![]()
![]()
![]()
3.1 Ecology and distribution of adults
3.2 Behaviour and catch rates
3.3 Reproduction and larval migration
3.4 Migration of juveniles
3.5 Recruitment and sexual maturity
3.6 Growth
3.7 Mortality
Garcia (1976) made a comprehensive study of the biology of the species in Ivory Coast. As the life cycle seems to be general, a summary of the main results (Garcia, 1978) will be presented here.
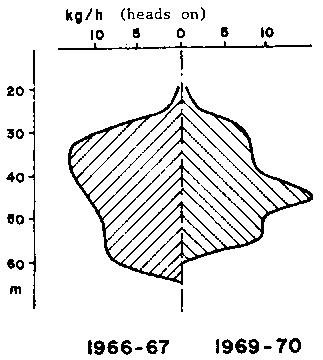
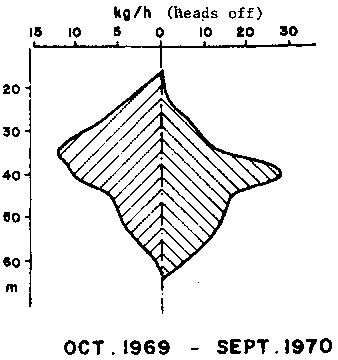
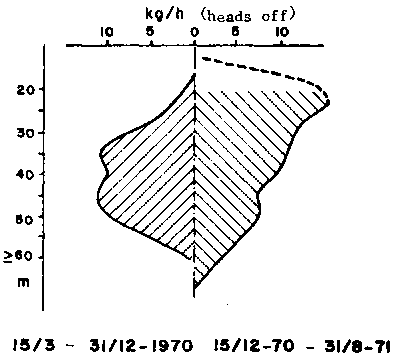
Figure 1 - Bathymetric distribution of catch rates by the Abidjan fleet in the Gulf of Guinea (after Garcia, 1977)
Radiale de GRAND - BASSAM
SASSANDRA - TABOU
GHANA - NIGERIA
TOGO - DAHOMEY
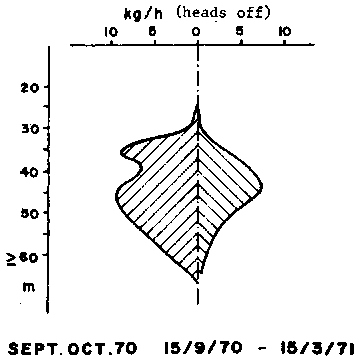
Table 1
SURFACE AREA OF SHRIMP FISHING GROUNDS IN THE IVORY COAST-ANGOLA SECTOR
|
COUNTRY |
STOCKS |
AREA IN SQUARE MILES |
|
IVORY COAST
|
Tabou-Sassandra |
250 |
|
Grand Lahou |
80 |
|
|
Bass am |
60 |
|
|
GHANA
|
Axim-Takoradi |
120 |
|
Ada-Keta |
80 |
|
|
TOGO |
|
90 (max.) |
|
BENIN |
|
130 |
|
NIGERIA
|
Lagos |
|
|
Rivers Benin to Pennington |
1 300 1/ |
|
|
Rivers Bonny to Calabar |
2 500 2/ |
|
|
CAMEROON |
|
190 |
|
EQUATORIAL GUINEA |
|
? |
|
GABON |
Cap Esterias |
200 |
|
CONGO |
|
No exploitable concentration |
|
ANGOLA |
|
No exploitable concentration |
|
TOTAL |
|
2 500 or 3 700 |
1/ Burukovski and Bulanenkov (1969)
2/ Raitt and Niven (1969)
The species can be found from Cape Hatteras, on the North American coast, to Cape Frio, on the East African coast (Perez Farfante, 1969). On either side of the Atlantic, Penaeus duorarum are found in areas where surface temperatures approach at least 24 °C during the year and seldom fall below 18 °C. This could be linked to metabolic requirements concerning notably the reproductive mechanisms.
Within these temperature limits the distribution of adults is controlled by salinities exceeding 35 percent. In the hydrologically suitable areas, the distribution is linked to the existence of lagoons or estuaries and sediments having fine particles (£ 50 µ) exceeding 25 percent. One of the important characteristics of shrimping grounds is the abundance of organic matter. The bathymetric distribution of the species is linked to the structure of the sediment: the valuable concentrations can be found in places where fine articles exceed 50 percent. Lower limit of the distribution - toward 60-65 m - is probably linked to the hydrological conditions.
In the regions where oscillations of the thermocline exist, the bathymetric distribution varies seasonally according to the rhythm of the oscillations. The shifting tends to maintain the species at the upper part of the layer of discontinuity, between temperature range of 23-25 °C and salinity of 35-35.5 percent. In the upwelling season, these waters disappear from the continental shelf and the shrimps are thus exposed, in some areas, to less favourable conditions (16-18 °C).
The bathymetric distribution of length and sexes is very characteristic (Figure 1) and different from those observed in the Gulf of Mexico. The small and medium sized individuals occupy the central part (30-45 m) of the area of distribution. They are the most abundant and their sex ratio is near normal. The average length increases while the abundance decreases offshore where males are predominant. On the other hand, the mean length increases while the abundance decreases toward the coast where females are more abundant. The bathymetric distribution of the sexes changes seasonally, probably in connexion with reproduction. Off the Niger delta however, the abundance is greater in the shallow areas (20-25 m).
