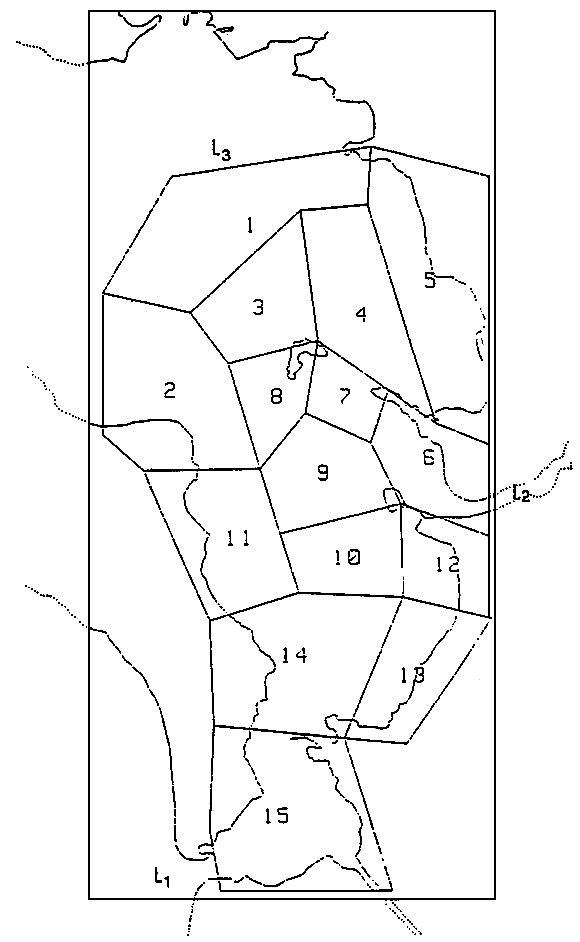
Figure 2 - Partition géographique du bassin de Marennes-Oléron
L1 : Limite océanique sud
L2 : Limite Charente
L3 : Limite océanique nord
2 - ÉQUATION D'ADVECTION-DISPERSION
En absence de processus d'ordre biologique. l'évolution de la concentration d'une variable pélagique dans la boîte i est calculés un schéma décentré-amont.
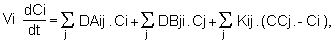
Vi est le volume de la boîte i, A désigne la quantité de la substance quitant la boîte i sous la condition que Dij soit négatif, et B la quantité entrant dans cette même boîte si Dij est positif. Le dernier terme C de l'équation traduit les échanges dispersifs dont le sens est fonction du gradient de concentration entre les boîtes adjacentes.
C - Conditions initiales et aux frontières du modèle
L'importance du forçage courantologique rend le modèle peu sensible aux valeurs de départ des variables pélagiques qui sont, pour cette raison, fixées approximativement d'après les mesures.
Le nombre d'huîtres par boîte et classe d'âge est estimé à partir des échantillonnages annuels réalisés pour deux classes de poids et par strate géographique, sur l'ensemble du Bassin, depuis 1984 (Bacher et al., 1986; Bodoy et al.,1987). Les poids de départ des individus de la première et de la seconde classe d'âge sont issus de valeurs moyennes mesurées sur le terrain. Les stocks de matière détritique sur le fond sont inconnus; en première approximation, leur niveau est pris égal à O dans toutes les boîtes.
Les scénarii aux limites géographiques du modèle concernent l'ensemble des variables pélagiques. Afin de gommer les perturbations météorologiques du signal, une année hydrobiologique-type est reconstituée à l'aide des mesures effectuées pendant plusieurs années dans deux stations du Bassin, celle de Boyard et de l'estuaire de la Charente (voir Fig.3).
Cependant, la constitution des scenarii à la limite océanique pose problème. II n'existe pas de mesures hydro-biologiques qui reflètent véritablement les condition du large. En raison de sa proximité avec la Charente, la station Boyard (la plus proche de l'Océan) reste influencée par les apports de ce fleuve. Les teneurs des diffèrentes variables hydro-biologiques mesurées en ce point ne sont donc pas en mesure d'alimenter le modèle dans son emprise géographique actuelle. Les pseudo-limites océaniques sont obtenues de la manière suivante : sous l'hypothèse que le Bassin soit un facteur d'enrichissement des eaux océaniques (pour les variables concernées par le modèle biologique), on admet à l'inverse que l'évolution temporelle des concentrations au large peut être représentée par une certaine « dilution» des concentrations mesurées à Boyard. La valeur du facteur de dilution est déterminée en calant les courbes simulées, par le modèle de production primaire, de l'azote minéral et du phytoplancton dans la boîte 8, aux données de Boyard (Fig.4).
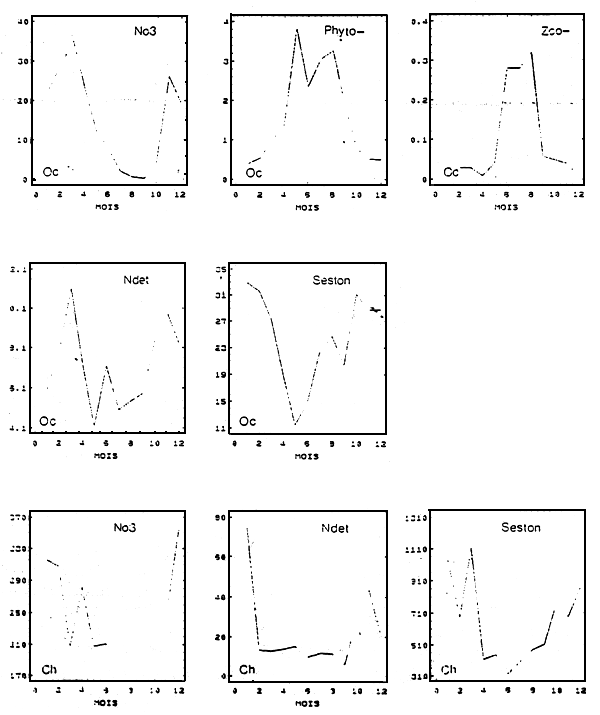
Figure 3 - Cycles annuels des conditions hydrobiologiques aux limites océaniques (Oc) et à la limite Charente (Ch). Les concentrations sont exprimées en μMN/l, à l'exception de celles du seston minéral qui sont exprimées en mgPs/l
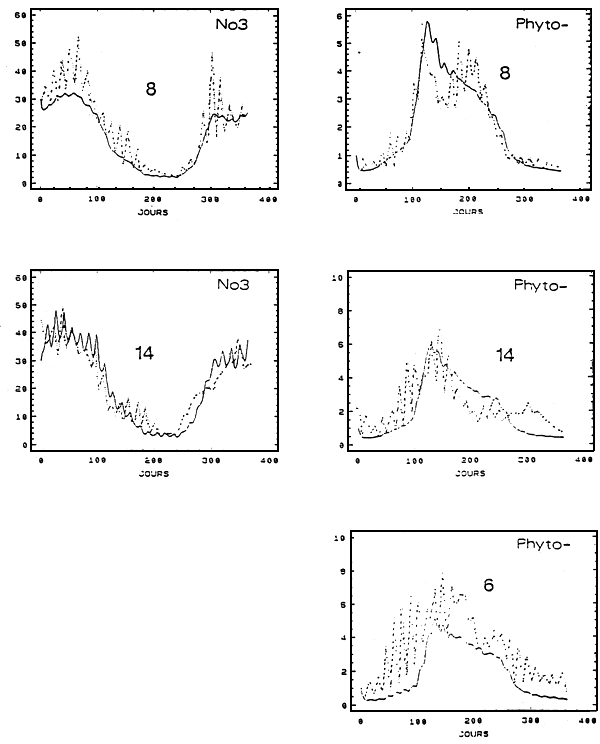
Figure 4 - Valeurs calculées (traits pleins) et mesurées (traits pointillés) des concentrations de nitrates (μM/l) et le phytoplancton
(μMN/l) sur une période d'un an.
8 : résultats du calage des simulations aux mesures à Boyard pour l'obtention des conditions aux limites
14 et 6 : comparaisons calculs/mesures au Chapus (14) et dans l'estuaire de la Charente (6)
II - RÉSULTATS
A - Simulation nominale
Les simulations du nitrate et du phytoplanction sont comparée à leurs valeurs mesurées au Chapus et dans l'estuaire de la Charente (phytoplancton uniquement pour cette dernière station). Ces confrontations entre les mesures et les calculs constituent un premier test de la qualité des résultats du modèle mais ne peuvent être utilisées pour juger strictement de sa validité. Si la valeur des paramètres des modèles d'azote et de croissance des huîtres a bien été déterminée en dehors du contexte actuel, le calcul préalable des séries temporelles de phytoplancton et de nitrate à la limite marine introduit un biais dans la procédure de validation.
A l'échelle de la saison, le calcul de l'évolution temporelle des nitrates de la boîte 14 s'ajuste aux données du Chapus (Fig.4). A l'instar de ce qui est observé, le modère également des oscillations à plus hautes fréquences (celle du cycle de marée), induites par les différences de mélange en vives-eaux et en mortes-eaux entre les eaux de Charente et océanique.
La chlorophylle prédite en boîte 6 (Charente) présente des valeurs systématiquement plus basses que ce qui est mesuré dans l'estuaire (Fig.4). On peut supposer qu'au niveau de l'estuaire de la Charente, les apports de phytoplancton par la rivière, ici négligés, et le développement d'espèces locales, adaptées aux fortes turbidités, expliquent cette sous-estimation des teneurs chlorophylliennes par le modèle. D'autre part, il est possible que le modèle surestime en moyenne les turbidités printanières et estivales, en réalité régulées, pour une bonne part, par les échanges verticaux en ce point du Bassin (Ravail et al., 1987). L'absence des fluctuations hebdomadaires provient de cette sous-estimation des concentrations(absence de contraste entre les eaux marines et de l'estuaire) et de l'absence de simulation des échanges avec le fond auxquels sont également soumises les cellules phytoplanctoniques (Ravail et al., 1987).
Le phytoplancton calculé boîte 14 retrouve un niveau semblable à celui mesuré au Chapus. Malgré un léger décalage en début d'année, les pics printaniers son synchrones. Les mois suivants les courbes simulées et estimées se croisent sans présenter de différences importantes. C'est avant tout, et une fois encore, sur les oscillations tidales que le modèle est en désaccord profond avec la nature.
B - Sensibilité de la production primaire et ostréicole aux apports de nitrates du fleuve Charente
1 - PRINCIPE
Pour une simulation donnée, le scénario à la limite Charente correspondant est obtenu en multipliant les concentrations de la situation nominale par un facteur constant (fN03).
Sur ce schéma, on procède suivant deux étapes. Dans un premier temps, les résultats concernant les modifications induites par ces perturbations sur l'évolution temporelle de la chlorophylle et du nitrate sont examinées dans une boîte de la zone ostréicole pour trois valeurs du facteur fNo3 (0.25, l et 2), ensuite la dimension spatiale du problème est abordée à l'aide de 10 simulations. Les valeurs prises par fNo3 sont alors, 0.05, 0.15, 0.25, 0.5, 0.75, 1, 1.25, 1.5, 2 et 3. La production phytoplanctonique annuelle, le poids maximal des huîtres des deux classes d'âge et la production ostréicole totale sont les paramètres pour chaque cas de figure.
2- ÉVOLUTION TEMPORELLE DU NITRATE ET DU PHYTOPLANCTON (FIG.5)
En raison de sa situation géographique (au milieu de la zone de culture), c'est dans la boîte 14 que sont examinées les différentes simulations des nitrates et du phytoplancton.
Quel qu, en soit le sens, la variation des niveaux de nitrates à limite Charente du modèle n'affecte guère le comportement printanier du phytoplancton: pour chacune des valeurs de fNo3 les pies calculés de chlorophylle restent synchrones et leurs valeurs (pour fNO3 croissant) évoluent autour de 6mgChla/m3 (Fig. 5). Il apparaît ainsi qu'à cette époque de l'année les apports d'Antoche (limite nord du modèle), marqués par la présence des eaux de Gironde (Dechamboy, 1977), maintiennent des niveaux de nitrates au-delà du seuil de limitation du développement phytoplanctonique. A la fin du printemps, la situation change, les eaux du large retrouvent des caractéristiques plus océaniques et rendent à la Charente et à ses apports de nitrate une partie du contrôle de la production primaire. Ces résultats sont en accord avec ceux obtenus par la biais de la mesure et de l'expérimentation in situ (Ravail et al., 1988). Sans que cet effet soit spectaculaire sur un plan instantané (l'écart maximum est < 1 mgChla/m3 pour les valeurs extrêmes de fNo3), il se prolonge néanmoins pendant les 3 mois d'été et peut, à ce titre, influer sur les capacités trophiques du Bassin et, par voie de conséquence, sur les performances de croissance des huîtres.
