![]()
![]()
![]()
Le comportement de certains pins en Amérique latine
LAMBERTO GOLFARI
LAMBERTO GOLFARI, Division des forêts, Celulosa Argentina S.A., Buenos Aires
Comme tous les autres végétaux, les conifères¹ ont des exigences, acquises au cours des temps dans le milieu où ils vivent, vis-à-vis du climat, du sol et des facteurs biotiques. Certaines espèces, comme Pinus durangensis et P. cooperi, veulent des hivers froids et supportent des températures descendant jusqu'à -27° C, tandis que d'autres, comme P. caribaea et P. hondurensis², n'ont pas besoin de saison froide et souffrent à des températures inférieures à - 4° C. Certaines espèces, comme P. khasya et P. insularis, préfèrent les étés chauds et humides et d'autres, P. patula et P. pseudostrobus par exemple, aiment les étés froids et humides. D'autres encore, comme P. radiata, veulent des étés frais et secs. Certaines, comme P. merkusii, ont une longue période de végétation et sont adaptées à des étés à jours courts, tandis que d'autres, comme P. engelmanni, ont une période de végétation plus courte et sont adaptées à des jours longs. Quelques-unes, comme P. elliottii et P. taeda, peuvent croître sur des sols mal drainés, mais la grande majorité exigent un bon drainage, c'est le cas par exemple d'Araucaria angustifolia et Cunninghamia lanceolata; certaines, comme P. cubensis, exigent des sols fortement acides, tandis que d'autres, comme P. halepensis et P. elliottii densa, poussent sur des sols neutres ou alcalins.
l Ce groupe comprend des espèces qui, bien qu'originaires de la zone tempérée, sont plantées en altitude dans la région tropicale, et aussi des espèces tropicales qui, habitant les montagnes, demandent un climat tempéré.
2 Cette désignation, incorrecte, est cependant préférée, car elle permet de distinguer P. caribaea de Cuba de P. hundurensis d'Amérique centrale qui ont des exigences écologiques et un comportement en plantation nettement différents.
Lorsqu'un pin ou un autre conifère est introduit dans une nouvelle région, son succès ou son échec dépendra donc de la mesure dans laquelle les conditions écologiques locales coïncident avec ses exigences. L'interaction de tous les facteurs dans le complexe écologique favorise un certain nombre d'espèces, suivant leurs réactions propres. Il peut arriver, dans le meilleur cas, qu'une espèce trouve les conditions optimums dans son nouveau milieu, ou encore qu'elle trouve des conditions assez bonnes en ce qui concerne certains facteurs et défavorables pour d'autres, ou enfin des conditions absolument défavorables.
Lorsque l'expérimentation est assez avancée, le forestier peut juger le comportement des différents conifères d'après certains indices: vitesse de croissance; port, de la cime surtout; longévité; sensibilité au froid; résistance aux insectes et aux maladies, etc. D'après ces indices, et en les replaçant dans le contexte des conditions du milieu, il est possible de choisir les espèces les mieux adaptées à chaque région et même de choisir les meilleures stations en les évaluant et en les classant suivant leur aptitude pour le développement des arbres en station excellente, bonne, marginale ou inutilisable.
Dans les régions où l'expérimentation est encore insuffisante, les forestiers ne peuvent que choisir les espèces dont le succès paraît le mieux assuré, en essayant de trouver des analogies entre la station d'origine de ces espèces et le nouveau milieu, en prenant en considération les facteurs suivants: pluviosité et sa répartition saisonnière; températures moyennes annuelle et mensuelle, et températures extrêmes; humidité relative; longueur des saisons humide et sèche; intensité lumineuse et durée d'éclairement; profondeur, texture, drainage, pente et pH des sols; conditions biotiques, etc.
Lorsqu'il est impossible d'obtenir des renseignements sur l'habitat de certaines espèces encore relativement mal connues comme celles de la région tropicale, il peut suffire d'examiner trois facteurs fondamentaux, étroitement liés: répartition des pluies, pluviosité moyenne annuelle et régime des températures³.
3 On emploiera ici dans une certaine mesure les facteurs et indices déjà adoptés par Pavari (1916), Troup (1932), Swain (1938) et Holdridge (1947) dans leurs classifications des climats établies spécialement pour des buts forestiers.
Les pins et autres conifères peuvent être répartis en trois grands groupes en ce qui concerne le régime des pluies:
1. Espèces qui demandent les pluies surtout en hiver et qui peuvent supporter une période de sécheresse estivale (fig. l).
