![]()
![]()
![]()
by
Thomas T. Veblen and Roberto Delmastro N.
Instituto de Silvicultura
Facultad de Ingeniería Forestal
Universidad Austral de Chile
Valdivia, Chile.
1. Introduction
The absence of Araucaria araucana (Mol.) Koch from the list of conservation priorities compiled by the FAO Panel of Experts on Forest Gene Resources (FAO, 1974) may be partially due to lack of knowledge of the value and of the current status of this species. The purpose of this report is to summarize the information available on the A. araucana gene resource in Chile and thus provide the Panel with a basis for evaluating the conservation needs of this species.
Of the 10 species of Araucaria, 2 are present in South America - A. angustifolia in southern Brazil and northeastern Argentina and A. araucana in Chile and Argentina (Pitcher, 1975). A. araucana is a massive tree which occasionally reaches heights up to 50 m and trunk diameters of over 2 m, although maximum heights tend more frequently to be 25 to 35 m and diameters at breast height 1 to 1.5 m. The juvenile form is quite distinct from the form of an adult tree. The juvenile stage has a conical shape and the whorled branches (which occupy the full length of the trunk) are horizontally oriented except for the extremes, which are turned upwards. In the adult form the lower branches are missing and the crown is much reduced in depth, giving it the appearance of an umbrella. The extremely straight cylindrical bole and whorled branches give the tree a majestic appearance which has made it internationally popular as an ornamental. The tortoise-shell-like bark is commonly 10 cm thick and occasionally as thick as 18 cm (Montaldo, 1974), which adds to the unique appearance of A. araucana. For taxonomic descriptions of the species see Dallimore and Jackson (1966) or Munoz Pizarro (1971).
2. Distribution and Habitat
The present distribution of A. araucana represents a small fraction of its distribution at the time of European colonization of Chile. The two principal centres of distribution in Chile (see Map 1) are: (1) in the coastal cordillera (Cordillera de Nahuelbuta) between 37°20' and 38°40'S latitude and (2) in the Andes between 37°30' and 39°30'S latitude (Montaldo, 1974; Schmithusen, 1960). On the Argentine side of the Andes, A. araucana extends as far south as 40°03' in the region of Lago Meliquina (Baeza and Llana, 1965; Schmithusen, 1960).
A. araucana is found under a relatively wide range of climatic and edaphic conditions. The sparse meteorological data available indicate average annual precipitation varies from 1 500 to 3 000 mm in the northern part of its range (Montaldo, 1974) and increases substantially at points further south reaching well over 4 500 mm at approximately 450 m at 39°30'S (Burschel, et al., 1970). Much of the winter precipitation at altitudes where A. araucana occurs is in the form of snow. The lack of a continental climatic regime in south-central Chile is reflected by the relatively mild range of mean monthly temperatures from -1° to 15°C reported for the A. araucana zone (Corfo, 1950). However, the daily range of temperatures in the higher altitudes of the south-central Andes is substantial with below freezing temperatures occurring even during the summer months. A. araucana grows under a similarly wide variation of soil conditions. In the coastal cordillera it is found on soils derived from Paleozoic and Precambrian granitic rocks and in the Andes on substrates ranging from Jurassic metamorphic and sedimentary materials to Quaternary volcanics. Characteristically the soils beneath A. araucana have an acid pH of 4.5 to 5.4 and tend to be sandy in texture. A. araucana is found on both deep, well developed soils and on incipient soils with A horizons of only 10 cm resting directly on sandy volcanic tuff deposits.
In the coastal cordillera A. araucana occurs mainly from 1 000 to 1 400 m although Montaldo (1974) reports a small stand at 600 m at approximately 38°38'S latitude. Commonly at altitudes of 1 100 to 1 250 m in the coastal cordillera A. araucana forms a mixed forest with Nothofagus dombeyi in which the common shrubs are Desfontainea spinosa, Pseudopanax laetevirens, Myoschilos oblonga and Drimys winteri var. andina. The A. araucana-N. dombeyi forests occupy all aspects other than southerly ones and occur on sandy well-drained soils.
The altitudinal distribution of A. araucana forest in the Chilean Andes is from 600 m to the timberline, although Nielsen (1963) reports some isolated individuals as low as 500 m which suggests that the present lower altitudinal limit of the forest is the result of deforestation. In the northern part of its distribution the upper forest limit occurs at 1 700 to 1 800 m and in the southern range at 1 500 to 1 600 m. Depending on topographic position and exposure, A. araucana forms the upper forest limit in alternation with Nothofagus pumilio. For example, on Volcán Quetrupillán (39°30'S) near its southern limit in Chile, A. araucana is present from 1 200 m upwards as an isolated tree in the zone of N. dombeyi forest and forms pure forests or mixtures with N. pumilio from 1 300 to 1 600 m. On south-facing slopes pure stands of N. pumilio occur while on the flat areas, ridges and slopes not definitely exposed to the south mixed stands of A. araucana and N. pumilio predominate. On some of the north-facing slopes and ridges pure stands of A. araucana are present. The most common shrubs in the A. araucana - N. pumilio forests of this zone are Drimys winteri var. andina, Myoschilos oblonga, Berberis buxifolia and other species of Berberis.
