![]()
![]()
![]()
Ariel E. Lugo et Sandra Brown
ARIEL E. LUGO est chef de projet à l'Institut des forêts tropicales, U.S. Forest Service, Rio Piedras (Porto Rico). Sandra Brown est chercheuse au Centre for Wetlands, université de Floride.
Les forêts tropicales influent sur le bilan du carbone dans l'atmosphère du globe de plusieurs façons: en stockant rapidement de grandes quantités de carbone, en le libérant sous forme de gaz carbonique ou anhydride carbonique dans l'atmosphère via la respiration, en exportant la matière organique vers les nappes aquifères profondes ou les écosystèmes océaniques, et enfin en réagissant aux interventions humaines: extraction de bois, déboisement ou aménagement en vue de la production ligneuse. Ceux qui pensent que les forêts tropicales sont des sources de carbone atmosphérique soutiennent que le rythme rapide du déboisement réduit l'accumulation de carbone et accroît la quantité de gaz carbonique libérée dans l'atmosphère par suite de la combustion et de la décomposition de la végétation. Ceux qui au contraire considèrent les écosystèmes forestiers tropicaux comme des dévoreurs de carbone soulignent la rapidité des successions végétales et la capacité élevée d'absorption nette de CO2 des forêts tropicales.
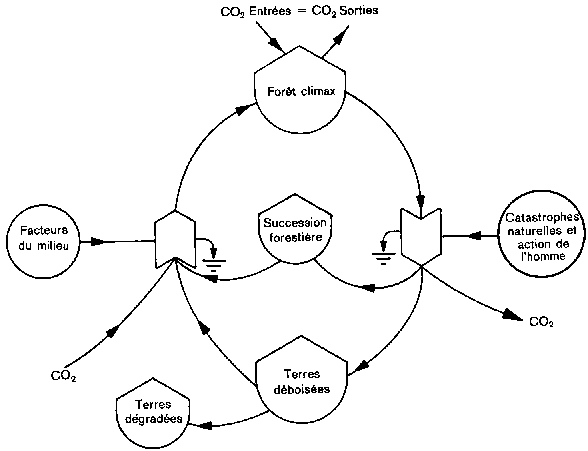
Une étude objective de la question doit prendre en considération les deux termes de la discussion, illustrés dans la figure 1 qui montre un diagramme des différents modes d'utilisation des terres et leurs incidences sur le bilan du carbone. Les forêts climaciques (en haut de la figure 1) sont censées réaliser l'équilibre entre l'absorption et la libération de carbone. Cette dernière l'emporte sur l'absorption lorsque l'homme ou les forces naturelles convertissent la forêt climacique en l'un des trois états possibles. L'état le moins perturbé est celui que l'on désigne sous le nom de forêt secondaire, ensuite viennent les surfaces coupées à blanc ou déboisées, et enfin les terres vouées à une utilisation permanente non forestière. La quantité nette de carbone libéré dans l'atmosphère s'accroît à chacune de ces étapes. Certains chercheurs supposent que c'est pratiquement le seul processus inter venant dans les régions tropicales, que 95 pour cent du carbone de la biomasse se retrouvent dans l'atmosphère sous forme de CO2 dans les zones déboisées et que les successions végétales ne jouent qu'un rôle négligeable.
A gauche, le diagramme montre le passage de la forêt perturbée à la forêt climacique, caractérisé par une absorption nette de carbone dont la rapidité dépend des conditions du milieu et du rythme de succession. Cette succession est lente dans les milieux arides, beaucoup plus rapide en milieu humide. La forêt secondaire se reconstitue évidemment plus vite qu'une zone entièrement déboisée, tandis qu'un terrain affecté à une utilisation non forestière a peu de possibilités de faire office de capteur net de carbone. Le bilan entre évolution descendante et évolution ascendante de l'occupation des terres, tel que le montre la figure 1, détermine si une région tropicale donnée produit ou absorbe du carbone. L'objet de la présente étude est d'examiner les forces et les facteurs qui peuvent faire pencher la balance d'un côté ou de l'autre.