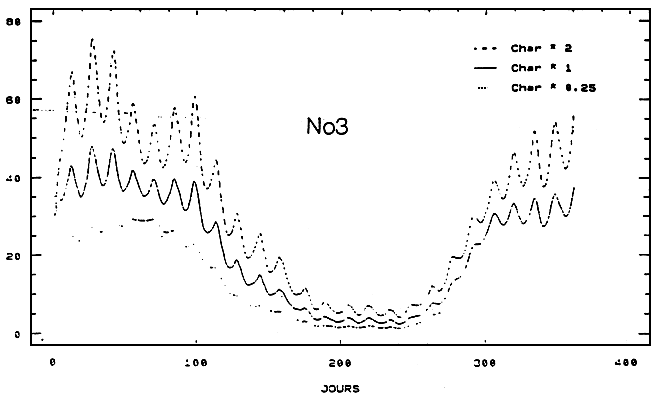
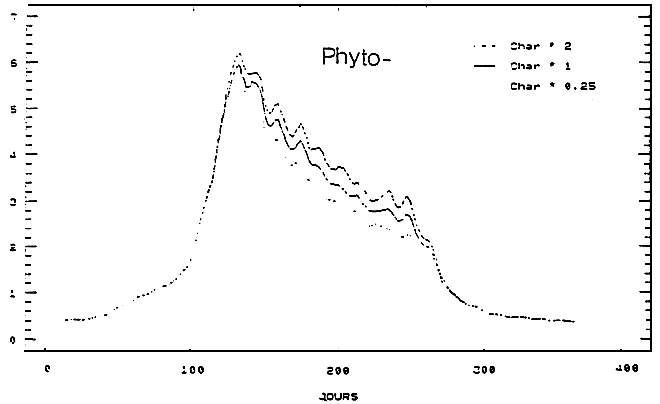
Figure 5 - Influences des apports de nitrates du fleuve Charente sur l'évolution temporelle des teneurs de nitrates et de chlorophylles au Chapus (boîte 14)
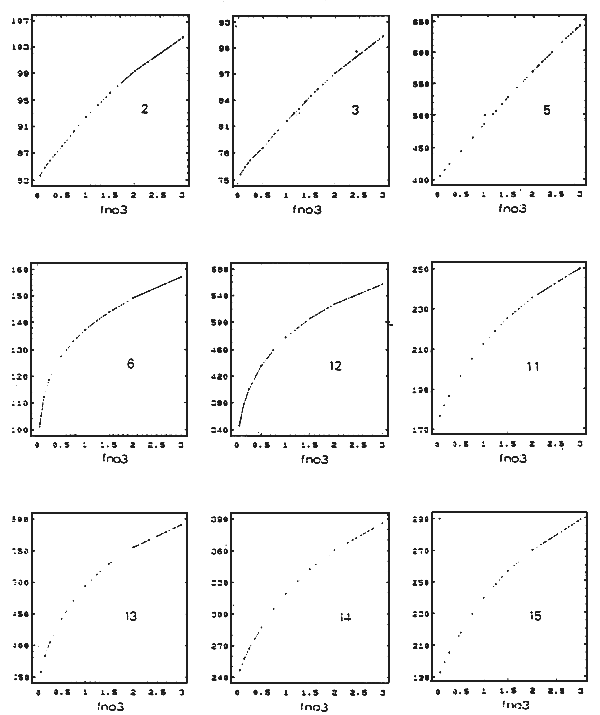
Figure 6 - Influence des apports de nitrates du fleuve Charente sur la production primaire brute annuelle (uM/l/an) de
différents secteurs du bassin
2, 3: nord-ouest; 5: nord-est, 6: estuaire: 12, 13: estran est; 14: central; 15: sud
FNO3: facteur multiplicatif du scénario de nitrates de la limite Charente
3 - ÉVOLUTION SPATIALE DE LA PRODUCTION PRIMAIRE ET OSTRÉCOLE
(Fig. 6)
La production primaire d'un secteur donné est d'autant plus marquée par les variations du régime de la Charente que ses concentrations de nitrate y sont sensibles, et qu'elles sont également proches du seuil de limitation de la croissance du phytoplankton. La région nord-ouest (boîtes 2 et 3) d'influence marine affiche une sensibilité faible (tableau 1), les concentrations de nitrates sont peu affectées par le changement des conditions à la limite est, et constante sur la gamme de variation du facteur fNo3, en raison du maintien de ces concentrations à un niveau limitant (pas de saturation). Dans la boîte nord adjacente (5), la marque des apports estuariens est plus prononcée et entraîne une réaction plus vive de la croissance du phytoplancton à leur variation, le transit relativement lent des masses d'eau sur ce secteur intensifie le pompage des nutriments et maintient la limitation malgré l'accroissement des rejets de Charente (forme linéaire de la réponse), au niveau de l'estuaire et dans la zone de production ostréicole, la non-linéarité des réponses met en évidence, d'une part, un processus de saturation de la production primaire par rapport à l'accroissement des flux de nitrate charentais pour les forte valeurs de fno3, et, d'autre part, une sensibilité de la production qui s'accroît lors de l'abaissement des rejets de Charente. La dépendance plus marquée de la boîte 13 et dans une moindre mesure de la boîte 14 met à jour le conditionnement particulier de ces secteurs par les régimes du fleuve.
Tableau 1 - Variation de la production ostréicole en fonction des apports de nitrates
| Boîte fNO3 | 3 % | 5 % | 6 % | 10 % | 11 % | 12 % | 13 % | 14 % | 15 % | |
| Phyto | 0,25 | -5,5 | -12,7 | -13 | -14,5 | -12,2 | -16,14 | -18 | -16,2 | -14 |
| 0,50 | -3,7 | -8,7 | -7,1 | -8,8 | -7,6 | -8,9 | -10,5 | -9,9 | -9,31 | |
| 1,50 | -3,5 | -8,5 | -4,8 | -6,4 | 15,9 | +5,8 | +7 | +7,2 | +7,36 | |
| 2 | -6,6 | -16,7 | -8,7 | -11,6 | 10,7 | +10,4 | +12 | +18 | +15,7 | |
| Huitre | 0,5 | -7,5 | -3,2 | -13,2 | -7,3 | -4,3 | ||||
| 1,5 | +5,7 | +2,5 | +9,7 | +5,5 | -3,3 |
Le comportement de croissance des mollusques est logiquement corrélé à celui du phytoplancton. Tant sur le plan des poids maximaux (Fig.7) que de la production totale (Fig. 8), la diminution des apports de Charente a un effet plus prononcé que son augmentation. La comparaison des résultats obtenus sur les différentes boîtes révèle également de relations plus étroites entre les apports de nitrate charentais et la croissance des mollusques sur l'estran est (boîte 13) et au centre du Bassin (boîte 14) (tableau 1) que dans les secteurs périphériques (boîtes 11 et 15). Un résultat qui revêt une importance particulière pour la boîte 14 qui est un secteur d'élevage relativement plus dense. Un accroissement de 50 % de ces apports estivaux (valeur prédite après la mise en place du barrage) entraînerait une augmentation de la production ostréicole entre 3 et 10 % de sa valeur nominale selon les secteurs considérés. Dans une situation de sécheresse (division des apports estivaux nominaux par 4), cette production subirait une baisse comprise entre 6 et 18 % de sa valeur actuelle.
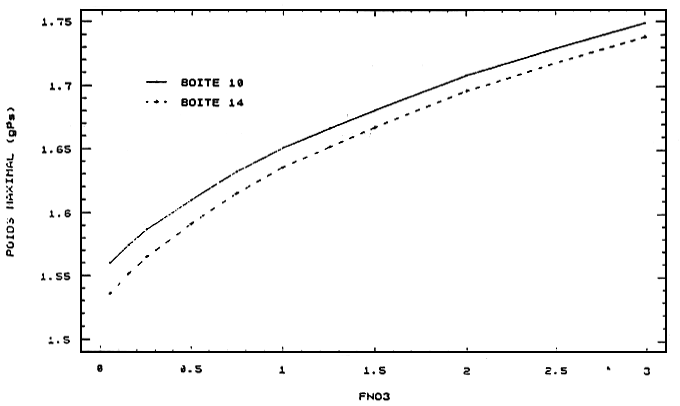
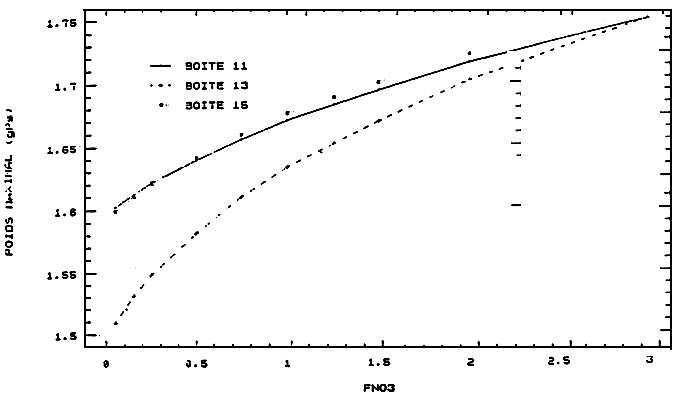
Figure 7 - Réponses de croissance des huîtres de la seconde classe d'âge aux variations des apports de nitrates du fleuve
Charente
FNO3: facteur multiplicatif du scénario de nitrates de la limite Charente
10, 11: nord; 13: estran est; 14: centre; 15: sud
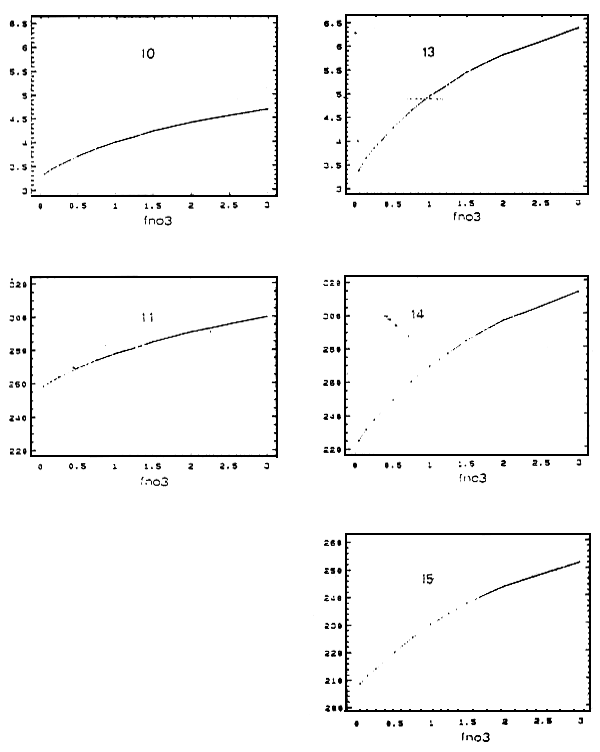
Figure 8 - Réponses de la production ostréicole annuelle (tonnes de poids sec) aux variations des apports de nitrates du
fleuve Charente
FNO3 : facteur multiplicatif du scénario de nitrates de la limite Charente
10, 11 : nord; 13 : estran est; 14 : cetre; 15 : sud
III - DISCUSSION
La saturation observée sur les différentes courbes de réponses de croissance des huîtres à l'enrichissement minime de la production ostréicole prédit pour un doublement des apports en nitrates de la Charente, semblent indiquer que le fleuve fertilise peu le bassin. Il paraît en effet illusoire de penser, au vu de ces résultats, que l'huître puisse retrouver une vigueur de croissance équivalente à celle des années passées en accroissant les débits de la Charente. On peut supposer, à l'instar d'Héral (1989), que les fortes turbidités et les faibles temps de résidence du bassin ne permettent pas l'utilisation efficace par le phytoplancton du surplus de nitrates en provenance de Charente, il existe un autre considérations d'ordre physique et biologique, dont la justesse n'est plus à discuter, il existe un autre responsable majeur de la censure des processus biologique. Une étude récente (Lazure, com. pers.), a permis de mettre en évidence que l'application des schémas classiques de calcul employés pour résoudre le transport particules. Replacé dans le contexte ?en boîtes? génère une accélération artificielle du transit des particules. Replacé dans le contexte actuel, ce biais induit une régulation accrue de la production primaire par la vitesse e déplacement des masses d'eau et corrélativement une sous-estimation du pompage des sels nutritifs par le phytoplancton.