In the northern part of its distribution (at approximately 38°20'S) A. araucana first appears at 800 m where it grows in a mixed forest with predominantly N. dombeyi and N. obliqua and less frequently with N. alpina (Nielsen, 1963). Above 1 000 m it mixes with N. pumilio which forms an intermediate layer approximately 10 m below that formed by A. araucana. This mixed forest is characterized by a luxuriant shrub layer of Desfontainea spinosa, Berberis spp., and Chusquea coleu and extends as high as 1 740 m on the north-facing slopes. A few kilometres to the south on the slopes of Volcán Llaima, A. araucana stands are present at 1 400 to 1 600 m and contain relatively small numbers of N. pumilio and N. antarctica. This volcano erupted during the previous century and thus the A. araucana forests occur as isolated patches separated by areas undergoing colonization by herbaceous plants (Montaldo, 1974).
Throughout its range, A. araucana is found at altitudes and on poor soils where very few other Chilean trees can grow. Its restriction to marginal sites may be due to an inability to compete successfully at lower altitudes and on better soils. However, the absence of A. araucana from some of the better sites may be partially explained by its elimination by cutting from the lower and more accessible sites. Nevertheless, the current restriction of the species to less favourable sites tends to support the theory that the distribution of A. araucana is relict and is currently undergoing range reduction as a result of natural processes. Auer (1951) and Kalela (1941) suggest a gradual diminution of precipitation has occurred over many decades in the southern Andes, which has allegedly resulted in an increase of N. antarctica scrub and of steppe vegetation types at the expense of A. araucana, while the fossil evidence indicates that the species had a much greater distribution during the Tertiary (Schmithusen, 1960). A. araucana cannot be considered a relict in the sense of not being able to reproduce under current climatic conditions. That this is not the case is clearly indicated by the presence of all age classes in A. araucana forests throughout its range.
3. The Current Status of A. araucana
A. araucana produces an excellent quality yellowish-white wood of great mechanical strength which is suitable for a wide variety of uses as long as it is not exposed to excessive humidity (Nielsen, 1963). Since the colonial period, the species has been heavily exploited as a construction timber, especially in the mining industry, and as a result the area of A. araucana forest has been drastically reduced (Elizalde, 1970; Montaldo, 1974). Exploitation usually takes place without any provision for restocking and it appears that natural regeneration is inadequate to maintain the resource. In addition to actual felling of the forest, the regeneration problem is exacerbated by the collection of seeds which were an important staple of the Mapuche Indians' diet during the pre-Hispanic period. Such collection continues on a large scale today.
In addition to negative human influences, A. araucana is plagued by a variety of other biotic pests. When seeds fall during March and April a large percentage is consumed by the slender-billed parakeet, rodents, and the introduced wild boar. Such seed predation is an important negative factor because of the very low viability of the seeds (Montaldo, 1974). A. araucana is also attacked by a wide variety of insects but generally only overmature trees or those already weakened by other factors are seriously damaged. More serious damage is done by the foliage-attacking fungus, Micronegeria fagi, which spends part of its life cycle on Nothofagus obliqua and N. alpina.
Due to the influence of this fungus as well as to the insect pests, the establishment of A. araucana plantations at altitudes where more rapid growth may be expected (i.e., under 600 m) would probably be difficult and costly in Chile. However, outside of the range of its insect and fungal pests the establishment of A. araucana plantations may be feasible. Although the increment is typically very slow on the poor sites where it occurs naturally, average annual diameter increments of 1 cm for the first 20 years of growth have been recorded for individuals occurring on better quality sites (Tortorelli, 1956, in Nielsen, 1963). Because of its capacity to grow on the poorest forest soils, A. araucana has been recommended as a reforestation species in Chile (Donoso, 1974) and deserves consideration as a reforestation species in other eroded mountain environments.
Since 1960 in Argentina the felling of A. araucana has been prohibited and the principal remaining stands fall under the jurisdiction of national parks (Erikson, 1975). In the case of Chile it is still legal to cut A. araucana as long as a management plan, which must include provisions for regeneration, is approved by the national forestry authorities (Chile, 1974). What provisions are required are not specified and, due to the lack of research on the ecology of this species, it is unknown which treatments will assure a safe and rapid regeneration. Parts of the central portions of the range of A. araucana lie within national parks but cannot be considered totally protected because of the cutting and grazing activities which sometimes occur illegally within Chile's national parks. Perhaps one of the greatest dangers to the A. araucana gene resource in Chile may now be the competing demands for the use of its mountain habitat for tourist and recreational developments.
4. Recommendations
While Araucaria araucana does not appear to be in imminent danger of extinction, the gene pool has long been and continues to be seriously impoverished. Outside of Chile A. araucana is widely used as an ornamental and some of the genetic potential of the species is preserved in such ornamental plantings. However, the provenances of these ornamental plantings are generally unknown and consequently the proportion of the genetic potential preserved in ornamental plantings cannot be determined.
