![]()
![]()
![]()
K.D. Singh
K.D. Singh es especialista en inventarios e informática del Servicio Forestal de la India.
El autor analiza en detalle tres tipos de patrones de variación espacial observados en la selva tropical. Dichos patrones revelan que el aparente caos de la vegetación en realidad tiene una estructura orgánica.
El presente artículo da a conocer algunos patrones comunes de variación observados en la selva tropical, principalmente en base a los actuales conocimientos, pero agregando además algunos materiales inéditos. Primero trata de los macropatrones, indicando su significación para la estratificación. Después, trata de los micropatrones y su importancia para la determinación de las dimensiones y de la forma de la unidad de muestra. Por último, trata de los patrones de variación, a escala intermedia, y de su función en la determinación del plan de muestreo, o sea, trazado en racimo o disperso, al azar o sistemático.
La selva tropical es el clímax natural de la vegetación, en la zona que se extiende hasta los grados 8-10, a uno y otro lado del ecuador. Los factores climáticos esenciales para su desarrollo son: una temperatura elevada uniforme (alrededor de 24-26°C) y una larga época de lluvias, de alrededor de 10 a 12 meses de duración.
Las principales zonas de selvas tropicales son:
Centro y Sudamérica - La cuenca del Amazonas (Brasil), que se extiende por el oeste hasta los primeros contrafuertes de la cordillera de los Andes, por el este hasta las Guayanas, por el sur hasta el Gran Chaco y por el norte a todo lo largo de la parte oriental de la América Central.
Africa - La cuenca del Congo (principalmente el Zaire y el Congo), Gabón y Camerún. Se extiende, por el oeste hasta el interior de Nigeria, Ghana, Costa de Marfil, Liberia y Guinea y, por el este, hasta Uganda.
Asia - Principalmente el sudeste de Asia, abarcando la península malaya, las islas de Sumatra, Borneo, Célebes y Nueva Guinea. Se extiende también, en parte, a la India, Sri Lanka, Birmania, Tailandia y Filipinas.
A pesar de que las selvas tropicales se encuentran en zonas muy distantes, los estudios ecológicos dan pruebas de que su semejanza es notable. Según Richards (1971), se componen de sinusias o comunidades de árboles semejantes, con características homogéneas y disposición espacial muy similar. Las especies de las correspondientes sinusias de diferentes regiones geográficas se asemejan, a su vez, por su forma biológica y también, en alto grado, por su fisonomía. El patrón fundamental de la estructura es, por lo tanto, igual en todas las zonas que cubre la selva tropical.
Estos descubrimientos demuestran que seria interesante investigar patrones comunes de variación que podrían ser significativos desde el punto de vista del diseño de los inventarios. Es claro que estos conocimientos pueden facilitar el intercambio de información inventarial entre regiones y acrecentar la eficacia de los diseños de inventarios.
Para planificar un inventario son de interés los patrones de variación a muchas escalas. Para tomar decisiones con respecto a la estratificación se trata de establecer macropatrones, o sea, cambios en el uso de la tierra, tipos de bosques y clases maderables. Para determinar las dimensiones y las formas de la unidad de muestreo es más critica la microvariación. Para elaborar un plan de muestreo hay que tener algunos conocimientos sobre el patrón de variación a escala intermedia.
Los párrafos que siguen contienen una breve reseña de los patrones de variación, a tres escalas.
Las causas de diferenciación dentro de las selvas tropicales se pueden dividir en dos grandes grupos: naturales y silvícolas (o culturales).
Entre los factores naturales, el clima es relativamente uniforme. El suelo y la topografía son, al parecer, las principales causas de variación. Algunos tipos comunes son:
- El clima de la selva tropical (generalmente en suelos de buen drenaje y altitud inferior a 500-800 m).
- Los manglares.
- La selva pantanosa.
- La selva que se inunda periódicamente.
- La selva ripiaría.