Les simulations obtenues dans des conditions nominales permettent de créditer le modèle d'une prédiction relativement correcte de l'évolution moyenne des teneurs en nitrates et en phytoplancton au niveau du bassin conchylicole. Il ne faudrait pas toutefois en déduire que l'on ait compris (même à cette échelle de temps) le processus de la croissance phytoplanctonique à Marennes. Il est ainsi probable que l'adéquation sur les niveaux de nitrates et de chlorophylle au Chapus soit en partie due au calage préalable du modèle sur les données de Boyard (voir Il), les stations Chapus et Boyard variant peu de ce point de Vue.
Quant aux écarts observés entre le calcul et la mesure des concentrations chlorophylliennes à l'échelle du cycle vives-eaux/ mortes-eaux de marée, ils proviendraient en partie de l'absence de simulation des échanges particulaires entre l'eau et le sédiment. Les variations de concentration à l'échelle du cycle de marée, initiées par les mélanges variable des eaux marine et estuarienne, sont accuentuées par les échanges verticaux, avec une tendance au dépôt en mortes-eaux et à l'érosion en vives-eaux. Cette amortissement des oscillations vives-eaux/ mortes-eaux reproduit par le modèle est de nature à fausser la forme effective de l'interaction entre les nitrates et le phytoplancton.
Il semble donc que les résultats concernant l'impact des rejets de nitrates du fleuve sur la production primaire et ostréicole n'aient qu'une relative valeur prédictive et ne soient pas transposable à la réalité du terrain.
CONCLUSIONS
En résumé, la modélisation de l'écosystème de Marennes-Oléron a permis de mettre en évidence :
un impact variable des rejets en nitrate du fleuve Charente sur la production primaire et ostréicole d'après la période et la région considérée;
une sensibilité plus marquée de la production ostréicole à la baisse qu'à la hausse des flux de nitrate charentais;
un contrôle important de la production primaire par l'hydrodynamisme.
Ces trois points, qui soulignent l'importance que revêt l'aspect dynamique dans l'étude du phénomène de production primaire, justifient pleinement la démarche modélisatrice adoptée pour le site de Marennes-Oléron. De plus, si le présent modèle n'a pu fournir que des tendances (et non pas des prévisions applicables au terrain) quant à l'incidence biotique de la Charente, il n'en demeure pas moins un outil de recherche qui a participé pleinement à l'élargissement de notre connaissance de l'écosystème conchylicole de Marennes-Oléron. Les auteurs engagés dans la modélisation d'éco-Système s'accordent sur la primauté d'une telle valorisation de l'outil (Keizer et al., 1987; Baretta et Ruardij, 1988; Klepper et Scholten, 1988), Il est en effet généralement admis que la complexité des systèmes abordés en écologie et l'état actuel de nos connaissances remettent en cause les capacités prédictives de ces modèles (Jorgensen, 1988; Chapelle, 1991). Cependant, la mise à jour des propriétés et des faiblesses du modèle, par la confrontation des simulations avec les mesures et par l'analyse de sensibilité permet de focaliser l'attention sur certain mécanismes et composants essentiels qui conditionnent l'évolution du système étudié, et de suggérer des priorités de recherche. En l'occurrence, la définition de la capacité trophique du bassin pourra être mieux précisée à partir des actions suivantes:
la mise au point d'un modèle courantologique fiable. La résolution des problèmes de diffusivité numérique n'implique pas nécéssairement l'abandon des modèles on boites au profit des outils plus précis, mais coûteux, développés par les hydrauliciens. Des améliorations peuvent être apportées à partir d'une réflexion sur la cohérence des échelles spatiales et temporelles et sur les schémas de calculs appropriés;
la représentation plus réaliste des variations temporelles et spatiales des concentrations particulaires. Outre le fait que le modèle n'est pas en mesure de reproduire correctement le signal dans sa composante liée au cycle de marée, il ne prend pas en compte les perturbations induites par les phénomènes climatiques accidentels (le vent, principalement) et la variation des régimes de la Charente (qui perturbent la circulation des masses d'eau et le transport particulaire (Bacher, 1989). La mise en place d'un modèle sédimentaire devrait permettre d'intégré de manière explicite différentes sources de variabilité du milieu (voir chapitre suivant);
l'acquisition de données sur le terrain. Il est apparu que les stratégies d'échantillonnage mises en place pour aborder la problématique des mollusques ne s'adaptaient pas toujours aux exigences du modèle de production primaire (an moins sur un plan spatial). Des campagnes de mesures sont en cours pour étudier la variabilité spatiale et temporelle des paramètres hydrobiologiques aux frontières nord et sud du bassin.
BIBLIOGRAOHIE
ANON - 1979 - Dispersion de la pollution dans les pertuis charentais - Études sur modèles mathématiques
Rapport d'essais
Laboratoire Central Hydraulique de France : 12 p.
BACHER C - 1989 - Capacité trophique du bassin de Marennes-Oléron : couplage d'un modèle de
transport particulaire et d'un modèle croissance de l'huître C. gigas
Aquat. Living Ressources - 2. 199–214
BACHER C., BAURD J.P., DESLOUS-PAOLI J.M., DRENO J.P., HÉRAL M., MAURER D. &
PROU J, - 1986
A methodology for stocks assessment of cultivated oyster along the French Atlantic coasts
J. Cons. int. Explor. Mer. - CM 1986/K : 36 p.
BARETTA J.W. & RUARDIJ P.- 1988 - Tidal flat estuaries (simulation and analysis to the Ems estuary). Springer Verlag Heidelberg - Ecological studies - Vol. 71
BAYNE B.L., WIDDOWS J. & THOMPSON R.J. - 1976 - Physiological integrations
In : Marine musseles : their ecology and phsiology
B.L. Bayne Ed. - IBP № 10 - Cambridge Univ. - Press,. 261–291
BIRD S.L. & HALL R. - 1988 - Coupling hydrodynamics to a multiple-box water quality model Technical report EL-88-7 - US army engineers waterways experiment station - Vicksburg - Miss.
BODOY A., BACHER C. & GEAIRON P. - 1987 - Estimation des stocks d'huître cultivés dans
le Bassin de Marennes-Oléron en 1986
Rapp. IFREMER - DRV 87-005 RATREM-71 p.
BROCK T.D. - 1981 - Calculating solar radiation for ecological studies
Ecol. Modell, - 14,1–19
CLOERN J.E. - 1987 - Turbidity as a control on phytoplancton biomass and productivity in estuaries
Cont. Shelf Res. - 7, 1367–1381
DECHAMBOY C., PONTIER L., SIRIOU F. & VOUVE J. - 1977 - Apport de la thermographie
infra-rouge aéroportée à la connaissance de la dynamique superficielle des estuaires (système
Charente - Seudre - Anse de l'Aiguillan)
C.R. - Acad. Paris - 284, 1269–1272
DESLOUS-PAOLI J.M. & HÉRAL M. - 1984 - Transferts énergétiques entre la nourriture disponible
dans le'un bassin ostréicole et l'huître Crassostrea gigas âgée l an
Haliotis - 14,79–90
FOSTER-SMITII R.L. - 1975 b - The effect of concentration of suspension on the filtration rate and pseudo-fecal production for Mytilus edulis L., Cerastoderma edule L. and Venerupis pullastra J. Exp. Mar. Ecol. - 17,1–22
HÉRAL M. - 1977 - Étude préliminaires des potentialités nutritives dans le Bassin de Marennes-Oléron. Océanexpo Bordeaux : 14 p.
HÉRAL M. - 1989 - Modélisation des écosystèmes conchylicoles : bassin de Marennes-Oléron Rapport de fin de contrat M.E.R.S. 86-JOOJI-IFREMER CNRS, 186 p.
HÉRAL M., DESLOUS-PAOLI J.M. & PROU J. 1986 - Dynamiques de Marennes-Oléron masses des huîtres (Crassostrea gigas et Crassostrea angulata) dans le bassin de Marennes-Oléron depuis un siècle. Note C.I.E.M. - CM 1986/F : 41.
HÉRAL M., RAZET D., MAESTRINTI S.Y. & GARNIER J. - 1980 - Composition de la matière organique particulaire dans les eaux du Bassins de Marennes-Oléron - Apport énergétique pour la nutrition de l'huître. Note au conseil International Exploration de la Mer - Comité océano. Bio. L. - 44; 20 p.
HÉRAL M., ROBERT J.M., TRUQUET J., BARBAROUX O., GARNIER J. & RAZET D. - 1981
Composition en éléments dissous de l'eau du bassin conchylicole de Marennes-Oléron.
Note au Conseil Internatioanl Exploration de la Mer - comité de la qualité de l'envrionnement
marin e58
JORGENSEN S.E. - 1988 - Fundamentals of Ecological Modelling. Developements in Environemental Modelling 9 - Elsevier, Amsterdam - Oxford - New York - Tokyo
KEIZER P.D., GORDON D.C. Jr, SCHWINGHAMER P., DABORN G.R. & EBENHOEH W. - 1987 - Comberland Basin Ecosystem Model : Structure, Performance and Evaluation. Can Tech. Report to Fish. and Aqua. Sci., № 1547
KLEPPER O. & SCHOLTEN H. - 1988 - A model of carbon flows in relation to macrobenthic food supply in the Oostershelde estuary. Balans - Nota 42
MENESGUEN A. - 1989 - Modélisation de la production primaire dans le bassin de Marennes-Oléron (France). ICES EMEM:№60
RAVAIL B., HÉRAL M. & MAESTRINI S.Y. - 1988 - Incidence des débits de la Charente sur la capacité biotique du Bassin ostréicole de Marennes-Oléron. Rapp. MRS 86–50051 - Oct. 1987 ; 23–63
RAVAIL B., HÉRAL M., MAESTRINI S.Y., ROBERT J.M., RAZET D. & PROU J. - 1987 Incidence de la diminution des débits de la Charente sur la production primaire du Bassin de Marennes-Oléron. Rapp. IFREMER D.R.V.№ 87008/RA/TREM
WIDDOWS J., FIETH P. & WORRALL C.M. - 1979 - Relationships between seston, available food and feeding activity in the common mussel Mytilus edulis. Mar. Biol. - 50 ; 195–207
ANNEXE 1
PRÉSENTATION DU MODÈLE DE PRODUCTION OSTRÉICOLE
1 - ÉQUATIONS DIFFÉRENTILELLES
Azote minéral (μM/1) :
dNo3/dt = minéralisation. Morge + excrétion. Zoo - rationphyto.Phyto
Azote Phytoplanctonique (μM/1) :
dPhyto/dt = (rationphyto-mortphyto). Phyto - rationzoo. Zoo - ingphytH / volume
Azote Zooplanctonique (μM/1) :
dZoo/dt = (rationzoo. assilmilzonn - xmortzoo - excretionzoo - Zoo
Azote organique détritique benthique (mM/m2) :
dMorgb/dt = Sédiment. (fluxfond. Morge. Hteau - Trelargage. Morgb) + (fecphyt + fecmorge)
/ Volume. Hteau
Azote vivant benthique ( mM/m2) :
dNhui/dt = (assiphyt + assimorge) / Volume. Hteau
Azote detritique de l'eau (μM/1) :
fMorge/dt = - DN03 - Dphyto - (Dmorgb + DNhui) / Hteau
Valeur énergétique de la chair des huîtres de 1 et 2 ans (calories) :
dHui1/dt = Sfg 1 - Dponte 1
dHui2/dt = sfg 2 - Dponte 2
Nombre d'huîtres de 1 et 2 ans :
dSto 1/dt = morthui1. Sto 1
dSto 2/dt = morthui2. Sto 2
Volume : volume de la boîte i (1)
Hteau : hauteur d'eau de la boîte i
fluxfond : taux de dépôt de la matière organique détritique (m.o.d) (j-1)
Trelargage : taux de remise en suspension de la m.o.d. benthique (j-1)
2 - LOIS DU MODèLE BIOLOGIQUE
Minéralisation : taux de reminéralisation de la matière organique (j-1)
minéralisation = B1. effetchaleur
rationphyto : taux de croissance brute du phytoplancton (j-1)
rationphyto = rationphytomax. effetselnut. effetlumière
rationphytomax = B11. effetchaleur
effetselnut = No3 / (No3 + B13)
effetchaleur = exp ( B3. Température)
Morphyto : taux de mortalité du phytoplancton (j-1) :
morphyto = B23. effetselnut + B24. (effetselnut)
rationzoo : zooplanctonique (j-1)
rationzoo = B41. effetchaleur. effetphyto
effetphyto = 1 - exp (-azotivlev. effetchaleur. max. (o. phyto-pytoseuil))
azotivlev = B42. 2S/(B17 + B18)
phytoseuil = B22. (B17 + B18)/28
assimilzoo : fraction assimilée de la ration (s.d)
assimilzoo = B43. exp (-B44. rationzoo)
excretionzoo : taux d'excrétion du zooplancton (j-1)
excretionzoo = max (rationzoo. assimilzoo. B45. B46. effetchaleur)
mortzoo : taux de mortalité du zooplancton (j-1)
mortzoo = B49
Si la boîte i est pourvue d'huîtres, on a alors :
avec k : classe d'âge de l'huître, Wkj : poids de l'individu (gPs) et TMP : température de l'eau
ingphyt : quantitié de phytoplancton ingéré (μM/h) :
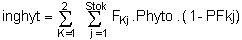
Fkj = B70. exp (B50. min (0.B51 - SES)). Wkj B71
PFkj = B52. (1-exp (B53. min (0.B54 - constHkj)) + (1 - B52). (1 - exp (B55. min (0.B56-constkj))
constkj = Fkj. SES
constkj : consomation de seston par unité de poids sec (mgPs/h/gPs) :
Fkj : taux de filtration individuel (1/h/huître)
PFkj : % de la nourriture consommée qui émise sous forme de pseudo-feces
SES : concentration de seston (mgPs/1)
feephyt : Quantité de phytoplancton biodéposé (μM/h) :
feephyto = ingphyt. (1 - AE)
AE = B57. TMP + B58
AE : efficacité d'assimilation du phytoplancton ingéré (s.d)
fecmorge : quantité de matière organique détritique biodéposée (μ/M/h) :
fecmorge = ingmorge. (1 - AE)
ingmorge : quantité ingérée de matière organique détritique (μM/h)
Sfgkj : gain de poids individuel des huîtres de la kème classe d'âge (calories/h/huître) :
Sfgkj = (assimorgek. B60 + assiphytk. B61) / Stok - Rkj
assimorgek = ingmorgek. AE
assiphytk =ingphytk. AE
Rkj = B62. TMP + B63
assimorgek : quantité de matière organique détritique assimilée par les huîtres de la Kème classe d'âge (μM/h/Stok)
assiphytk : qauntité de phytoplancton assimilé par les huîtres de la Kème classe d'âge (μM/h/Stok)
Rkj : perte calorique individuelle due à la respiration (calories/h/huître).