Considering the high quality of its wood and its potential for use in the reforestation of marginal forest sites, the following actions are recommended to assure the preservation and non-destructive use of this forest genetic resource.
A range-wide reconnaissance should immediately be undertaken to assess the extent of the A. araucana resource and its status. All potential threats to the survival of local populations must be evaluated with special attention paid to the possibly negative influences of recreational developments. Based on this reconnaissance a series of recommendations to assure in situ conservation should be presented to the national authorities responsible for national parks and forest reserves in Chile.
Guided by the results of this reconnaissance, a basic ecological research programme should be initiated in order to provide the scientific knowledge required for the preservation-management of A. araucana.
A range-wide seed collection programme should be undertaken to provide materials for ex situ conservation and possible future provenance trials.
5. References
1. Auer, V. 1951 Consideraciones científicas sobre la conservación de los Recursos Naturales de la Patagonia. Inst. Suelos y Agrotécnica, Pub. 20. Buenos Aires. 36 pp.
2. Baeza, V.M. & A.L.G. Llana. 1965 Las coníferas chilenas. Unpubl. ms. Valdivia, Chile. 24 pp.
3. Burschel, P., O. Martínez & B. Muller-Using. 1970 Fundamentos Ecológicos Básicos de Regeneración Natural para la Planificación del Aprovechamiento Sostenido de Bosques de Coigüe y Rauli. Mimeo. Universidad Austral de Chile, Valdivia. 85 pp.
4. Chile. 1974 Legislación Forestal. Corporación Nacional Forestal. Santiago. 68 pp.
5. CORFO. 1950 Geografía Económica de Chile. Vol. 1. Corporación de Fomento. Santiago. 428 pp.
6. Dallimore, W. & A. Jackson. 1966 A Handbook of Coniferae and Ginkgoaceae. London. 729 pp
7. Donoso, C. 1974 Dendrología: árboles y arbustos chilenos. Univ. de Chile. Fac. de Ciencias Forestales. Manual No. 2. Santiago 142 pp.
8. Elizalde, R. 1970 La Sobrevivencia de Chile. Santiago. 492 pp.
9. Erikson, W. 1975 Disruptions in the Ecosystems of the Steppe and Forest Regions of Patagonia by Climate and Man. Applied Sciences 6: 127–142.
10. FAO 1974 Report of the Third Session of the FAO Panel of Experts on Forest Gene Resources. Rome. 90 pp.
11. Instituto Forestal. 1960 Clasificación Preliminar del Bosque Nativo de Chile. Informe Técnico No. 27. Santiago. 19 pp.
12. Kalela, E.K. 1941 Uber die Holzarten und die durch die klimatischen Verhaltnisse verursachten Holzartenwechsel in den Waldern Ostpatagoniens. Ann. Acad. Scient. Fenn. Ser. A 1 2. Helsinki.
13. Montaldo, P.R. 1974 La Bio-Ecología de Araucaria araucana (Mol.) Koch. Inst. Forestal Latino-Americano de Investigación y Capacitación. Bol. No. 46–48: 1–55. Mérida.
14. Muñoz Pizarro, C. 1971 Chile: plantas en extinción. Santiago. 248 pp.
15. Nielsen, U. 1963 Crecimiento y propiedades de la especies Araucaria araucana (Mol.) Koch. Unpubl. thesis, Univ. Austral de Chile. Valdivia. 77 pp.
16. Pitcher, J.A. 1975 Report on an FAO Project to Establish International Provenance Trials of Araucaria angustifolia (Bert.) O. Ktze. Forest Genetic Resources Information, No. 4: 59–64.
17. Schmithüsen, J. 1960 Die Nadelhozer in den waldgesellschaften der sudlichen Anden. Vegetatio 10: 313–327.
18. Tortorelli, L.A. 1956 Maderas y bosques argentinos. Buenos Aires. 3 vols.
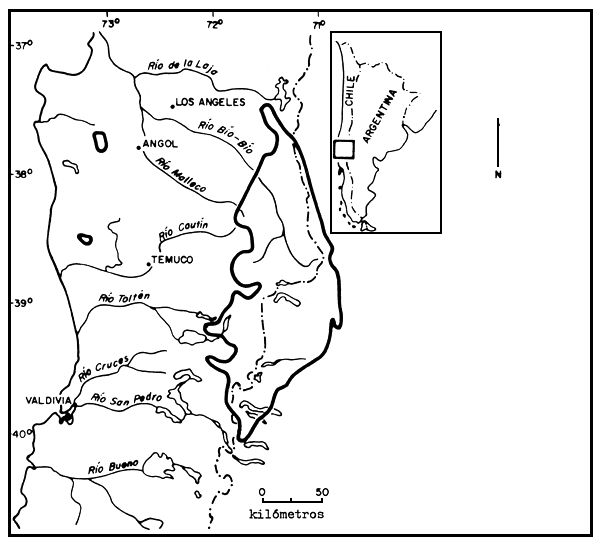
Map 1. The present range of Araucaria araucana. (After Montaldo, 1974, and Instituto Forestal, 1966.)
![]()
![]()
![]()