On the coast of Africa, high catch rates can be obtained, either during the day or night. Obtaining maximum catch rates in the day-time is extremely frequent in the Ivory Coast, Nigeria and Senegal (Garcia, 1977; Raitt and Niven, 1966). In Gabon, the best catch rates are observed during the night (Fontana and Bâ, 1972).
This apparent behaviour depends on the area, time and depth. It is admitted that the activity is mainly nocturnal and that apparent behaviour is only a manifestation of nycthemeral modifications of the vulnerability of shrimps. Such modifications helped by the turbidity of the waters are supposed to be the result of vertical migrations placing shrimps periodically out of reach of the trawls (hypothesis put forward by Le Guen and Crosnier, 1968, in Congo). Fontana and Bâ (1972) have stated that when trawling against the wind, it is possible to obtain higher catch rates (by 25 percent) than trawling with the wind.
In the Ivory Coast, the proportion of mature females in the catches increases from October after the cold season, when the water gets warmer. From October to December, higher percentages of females are observed in the catch. These decrease irregularly after December until May-June. Occasionally higher percentages of females are observed in May-June.
The abundance of post-larvae at the mouth of lagoons follows nycthemeral, lunar and seasonal rhythms. Larvae are present only during the night. The nocturnal abundance curve is unimodal or bimodal according to the lunar phase and the tidal cycle. The overall abundance depends on the lunar phase: it is maximum at new moon, minimum at full moon and intermediate during quarters. These variations reflect either a lunar rhythm of egg-laying, or more likely tidal rhythm with interaction by lunar luminosity, involving a decline in abundance of surface larvae during full moon.
The size of post-larvae at the same state of development varies seasonally with the abundance of plankton and the temperature (inverse relation).
Juveniles migrate when they are 3 to 4 months old after a period of 2.5 to 3 months in the lagoon. There exists a nycthemeral and tidal rhythm: migration takes place by night at ebb-tide. Seasonal variations of annual amplitude are obviously linked to the cycle of reproduction and to the external factors governing it. Peak migration takes place at spate-time, when salinity is low and the currents rapid. The most important catches are recorded from November to May in the Ebrié lagoon (Ivory Coast); from January to May in the Tagba lagoon (Ivory Coast), and from February to May in the Lagos lagoon (Nigeria) (Obakin, 1969 and 1970).
The limits of distribution of juveniles in lagoons are influenced by the sea (tidal currents, saline wave). The length at migration varies seasonally, according to regions it is 16-17 mm (carapace length) in Ivory Coast. On the average, it is slightly smaller in Nigeria (13-15 mm; Obakin, 1969 and 1970). The length at migration also varies, from one sector of a lagoon to the other, and from one lagoon to the other, in relation to the salinity of the environment. It also varies from one year to the other in the same lagoon depending on the annual rainfall and, consequently, with the salinity of the environment. This effect of salinity on the length at migration is supposed to be effective through a shortening of the duration of the stay in the lagoon when salinity gets low.
In the Ivory Coast, the recruitment at sea, estimated from the percentage of small sized individuals (CL under 22 mm) in the marine catches, agrees with the annual mean curve of migration out of the lagoons. It is maximum from February to April. It is also the case in Nigeria (Bayagbona, Sagua and Afinowi, 1971) and in Gabon (Fontana and Bâ, 1972). The comparison of mean annual cycles of spawning, migration and recruitment made it possible to reconstruct the life cycle in Ivory Coast (Figure 2). If post-larvae entering the lagoon at 3-4 rostral spine stage are three weeks old, peak migration takes place toward 3-4 months of age when carapace length has reached 16-17 mm. The first modes (18 mm, CL) in the length frequency distribution of catches at sea are nearly 4 months old. The time elapsing between migration and recruitment from the fished population is very short. The size at first maturity (30 mm, CL in Ivory Coast) is reached in about 6-7 months. The smallest mature female found in Nigeria by De Vries and Lefevere (1966) measured 28 mm (CL). Shrimps can live up to at least two years.
Figure 2 - Reconstruction of the sequence of life cycle in Ivory Coast (after Garcia, 1977)
Various methods including tagging were used for growth studies in the Ivory Coast. They all produced consistent results. Growth is rapid during the warm season, highly disturbed during the cold season and apparently during the spawning season too. No significant difference has been observed in the mean growth between shrimps on the African and American coasts.
The annual mean growth curve can be represented by the following parameters:
|
|
Tagging |
Petersen’s Method |
|
|
Males
|
K months |
0.30 |
0.26 ± 0.50 |
|
CL¥ |
32.3 mm |
34.4 mm ± 2.0 |
|
|
Females
|
K months |
0.18 |
0.16 ± 0.05 |
|
CL¥ |
51.7 mm |
51.7 mm ± 3.4 |
|
In the Ivory Coast, tagging made it possible to estimate natural mortality (M) equal to 0.25/month; the estimate of M obtained from cohort analysis is M = 0.21/month. Seasonal variations of catchability have been made evident.
![]()
![]()
![]()
{kind=link}