Dpontekj : énergie individuelle consacrée à l'effort de ponte (calories/h/huître)
Dpontekj = (B64. Wkj B65) (1/B66)
morthuil.2 : taux de mortalité des populations H1 et H2 (s.d)
morthuil = B92 : morthui2 = B93
3 - LISTE DES PARAMÈTRES
| B1 | : | Taux de reminéralisation de l'Azote Org. Det. à 0°c | 0.04 j-1 |
| B3 | : | Taux d'augmentat° des vitesses selon la température | 0.07°C-1 |
| B11 | : | Taux max. de croissance brute du phytoplancton à 0°c | 0.5 j-1 |
| B13 | : | Constante de Michaelis du Phyto-pour l'azote | 0.8 μMN/1 |
| B17 | : | Quota min. d'azote du phytoplancton | 15 gN/gChla |
| B18 | : | Quota max. d'azote du phytoplancton | 25 gN/gChla |
| B22 | : | Seuil d'échappement de la prédation | 0.5μgChla/1 |
| B23 | : | Taux min. de mortalité naturelle du phytoplancton | 0.01 j-1 |
| B24 | : | Taux max. de mortalité naturelle du phytoplancton | 0.1 j-1 |
| B41 | : | Ration relative maximum du zooplancton à 0°c | 0.25 j-1 |
| B42 | : | Delta d'Ivlev du zooplancton à 0°c | 0.12μChla-1 |
| B43 | : | Taux d'assimilation du zooplancton | 0.6 s.d |
| B44 | : | Taux relatif de décroissance d'assimilation selon la ration | 0.5 s.d |
| B45 | : | Fraction de la nourriture assimilée excrétée á 0°c | 0.15 s.d |
| B46 | : | Taux d'excretion basal du zooplancton à 0°c | 0.025 j-1 |
| B49 | : | Taux basal de mortalité du zooplancton à 0°c | 0.035 j-1 |
| B50 | : | coceflicient de la fonction de colmatage branchial | 0.07 s.d |
| B51 | : | Seuil de déclanchement du colmatage branchial | 200 mg/1 |
| B52 | : | Taux maximal de production de pseudo-fécès en présence de mécanismes sélectifs | 0.4 s.d |
| B53 | : | Exposant de la loi de production de pseudo-fécès | 0.15 s.d |
| B54 | : | Seuil de déclanchement de l'émission de pseudo-fécès | 10 mgPs/h/g Ps |
| B55 | : | Exposant de la loi de production de pseudo-fécès | 0.01 s.d |
| B56 | : | Seuil d'affaiblissement des processus sélectifs pré-ingestif | 100 mgPs/h/gPs |
| B57 | : | Pente de la fonction AE = f (TMP) | 0.033 C-1 |
| B58 | : | Efficacité d'assimilation à 0°C | 0.033 s.d |
| B60 | : | Coefficient de conversion calorique de la matière organique détritique | 0.4 cal/mgPs |
| B61 | : | Coefficient de conversion calorique de la chlorophylle | 0.684 cal/μgChla |
| B62 | : | Pente de la fonction R = f (TMP) | 0.032 mlO2/h/gPs/°C |
| B63 | : | Ordonnée à l'origine de la fonction R = (TMP) | -0.022 ml02/h/gPs |
| B64 | : | Taux de ponte pour un individu de 1gPs | 0.27 s.d |
| B65 | : | Exposant allométrique de la ponte | 1.28 s.d |
| B66 | : | Durée de la ponte | 58 jours |
| B92 | : | Taux de mortalité des huîtres de 1 an | 1.5 10-3 j-1 |
| B93 | : | Taux de mortalité des huîtres de 2 ans | 3.4 10-4 j-1 |
Par Mr. Cédric BACHER
France
ABSTRACT
L'influence du stock d'huîtres (Crassostrea gigas) cultivées dans le Bassin de Marennes-Oléron, sur les performances de croissance individuelles a été étudiée grâce à un modèle couplant des mécanismes biologiques et hydrodynamiques. Ce modèle comporte une équation du bilan énergétique d'un individu moyen et des termes de transport advectif de la matière particulaire organique et inorganique entre les 12 boîtes définies dans le découpage spatial du bassin. Une analyse de sensibilité permet de calculer l'influence de la répartition du stock dans ces boîtes sur la croissance moyenne individuelle dans chaque boîte. Il apparaît ainsi que cette influence est surtout notable dans le sud du bassin. De même, l'impact du stock des populations de compétiteurs filtreurs a été calculé et est apparu négligeable par rapport à l'impact précédent. Enfin, la différence de réponses selon les zones géographiques a été expliquée par l'existence d'une circulation résiduelle de marée dirigée vers le sud. Le milieu est ainsi appauvri au fur et à mesure du transit des masses d'eau au-dessus des populations d'huîtres.
A model coupling biological and hydrodynamical processes is used in order to study the influence of the amount of cultivated oysters (Crassostrea gigas) on growth performance in the Bay of Marennes-Oléron. The model is based on an energetic budget equation of a mean oyster and on a transport équation of particulate organic and inorganic matter through 12 spatial boxes. Sensitivity analysis is performed and allows computation of the impact of the distribution of oysters among the boxes on the growth performance of a mean oyster in each box. It is shown that, mainly in the south of the bay, individual growth is affected by the increase in the stock. The same approach also shows that the response of individual growth to the amount of natural benthic filter-feeding population is negligible compared with the preceding effect. Finally, the spatial variability of these responses is explained by the tidally induced residual circulation of the water throughout the bay from north to south. The water column is thus impoverished as the organic matter is filtered by the oyster populations.
INTRODUCTION
Les relations trophiques entre une population d'huîtres (Crassostrea gigas), cultivées dans le Bassin de Marennes-Oléron, et la nourriture potentielle, on été modélisées en tenant compte des caractéristiques du milieu estuarien (Bacher, 1989). Un modèle en boîtes (Fig. 1) permet de coupler le transport du matériel particulaire et son assimilation par la population d'huîtres. Les échanges entre boîtes dépendent de leur volume, de l'excursion de marée et des débits résiduels calculés à partir des résultats d'un modèle numérique courantologique. La consommation, l'assimilation et la respiration sont les termes d'une équation de bilan énergétique permettant de simuler la croissance des huîtres (Fig. 2). Le modèle met en évidence un gradient de croissance selon un axe sud-nord et l'interaction stock-performance de croissance (Bacher, 1989).
Figure 1 - Structure en boites adaptées sur la zone de production ostréicole
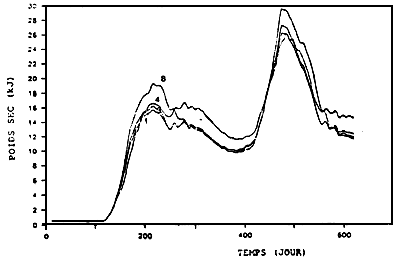
Figure 2 - Croissances de huîtres simulées en différentes boîtes, selon un gradient Nord-Sud (boîtes 8, 4,3,1)
Une analyse plus fine des différents niveaux d'interaction entre la croissance des huîtres et les stocks d'huîtres ou de compétiteurs de chacune des boîtes, est présentée au moyen d'une analyse de sensibilité, consistant à mesurer le comportement des variables d'état en fonction des erreurs d'estimation ou de mesure des paramètres du modèle (Platt et Radach, 1981), ou de la structure de ce dernier (nombre de compartiments, algorithmes de résolution des équations différentielles, valeurs initiales, du pas de temps (Recknagel, 1984).
De façon générale, les buts de l'analyse de sensibilité sont les suivants:
calculer la précision de variables d'état (sortie) en fonction de celle des paramètres (Majkowski et al., 1981). Certains auteurs parlent de l'étude de la propogation de la propagation de l'erreur (Huson, 1982) ou de l'analyse des erreurs («error analysis», Bax, 1985);
définir des axes de recherche, orienter l'acquisition de données de façon à affiner la valeur prédictive du modèle et diminuer sa sensibilité aux erreurs d;échantillonnage (Majkowski, 1982). Selon Argentesi et Olivi (1976), Summers et McKellar (1981), il s'agit du problème inverse du précédent, à savoir que la précision des paramètres mesurés est calculée en fonction de la précision du modèle. Johnson et al. (1987) testent les interactions entre paramètres O'Neil et al. (1980) montrent que les corrélations entre certains paramètres permettent de diminuer la variance des variables simulées, d'où l'intérêt de mesurer ces paramètres simultanément;
3.comparet le fonctionnement d'écosystèmes étudiés, avec un même modèle, selon les valeurs des coefficients de sensibilité qui permettent de hiérarchiser les paramètres et les paramètres et les variables de sortie. Pour Platt et Radach (1981), le calcul des coefficients de sensibilité, en permettant de visualiser l'ensemble des interactions du système, est à comparer à d'autres outils du modélisateur, comme l'analyse des systèmes («system analysis» ou «loop analysis»);
étudier l'impact d'une perturbation du milieu, introduite sous la forme de variation de paramètres, de conditions initiales ou de variables forçantes (Platt et Radach), 1981 - Kelly et Spofford, 1977);
valider le modèle (Majkowski, 1982 - Recknagel, 1984). Argentesis et Olivi (1976) concluent ainsi à inadéquation du modèle du lac Endine, trop sensible à certains paramètres difficiles à estimer. Pour Fedra et al. (1981), l'utilisation du modèle est indissociable de l'analyse de sensibilité. Celle-ci permet de prendre en compte l'incertitude des données pour estimer un ensemble (au sens mathématique) de vecteurs paramètres possibles, qui produisent autant de résultats (simulations) acceptables. La prédiction se présente alors sous forme d'une distribution multidimensionnelle des variables de sortie. Les distributions des vecteurs paramètres et des variables de sortie sont une caractéristique mathématique de la structure du modèle qui introduit le problème d'identification des paramètres (définition, justification des mécanismes, estimation), et de la validité du modèle.