2. Espèces exigeant une pluviosité uniformément répartie tout au long de l'année (fig. 2).
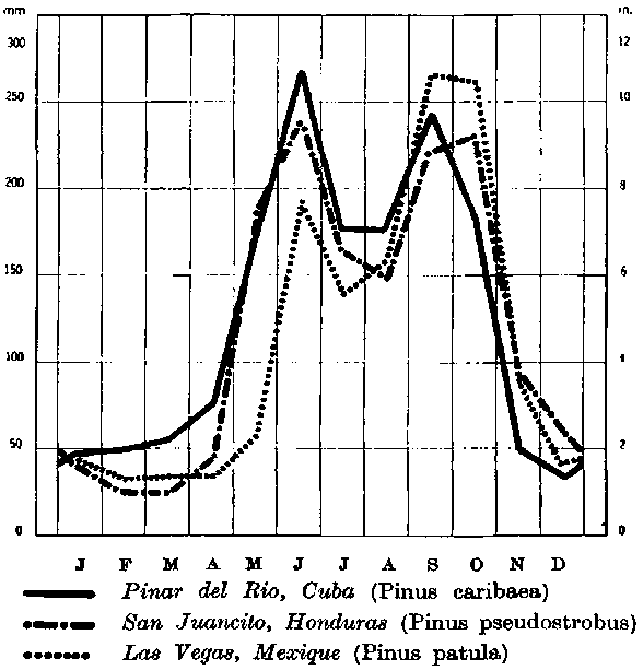
3. Espèces exigeantes en eau surtout en été et qui peuvent supporter un hiver nettement sec (fig. 3).
FIGURE 1. - Pluviosité moyenne mensuelle pour trois stations, correspondant à trois espèces différentes typiques des régions méditerranéennes ou à pluies d'hiver marquées.
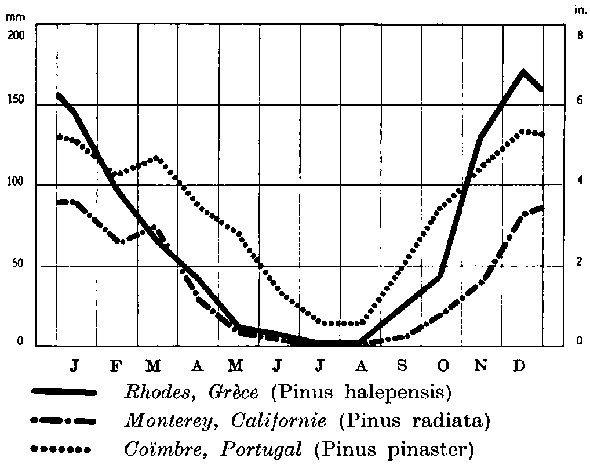
FIGURE 2. - Pluviosité moyenne mensuelle pour trois stations, correspondant à trois espèces différentes typiques des régions à régime de pluviosité uniformément répartie clans l'année.
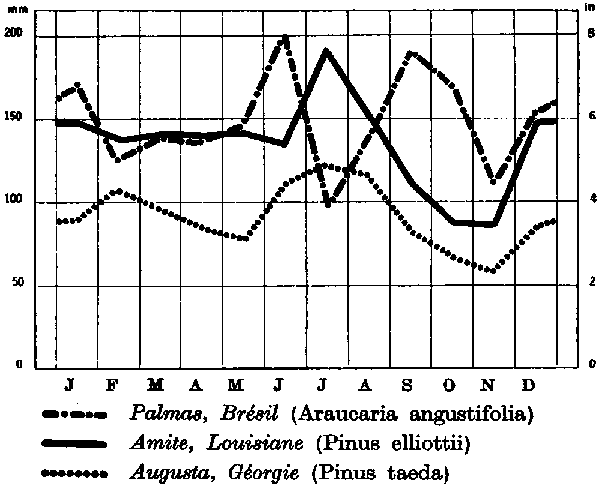
FIGURE 3. - Pluviosité moyenne annuelle pour trois stations, correspondant de trois espèces différentes typiques des régions de mousson ou de pluies d'été marquées.
Les espèces à pluviosité hivernale, ou du type méditerranéen, ont en règle générale deux périodes de repos une due au froid d'hiver et l'autre à la sécheresse d'été. La période de végétation des espèces qui exigent une pluviosité uniforme ou constante est limitée en général par les froids d'hiver. D'autre part, pour les conifères qui préfèrent les pluies d'été, type climat de mousson, la croissance est habituellement ralentie par la sécheresse et non par le froid, puisque dans les régions tropicales ou subtropicales, la variation. saisonnière de température est faible4.
4 Presque tous les conifères proviennent de régions soumises à l'un des trois régimes pluviaux mentionnés ci-dessus, mais il y a quelques exceptions. Par exemple, Araucaria angustifolia, dont l'habitat principal est le sud du Brésil et la région frontière avec l'Argentine, peut également vivre plus au nord dans de petits îlots montagneux des Etats de São Paulo et Minas Gerais avec des pluies d'été et des hivers assez humides. Une autre espèce, P. pinaster, qui exige normalement des pluies d'hiver présente une race adaptée à une pluviosité uniforme dans la partie nord-ouest de son aire, c'est-à-dire les Landes, en France. De même, P. elliottii, qui préfère :normalement une pluviosité bien répartie s'est adapté aux conditions de la Floride centrale, à la limite méridionale de son aire, où la pluie tombe en été. Il existe des cas limites dans les climats de transition. Une autre catégorie est représentée par les espèces de la Sierra Madre occidentale au Mexique, P. durangensis, P. engelmanni P. cooperi et P. arizonica, et aussi par les conifères dé la Chine du centre et du Sud, Cunninghamia lanceolata et P. massoniana dont la période de repos hivernal est due à une combinaison du froid et de la sécheresse.
Dans le nouvel habitat où l'on se propose d'introduire une espèce, il doit y avoir la même concordance entre le climat et la période de végétation de cette espèce que dans son milieu d'origine, car, s'il n'y a pas concordance entre le rythme végétatif et le climat, les chances du succès de l'introduction sont limitées. Les signes les plus nets d'incompatibilité entre les écologies sont: croissance rabougrie; port défectueux; mauvaise formation de la cime; feuillage partiellement desséché; floraison précoce; symptômes de nanisme; aspect souffreteux; sensibilité aux attaques des insectes et aux maladies; mort prématurée, etc.