FORÊT TROPICALE MIXTE A SURINAME le problème du carbone est tout aussi complexe
Superficies boisées et carbone accumulé à Porto Rico à diverses époques
|
Date |
Superficie boisée (10³ km²) |
Carbone accumulé1 (106 t) |
Source |
|
Epoque de la découverte (1493) |
28,500 |
93,8 |
Zon & Sparhawk, 1923 |
|
1916 |
1,787 |
19,7 |
Zon & Sparhawk, 1923 |
|
1950 |
2,352 |
26,0 |
Programme d'inventaire du Département des ressources naturelles |
|
1973 |
3,758 |
41,5 |
Programme d'inventaire du Département des ressources naturelles |
1
L'estimation pour 1493 est basée sur la répartition des biozones de Porto Rico et sur l'accumulation de biomasse dans les plantes et la litière, d'après Brown et Lugo (1980). Pour les autres années, on a admis un taux de déboisement et une rapidité de croissance identiques dans toutes les biozones.
2 En admettant que 95% de l'île étaient boisés.
1. Diagramme de flux illustrant les processus de changement dans l'utilisation des terres qui influent sur le bilan du carbone. Le déboisement entraîne une libération temporaire de carbone, qui peut être contrebalancée par une absorption de carbone au cours de la succession de végétation. Les cercles représentent les forces extérieures qui entraînent la perturbation de la forêt ou sa régénération. La case inférieure (emmagasinage) représente des terres que l'on ne laisse pas se reboiser et ne fait par conséquent apparaître aucun processus de succession. Les flèches symbolisent les flux de matière ou d'énergie.
Il existe un état d'équilibre stable du carbone atmosphérique lorsque les humains dépendent entièrement de l'énergie solaire pour leur survie (figure 2). Dans ces conditions, l'homme est un simple consommateur; il peut recourir, pour se procurer sa nourriture, à la culture itinérante traditionnelle, à la chasse ou à la cueillette. Le bilan du carbone se maintient parce que l'intensité des activités humaines n'est pas suffisamment forte pour l'emporter sur la reconstitution de la végétation. La succession forestière absorbe facilement le CO2 produit par le défrichement périodique de la végétation. En général, on considère la végétation comme étant à un stade climacique stable; les grandes étendues de forêt vierge amortissent l'effet des activités humaines. Ces forêts sont-elles aussi en équilibre avec l'atmosphère en ce qui concerne le carbone?
Que les forêts climaciques soient ou non en parfait équilibre avec l'atmosphère pour ce qui est du carbone, c'est là question qui n'a pas encore reçu de réponse définitive. On admet en général qu'il y a équilibre, mais tous les écosystèmes étant ouverts, on ne doit pas s'attendre à ce qu'ils soient parfaitement équilibrés. Des peuplements de forêt climax; de la forêt expérimentale de Luquillo à Porto Rico, par exemple, accusaient une perte faible mais mesurable de matière organique dissoute transportée vers les écosystèmes situés en aval (Odum, 1970). Odum a relevé jusqu'à 15 mg/l de matière organique dissoute dans les eaux de ruissellement lors de périodes de fortes pluies, mais il n'a fait que quatre mesures directes. Lors d'une étude de bassins versants dans les forêts de plaine du Guatemala qui a duré une année, Brinson (1973) a mesuré des concentrations élevées de matière organique dissoute dans les cours d'eau (jusqu'à 36,8 mg/l) et constaté qu'elles étaient proportionnelles au débit des cours d'eau.