Dans le cas présent et étant données les hypothèses simplificatrices sur lesquelles repose le modèle (conditions moyennes aux limites, découplage entre les maillons primaire et secondaire, non prise en compte de l'interface eau sédiment), il n'est pas question de prédire l'évolution du système dans des conditions radicalement différentes des conditions imposées. En attendant son développement ultérieur, qui intégrera les niveaux négligés jusque là, le modèle est capable d'estimer l'importance des interactions entre les différents compartiments au voisinage de la situation moyenne définie a priori.
S'il existe de solutions analytiques, faisant intervenir les dérivées partielles des variables d'état par rapport aux paramètres, adaptées aux modèles linéaires ou en régime permanent (Kelly et Spofford, 1977 - Halfon, 1977), par contre le simulations sont utilisées dans le cas dynamique non linéaire (Majkowski, 1982). Ces auteurs opposent également les calculs «paramètre» à l'approche globale (ou statistique; Bax, 1985) consistant à tirer aléatoirement des valeurs de paramètres dans des distributions indépendantes (méthode de Monte-Carlo), triangulaire (Bax, 1985), rectangulaire ou normale (Huson, 1982). Les interactions entre paramètres sont néanmoins calculées en fonction de leurs corrélations définies a priori par O, Neil et al, (1980), ou testées par analyse de variance sur leurs effets croisés (Johnson et al., 1987). L;étude multi-paramètres est facilitée par l'utilisation des techniques d'analyse multivariée, telles la régression multiple (hiérarchisation des paramètres pour chaque variable expliquée, Majkowski et al., 1981) ou l'analyse en composantes principales (liens entre variables expliquées; Bax, 1985 - Iluson,1982).
MÉTHODE
(A) - PRINCIPE
Pratiquement, pour un paramètre p dont on veut étudier l'influence, un tirage aléatoire et réalisé à partir d'une loi normale N(po.s). po étant la valeur retenue dans le modèle, et s l'écart-type de la loi. La valeur de s est choisie arbitrairement dans la mesure où il, ne s'agit pas d'étudier l'impact de l'incertitude de l'estimation du paramètre p sur les prédictions du modèle. Le modèle est utilisé avec une nouvelle valeur de p.
La comparaison des nouveaux résultats et des premiers nécessite le calcul d'un critère d'écart. Selon le but de l'étude, plusieurs critères peuvent être proposés (Recknagel, 1984). Pour mesurer l'impact sur la croissance, le critère suivant a été retenu:
F1 = ∑ (W1(t) - W0(t))2
qui représente l'écart moyen sur deux années de croissance, W0, W1 désignant le poids individuel à l'instant t dans la simulation originale et la simulation du système perturbé.
Le critère choisi est calculé dans chaque boîte où la croissance est simulée, pour un nombre N de simulations avec des valeurs différentes du paramètre p d'une simulation à l'autre. L'étude statistique de l'évolution du critère en fonction de différents paramètres permet de hiérarchiser les impacts d'une fluctuation de ce paramètres sur la croissance.
Appelons (x0l…x0n) l'ensemble des valeurs moyennes des stocks de chaque classe d'âge dans les boîtes. (x1……xn) la réalisation d'un tirage aléatoire de n valeurs indépendantes et (y1……yq) l'ensemble des valeurs de la fonction critère choisie résultant de la modification des stocks d'huîtres et de compétiteurs. Puisque le critère se rapporte à la croissance sur deux ans, q est le nombre de boîtes où cette croissance est simulée. On écrit alors:
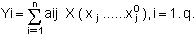
Appelons s(yi) l'écart-type de yi et s(xi) l'écart-type de xi. On a:
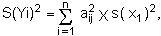
si (xj- xnj) est petit (développement limité du ler ordre). Il revient au même d'écrire:
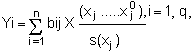
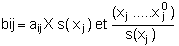
bij exprime la sensibilité de la croissance à un taux de fluctuation du stock constante d'une boîte à l'autre; en effet:
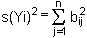
chaque terme b2ij représentant la contribution de la fluction de xj à la fluctuation de la croissance dans la boîte i.
Les bij calculés par régression linéaire de yi fonction des xi', à partir des résultats de N simulations. La matrice B- (bij) forme la matrice de sensibilité.
(b)- SENSIBILITÉ AU STOCK D'HUITRES
Ce type de calcul a été dans un premier temps pour quantifier l'influence du stock d'huîtres et de sa répartition sur la croissance. Le stock d'huîtres varie dans chaque boîte et pour chaque classe d'âge selon une loi normale. Le tirage aléatoire de N conditions de stocks est réalisé de façon indépendante dans chaque boîte. La matrice B= (bij) désigne la matrice des coefficients de sensibilité des croissances simulées dans chaque boîte aux fluctuations des stocks. L'indice i représente le numéro de la boîte l'indice identifie la boîte et la classe d'âge dont le stock varie. La matrice B peut être analysée comme un tableau de contingence dont les cases représentent la répartition de la sensibilité globale du système. Une analyse factorielle des correspondances (AFC) permet alors de visualiser sur un plan factoriel l'essentiel de la sensibilité du système.
(c) - SENSIBILITÉ AUX STOCKS DE COMPÉTITEURS
L'influence des trois espèces de compétiteurs, Mytilus, Crepidula fornicata et Cerastoderma edule sur la croissance de Crassostrea gigas, a été étudiée à l'aide de l'analyse de sensibilité. initialement, les compétiteurs sont entrés sous forme de variables forçantes d'assimilation dans chaque boîte du modèle. Pour chaque espèce, l'assimilation annuelle totale a été estimée. L'assimilation journalière totale en été déduite puis modulée de façon à reproduire l'évolution saisonnière classiquement reliée à la température. Les pourcentages de répartition dans les boîtes ont été appliqués aux estimations journalières. Les variables explicatives prises en compte dans cette étude sont donc pourcentages par espèce et par boîte. Les tirages aléatoires indépendants de nouveaux pourcentages, dont les moyennes sont les valeurs d'origine, permettent de lancer de nouvelles simultations et de mesurer l'impact de la répartition et du stock de compétiteurs sur la croissance des huîtres. La variable expliquée est le poids final au bout de deux années de croissance. Les coefficients de régression de la variable expliquée par rapport aux variables explicatives sont calculés à partir de 100 simultation puis analysés par une AFC.
RÉSULTATS
(a) - SENSIBILITÉ AU STOCK D'HUÎTRES
L'analyse de sensibilité fournit la pente de la courbe: croissance= f(stock) (effets marginaux). Les résultats de 100 simulations concernant l'impact du niveau du stock sur la croissance ont été dépouillés par un calcul de coefficients e régression (tableau 1) et une analyse factorielle des correspondances. Les huîtres sont réparties dans 6 boîtes (1, 3, 4, 5, 8, 11) selon deux classes d'âge.
L'axe l oppose les boîtes où l'interaction est la plus faible (nord) à celle de sensibilités fortes (sud) (Fig.3). Sur l'axe 2, il apparaît que les boîtes nord-est sont moins sensibles à l'évolution du stock. D'autre part, la proximité entre stocks et croissances illustre la sensibilité de la croissance d'une boîte donnée aux fluctuations du stock dans la même boîte et dans les boîtes situées en amount, e'est-à-dire plus au nord. La proximité des modalités des modalités des stocks 1 an et 2 ans d'une même boîte indique que les influences relatives de ces stocks sur les croissances de l'ensemble des boîtes sont comparables.
Les coefficients de régression sont ainsi tous négatifs et passent de la valeur-0.12 à-0.007 en moyenne pour la classe d'âge 1 de la boîte 8. Ces coefficients sont environ 2 fois plus élevés (en valeur absolue) pour la classe 2 ans, ce qui illustre l'amplification de l'impact de l'augmentation du stock au fur et à mesure de la croissance.
Tableau 1- Coefficients de régression des performances de croissances au bout de 2 ans) en fonction des fluctuations du stock des classes d'âge de Crassostrea gigas dans les boîtes 1, 3, 4, 5, 8, 11. Les variables explicatives et expliquées sont centrées et réduites.
| Performances de croissance | |||||||
| Stock | 1 | 3 | 4 | 5 | 8 | 11 | |
| 1 an | 1 | -0.191 | -0.052 | -0.010 | -0.006 | -0.005 | -0.006 |
| 3 | -0.139 | -0.043 | -0.010 | -0.004 | -0.004 | -0.003 | |
| 4 | -0.122 | -0.038 | -0.013 | -0.004 | -0.003 | -0.004 | |
| 5 | -0.114 | -0.036 | -0.013 | -0.018 | -0.009 | -0.008 | |
| 8 | -0.119 | -0.037 | -0.015 | -0.012 | -0.016 | -0.010 | |
| 11 | -0.128 | -0.040 | -0.016 | -0.009 | -0.006 | -0.022 | |
| 2 ans | 1 | -0.334 | -0.091 | -0.020 | -0.006 | -0.004 | -0.007 |
| 3 | -0.248 | -0.078 | -0.016 | -0.005 | -0.003 | -0.006 | |
| 4 | -0.207 | -0.068 | -0.025 | -0.007 | -0.005 | -0.009 | |
| 5 | -0.199 | -0.062 | -0.023 | -0.032 | -0.015 | -0.014 | |
| 8 | -0.206 | -0.066 | -0.023 | -0.019 | -0.029 | -0.018 | |
| 11 | -0.221 | -0.068 | -0.024 | -0.012 | -0.011 | -0.037 | |
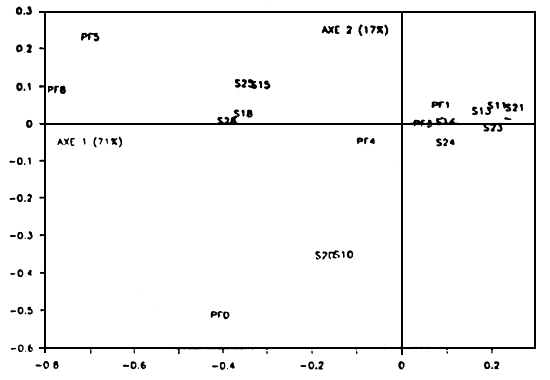
Figure 3 - Analyse factorielle des correspondances appliquée aux coefficients de régression des croissances finales en fonction des fluctuations du stock dans chaque boîte. Le stock a été réparti entre les boites 1, 3, 4, 5, 8, 11. Sij: stock de la classe d'âge i dans la boîte numéro j (la boîte II est renumérotée (?). PFj; taille à la fin de la deuxième année de croissance dans la boite j (la boîte II est renumérotée (?). Les axes 1 et 2 expliquent 88% de la sensibilité totale du modèle aux fluctuations des stocks. La proximité des modalité stock-performance de croissance indique le degré d'influence du stock sur la croissance (boîte 5, 8). Les performances proches de l'origine sont sensibles à l'ensemble des stocks (PFI, PF3). La boîte II se singularise sur l'axe 2
(b)- SENSIBILITÉ AUX STOCKS DE COMPÉTITEURS
Le tableau 2 montre les répartitions des espèces dans les boites et les coefficients de régression issus de l'analyse de sensibilité (resp). Les axes 1 et 2 de l'AFC expliquent 91% de la sensibilité globale représentée dans la table de contingence (75 et 16% resp.-Fig. 4). Les performances de croissance s'échelonnent sur l'axe l selon un axe nord-sud (boîtes 8, 5, 4, 3, 1). L'axe 2 oppose la boîte 11 aux autres boîtes. Les croissances sont perturbées par les fluctuations de stocks de compétiteurs dans les mêmes boîtes, lorsque ces stocks sont présents. Ainsi, les croissances dans les boîtes 5 et 8 sont proches des espèces 1 et 3 (moule et coque resp.). La seule espèce présente dans la boîte 11 est la coque et la performance de croissance des huîtres dans cette boîte ne dépend que de cette espèce. La position des croissances de boîtes 1, 3, 4 près du centre de gravité du nuage des points montre que les huîtres de ces boîtes subissent l'influence des compétiteurs dans ces mêmes boîtes avoisinantes (1, 2, 3, 4, 5, 6). Les coques de la boîte 6 ont surtout un impact sur la croissance dans la boîte 4, tandis que les deux espèces présentes dans la boîte 4 (crépidule, coque) ont des effets sensibles sur la croissance dans la boîte 3.
Les coefficients de régression restent malgré tout assez faibles. Seuls deux d'entre eux, relatifs à la croissance dans la boîte l, ont une valeur absolue supérieure à 0.1. Lorsque plusieurs espèces sont présentes dans la même boîte (moule et coque dans la boîte 8, coque et crépidule dans la boîte 4), leurs effets sur la croissance sont comparables.