On peut donner quelques exemples. Au pinetum de Placerville (Californie), dans un climat typiquement méditerranéen, aucun des pins du Mexique ou de l'Amérique centrale, originaires d'un climat de mousson, ne peut prospérer; on a essayé: P. patula, P. hondurensis, P. pseudostrobus, P. oocarpa, P. montezumae, P. strobus chiapensis, P. durangensis, P. michoacana, P. douglasiana et d'autres encore.
Au contraire, P. parryana, P. muricata et P. contorta latifolia poussent de façon satisfaisante; ils proviennent de la partie septentrionale de la Basse-Californie, seule région du Mexique à été sec et pluviosité hivernale, c'est-à-dire à climat analogue à celui de Placerville.
Dans la région centrale du Chili, soumise à un régime de pluies d'hiver et d'étés secs, on a introduit avec succès P. radiata originaire d'une bande côtière en Californie où le climat est semblable. Plus au sud, dans la région de Valdivia, dans les stations plus arrosées, Pseudotsuga menziesii réussit mieux, de même que dans la région montagneuse des Provinces argentines de Neuquén, Rio Negro et Chubut, à proximité de la frontière chilienne, tandis que dans les stations plus arides avoisinantes P. ponderosa, P. lambertiana, et P. contorta var. latifolia réussissent bien. Toutes ces espèces proviennent de l'ouest des Etats-Unis où le climat est semblable à celui des stations susmentionnées.
Dans l'Etat de Minas Gerais, au Brésil, en haute altitude, sous un régime de mousson avec une courte période de sécheresse d'hiver, P. radiata pousse presque sans arrêt et présente de longues branches minces, peu ramifiées, avec parfois l'extrémité sèche. (Jette anomalie, observée également par Lamprecht (1959) au Venezuela et García Díaz (Wadsworth, 1960) en Colombie, est probablement due au fait que le climat dans ces régions ne convient pas à cette espèce. Un autre cas d'incompatibilité climatique concerne une place d'expérience de P. elliottii (espèce qui préfère une pluviosité uniformément répartie) à Augustine (Belice) dans une région à climat de mousson avec des hivers chauds et secs. A six ans et demi, les plants avaient moins de 3 m de haut et étaient irrégulièrement branchus avec des pousses fortement lignifiées, parfois sèches à l'extrémité. Comme il n'y a aucune raison d'attribuer cette anomalie au sol, on pense que l'espèce souffre de la sécheresse qui dure de janvier à mai et de l'absence des conditions hivernales auxquelles elle est accoutumée.
FIGURE 4. - Graphique montrant la relation entre le régime pluvial convenant à certains conifères et les chances du succès de l'introduction dans d'autres pays.

Conifères méditerranéens
Des expériences ont montré que les espèces autochtones dans les régions à pluies d'hiver peuvent être introduites avec succès dans des régions analogues et, jusqu'à un certain point, dans celles où la pluviosité est uniformément répartie, pourvu que les autres conditions écologiques soient favorables5; mais leur introduction dans les régions de mousson est un échec complet (fig. 4).
5 Outre l'importance de la pluviosité, de son régime, et de celui des températures, les conditions de sol représentent aussi un facteur décisif pour l'introduction d'une espèce en dehors de son aire d'origine.
P. radiata, originaire d'une région à climat de type méditerranéen avec hiver frais, est un bon indicateur. A Concepción (Chili), où les conditions sont analogues et à certains égards meilleures que dans l'aire naturelle, ce pin pousse extraordinairement vite, avec une forme excellente et il est très longévif. Il pousse aussi assez bien sur la côte sud de l'Uruguay et dans les zones isolées de la région côtière du sud-est de la Province de Buenos Aires en Argentine, où la pluviosité est uniforme et modérée, les étés frais et les sols légers, profonds et bien drainés. Par contre, dans la région du Nord et de l'Ouest, à pluviosité bien répartie mais à étés plus chauds et plus humides, où les sols sont superficiels et argileux, avec drainage médiocre ou nul, cette espèce est moins brillante et sa croissance plus irrégulière, avec un tronc parfois excessivement branchu, fourchu ou flexueux (Barret & Garbosky, 1960). Plus au nord, les conditions deviennent progressivement encore plus défavorables et on constate une croissance lente, des déformations et des attaques sévères de Diplodia. P. radiata est incapable de survivre dans la Province de Misiones (Argentine), en zone subtropicale avec pluies abondantes et uniformément réparties; il y meurt en quelques années. Cela se produit aussi dans les Etats de Sao Paulo (Brésil), et dans la Province de Tucumán (Argentine), dans la région à pluies d'été bien marquées. Dans le dernier cas, les facteurs défavorables, outre l'été humide, sont probablement la température élevée et le régime photopériodique différent.
Ce résultat a été confirmé par des expériences entreprises en Afrique du Sud (Poynton, 1957). Les tentatives faites pour étendre les plantations de P. radiata du sud-ouest de la Province du Cap - pluies d'hiver ou uniformément réparties - vers le nord-est, c'est-à-dire l'Etat libre d'Orange, le Natal et le Transvaal, avec des étés très humides, ont échoué; en raison des conditions écologiques défavorables, les arbres ont été sévèrement attaqués par Diplodia et la plantation a dû être abandonnée.