De fortes concentrations de matière organique dans les eaux sont généralement associées aux écosystèmes de zones marécageuses. Ainsi, les cours d'eau drainant des bassins versants qui comportent des superficies importantes de terrains marécageux montrent des concentrations de matière organique dissoute de 3 à 72 mg/l, tandis que ceux qui drainent des bassins versants avec peu de terrains marécageux ont des concentrations de 1,8 à 12,4 mg/l. Les mangroves à manglecito de Floride exportent environ 0,7 g/m²/j de matière organique (Twilley, 1980).
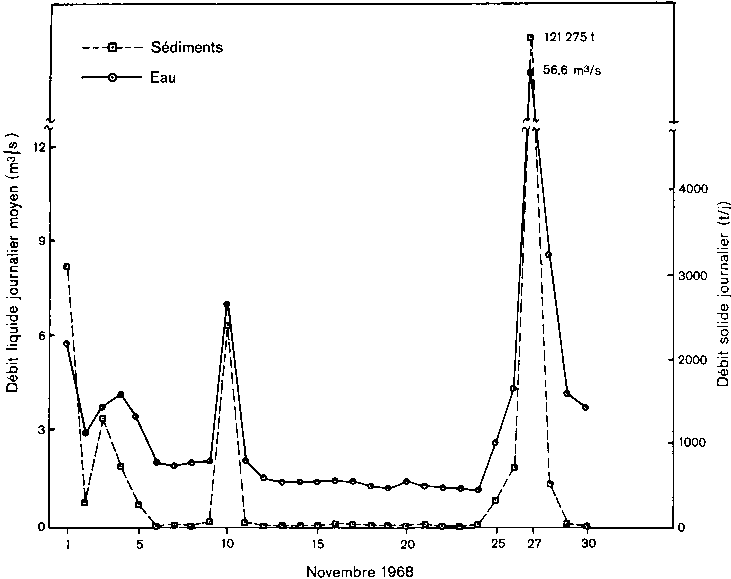
2. Débits et transport de sédiments du Rio Tanamá (Porto Rico) au cours du mois de novembre 1968.
Ces données fournissent généralement, lorsqu'elles sont extrapolées à l'échelle mondiale, des quantités absolues de carbone peu élevées. On a estimé que le bassin de l'Amazone, par exemple, exportait 0,2×1014 g (soit 20 millions de t) de carbone par an (Williams, 1968). Mais cette estimation, basée sur des mesures de concentration de carbone dissous effectuées à la saison sèche et sur le débit annuel du fleuve, a été récemment révisée et portée à 1014 g (100 millions de t) de carbone par an (Richey et al., 1980).
Ces résultats indiquent que même des écosystèmes «en équilibre» exportent de la matière organique vers les écosystèmes aquatiques situés en aval, dans lesquels le carbone peut se trouver retenu pendant longtemps. Il est encore impossible de préciser l'importance sur le plan mondial de ces quantités de matières organiques transportées, parce que les données disponibles ne permettent pas de calculs fiables. Le calcul de Williams (1968) pour l'Amazone, par exemple, se fonde sur 12 mesures, dont aucune en période de hautes eaux. L'estimation de Richey et al. (1980) s'appuie sur un échantillonnage réalisé au début de la saison des pluies, au moment où les rivières montaient et en pointe de crue. En ce qui concerne les cours d'eau pour lesquels on dispose de données, il est certain que l'exportation de matière organique est proportionnelle au débit. Cependant, plus importants encore sont les faits inhabituels qui échappent à l'observation, telle une rivière à Porto Rico (figure 2) qui en un seul jour (27 novembre 1968) a déversé 70 pour cent de son débit solide annuel et 10 pour cent de son débit liquide. Tant que l'on n'aura pas pris en considération ces phénomènes, en particulier dans les tropiques humides, il sera difficile de réfuter l'hypothèse selon laquelle il existerait un vaste capteur mondial de carbone, sous forme de matière organique produite dans les écosystèmes terrestres qui s'écoule vers l'océan par l'intermédiaire des nappes aquifères et des fleuves à l'état de carbone dissous. S'il est exact qu'une partie de cet apport organique est utilisée par les organismes marins, il est possible également que le CO2 produit par la respiration de ces organismes n'atteigne pas l'atmosphère.