L'impact des abondances de compétiteurs sur la croissance des huîtres est marqué par un important gradient nord-sud. Dans une moindre mesure, on observe également une opposition est-ouest. Qualitativement, ces résultats sont à rapprocher de la compétition intraspécifique déjà étudiée qui montrent l'influence des boîtes amont sur la croissance en aval (sud).
| Stocks de compétiteurs | |||||||||||
| C.E. | |||||||||||
| Performance | M.E. | C.E. | |||||||||
| de | 8 | 4 | 1 | 2 | 3 | 4 | 5 | 6 | 8 | 11 | 12 |
| Croissance | (100%) | (100%) | (100%) | (100%) | (100%) | (100%) | (100%) | (100%) | (100%) | (100%) | (100%) |
| 1 | -0,153 | -0,112 | -0,037 | -0.036 | -0,035 | -0,063 | -0,029 | -0,030 | -0,029 | 0,030 | -0,030 |
| 3 | -0,064 | -0,047 | -0,014 | -0,014 | -0,015 | -0,027 | -0,012 | -0,013 | -0,012 | -0,012 | -0,013 |
| 4 | -0,026 | -0,018 | -0,003 | -0,004 | -0,004 | -0,010 | -0,004 | -0,005 | -0,005 | -0,005 | -0,005 |
| 5 | -0,021 | -0,005 | -0,001 | -0,001 | -0,001 | -0,003 | -0,006 | -0,003 | -0,004 | -0,002 | -0,002 |
| 8 | -0,028 | -0,004 | -0,001 | -0,001 | -0,001 | -0,002 | -0,003 | -0,002 | -0,005 | -0,002 | -0,002 |
| 11 | -0,016 | -0,007 | -0,001 | -0,001 | -0,001 | -0,003 | -0,003 | -0,004 | -0,003 | -0,007 | -0,004 |
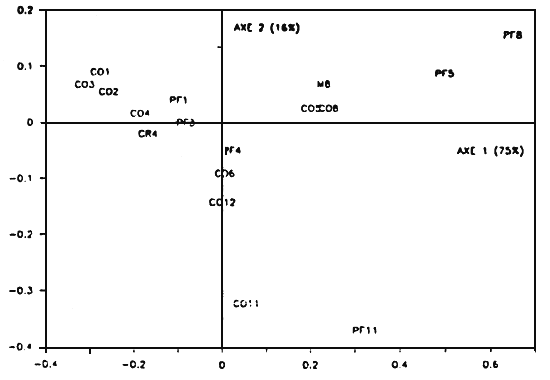
Figure 4- Analyse factorielle des correspondances appliquées aux coefficients de régression des croissances finales en fonction des fluctuations des stocks de compétiteurs dans chaque boîte. Les axes 1 et 2 expliquent 91% de la sensibilité totale du modèle. Xn: stock de l'espèce X dans la boite nX - M (moule), CR (crépidule), CO(coque). PFj: taille à la fin de la deuxième année de croissance dans la boite j.
DISCUSSION
L'analyse de sensibilité aux stocks d'huîtres et de compétiteurs benthiques permet d'approfondir les résultats préliminaires du modèle concernant l'impact du stock sur les performances de croissance des huîtres dans le bassin de Marennes-Oléron (Bacher, 1989). It apparaît ainsi que la répartition du stock a un effet non négligeable sur ces performances, et que cet effets' accroît selon I'axe nord-sud correspondent au sens de parcours des masses d'eau (Bacher, 1989). D'après la comparaison des coefficients de régression des deux études, il est évident que I'impact de la compétition entre espèces est bien inférieur à la compétition intraspécifique. Cela confirme l'hypothèse adoptée d'après la comparaison des stocks de moules, coques, crépidules (15.000 tonnes) et du stock d'huîtres (80.000 tonnes). It est également clair que la modélisation de toutes les espèces de filtreurs n'est pas nécessaire. On peut se contenter des variable forçantes introduites dans chaque boîte.
Au niveau du stock actuel l'existence de ce gradient de croissance est la confirmation d'une capacité trophique propre au bassin, dont la limite est prés d'être atteinte. Environ 10% de la nourriture transitant dans le bassin sous l'action de la marée sont ainsi utilisés pour la croissance, malgré un temps de résidence des masses d'eau relativement faible (entre 4 et 10 jours).
Au problème ralentissement de croissance sont associés des risques de mortalité accidentelle accrus, d'épizooties, de d'éséquilibres entre les divers secteurs de production liés à affaiblissement de l'organisme (Héral, comm.pers.). D'autres auteurs soulignent la nécessité du calcul de la capacité de sites de production à supporter les stocks cultivés. Carver et Mallet (sous presse), Incze et al (1981) estiment les productions potentielles en rapportant flux de nourriture et taux de filtration des populations. Pour les populations naturelles, l'importance des apports exogènes susceptibles d'être perturbés, peut entraîner le déséquilibre du système. Ainsi, d'après le modèle de Chardy (1987), 16% des apport exogènes (dus à la marée) sont assimilés par les filtreurs du système benthique dans le golfe normand-breton. Weigert et Penas-Lado (1982) soulignent également la dépendance du système côtier de la Ria de Arosa à l'égard d'un phénomène d'upwelling.
La prise en compte de la variabilité spatiale de la croissance et de la distribution de nourriture, à J'aide d'un modèle en boîte, justifiée dans cette élude par le problème de la gestion des stocks d'huîtres cultivées, est une caractéristique des écosystèmes littoraux soumis à l'action de la marée. II en résulte un couplage entre les phénomènes biologiques d'une part, les mécanismes de transport d'autre part (Uncles, 1983 - Kremer et Nixon, 1978 - Harris et al., 1984). Le calcul simultané, en utilisant l'analyse de sensibilité de l'impact d'une perturbation de l'environnement en divers points sur les variables de sortie dans l'ensemble des boîtes, s'avère un outil précieux. Kelly et Spofford (1977) l'appliquent par exemple à un problème de gestion des rejets de nitrates et phosphates dans l'estuaire du Delaware. II faut cependant préciser les limites de telles études au voisinage des valeurs mesurées. L'analyse de sensibilité fournit en général les effets marginaux des variations de l'environnement, lorsque le manque de données ne permet pas de valider le modèle en dehors du domaine de validité des hypothèses. Dans le cas présent, il est nécessaire de rappeler de rappeler que la production primaire n'a pas été modélisée. La nourriture est considérée comme variable forçante à partir des concentrations et flux advectifs aux limites géographiques du modèle. Le découplage entre productions primaire et secondaire limite alors l'utilisations du modèle à des niveaux de stocks voisins des niveaux mesurés.
BIBLIOGRAPHIE
ARGENTESI, F & OLIVIL - 1976 - Statistical sensitivity analysis of a stimulation model for the
biomass-nutrient dynamics in aquatic ecosystems.
In: roc. Summer Computer Simulation Conf.- 4th Simulation councils - La Jolla - California:
pp. 389–396
BACHER C. 1989 - Capacité trophique du bassin de Marenne-Oléron:couplage d'un modèle
de transport particulaire et d'un modèle croissance e l'huître Crassostrea gigas
Aquat. Living Res - 2(4): 189–206
BOX N.J. - 1985 - Global sensitivity analysis of SKEBUB parametrized for Balsfjord, a fjord in
Northern Norway.
NWAFC Processed Report 85-07 - National Marine Fisheries Services - US Department of
Commerce: 40 pp.
• CARVER C.E.A & MALLET A.L. - 1989 - Assessing the carrying capacity of a coastal inlet in
terms of mussel culture.
Can J. Fish. Aquat. Sci. - Sous presse
CHARDY P. 1987 - Modèle de simulation du système benthique des sédiments grossiers du golfe
normand-breton (Manche)
Occanol Acta - 10(4): 421–434
FEDRA K., VANSTRATEN G. & BECK M, B. - 1981 - Uncertainly and arbitrariness in ecosystems
modelling: a lake modelling example
Ecol Model - 13:87–110
GABRIEL, K.R. - 1971 - The biplot graphic display of matrice with application to principal component
analysis
Biometrika - 58 (3): 453–467
HALFON E. 1977 - Analytical solution of the system sensitivity equation associated with a lincar
regression
Ecol. Model - 3: 301–307
HARRIS.R. W., BALE A.j., BAYNE B.J., MONTOURA REC., MORRIS A. W., NELSON L. A.,
RADFORD P.J., UNCLES R.J., WESTON S.A. & WIDDOWS J. 1984
A preliminary model of the dipersal and biological effect of toxins in the Tamar estuary
England - Ecol. Model - 22: 253–284
HUSON L. W.- 1982 - A graphical aid to multivariate sensitivity analysis
Ecol. Model. - 16:91–98
INCZE L.S., LUTZ R. A. & TRUE E - 1981 - Model carrying capacities for bivalve molluscs
in open, suspended-culture systems
I. World Maricul. Soc. - 12:143–155
JOHNSON D.H., SPARLING D.W. & COWARDIN L.M. - 1987
A model 0f the productivity of the Mallard duck
Ecol. Model. - 38: 257–275
KELLY R.A. & SPOFFORD W.O.Jr - 1977
Applications of an ecosystem model to water quality management: the Delaware estuary
In: Ecosystem modelling in theory and practice: an introduction with case histories: 419–443
Ed. by C.A.S. Hall and J.W. Day, Jr. John Wiley and Sons: 684 pp.
KREMER J. & NIXON S.W. - 1978 - A coastal marine exosystem - Simulation and analysis
Ecol. Stud. 24 - Ed. by W.D. Billings, F. Golley, O.L. Lange & J.S. Olson - Springer Verlag:
250 pp.
MAJKOWSKI J., RIDGEWAY J.M. & MILLER D.R. 1981 - Multiplicative sensitivity analysis
and its role in development of stimulation models
Ecol. Model. - 12: 191–208
MAJKOSKI J. 1982 - Usefulness and applicability of sensitivity analysis in a multipeeices
approach to fisheries management
In Theory and management of tropical fisheries - Ed. by D. Pauly and G.I, Murphy. ICLARM
Conference Priceedings - 9.360 pp.
O'NEIL R.V., GARDNER R.H. & MANKIN J.B. - 1980 - Analysis of parameter error in a nonlinear
model
Ecol. Model. - 8: 297–311
PLATT T. & RADACH G. - 1981 - Sensitivity analysis
In Mathematical models in biological oceanograply:99.37–46
Ed. by T. Platt, K.H. Mann and R.E. Ulanowicz - Unesco Press:156 pp.
RECKNAGEL F. - 1984 - A comprehensive sensitivity analysis for an exological simulation model
Ecol. model. - 26: 77–96
SUMMERS J.K. & McKELAR ILN. Jr. - 1981 - A sensitivity analysis of an ecosystem model of
estuarine carbon flow
Ecol. Model. 13: 283–301
UNCLES R.J. - 1983 - Modeling tidal stress, circulation and mixing in the Bristol Channel as a
prerequisite for ecosystem studies
Can. J. Fish. aquat. Sci. - 40 (Suppl. 1): 8–19
WEIGERT R.G. & PENAS-LADO E. - 1982 - Original exploitation, by mussel rafts, of the Ria de
Arosa - Sapin: predictions of a first-generations model
Marine Ecosystem Modelling - Proceedings from a Workshop NOAA: 159–171
By Mr. Juan Carlos MARTINEZ COLL
Spain
ABSTRACT
The aquaculture firms face a class of legal problems with economic incidence that can be modeled upon the Coase's theorem. That is the case when other industries emissions pollute rivers or lagoons. These models are useful also when several firms compete for the exploitation of the lagoon with assigned quotas or zones. The conflicts among different farms and the possibility of overfishing the lagoon can be modeled with Game Theory methods.
INTRODUCTION
An important portion of the job in the aquaculture firms is to be made at the law courts and the negotiation tables. In order to portect the fauna in rivers and lagoons from pollution and overfishing, the aquaculturists will have to convince judges and to bargain with other firms about property rights.
The costs inherent to any transaction or negotiation process may block the way to any agreement. On the other hand, these firms have frequently to confront long-established interests, where the social and individual efficiency will play a convincing and conclusive role.
Although the law is to be considered the basic and vital framework within aquaculture business is to be conducted, the behavior of our competitors could make necessary to consider the convenience of certain uncontrolled activities. Could it be necessary, under some circumstances, to harvest over the lagal quotas?
Some Law and Economic models could be useful to understand this kind of problems. The Coase's Theorem and the Game Theory, although too much general and abstract, could suggest some answers to these questions.