P. pinaster, autre espèce du type méditerranéen, a été introduite avec succès dans la région côtière du sud de l'Uruguay, ou les pluies sont modérées et bien réparties. Il ne réussit pas bien cependant dans le nord de l'Argentine ou le sud du Brésil.
Conifères des régions à pluviosité uniforme
L'expérience a montré que les espèces originaires des régions où la pluviosité est uniformément répartie tout au long de l'année peuvent être introduites avec succès dans des régions analogues et aussi, sous certaines conditions., dans les régions à pluies saisonnières avec maximum d'été, mais qu'elles s'adaptent rarement aux régions à pluies d'hiver (fig. 4).
Araucaria angustifolia, espèce sud-américaine de basse montagne demandant une pluviosité uniforme, a été récemment essayée en dehors de son aire naturelle. Dans les régions avec la même pluviosité et le même régime, quoique avec des températures un peu plus élevées, par exemple dans le nord de la Province de Misiones, les résultats sont excellents. Mais avec des pluies intermittentes et des hivers secs, les arbres poussent plus lentement (São Paulo) ou restent rabougris (Tucumán).
On cultive également avec succès P. elliottii en Argentine depuis le delta du Paraná et le nord de la Province de Buenos Aires, jusqu'à, la Province de Misiones; dans Le centre et le nord de l'Uruguay au Brésil, clans le nord-est de l'Etat de Paraná et le sud-ouest de celui de São Paulo, toutes régions où la pluviosité est uniformément répartie. Au contraire, dans le centre et le nord de l'Etat de São Paulo, avec des pluies saisonnières et un hiver nettement sec, la croissance est lente et irrégulière et il est peu probable que cette espèce donne satisfaction dans de telles conditions.
En Afrique du Sud (Poynton, 1957), P. elliottii a été cultivé avec succès dans diverses conditions écologiques, sur la côte sud de la Province du Cap à pluviosité uniforme, au Natal où le régime pluvial est intermédiaire entre les précipitations continues et intermittentes, et au Transvaal où les pluies sont saisonnières. Les meilleurs résultats sont obtenus au Transvaal oriental, dans les régions où les pluies d'été sont abondantes, le climat adouci par l'altitude, les températures moyennes de 18° C dans le mois le plus chaud et 10° C dans le mois le plus froid. Ici, le sécheresse d'hiver est tempérée par la température fraîche et les brouillards qui entretiennent un état hygrométrique élevé tout au long de l'année.
En Australie (Bednall, 1957; New South Wales Forestry Commission, 1957; Rogers, 1957), P. elliottii est cultivé sur la côte orientale depuis la Nouvelle Galles du Sud (pluviosité uniforme) jusqu'au centre de la côte du Queensland, au nord de Rockhampton (pluies d'été); cependant la principale région de culture se trouve dans le sud du Queensland près de Brisbane, où cette espèce montre la croissance la plus forte dans les régions à pluies abondantes et à régime intermédiaire entre uniforme et saisonnier. Elle n'est pas plantée dans les régions à climat méditerranéen du sud de l'Australie, sa croissance y étant très lente et bien inférieure à celle de P. radiata.
Conifères des climats de mousson
Les expériences ont montré que les espèces originaires des régions à fortes pluies d'été peuvent être introduites avec succès dans les régions à climat analogue et parfois aussi dans celles où la pluviosité est uniformément répartie, mais qu'elles réussissent rarement dans les zones à pluies d'hiver (fig. 4).
Dans le centre et le nord de l'Etat de Sao Paulo, avec des pluies d'été, les espèces qui semblent les plus intéressantes sont: P. hondurensis, P. caribaea, P. insularis dans la zone subtropicale et à une altitude de 500 à 700 m, et Cunninghamia lanceolata, P. patula, Cupressus lusitanica, Cryptomeria japonica à une altitude de 700 à 2 000 m. Toutes ces espèces sont indigènes dans des régions à climat de mousson.
Dans les régions à pluviosité uniforme et abondante, comme la Province de Misiones (Argentine), la moyenne annuelle étant 1 700 mm, P. hondurensis et P. caribaea se révèlent très intéressants et ont une croissance bien supérieure à P. elliottii et Araucaria angustifolia; cependant, P. hondurensis y montre un port beaucoup plus variable que dans l'Etat de Sao Paulo, au Brésil, bien que les mêmes graines eussent été employées dans les deux cas. Il semble que cette irrégularité dans la forme soit due, dans la Province de Misiones, à l'absence d'une période sèche d'hiver que l'espèce trouve aussi bien dans l'Etat de Sao Paulo que dans sa région d'origine: cela détermine une croissance presque ininterrompue. Ce dernier trait est un désavantage en ce sens que les froids d'hiver peuvent endommager les jeunes feuilles ou pousses qui n'ont pas eu le temps de s'aoûter. Plus au sud, dans les Provinces de Santa Fé et Buenos Aires, avec une pluviosité modérée et uniformément répartie (900 à 1 000 mm) et un climat tempéré, P. hondurensis et P. caribaea sont inférieurs à P. elliottii et par d'autres signes témoignent d'une mauvaise adaptation à ce milieu.