- La selva montañosa (que suele encontrarse a más de 500-800 m de altura)
- La selva de los podsoles tropicales.
- La selva de los arenales de la costa.
Los factores culturales (influencias positivas o negativas del hombre) son causa de ulteriores cambios de la vegetación de las tropicales naturales, que se pueden dividir grosso modo en los siguientes tipos:
- La selva tropical primaria (donde la influencia del hombre es menor).
- La selva explotada.
- La selva secundaria (resultante de los cultivos migratorios abandonados).
- La vegetación mixta forestal-pastoral.
- El bosque manejado (natural y artificial).
- Otras clases de usos no forestales de la tierra.
Hay claras diferencias, entre los tipos mencionados, en cuanto a los pisos, las especies, el número de árboles y el volumen por unidad de superficie. Cabe observar también que, mientras mayor es la limitación del crecimiento debida a las condiciones, mayor es la tendencia a la uniformidad.
Los estudios realizados revelan que es posible identificar, en su mayoría, los macropatrones, tanto en los bosques naturales como en los artificiales, empleando aerofotografías convencionales, a una escala de 1/20000 o menos.
Como la selva tropical primaria ocupa una superficie relativamente grande, en comparación con los demás tipos de bosques, se han hecho varias investigaciones para explorar las posibilidades de una mayor estratificación. Nyyssönen (1961), Loetsch et al. (1973) han reseñado tales estudios. Al parecer, es relativamente difícil distinguir un mayor número de tipos ecológicos empleando aerofotografías, porque la vegetación que cubre el terreno en la selva tropical se ve muy uniforme en grandes extensiones, sin cambios ni deslindes.
CUADRO 1. - VENTAJAS DE LA ESTRATIFICACIÓN 1
|
Método de muestreo |
Estimado de precisión de las regiones del reconocimiento (±) |
|||||||
|
1 |
2 |
3 |
4 |
5 |
6 |
7 |
8 |
|
|
Porcentaje |
||||||||
|
No estratificado |
6,7 |
8,1 |
9,6 |
6,6 |
7,2 |
7,9 |
10,4 |
10,0 |
|
Estratificado (post) |
4,5 |
6,8 |
7,6 |
4,0 |
3,7 |
4,2 |
5,4 |
5,6 |
1 Carácter: Estimado de la precisión del parámetro del volumen industrial neto de todas las especies de 18 pulgadas (45 cm) o más de diámetro (nivel de confianza, 95 por ciento).
Por otra parte, se ha descubierto la posibilidad de delinear groseramente clases según el volumen y el área basal, valiéndose de la correlación que hay entre la densidad de las copas de los árboles dominantes y el volumen bruto por unidad de superficie de todas las especies. Un análisis piloto, realizado por el autor en un reconocimiento hecho en Laos en 1969, basándose en datos de 185 parcelas, reveló que el cuadrado del coeficiente de correlación múltiple (R2) era de 0,64. Se observó también una estrecha relación. en el reconocimiento del valle del Amazonas, realizado por la FAO.
1. RACIMO DE NUEVE PUNTOS. RECONOCIMIENTO DEL SARAWAK

CUADRO 2. - ANÁLISIS DE LA VARIANZA DEL CARÁCTER VOLUMEN BRUTO/ACRE1 DE TODAS LAS ESPECIES DE 18 PULGADAS (45 CM) O MÁS DE DIÁMETRO