1- POLLUTION AND EFFICIENCY
The law may be in contradition with the economic efficiency. If the exploitation of a lagoon is shared by a system of quotas, some of these quotas may be assigned to inefficient firms. If a paper industry has the right to pollute a river, it may be preventing a more efficient fish hatchery firm to develop its activity.
The conflict between law and efficiency may be solved easily if the quotas or property rights are exchangeable and the transactions are costless. This statement is know as Coase's Theorem and requires also a clear and unambiguous definition of the initial legal assignment of rights.
Let us suppose that a fish hatchery firm and a paper industry are struggling for the use of a river. Let us suppose that the appropriative rights are clearly set and can be exchangeable. One of both firms is the most efficient so that there may be four cases, as shown in Table 1.
Table 1
| The property right is owned by the | |||
| Fish hatchery firm | Paper industry | ||
| The most efficient is the | Fish hatchery firm | Case 1 | Case 2 |
| Efficient. There is not transaction | Inefficient.The fishery will buy the right | ||
| paper industry | Case 3 | Case 4 | |
| Inefficient.The paper industry will buy the right | Efficient.There is no transaction | ||
Case 1 offer no problem. The property right is owned by the fish hatchery firm that is also the most efficient. With the support of the law, it will prevent the paper industry from pollute the river. In case 4, the paper industry has the right over the river so that the aquaculturist firm will have to retract.
Let us see case 2 more carefully. The appropriative right is owned by the paper factory, maybe for historical reasons, while the fish hatchery would be more efficient. The concept that «first in time is first in right» is usual in national laws in many countries, and goes against the newest (and frequently more efficient) enterprises. If the fish hatchery benefits are valued in, say, 10, and the benefits obtained by the paper firm will be able to buy the competitor right for a price between 8 and 10. Both firms would improve their position with the exchange : the paper factory, without working, would get a net benefit equal or superior to that it used; the aquaculturist firm, that at first was not allowed to develop its activity, would get some benefit, between 0 and 2.
Case 3 is symmetrical to the previous one. The paper factory, more efficient, will buy the appropriative right to the fish hatchery firm.
Conclusion : regardless of the specific initial assignment of the property rights, the final outcome is that the more efficient firm will develop its activity.
2- EFFICIENCY AND EQUITY
This optimistic conclusion is based upon two unlikely requirements : that the legal rights are well-defined and that are marketable without costs. To see what the costs for the transaction may be, let us think that an inefficient paper industry is polluting a river and someone is promoting an agreement among all the individuals damaged by the pollution to buy the industry rights or to construct a purifying plant. How could be identified all the anglers? How much should pay each of them? The problem of «free-riders», people trying to get the benefit of a situation without paying its share, would make the agreement almost impossible.
So, the law has three important roles to make easy the achievement of the efficiency. First, establishing unambiguous rights; second, making these rights easily marketable without costs; third, when these costs are unavoidable, granting the rights to the more efficient firm.
This last statement raise other kind of difficulties. It is an accepted objective for the law the achievement of the efficiency? Another different criterion for the judge may be the equity. When the exploitation of a lagoon is to be shared by different firms, what criterion could be used for the assignment of the quotas? The equity may be contradictory to the efficiency. A greater firm is able to manage a lagoon more efficiently than a lot of smaller fisherman. The efficiency criterion would lead to assign the property right on the lagoon to the greater firm, the equity criterion would load to an inefficient partition of the lagoon in zones or quotas.
In order, to conciliate efficiency and equity, it can be suggested a couple of solutions. One, the quotas may be marketable, so that the more efficient firm could buy their quotas to the smaller fishermen that would participate somewhat in the benefits. Two, the exploitation of the lagoon may be put up for auction; the most efficient firm will be able to offer a greater amount of money that could be earmark to state aid or assistance for the smaller ones.
3- BREAKING THE LAW
We must now look at the possibility to transgress the law. We can overfish and others can do it also; the paper industry can try to pollute the river surreptitiously. Although and individual where absolutely law-abiding, a rational behaviour must take into account that others are not so scrupulous.
Suppose that the exploitation of a lagoon is to be shared by several fishermen under a system of quotas. Each fisherman has to choose between two strategies, Overfish or Righteous. No one can know if the other fisherman are being righteous or are overfishing.
From the point of view of any fisherman there are four possible situations, shown in Matrix 1. They have been ordered from the best one (1) to the worst (4). What is the most rational choice? What should recommend an expert on Game Theory?
MATRIX I
(Prisoners'Dilemma)
| Other fishermen | |||
| Righteous | Overfish | ||
| My firm | Righteous | 2 | 4 |
| Overfish | 1 | 3 | |
This Matrix is called «Prisoners' Dilemma» and is one of the more intensively studied by Game theorists. It hides the paradox that a rational choice leads to a very bad result.
If the others fisherman are being righteous, the best choice is Overfish; but if the others are overfishing the best best choice is also Overfish ! Whatever decision had been taken by the others my best choice is to overfish ! If all the fishermen adopt a rational decision, the outcome will be all-Overfish, although they had been far better if they had chosen Righteous.
4- MAKING THE LAW
Frequently the aquaculturist will have the possibility to influence the regulations. This is the case when two or more part negotiate a convention or agreement. Let us take as an example the negotiations between the Kingdom of Morocco and the CEE on the exploitation of Moroccan fisheries.
At any negotiation, each side will try to get the best conditions and will say that the other proposals are unacceptable. Each player bus two possible strategies to choose, to be Hard or Soft. Hart means to be strict, inflexible, without making concessions. Soft would imply offering new options, to give way, to yield ground. As before, there are four possibilities, shown in Matrix 2.
MATRIX II
(Chicken)
| C.E.E. | |||
| SOFT | HARD | ||
| Morocco | SOFT | 2 | 3 |
| HARD | 1 | 4 | |
This matrix is called «Chicken» or «Hawk-Dove» and is and well known in Game Theory literature. Many situations in everyday life can be modeled upon one of these two matrices.
The best outcome for the Kingdom of Morocco is reached by choosing Hard when the CEE had chosen Soft. But the worst choice for both is to play Hard simultaneously. Timing is a very important factor in this game. The first player will choose the Hard strategy and the other will be obliged to Soft.
5- THE POSSIBILITY OF COOPERATION
The strategies considered before-Righteous, Overfish, Soft, Hard-are too much simple. In real world, situations are repeated on time, people met one another day after day, and can recollect the behavior of the competitors. The individuals will show frequently different behaviour accordingly with the strategy of the other contender.
Taking this into account, there are other kind of strategies, the reactive strategies, to the considered. Two of them will be considered here: Tit for Tat and Bully. Tit for Tat means to answer Soft to Soft, Righteous to Righteous, Hard to Hard and Overfish to Overfish. Other usual strategy, Bully, lie in answering Hard to Soft and Soft to Hard.
In computer simulations, the most successful strategy in the Prisoners' Dilemma environment has came out to be Tit for Tat. In the problem considered here, the tendency to overfishing, this result suggests that fishermen will behave cooperatively, righteously, as far as the other fishermen act properly, but will change to the overfish behavior when the others do. The Bully strategy, on the other hand, yields not good results under the Prisonners'Dilemma conditions but is highly recommended under the Chicken environment.
Anyway, the search of individual profit does not guarantee the best individual or social outcome individual or social outcome in these environments. It is just in these circumstances where the ethical impulse would play a highly positive role leading to social mutual cooperation.
6- CONCLUSIONS
The efficiency of aquaculture enterprises is a very important weapon in lawsuits. When less efficient firms have properly rights on the river or lagoon, the efficiency will enable to identify them; when rights are ambiguous or not well established, the reason of a higher efficiency could be convincing enough for a judge.
the auction of the quotas with subsequent spread out of the benefits.
The temptation to overfish in shared lagoons can be modeled as a Prisoner's Dilemma, Although at first sight it were rational an unrighteous behaviour, the experts in game theory suggest a Tit for Tat strategy : to behave as others.
The negotiation of rights or rules will require a Hard or Soft choice that can be modeled as a Chicken game. The recommended strategy is Bully, to display the opposite behaviour as the contender, although the best outcome is not guarantee.
REFERENCES
COASE R.H. - 1960- The problem of Social Cost
Journal of Law and Economies - 3
ESTEVE SECALL R. & NARVAEZ BUENO A. - 1989 - «La Acuicultura como sector de futuro»
Acuicultura y Economia - R. Esteve, A. Narvaez, G. Ruiz y A. Ruiz - Coordinadores
Ministerio de Agricultura, Pesca y Alimentacion - Madrid
GORDON H. SCOTT - 1954 - «The Economic Theory of Common-property Resource : the
Fishery»
J. Polit. Econ. - v. 62 - Abril 1954
HIRSHLEIFER J. - 1978 - «Economics from a Biological Viewpoint»
Journal of Law and Economics - v. 20 - Abril 1978
HIRSHLEIFER J. - 1978 - «Competition, Cooperation and Conflict in Economics and Biology»
Am. Economic Rev. - v 68 - Mayo 1978
HIRSHLEIFER J. - 1982 - «Evolutionary Models in Economics and Law Cooperation versus
Conflict Strategies
Research in Law and Economics v. 4
HIRSHLEIFER J. & MARTINEZ COLL J.C. - 1988 - «What Strategies Can support the
Evolutionary Emergence of Cooperation»
J. Of Conflict Resolution - v. 32 - 2 - Junio 1988
HIRSHLEIFER J. & MARTINEZ COLL J.C. - «Selection and Mutation in Evolutionary Games»
Revista Espanola de Economia - Forthcoming
MARTINEZ COLL J.C. - 1986 - Bioeconomia
Secretariado de Publicaciones de la Universidad de Malaga - Malaga
MARTINEZ COLL J.C. - 1-1990 - «La Economia Natural, la Economia Politica y la Pesqueria,»
en Curso de lntrocuccion a la Economia de la Acucuiltura - G. Ruiz Bravo y A. Ruiz Molina-Doordinadores
- Ministerio de Agricultura - Pesca y Alimentacion - Madrid
MARTINEZ COLL J.C. & •HIRSHLEIFER J. - 1991 - «The Limits of Reciprocity : Solution
Concepts and Reactive Strategies in Evolutionary Equilibrium Models»
Rationality and Society - Enero 1991
NARVAEZ BUENO A. - 1990 - «Evaluacion de la actividad acuicola»
en Curso de Introduccion a la Economia de la Acuicultura - G. Ruiz Bravo y A. Ruiz Molina-Coordinadores
- Ministerio de Agricultura, Pesca y Alimentacion - Madrid
RUIZ BRAVO de MANSILLA G. & A. - 1986 - La economia de la empresa acuicola
Ministerio de Agricultura, Pesca y Alimentacion - Madrid
By Mr. M. BELKHIR & Mr. S.E. JORGENSEN
1 - OBJECTIVES OF THE PROJECT
Lakes and lagoons are often used to promote aquaculture activities.
The scope is to get the optimum economical benefit of such activities without production of significant adverse effects from pollution, which may even course an economical negative effect caused by low oxygen concentrations in the water.
The idea behind this project is to develop a model for the lagoon of Monastir. The model should be able to:
to apply the results of 1 – 3 to optimize the aquaculture operation.
The idea is to use this case as an example and transfer through this case study the know-how for development of such models of the application of lakes and lagoons for aquaculture to other case studies in Tunisia and North Africa.
2- DESCRIPTION OF THE PROJECT
The developed model will consist of three submodels:
the first model relates growth rate of the cultivated aquatic organisms with the forcing functions, i.e. the temperature and the feeding rate, the latter being controlled by man. The utilization of the food and the growth rate are known from the literature. The amount of pollutants coming from the aquaculture is also a part of this submodel, i.e. the amount of ammonia excreted and the amount of faeces and undigested feed. This information is used in submodel 3. The water quality influences also the growth of the fish; this is the results of submodel 3. The interrelations of the 3 submodels are shown in fig. 1;
the costs such as food, electricity for pumping, oxygen etc… are calculated in submodel 2 as
well as total daily costs and costs per kg of the production. The idea is obviously to maintain
the latter one as low as possible. The costs of food are based upon the cost per kg and the food
consumption, which is a result of submodel number 2.
The other variable costs are related to the maintenance of an acceptable water quality, which is
the results coming from submodel number 3. See again fig. 1;
submodel 3 is concerned with the water quality. If the oxygen concentration is below a certain level aeration is required, and this information is used in submodel 2. As the aquacultures are a part of natural systems it is necessary in this submodel to include the natural processes such as phytoplankton growth and oxygen production, a coarse description of the hydrodynamics (the circulation pattern), the grazing, the detritus mineralization in the natural part of the ecosystems, the uptake of nutrients and biodegradable organisms. This submodel contains an «alarm» which will be used in the two other submodels to increase aeration and/or reduce feeding rate.