Exceptionnellement, des conifères de climat de mousson peuvent être introduits avec succès dans des régions à pluies d'hiver, par exemple pour Cupressus lusitanica à Bussaco (Portugal). Il faut noter cependant que le climat de Bussaco n'est pas méditerranéen, mais de transition avec le régime à répartition uniforme, ce qui se manifeste par la brièveté de la période sèche en été. Les autres conditions favorables sont la forte pluviosité annuelle (1500 mm) et des températures analogues à celles des régions montagneuses du Guatemala et du Mexique où cette espèce est indigène.
Le tableau 1 donne la pluviosité moyenne annuelle pour les régions d'origine des différents conifères. Lorsqu'ils sont introduits dans une nouvelle station, ils doivent avoir une hauteur de pluie au moins égale a celle qu'ils reçoivent dans leur pays d'origine. P. radiata, par exemple, a le plus fort accroissement au Chili et en Nouvelle-Zélande, dans des régions où la pluviosité annuelle de 1 100 à 1700 mm est bien supérieure à celle de la région d'origine.
Parfois, une espèce poussera dans des régions à pluviosité plus faible, mais, dans ce cas, le déficit de pluie est compensé par des brouillards côtiers ou de montagne, une température moyenne plus froide ou un sol plus profond. Par exemple, P. elliottii est cultivé au Queensland (Rogers, 1957) dans des zones assez basses où la pluviosité annuelle varie entre 1300 à 1550 mm, comme dans son aire d'origine. A Passchendaele cependant, dans le même Etat, où il est planté à une altitude de 900 m, la pluviosité annuelle est à peine de 850 mm; dans ce cas, le déficit de pluviosité est compensé par la température plus fraîche due à l'altitude.
TABLEAU 1. - DONNÉES CLIMATIQUES POUR CERTAINS CONIFÈRES DANS LEUR AIRE NATURELLE
|
Espèce |
Température moyenne du mois le plus froid (°C) |
Température moyenne du mois le plus chaud (°C) |
Température minimum absolue1 (°C) |
Précipitation moyenne annuelle (°C) |
|
Araucaria angustifolia |
8-14 |
18-23 |
- 8 |
1 250-2 200 |
|
A. cunninghamii |
9-16 |
20-26 |
- 6,5 |
1 000-1 500 |
|
Cunninghamia lanceolata |
0-15 |
26-29 |
- 20 |
900 1 800 |
|
Cuppressus lusitanica |
8-12 |
14-17 |
- 8 |
1 100 2 200 |
|
Pinus ayacahuite |
4 10 |
12-18 |
- 26 |
800 2 400 |
|
P. canariensis |
2-12 |
16-20 |
- 8 |
350- 600 |
|
P. caribaea |
22 |
28 |
4 |
1 000 1 600 |
|
P. cubensis |
18-22 |
24-28 |
1 |
1 150-1 500 |
|
P. douglasiana |
10 17 |
15-22 |
- 4 |
1 000-2 600 |
|
P. durangensis |
4-7 |
15-18 |
- 25 |
600-1 100 |
|
P. elliottii |
9-17 |
26-28 |
- 20 |
1 100-1 650 |
|
P. elliottii densa |
15-21 |
26-28 |
- 8 |
1 000-1 500 |
|
P. engelmanni |
5-9 |
17-19 |
- 20 |
600-1 000 |
|
P. halepensis |
2-10 |
20-28 |
- 10 |
380- 900 |
|
P. hondurensis |
16-25 |
22-28 |
5 |
1 000-3 500 |
|
P. insularis |
12-20 |
16-24 |
4 |
1 400-2 500 |
|
P. khasya |
13-22 |
18-25 |
- 5 |
1 200-2 000 |
|
P. massoniana |
2-15 |
26-29 |
- 18 |
1 000-1 500 |
|
P. merkusii |
15-25 |
20-28 |
0 |
1 200-2 200 |
|
P. michoacana |
13-20 |
18-26 |
- 2 |
800-1 500 |
|
P. montezumae |
8-13 |
11-19 |
- 7 |
900-2 200 |
|
P. occidentalis |
13-22 |
17-25 |
- 1 |
1 000-1 600 |
|
P. oocarpa |
11-19 |
16 24 |
- 3 |
700-1 600 |
|
P. palustris |
7-17 |
26-28 |
- 20 |
1 160-1 600 |
|
P. patula |
7-11 |
12-17 |
- 10 |
1 200-2 000 |
|
P. pinaster |
2-9 |
18-25 |
- 10 |
750-1 250 |
|
P. pseudostrobus |
7-16 |
12-21 |
- 6 |
1 100-3 000 |
|
P. radiata |
10-11 |
16,5 18 |
-_ 7 |
420- 700 |
|
P. roxburghii |
5-14 |
16-25 |
- 9 |
800-2 000 |
|
P. strobus chiapensis |
11-20 |
18-24 |
- 3 |
1 200-5 800 |
|
P. taeda |
2-15 |
24-26 |
- 23 |
920-1 550 |
¹ Les chiffres de cette colonne se réfèrent aux minimums absolus relevés pour chaque espèce dans son aire naturelle. Par exemple, le chiffre minimum pour Pinus ayacahuite est - 26 °C à Chihuahua (Mexique) où vit la variété septentrionale. Mais la variété méridionale pousse à Chiapas (Mexique) et au Guatemala dans des régions où le minimum absolu est - 7° G. Une telle disparité est également constatée pour d'autres espèces à large distribution, entre autres, P. taeda, Cunninghamia lanceolata.