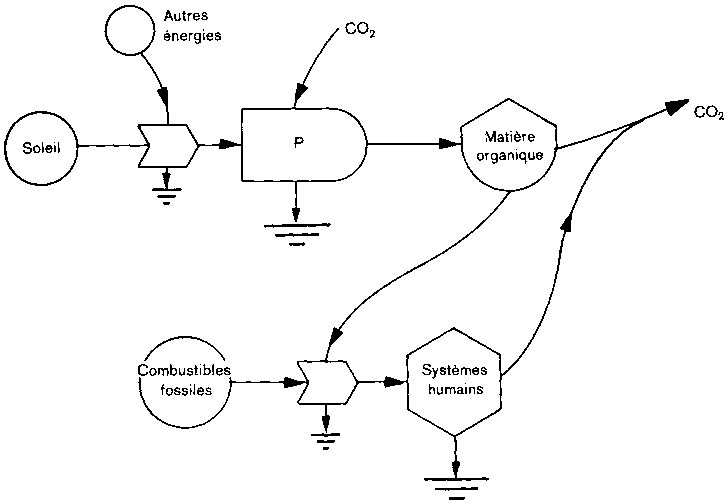
Dés lors que les hommes disposent de sources d'énergie supplémentaires, la densité démographique peut s'accroître au-delà de la capacité de charge naturelle, et ils sont alors en mesure d'exploiter plus pleinement les systèmes naturels (figure 3). Nous soutenons que cela conduit à des oscillations à long terme dans l'absorption et la libération nettes de carbone par la couverture végétale. Des exemples empruntés à deux pays en témoignent.
A Costa Rica, pays qui entre actuellement dans une phase de rapide mise en valeur des terres en raison d'un emploi croissant de combustibles fossiles, le déboisement progresse vite et ce pays devient sans doute un producteur net de carbone atmosphérique. A Porto Rico, le déboisement est intervenu beaucoup plus tôt, et les combustibles fossiles ont depuis longtemps remplacé le bois comme principale source d'énergie. Porto Rico a aujourd'hui davantage de forêts qu'au début du siècle (voir tableau). Le recours aux combustibles fossiles et la dépendance croissante à l'égard des aliments importés ont, semble-t-il, permis aux terres agricoles de se reposer et à la forêt de se reconstituer selon une succession rapide. Il est probable qu'à l'heure actuelle les terres boisées de Porto Rico sont des capteurs nets de CO2, de même que celles des régions tempérées du globe où des phénomènes analogues se sont produits.
Les changements dans l'utilisation des terres sont l'élément clef pour comprendre les échanges de carbone entre la couverture végétale et l'atmosphère. Porto Rico et Costa Rica sont des exemples des deux termes extrêmes d'une oscillation de longue durée dans l'utilisation des terres, qui est le résultat de la manière dont les hommes utilisent leur énergie fossile et autres ressources naturelles.