3- SCHEDULE
The development of this model requires an experienced team in modelling, but it is an advantage to train a local staff already during the development of the model; it is proposed to follow a schedule along the following lines:
the model is transferred to Tunisia and a modelling course is given in Tunisia over a period of 3 weeks. The course should give a general idea about modelling but will particularly focus upon the application on the developed model.
Based upon this training course participants from other countries should understand the background of this particular model so well that they would be able to implement it for other cases of interest, including modifying the model for such cases with only minor assistance.
Number of participants : 15–20 individuals from all North African countries, 2 or 3 of them from the local staff, who have participated in the development of the model in Denmark.
4- PROJECT ECONOMY
The project consists of:
training course in the application of the agriculture model.
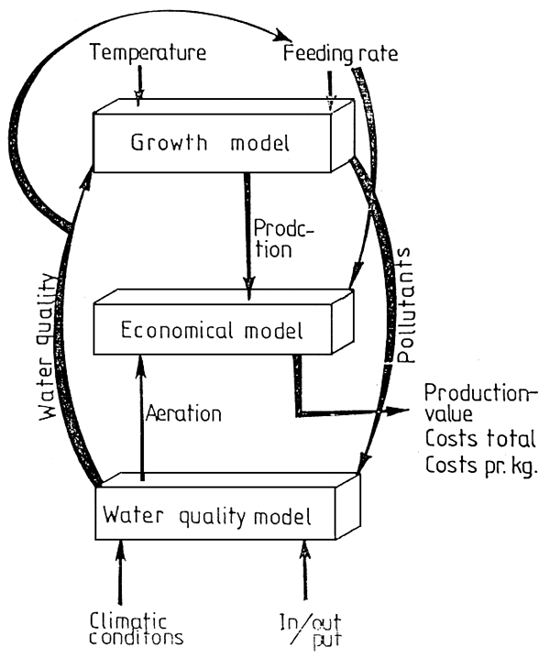
MER MÉDITERRANÉE
Lagoon of Monastir / TUNISIA (Centre National d'Aquaculture)
Par Mr. Zair ABDEL NASSER
Algérie
1 - INTRODUCTION
Le lac Mellah, qui s'identifie comme site unique en Algérie, est une cuvette côtière de 860 ha s'ouvrant sur la mer par un chenal long de 900 mètres. Orienté nord-nord-est, Mellah reçoit, outre l'eau salée rentrant aux pleines mers, les eaux douces des oueds ER-RIKBET, EL MELLAII, EL-BELAROUG et SATA.
Le cycle hydrologique du lac Mellah est dépendant des phénomènes météorologiques (précipitations évaporations et marées barométriques), par conséquent les échanges hydrauliques peuvent être variables dans le temps et dans l'espace, d'ou la variabilité de la salinité de ses eaux qui évolue d'un minimum de 23% fin mai à un maximum de 35% fin septembre.
Du fait de la faible profondeur du lac, les variations annuelles de la température sont comprises entre 10° et 30°c alors que l'oxygène dissous peut atteindre des valeurs maximales de l'ordre de 80% en surface et avoisinant le zéro dans les zones profondes du lac.
Le chenal qui constitue le «poumon» du lac présente un profil sinueux avec des profondeurs allant de 0.30 à 2 mètres. La couche sédimentaire du chenal est constituée de vases et de sables dont l'origine est vraisemblablement due aux transferts littoraux et éoliens ainsi qu'aux apports terrigène.
Quant au plan d'eau, la bathymétrie présente un profit décroissant des berges jusqu'aux zones centrales du lac. Les plus grandes profondeurs se situent autour de 6 mètres avec une grande superficie de l'ordre de 60% de la totalité, ayant des profondeurs >- 4 mètres. Le substrat du lac est recouvert d'une boue plus ou moins sableuse ou argileuse, elle-même surmontée d'une épaisseur de vase, présentant une grande zone centrale caractérisée par des boues riches en substances organiques et pauvre en oxygène.
Les espèces vivantes dans le lac Mellah, très courantes en Méditerranée, effectuent des migrations trophiques de la mer vers le lac et des tropismes génétiques dans le sens inverse. Néanmoins, certaines espèces tels que le loup et le mulet se reproduisent, malgré tout, dans le lac. Ces pontes semblent avoir lieu avec un décalage de deux mois par rapport à la mer.
II - GESTION ET AMENAGEMENT ACTUELS DU LAC MELLAH
La gestion actuelle du lac Mellah ne peut pas être considérée comme un modèle type d'aménagement lagunaire côtière. En effet, les quelques actions qui y sont entreprises peuvent se résumer en ce qui suit:
exploitation de la palourde.
II.1 ÉCLOSERIE DU LOUP
Le projet de l'écloserie du loup, à 80% de réalisation, implanté sur la berge Est du lac, aurait pour objectif la production de 200.000 alevins de loup quand l'exploitation aura atteint son rythme de croisière
Ces alevins seront destinés:
- au prégrossissement, puis au repeuplement du lac;
- l'approvisionnement de promoteurs privés;
- un élevage intensif en cages flottantes au niveau du lac.
En outre, une partie de ces alevins serait destinée au lac Oubeïra, à titre pilote, après acclimatation où le loup jouerait un rôle de police vis-à-vis des cyprinidés.
Nous estimons que l'introduction de ce prédateur est avantageuse car il ne se multiplie pas en eau douce et, donc, son développement reste contrôlable.
II est à noter que des individus de taille marchande ont été déjà pêchés au niveau du lac qui est à eau douce.
Par ailleurs, l'écloserie jouerait un rôle de formation.
II.2 PêCHE SELON LE SYSTèME BORDIGES
La pèche au niveau du lac, est représentée par deux installations actionnées alternativement le long de l'année en relation avec le tropisme du poisson.
A-Bordigues d'été
- Situation : aval du chenal
- Espèces ciblées : Mulets
- Fonctionnement : lors des courants d'appel
B- Bordigues d'hiver
- Situation: amont du chenal.
- Espèces ciblées: loups, daurades, soles, anguilles.
- Fonctionnement : lors des courants d'appel.
II.3 ÉLEVAGE DE MOULES
L'introduction de cet élevage est récente. Les techniques utilisées au niveau du lac Mellah sont celles pratiquées sur l'étang de Thau (France).
Au nombre de 62, 45 seulement sont équipées et fonctionnelles, et disposées en 4 rangées parallèles sur des fonds de 4 à 5 mètres.
Les pousses des coquillages sont excellentes et la quilité de la coquille et de la chair très bonne Cependant, l'élevage n'a pas atteint un niveau significatif avec des productions de l'ordre de 15 kg/ha/an.
II.4 EXPLOITATION DE LA PALOURDE
II s'agit d'une activité récente qui date de 1990. La récolte se pratique manuellement à l'aide d'un rateau classique doté d'une poche en filet de 1 cm de maille étirée.
Hormis les zones dites de géniteurs et de reparquages, l'exploitation de la palourde s'effectue sur les berges du lac, et essentiellement au niveau de la zone dite sablière (sud du lac). Exploité par des vacanciers, des productions de l'ordre de 250 kg/J ont été obtenue, dont la taille des individus est supérieure à 25 mm.
Des essais de récolte avec une drague ont été effectués mais n'ont pas donné les résultats escomptés; ceci est dû à l'instabilité de l'engin utilisé et à la nature du substrat occasionnant l'obstruction de l'ouverture de la drague et un colmatage du maillage.
La gestion actuelle du lac Mellah, fait ressortir des productions représentant des rendements faibles de l'ordre de:
- 30% pour les poissons (toutes espèces confondues);
- 10% pour la palourde;
- 7.5% pour les moules.
III- PERSPECTIVES DE DÉVELOPPEMENT DU LAC A DES FINS DE PECHES ET D'AQUACULTURE
Nos perspectives prennent en cosidération les différentes études menées sur le lac, notamment celles entreprises par quelques experts de MEDRAP l et par nos universités qui, d'ailleurs convergent vers les mêmes conclusions.
Le projet de développement du lac Mellah nécessite un certain nombre d'études et de travaux d'aménagement programmés dans un ordre logique pour assurer une exploitation optimale.
Parmi les raisons qui font que le lac Mellah n'est pas à son point fort de développement productif de l'aquaculture extensive, on peut citer:
- le mauvais échange hydraulique entre le lac et la mer;
- l'inadéquation entre les activités développées et les possibilités offertes, notamment, en matière de conchyliculture.
Les actions à entreprendre pour une mise en valeur optimale du lac peuvent se résumer essentiellement par:
- l'augmentation du virage du chenal, provoquant une sédimentation de algues en amont pouvant être dragués facilement et les opérations de déssablement pourraient s'opérer aisément;
- la mise en place d'écluses à l'entrée du chenal permettant une régularisation des échanges hydrauliques.
En matière d'infrastructure, les actions à entreprendre sont:
- ammenée de l'énergie électrique au niveau du lac permettant le fonctionnement de l'écloserie du loup;
- création d'une écloserie de reproduction d'huîtres et de palourdes;
- réaménagement des bordigues associées au tropisme trophique du poisson.
Un aménagement adapté permettrait au lac d'atteindre un niveau de production de l'ordre de 1.500 à 2.000 tonnes/an (toutes filières aquacoles confondues).
Le plan d'aménagement du lac se chargerait de développer:
- la mytiliculture au niveau de la zone A du lac, en créant:
- l'exploitation de la palourde au niveau de la zone B en créant:
- un élevage de loup en cages flottantes au niveau de la zone C (zone centrale du lac);
- un pare mytilicole et un pare ostréicole de 60 table au niveau de la zone D;
- un pare à palourde avec ensemencement de parcelles à partir de l'écloserie au niveau de la zone E du lac;
- réaménager et développer la pêche lagunaire en utilisant le monofilament comme engin de capture avec des ensemencements d'alevins chaque année.
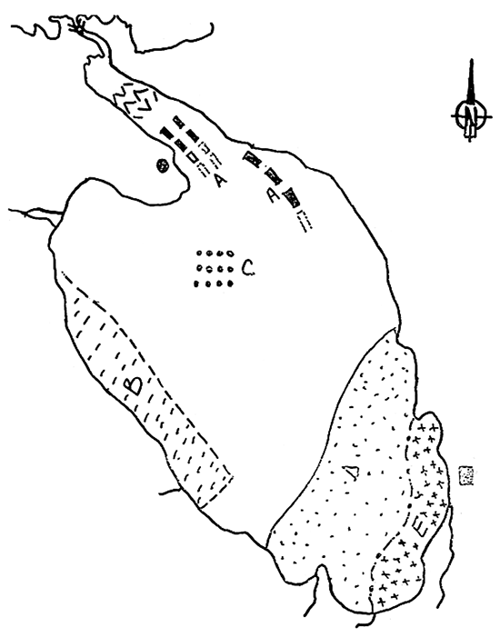
Échelle 1/25000ème
Bordigues
Tables conchylicoles
Traitement de coquillages
Exploitation de palourdes
Cages flottantes
Parc mytilicole et ostréicole
Parc à palourdes
Écloserie Huître + palourde
Représentation du plan de développement du lac Mellah
Échelle 1/25000ème
Présentation générale du lac Mellah
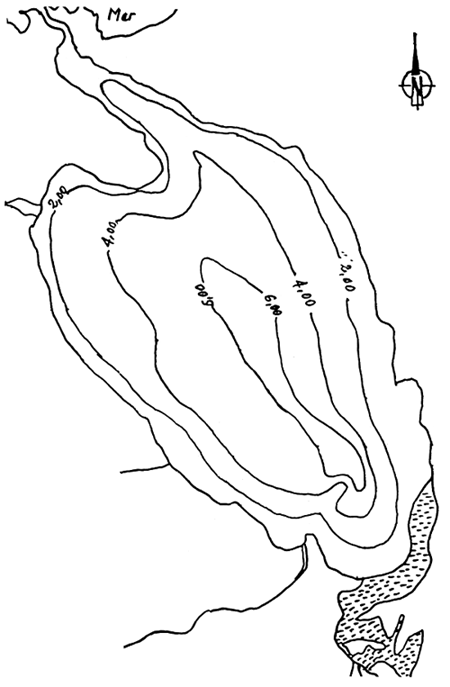
Échelle 1/25000ème
Écloserie loup
Bordigues
Tables conchylicoles
Zone dite sablière
Zone de géniteurs
Zone de reparquage
Représentation des activités actuelles au lac Mellah
![]()
![]()
![]()