Dans les régions où il tombe moins de pluie que dans le pays d'origine et où les autres facteurs ne sont pas favorables, il est douteux que les conifères réussissent. Dans cet :Etat, les diverses espèces sont souvent rabougries, avec un port conique, très branchu, et sont plus sensibles aux.
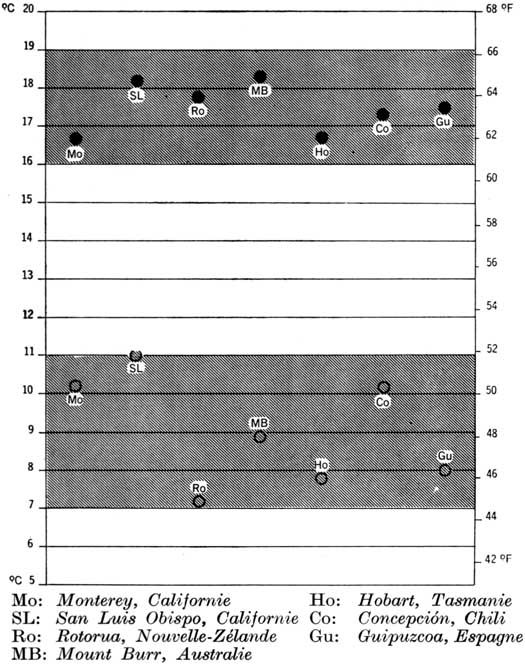
Les valeurs qui représentent le mieux les exigences thermiques de chaque espèce et peuvent être employées comme référence sont les températures moyennes annuelles, du mois le plus froid, du mois le plus chaud, et minimums absolues (tableau 1). La figure 12 montre la gamme des températures moyennes annuelles qui conviennent pour certains conifères dans leur habitat naturel. Evidemment, les espèces à large répartition géographique, comme P. taeda, P. pseudostrobus et P. merkusii, admettent une gamme de température plus large, en fonction des variations en latitude et en altitude 6. Au contraire, les espèces confinées à des stations particulières, comme P. caribaea et P. radiata, admettent une gamme de température limitée, bien qu'elles puissent s'adapter á des limites plus larges, puisqu'elles peuvent être cultivées avec succès dans des régions un peu plus froides ou plus chaudes, pourvu que les différences de température ne soient pas trop grandes. Par exemple, Araucaria angustifolia est adapté dans son habitat naturel à une température moyenne annuelle de 10° C, mais peut être cultivé avec succès dans la Province de Misiones, en dehors de son aire naturelle, dans des régions où la température atteint 20° C; par contre, il est très peu probable que l'on puisse y installer P. patula en raison de ses exigences en température très différentes.
6 Comme l'ont montré expérimentalement Wakeley (1944) pour P. taeda aux Etats-Unis et Rycroft et Wicht (1947) pour P. pinaster en Afrique du Sud, il est à peu près certain que d'autres espèces à aire étendue sont formées de races géographiques ou écotypes avec différentes vitesses de croissance. Le choix de la race la meilleure pour chaque région est un autre problème que seule l'expérimentation pourra résoudre.
D'autres valeurs de température importantes sont la température moyenne du mois le plus froid et la température moyenne du mois le plus chaud, utilisées par Swain (1938) pour son index climatique.
En prenant ces valeurs comme base, les expériences actuelles montrent; que les espèces à été chaud peuvent être transportées avec succès dans des régions à été plus froid, mais le succès est moins assuré en transférant un conifère à été froid dans une région à été chaud. Par exemple, P. elliottii est indigène dans une région où la température moyenne du mois le plus chaud est située entre 26° C et 28° a, mais il peut être introduit avec succès dans des régions où la température moyenne du mois le plus chaud est seulement 19° C comme à Graskop, dans le Transvaal oriental; tandis que P. patula, espèce de régions froides où la température moyenne du mois le plus chaud est situé entre 12° C et 16° C, ne peut réussir sur la basse côte du Zoulouland à étés chauds et avec une température moyenne de janvier de 24° C à 25° C. La même espèce pousse mal à Gourie, dans les montagnes du centre de la Jamaique (Lamb et al.; 1960), où la température moyenne d'été varie entre 20° C et 23° C.
Un autre facteur à ne pas négliger est la différence entre les extrêmes des températures saisonnières. P. massoniana et P. elliottii sont indigènes dans des régions à hivers assez froids et ii étés chauds, la différence entre la température moyenne du mois le plus froid et celle du mois le plus chaud étant 28° C pour La première espèce et 18°C pour la seconde. Aucune de ces espèces ne réussit en Malaisie ou aux îles Fidji (Barnard & Beveridge, 1957; Cottle, 1957) sous un climat tropical humide, avec un écart de moins de 5° Ci entre les extrêmes de températures moyennes mensuelles. Des espèces tropicales, comme P. hondurensis et P. caribaea, conviendraient mieux pour ces régions.