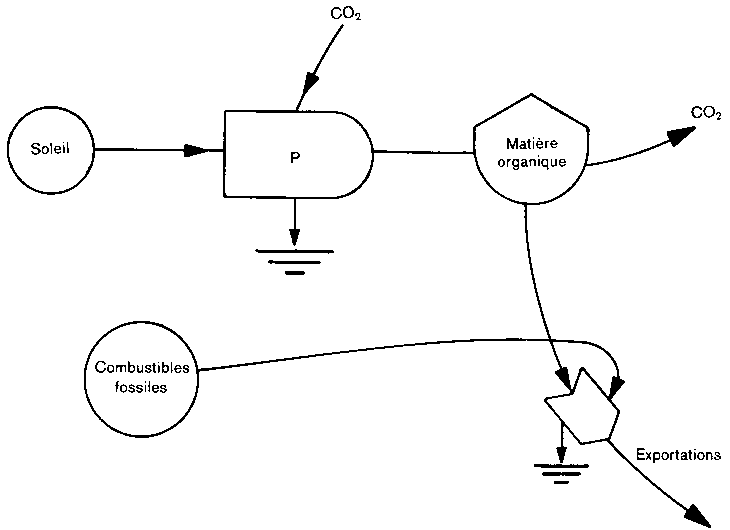
Dans les cas d'exploitation extrême des forêts (figure 4), comme cela se produit dans certaines régions tropicales humides, de grandes quantités de biomasse forestière peuvent être converties en CO2, mais cette situation peut paradoxalement conduire à une absorption du carbone, parce que ce n'est pas toute la biomasse qui est convertie en CO2, et que la succession forestière peut être assez rapide pour entretenir un écosystème jeune et dynamique avec un rythme rapide d'absorption nette de carbone.
La destination des bois coupés dans les forêts tropicales détermine la rapidité avec laquelle la matière organique retourne à l'atmosphère sous forme de CO2. La production actuelle de bois ronds dans les forêts mondiales est considérable (de l'ordre de 1,8 milliard de t), et elle s'accroît encore, la production des forêts tropicales entre pour environ 40 pour cent dans ce total. Cependant, plus de 70 pour cent de la production mondiale de charbon de bois et de bois de feu proviennent des forêts tropicales. On estime que ces productions représentent 90 pour cent du bois coupé dans la zone tropicale, soit 600 millions de t, chiffre en accroissement. Ces données indiquent que les bois tropicaux sont surtout employés comme source d'énergie, et qu'en conséquence la biomasse est rapidement renvoyée à l'atmosphère sous forme de CO2, au lieu de rester longtemps emmagasinée sous forme de bois de construction ou autres. En outre, l'utilisation de l'énergie tirée du bois sous les tropiques, est sans doute plus importante que ne l'indiquent ces statistiques, en raison des grandes quantités de bois utilisées localement et non recensées.
Par contre, lors du défrichement en vue de cultures traditionnelles sur brûlis, de grandes quantités de bois restent sur le sol, où leur décomposition est extrêmement lente. A Costa Rica, par exemple, Ewel et al. (1980) indiquent que 40 pour cent de la biomasse primitive était restée sur place, principalement sous forme de matière organique du sol et de bois.
Dans les milieux soumis à de fortes contraintes naturelles, tels que les régions très sèches ou très froides, les populations humaines, bénéficiant de sources extérieures d'énergie, risquent de déboiser les terres d'une manière irréversible et créer ainsi des conditions qui en font incontestablement des sources nettes de CO2. C'est ce qui doit se produire, dans des endroits comme Haïti et le Sahel. Dans ces milieux, la succession forestière n'est pas assez rapide pour compenser l'exploitation par l'homme, intensifiée par une population en expansion que favorisent des apports énergétiques de l'extérieur. En considérant leur état de dégradation, ces zones ont peu de chances de faire vivre leurs populations dans l'avenir lorsque les combustibles fossiles bon marché auront complètement disparu.
Il est plus facile de déceler les mécanismes et les raisons qui font que les écosystèmes tropicaux sont des dévoreurs plutôt que des producteurs de carbone. Paradoxalement, même le déboisement peut aboutir à une absorption de carbone si la succession est assez rapide et qu'une partie de la biomasse initiale reste sur place. Les écosystèmes climaciques peuvent aussi capter lentement le carbone via l'exportation de matière organique vers les écosystèmes aquatiques situés en aval. Ce processus est probablement plus important dans les milieux humides et dans les zones perturbées où l'écosystème peut être plus sensible au lessivage. L'analyse des données concernant le volcan de Mauna Loa (Hawaï) que nous avons effectuée indique également que la biosphère agit comme un capteur de carbone, de la même façon que se sont constituées dans le passé les grandes accumulations de carbone fossile.