|
Fuente de variación |
Grados de libertad |
Suma de los cuadrados |
Promedio de la suma de los cuadrados |
Componentes de la varianza |
|
Entre muestras |
81 |
431448800 |
5326528,3 |
s b2 = 474223,86 |
|
Dentro de cada muestra |
656 |
694383300 |
1058511,1 |
s w2 = 1058511,10 |
1 Media: 2031 pies3/acre (142 m3/ha).
CUADRO 3. - EFECTO DE LAS DIMENSIONES Y DE LA FORMA DE LA PARCELA SOBRE EL COEFICIENTE DE VARIACIÓN DEL VOLUMEN TOTAL DE TODAS LAS ESPECIES
|
Tipo de bosque |
Volumen medio |
Estimado del coeficiente de variación |
Número de muestras |
|||
|
Parcela cuadrada |
Parcela rectangular |
|||||
|
03 ha |
0 6 ha |
0,3 ha 10X300 m |
0,6 ha 20X300 m |
|||
|
m3/ha |
...Porcentaje... |
|||||
|
Selva tropical |
393 |
98 |
82 |
75 |
48 |
14 |
|
Bosque deciduo |
177 |
63 |
60 |
60 |
58 |
25 |
FUENTE: Informe FAO, Forest resources survey, India, 1970.
CUADRO 4. - MEDIA Y COEFICIENTE DE VARIACIÓN DE ÁREA BASAL/HA, A NIVEL DE GRUPOS Y DE ESPECIES, EN ÁRBOLES DE 18 PULGADAS (45 CM) O MÁS DE DIÁMETRO (83 MUESTRAS. UNIDAD 1)
|
Grupos/especies |
Media área basal |
Coeficiente de variación |
|
...Metros2/ha... |
...Porcentaje... |
|
|
De todas las especies |
14,70 |
27 |
|
Dipterocarpáceos |
9,89 |
32 |
|
Merantis roja |
4,10 |
40 |
|
Shorea parvifolia |
2,46 |
111 |
|
Shorea sagittata |
1,52 |
122 |
|
Shorea rubro |
0,95 |
142 |
|
Shorea macroptera |
0,35 |
187 |
|
Merantis amarilla |
1,28 |
85 |
|
Shorea collaris |
1,14 |
234 |
|
Kapurs |
1,21 |
141 |
|
Dryobalanops oblongifolia |
2,37 |
202 |
|
Dryobalanops aromatica |
1,59 |
191 |
|
Keruings |
1,16 |
82 |
|
Dipterocarpus pachyphyllus |
0,086 |
270 |
|
No dipterocarpáceas |
4,68 |
39 |
|
Especies de escasa densidad |
2,12 |
50 |
|
Especies de densidad mediana |
1,23 |
67 |
|
Especies densas |
0,93 |
68 |
FUENTE: Informe FAO, A description of the data processing system... Malaysia, 1973.
Se ha empleado una clasificación grosera, según la densidad, en varios inventarios hechos en el Asia, el Africa y la América Latina. La mayor precisión, de que se da noticia en el reconocimiento hecho por la FAO en Sarawak, puede apreciarse en el Cuadro 1.
El patrón de mesovariación, o sea, la variación entre el macro y el micro nivel, es un concepto relativo que depende de la superficie del reconocimiento y de las dimensiones y formas de la unidad de muestreo. El objeto es estudiar las variaciones entre las unidades de muestreo.
En las primeras etapas del inventario forestal del Sarawak, se realizó un análisis de varianza para descubrir la componente relativa de variación dentro de las muestras y entre las mismas y determinar el número óptimo de parcelas en un racimo. Se contaba con datos de dos unidades (1 y 3) cuya superficie era de 176000 y 225000 ha, respectivamente. La unidad de muestreo consistía en un racimo de nueve puntos, según puede observarse en la Figura 1.
2. CORRELOGRAMA DE TRES ESPECIES Y UN GRUPO DE DIEZ ESPECIES DEL BOSQUE SEMIPERENNE DEL CAMERÚN
Se hizo una cubicación con un relascopio cuyo FAB era de 10 pies3/acres (2,3 metros2/ha). Las muestras eran 50-100 y estaban distribuidas uniformemente en las dos unidades de reconocimiento. Los resultados aparecen en el Cuadro 2.
El valor de R2, según la fórmula s b2/(s b2 + s w2), resultó ser de 0,3, siendo relativamente pequeña la variación entre muestras en comparación con la variación dentro de cada muestra. Una variación igualmente elevada dentro de cada unidad de muestreo la observó también el autor, al analizar los datos del reconocimiento hecho por la FAO en la Malasia peninsular.