La résistance au froid est un autre facteur à prendre en considération, car il peut limiter OU empêcher la culture d'une espèce dans une région donnée. Le tableau 1 donne les températures minimums pour différents habitats. Il faut noter que, contrairement, aux eucalyptus qui dans les pays où ils sont introduits, sont aussi sensibles au froid que dans leur aire d'origine, les conifères sont en général plus plastiques. P. hondurensis et P. caribaea, par exemple, supportent dans la Province de Misiones, des températures dé - 4° C, plus basses que celles de leur pays d'origine où le minimum de + 5° C est rarement atteint. La résistance au froid de chaque espèce dépend de divers facteurs tels que l'altitude et l'exposition de la station, les gelées, le brouillard, l'humidité du sol et les vents dominants. Elle dépend également de l'adaptation de l'espèce aux conditions écologiques de la station où elle pousse. Par exemple, au Tanganyika (Parry & Willian, 1957), sur le plateau d'Elton à 2 700 m, P. patula est aussi résistant au froid que dans son habitat normal et supportera une température de - 11° C, alors que dans la Province de Misiones, il est endommagé par un froid de - 5° C.
Les exigences et les tolérances climatiques d'un certain nombre de conifères ont été définies en fonction des conditions écologiques de leur aire naturelle et des stations où ils ont été introduits avec des résultats variant de la pleine réussite à l'échec total.
Le succès de l'introduction d'une espèce dépend surtout du degré de similitude entre le climat de l'aire naturelle et celui des stations d'introduction. Les données de base pour déterminer cette similitude sont: la répartition saisonnière des pluies; la pluviosité totale annuelle; les températures moyennes du mois le plus froid et du mois le plus chaud et les minimums absolus. Les conifères peuvent être répartis, suivant le régime des pluies de la région d'origine, en trois groupes correspondant aux régions à pluies d'hiver, à pluies bien réparties et à pluies d'été. Il faut noter que chaque groupe, suivant son origine, ne peut, sauf exceptions, réussir dans les régions correspondant au régime pluvial des autres groupes.
La comparaison de ces données climatiques fournit une base solide pour savoir si une espèce est susceptible de réussir dans une région donnée. Par exemple, les conditions climatiques idéales pour quelques espèces sont les suivantes:
Pinus radiata
pluviosité annuelle de 1100 à 1 700 mm tombant surtout dans la moitié la plus froide de l'année; été sec;
température moyenne du mois le plus froid: 7° C à 11° C;
température moyenne du mois le plus chaud: 16,5° à 18,5° C;
minimums absolus, si possible pas inférieurs à - 7° C.
Araucaria angustifolia
pluviosité de (1 250 à 2 200 mm), uniformément répartie tout au long de l'année; c'est-à-dire sans saison sèche;
température moyenne du mois le plus froid: 8° C à 14° C;
température moyenne du mois le plus chaud: 18° a à 23° C;
minimums absolus, pas inférieurs à - 12° C.
Pinus hondurensis
pluviosité entre 1 000 et 3 500 mm, surtout dans les six mois les plus chauds;
hiver sec;
16° C à 25° C;
22° C à 28° C;
minimums absolus, pas inférieurs à - 8° C.
Avec des renseignements plus complets, il sera possible de préciser pour chaque espèce et, le cas échéant, ses écotypes, les conditions climatiques optimums et aussi les limites entre lesquelles elles peuvent être employées pour le reboisement.
BARNARD, R. C. & BEVERIDGE, A. E. 1957. Exotic trees in the Federation of Malaya . Seventh British Common wealth Forestry Conference, Canberra, Australia.
BARRET, W. G. & GARBOSKY, A. J. 1960. Efectos del suelo en el crecimiento de Pinus radiata en el norte de la provincia de Buenos Aires. Rev. Invest. for. B. Aires, 2: 67-88.
BEDNALL, B. H. 1957. Exotic forest trees in South Australia. Adelaide, Government Printer.
CHAMPION, H. & BRASNETT, N. V. 1960. Le choix des essences forestières pour les boisements et reboisements. Rome. Collection FAO. Mise en valeur des forêts, N° 13.
Conifers of Latin America used in forestry in the United States: official national statement. 1961. FAO Latin American Conifer Seminar, Mexico, D.F.
COTTLE, G. W. 1957. Exotic forest trees in the British Commonwealth: Fiji. Fiji, Forestry Department, Seventh British Commonwealth Forestry Conference, Canberra, Australia.
Cozzo, D. 1961. Informaciones sobre el crecimiento en la Argentina de varias especies del género Pinus. Rev. for. argent., 5: 3-20.
DE PHILIPPIS, A. 1951. Ecologie forestière et phytoclimatologie. Unasylva, 5: 11-15.
DYSON, W. G. 1961. The growth of Latin-American conifers in the Kenya highlands. Document, FAO Latin American Conifer Seminar, Mexico, D.F.
FLINTA, C. M. 1960. Prácticas de plantación forestal en América Latina. Rome. Collection FAO. Mise en valeur des forêts, N° 15.
FOOT GUIMÃRES, R. 1958. Plantío expérimental de coníferas no interior do Estado de São Paulo. An. bras. Econ. flor., Inst. nac. Pinho, 10: 191-207.
GIORDANO, G. 1956. The Mediterranean region. Dans Haden-Guest, S. A world geography of forest resources. New York, Ronald Press. American Geographical Society Special Publication N° 33.
GOLFARI, L. 1959. Notas sobre el cultivo de pinos y de otras coniferas en la Argentina. Rev. argent. Agron., 26 (3, 4): 65-99.
GOOR, A. Y. 1957. Les méthodes de plantations forestières en zones arides. Rome. Collection FAO. Mise en valeur des forêts, N° 6.
HILEY, W. E. 1959. Conifers: South African methods of cultivation. London, Faber and Faber.
HOLDRIDGE, L. R. 1947. Determination of world plant formation from simple climatic data. Science, 106: 367-368.