Expérimentant sur des microcosmes fermés, Odum et Lugo (1970) ont constaté que chaque microcosme atteignait et maintenait une concentration de CO2 «atmosphérique» différente. Cet équilibre «atmosphérique» dépend des proportions relatives des plantes et des consommateurs dans le microcosme. Cela conduit à des spéculations quant à l'influence possible du biote sur des phénomènes climatiques tels que les glaciations par son action de régulation du CO2 atmosphérique. A l'heure actuelle les activités humaines intenses, fondées sur des combustibles fossiles abondants, font à nouveau pencher le bilan mondial du carbone vers une production nette de carbone, et peut-être vers un réchauffement du globe. Cependant, il semble y avoir dans les biotes terrestres une capacité d'absorption du carbone suffisante pour contrarier lentement cette tendance. En fait, on peut supposer qu'avant l'ère des combustibles fossiles, il devait y avoir diminution du CO2 dans l'atmosphère; notre analyse concernant le Mauna Loa montre en effet un taux net d'absorption de carbone par la biosphère.
Si notre analyse est correcte, il semble que, aussi longtemps que les forêts tropicales seront aménagées en vue de la production ou qu'on les laissera se régénérer naturellement, et à la condition que les surfaces coupées ne représentent qu'une faible fraction de la superficie boisée totale, le bilan du carbone sera maintenu. Des difficultés surgiront toutefois à l'échelon local, où les populations bénéficiant d'apports extérieurs d'énergie risquent d'anéantir l'écosystème forestier.
Il est difficile de trouver des preuves à l'appui de ce raisonnement. C'est justement la nécessité de réunir les informations qui manquent pour arriver à un tel niveau de compréhension qui motive l'intérêt que nous portons personnellement à cette question.
BRINSON M.M. 1973. The organic matter budget and energy flow of a tropical lowland aquatic ecosystem. Ph.D. thesis. University of Florida, Gainesville. 251 p.
BROWN, S. & A.E. LUGO. 1980. Preliminary estimate of the storage of organic carbon in tropical forest ecosystems. In S. Brown, A.E. Lugo and B. Liegel, eds. The role of tropical forests on the world carbon cycle. Proc. Symposium at the Institute of Tropical Forestry, Rio Piedras, Puerto Rico. U.S. Department of Energy Washington, D.C. (Sous presse). p. 65-117.
EWEL, J., C. BERISH, B. BROWN, N. PRICE & J. RAICH 1980. Slash and burn impact on a Costa Rican wet forest site. Ecology. (Sous presse).
ODUM, H.T. 1970. Summary: An emerging view of the ecological system at El Verde. In H.T. Odum and R.F. Pigeon, eds. A tropical rain forest. U.S. Atomic Energy Commission. NTIS, Springfield, Va. Ch. I-10.
ODUM, H.T. & A. LUGO. 1970. Metabolism of forest-floor microcosms. In H.T. Odum and R.F. Pigeon, eds. A tropical rain forest. U.S. Atomic Energy Commission. NTIS, Springfield, Va. Ch. I-3.
RICHEY, J.E., J.T. BROCK, R.J. NAIMAN, R.C. WISSMAR & R.F. STALLARD. 1980. Organic carbon: oxidation and transport in the Amazon River. Science, 207: 1348-1351.
TWILLEY, R. 1980. Organic matter experts from black mangrove forests in south Florida. Ph.D. dissertation. Univ. of Florida, Department of Botany. (En préparation).
WILLIAMS P.M. 1968. Organic and inorganic constituents of the Amazon River. Nature, 218: 937-938.
ZON, R. & W.N. SPARHAWK. 1923. Forest Resources of the World. McGraw-Hill Book Co. New York. 2 vols. 997 p.
![]()
![]()
![]()
{kind=link}
{kind=link}
{kind=link}
{kind=link}