En la selva tropical del valle del Amazonas, Ramos y col. (1972) obtuvieron un R2 de 0,3 en un caso y de 0,5 en otro. La unidad de muestreo consistía en cuatro rectángulos, poco espaciados, de 250 X 10 m, separados por una distancia de 10 metros desde el centro del racimo, en las cuatro direcciones.
El análisis, a nivel de grupos de especies, (31 grupos en total) en los bosques de Sarawak (unidad 2), dio una correlación intraclase del orden del 0,2 ó menos, en todos los grupos salvo uno, un grupo de especies no dipterocarpácea, donde el R2 era igual a 0,5.
Boon (1962) estudió el patrón de variación de Eusideroxylon zwageri, dividiendo la superficie del reconocimiento en cuarteles de dimensiones variables: 2 X 1, 4 X 1 y 4 X 2 km. Cada cuartel se subdividió en dos subcuarteles para comparar la varianza entre cuarteles y dentro de cada uno. El análisis reveló que la varianza entre cuarteles no era significativamente diferente de la varianza dentro de cada cuartel. Lo dicho se observó en todos los cuarteles y en las parcelas de todas las dimensiones.
Giudicelli y otros (1972) estudiaron la correlación espacial del número de árboles de tres especies de importancia y de un grupo de especies, en una superficie boscosa de 1500 ha, en el Camerún, empleando una parcela de 50 X 50 m. El correlogramo observado por dichos autores, que aparece en la Figura 2, revela una gran variabilidad a corta distancia con respecto a todas las especies. Además, el correlogramo revela concavidad hacia el origen. En tales casos, es probable que el muestreo sistemático sea más eficaz que el muestreo al azar.
Los resultados mencionados ponen de manifiesto una característica importante de la selva tropical: su alta variabilidad dentro de pequeñas extensiones y su relativa uniformidad en grandes extensiones.
Lo dicho parece indicar que el empleo de un mismo tipo de muestreo por racimos no producirá. una gran pérdida de precisión.
La complejidad de la microvariación, que hay que maestrear en un inventario. depende en gran medida del objetivo de éste y de la separación de las especies. Si sólo hay que estimar los parámetros a nivel de todas las especies, el patrón de variación es relativamente sencillo.
Para estudiar la influencia de las dimensiones y de la forma de la unidad de muestreo sobre la precisión, se realizó un reconocimiento piloto en la India. Había en total 39 muestras, distribuidas en dos tipos de bosque, en una superficie de alrededor de 80000 ha. Los resultados revelaron que, en la selva tropical, tanto las dimensiones como la forma ejercían una influencia importante sobre el coeficiente de variación. Por otra parte, en la selva de especies deciduas adyacente, las dimensiones y la forma producían escasos efectos sobre la precisión, como pueden observarse en el Cuadro 3.
Los experimentos de reconocimiento, realizados en la selva de Indonesia, con parcelas rectangulares de 20 m de ancho y longitud variable, revelaron que el coeficiente de variación disminuye exponencialmente con el aumento de la superficie de las parcelas. Tendió a estabilizarse en torno al 16 por ciento, cuando la superficie era superior a 1,5 ha (véase Nyyssönen, 1961).
A nivel de especie, la disposición espacial en la selva se complica mucho. Según dice Richards, al principio tiene la apariencia desconcertante de una vegetación caótica. Investigaciones detalladas, realizadas por varios ecólogos, revelan, no obstante lo dicho, que la selva, como muchas otras comunidades ecológicas, tiene una estructura orgánica. Las plantas de que se compone forman un número limitado de sinusias, cuya disposición en el espacio, aunque compleja, es discernible. Esta disposición se repite como un patrón, sólo con un escaso grado ción de selva tropical (Richards, 1971). de variación, a través de toda forma.