HUECK, K. 1953. Distribução e habitat natural do pinheiro do Paraná, Araucaria angustifolia. São Paulo, Universidade de São Paulo, Faculdade de Filosofia, Ciéncias e Letras. Boletín N° 156.
JACOBS, M. R. 1961. Eucalyptus as an exotic. Document Deuxième conférence mondiale de l'eucalyptus, São Paulo, Brésil.
KÖPPEN, W. 1931. Climatología. México, D.F., Fondo Cultura Econórnica.
LAMB, F. B., BRISCOE, C. B. & ENGLERTH, G. H:. 1960. Recent observations on forestry in tropical America. Caribbean Forester, 21 (1, 2): 46-59.
LAMPRECHT, H. & FINOL, H. 1959. Programma de estudios sobre coníferas exóticas en los Andes venezolanos. Mérida Venezuela, Instituto Forestal Latinoamericano Boletín N° 4.
LINDSAY, A. D. 1932. Report on Monterey pine (Pinus radiata D. Don) in its native habitat. Canberra, Commonwealth Forestry Bureau. Bulletin N° 10.
LITTLE, E. L. & DORMAN, K. W. 1954. Slash pine (Pinus elliottii) including South Florida slash pine, nomenclature and description. Asheville, North Carolina, Southeastern Forest Experiment Station. Station Paper N° 36.
LOOCK, E. E. M. 1950. The pines of Mexico and British honduras. Pretoria, Department of Forestry. Bulletin N° 35.
MARTÍNEZ, M. 1948. Los pinos mexicanos. México, D.F. Edic. Botas.
McLAREN Dow, T. 1961. Introductory research in silviculture in Latin American pines in Northern Rhodesia. Document, FAO Latin American Conifer Seminar Mexico, D.F.
MÉTRO, A. 1954. Les eucalyptus dans le reboisement. Rome. Collection FAO. Etudes des forêts et des produits forestiers, N° 11.
NEW SOUTH WALES. FORESTRY COMMISSION. The principal exotic forest trees in New South Wales. Sydney, Forestry Commission of New South Wales. Seventh British Commonwealth Forestry Conference, Canberra, Australia.
PARRY, M. S. 1956. Les méthodes de plantations forestières en Afrique tropicale. Rome. Collection FAO. Mise en valeur des forêts, N° 8.
PARRY, M. S. & WILLIAN, R. L. 1957. Exotic forest trees in the British Commonwealth: Tanganyika. Seventh British Commonwealth Forestry Conference, Canberra, Australia.
POYNTON, R.J. 1957. Notes on exotic forest trees in South Africa. Pretoria, Government Printer. Seventh British Commonwealth Forestry Conference; Canberra, Australia.
RYCROFT, H. B. & WICHT, C. L. 1947. Field trials of geographical races of Pinus pinaster in South Africa. Pretoria,. Department of Forestry.
ROGERS, L,. J. 1957. Exotic forest trees in Queensland. Queensland, Forestry Department. Seventh British Commonwealth Forestry Conference; Canberra, Australia.
ROYAUME-UNI. METEOROLOGICAL OFFICIE. 1958. Tables of temperature, relative humidity and precipitation for the world. Parts I, II, III, IV, V, VI. London,
SCHWERDFEGER, F. 1953. Los pinos de Guatemala, Roma, Informe FAO/PAAT N° 202.
SCOTT, C. W. 1960. Pinus radiata, essences exotique. Unasylva, 14: 6-16.
SMITHSONIAN INSTITUTION. 1944-47, World weather records. Washington, D.C. Smithsonian Miscellaneous Collection, Vols. 79, 90, 105.
SWAIN, E. H. F. 1938. Climatic index. Sydney, Forestry Commission of New South Wales.
THORNTHWAITE, C. W. & HARE F. K. 1955. Classification climatologique et foresterie. Unasylva, 9: 55-63.
TROUP, R. S. 1932. Exotic forest trees in the British Empire. Oxford, Clarendon Press.
U AUNG DIN. 1958. L'utilisation des pins dans les régions tropicales. Unasylva, 12: 121-133.
VIVÓ, J. A. & GOMEZ, J. C. 1946. Climatología de México. México, D.F., Instituto Pan-Americano de Geografía y Historia. Publicación N° 19.
WADSWORTH, F. H. 1960. Records of forest plantation growth in Mexico, the West Indies, and Central and South America. Caribbean Forester, 2:1 (Supp.), '270 p.
WAKELEY, P. C. 1944. Geographie source of loblolly pine seed. J. For., 42: 23-32.
WHITE, K. J. 1960. Latin American conifers in Australia. Document, FAO Latin American Conifer Seminar, Mexico, D.F.
PROCHAINES RÉUNIONS FAO
|
Consultation mondiale sur le contre-plaqué et les panneaux |
Salzbourg (Autriche), juillet |
|
Consultation mondiale en matière de génétique forestière et d'amélioration des semences |
Stockholm (Suède), 23-30 août |
|
Cinquième Conférence de la technologie du bois |
Madison, Wisc. (Etats-Unis), 16-17 septembre |
|
Commission des forêts pour l'Europe |
Genève, septembre |
|
Comité du bois de la CEE |
Genève, 7-11 octobre |
|
Conférence de la FAO - Comité des forêts |
Rome, 31 octobre- 15 novembre |
![]()
![]()
![]()
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}