Heinsdijk (1961) investigó la disposición espacial de las especies en el valle del Amazonas. Observó que la distribución de muchas especies se acerca al patrón de distribución de Poisson, pero con muchas excepciones, cuando las unidades de muestreo tenían un número elevado de especies particulares, lo que parece indicar una tendencia a la distribución contagiosa.
Empleando los datos de los reconocimientos del valle del Amazonas, de Surínam y de Sarawak, el autor hizo una serie de investigaciones. Trazó el número medio de árboles por unidad de muestreo (m) y el coeficiente de variación (cv) de las 10-20 especies más importantes y estudió la relación que había entre los dos parámetros.
En una estricta distribución de Poisson, los puntos cuya relación es de 100/ y cv deben caer en una línea recta con 45° de inclinación hacia el eje X. Esto se debe a una característica importante de la distribución de Poisson, la de que la media es igual a la varianza. En todos los casos (véase Figura 3), las especies tendían a la desviación ascendente, con respecto a la relación en línea recta esperada, indicando tendencia a la agrupación. Es claro que en estos casos la expresión 100/
y cv deben caer en una línea recta con 45° de inclinación hacia el eje X. Esto se debe a una característica importante de la distribución de Poisson, la de que la media es igual a la varianza. En todos los casos (véase Figura 3), las especies tendían a la desviación ascendente, con respecto a la relación en línea recta esperada, indicando tendencia a la agrupación. Es claro que en estos casos la expresión 100/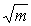 no es una guía muy satisfactoria para estimar el coeficiente de variacíon.
no es una guía muy satisfactoria para estimar el coeficiente de variacíon.
Las investigaciones de Boon (1962) sobre Eusideroxylon zwageri, revelan que esta especie tiene una notable tendencia a la agrupación.
Jack (1961) estudió la disposición espacial de las siete especies comerciales más importantes de la selva semiperenne de Ghana, empleando unidades de muestreo de varias superficies y formas. En Triplochiton scleroxylon, Khaya ivorensis y Entandrophragma cylindricum, observó una discrepancia significativa, con respecto a la distribución de Poisson y un buen ajuste con la distribución binomial negativa. Entandrophragma utile y Mimusops heckelli revelaron tendencias a la agrupación, pero la observación sólo pudo hacerse en parcelas de superficie superior a 2,0 ha; en cambio Clorophora excelsa, con cambio de la superficie de la parcela reveló un patrón que fluctuaba entre el aleatorio y el contagioso.
Giudicelli y otros (1972), en un tipo de selva semejante, en el Camerún, observaron una distribución contagiosa pronunciada en Triplochiton scleroxylon y Terminalia superba, pero una distribución aproximadamente aleatoria, en Entandrophragma cylindricum. Observaron también que el patrón de distribución tendía a hacerse regular cuando se agrupaban 10 especies de importancia.
Los estudios de distribuciones diferentes a las de Poisson se complicaban debido a que el patrón revelado por los datos observados se confundía en parte con la superficie y la forma de la parcela. Según Greig-Smith (1964), muchos tipos de patrones no aleatorios parecen aleatorios cuando las parcelas de muestreo son muy pequeñas o muy grandes y no aleatorios cuando su superficie es mediana. Lo dicho revela la necesidad de tener cuidado al sacar conclusiones generales sobre los patrones de distribución de las especies. Las consideraciones que preceden, sumadas a los estudios ecológicos citados, revelan a las claras, no obstante lo dicho, que dentro de la gama de superficies de parcelas empleadas, muchas especies dominantes de la selva revelan una distribución contagiosa.
3. RELACIÓN ENTRE EL COEFICIENTE DE VARIACIÓN (CV) Y 100/
El inventarista no se interesa tanto en los patrones como tales como en sus consecuencias para el diseño del reconocimiento. En poblaciones puras, distribuidas al azar (por ejemplo Poisson), la planificación es relativamente sencilla. En este caso, ha precisión del estimado depende sólo del número de individuos contados. El error normal del estimado es siempre igual, ya sea que se empleen muchas parcelas pequeñas o pocas grandes, o incluso una sola parcela grande (Greig-Smith 1964). La forma de la parcela no produce ningún efecto. De manera semejante, la precisión del reconocimiento casi no se ve afectada cuando las parcelas se disponen o al azar o sistemáticamente, 0 en racimos 0 no.
La situación se complica en las distribuciones contagiosas. Ya se mencionó una de las consecuencias, a saber. la falta de correlación definida entre la densidad de las plantas y el coeficiente de variación. Segundo, tanto la superficie como la forma de la unidad de muestreo influyen en la precisión de los estimados. Se observa comúnmente que, con una misma intensidad de muestreo, parcelas pequeñas, pero bien dispersas, dan una precisión mayor que parcelas grandes, compactas, poco numerosas (Dawkins, 1952; Lanly y Vannière, 1969; Greig-Smith, 1964).
Las especies difieren en su respuesta al aumento de la superficie de las parcelas. Su ecología diferencial se puede correlacionar con su disposición espacial. Triplochiton y Terminalia, que tienen ambas notable distribución contagiosa, revelan, relativamente hablando, más perdida de precisión con el aumento de la superficie de las parcelas que Entandrophragma utile, cuya disposición espacial se aproxima a la distribución de Poisson (véase Figura 2). Eusideroxylon zwageri y Khaya ivorensis, que revelan ambas tendencia a la agregación, se comportan de manera semejante.
Tratándose de la distribución contagiosa, las unidades de muestreo alargadas producen estimados más precisos que las compactas de igual superficie. Este hecho se puede visualizar fácilmente: Aquellas tienen más probabilidades que éstas de formar manchas' tanto densas como ralas.
Las investigaciones de Jack (1961) revelan que las unidades de muestreo largas y angostas, colocadas transversalmente con respecto al drenaje local, arrojan resultados más precisos que las relativamente compactas. Experiencias semejantes con los estudios ecológicos se han dado a conocer también por parte de muchos investigadores, por ejemplo, Greig-Smith (1964, p. 29) y Aubréville (1971).
Hasta la fecha, la atención se ha concentrado en el estudio de la distribución contagiosa solamente, debido a su alta frecuencia en la selva tropical. Hay muchas otras posibilidades de disposición de las plantas en el espacio. Por ejemplo, la «distribución regular», ya mencionada en relación con la distribución de a grupos de 10 especies». En relación con los estudios mencionados. al parecer, un inventario en la selva tropical, a nivel de especies individuales, probablemente seria muy caro, en comparación con uno de «todas las especies». Como alternativa es interesante averiguar qué sucede a nivel de grupos de especies. El efecto probable de la agrupación de las especies es una disminución del coeficiente de variación.
La media del coeficiente de variación de varias especies y niveles de agrupación de especies, en los bosques mixtos de dipterocarpáceas de Sarawak que aparece en el Cuadro 4, muestra a las claras que se logra hacer economías en el muestreo, operando a nivel de grupos de especies. En comparación con las especies consideradas individualmente a nivel de grupo hay mucho menos variabilidad. El número de unidades de muestreo en blanco disminuye, y la simetría de la curva de frecuencia decrece. El efecto de la superficie y de la forma de la unidad de muestreo siguen siendo notables y, como ya se dijo, esto continúa sucediendo incluso a «nivel de todas las especies».
AUBRÉVILLE, A., 1971, Regeneration patterns in the closed forests of Ivory Coast. En World vegetation types ed. by S.R. Eyre, p. 41. London, Macmillan.
BOON, D.A., 1962, Plot size and variability. Delft, International Training Centre for Aerial Survey. ITC publication, Series B, No. 17.
COX, F., 1971, Dichtebestimmung und Strukturanalyse von Pflanzenpopulationen mit Hilfe von Abstandsmessungen. Mitt. BundForsch-Anst. Forst-u. Holzw., 87.
DAWKINS, H.C., 1952, Experiments in low percentage enumerations of tropical high forest with special reference to Uganda. Emp. for. rev. 31:131.
DAWKINS, H.C., 1958, The management of natural tropical high forest with special reference to Uganda, Oxford, Imperial Forestry Institute. Paper No. 34.
GIUDICELLI, X., LANLY, J.P., OUAKAM, J.B. & PIETRY, M., 1972, Application de la theorie des processus aléatoires a l'estimation de la précision d'un inventaire forestier par échantillon nage systématique. Annls. sci. for., 30:267.
GREIG-SMITH, P., 1964, Quantitative plant ecology. London Butterworths.
HEINSDIJK, D., 1961, Forest survey in the Amazon valley. Unasylva 15:167.
JACK, W.H., 1961, The spatial distribution of tree stems in a tropical high forest. Emp. for. rev. 40:234.
KEMPTHORNE, O., 1957, An introduction to genetic statistics. New York, Wiley.
LANLY, J.P. & VANNIÈRE, B., 1969, Précision d'un inventaire forestier en function de certaines de ses caractéristiques. Revue Bois Forêts Trop. 125:35.
LOETSCH, F. & HALLER, K.E., 1973, Forest inventory. Vol. 1. München, BLV Verlagsgeselischaft.
LOETSCH, F., ZÜHRER, F. & HALLER, K.E., 1973, Forest Inventory. Vol. 2. München, BLV Verlagsgeselischaft.
MATÉRN, B., 1960, Spatial variation. Meddn. St. SkogsforskInst., 49(5).
MATÉRN, B., 1972, Poisson process in the plane and related models for clumping and heterogeneity. Symposium lecture on Analysis of spatial patterns and ecological relations.
MEYER, H.A., 1953, Forest mensuration. State College, Pa.
NYYSSÖNEN, A., 1961, Survey methods of tropical forests, Rome, FAO.
RAMOS, A.A., VIEIRA, A.N., VIVACQUA, C.A., ALENCAR, J.C., BARROS, J.C.M. & PÉLLICO NETTO, S., 1972, Inventario florestal do distrito agropecuario da zona Franca De Manus. Revta flor. 4(1).
RICHARDS, P.W., 1952, The tropical rain forest, p. 10, 249. Cambridge, University Press
RICHARDS, P.W., 1971, The structure of tropical rain forest. En World vegetation types, ed. by S.R. Eyre, p. 26. London, Macmillan.
SCHIMPER, A.F.W., 1903, Plant-geography upon a physiological basis. Tr. by W.R. Fischer. Ed. by P. Groom and I.B. Balfour. Oxford, Clarendon Press.
WECK, J. & WIEBECKE, C., 1961, Weltforstwirt schaft und Deutschlands Forst- und Holzwirtschaft. München.
BRASIL
Report to the Government of Brazil on the forest inventory in the Amazon valley. 1957, FAO/ETAP Report No. 601.
Report to the Government of Brazil on forest inventories in the Amazon. 1965, FAO/EPTA Report No. 2080.
GUYANA
Inventory of the Ebini-ltaki area. 1970, Guyana. Forest Industries Development Survey Technical Report No. 9. FO-UNDP/SF GUY/9.
INDIA
Forest resource survey. 1970, India. Preinvestment Survey of Forest Resources. Technical Report No. 1. FO-UNDP/SF IND/23.
MALASIA
A description of the data processing system used in the inventory of selected areas of the mixed dipterocarp forest in Sarawak. 1973, Malaysia. Forestry and Forest Industries Development Project. Working Paper No. 22. FO: DP/MAL/72/009.
The timber species of the mixed dipterocarp forest of Sarawak and their distribution. 1973, Malaysia. Forestry and Forest Industries Development Project. Working Paper No. 21. FO: DP/MAL/72/009
![]()
![]()
![]()
{kind=link}
{kind=link}