![]()
![]()
![]()
by
Veikko Koski
The Finnish Forest Research Institute
Unioninkatu 40 A
SF-00170 Helsinki 17
Finland
INTRODUCTION
The stated goals of seed orchards are to produce genetically improved seed of forest trees, to promote frequent seed crops, and to facilitate harvesting. On an international scale, specific rules on minimum standards for tree seed orchards have only fairly recently been published, as guidelines for countries participating in the OECD 1 Scheme for the Control of Forest Reproductive Material Moving in International Trade (OECD 1974). Although these rules specify that self-pollination and outside pollen contamination should be minimized, they do not give any quantitative guidelines or establish any specified requirements. The responsibility of defining specific criteria for seed orchards belonging to the various categories recognized in the Scheme, is left to the national authorities responsible for its implementation.
It is natural that no detailed, universal requirements can be specified which are common to all seed orchards; various categories and types of seed orchards, biological characteristics of different tree species and environmental conditions, will all have to be considered. On the other hand, our knowledge of quantitative genetics as well as of flowering biology of many forest tree species, has increased substantially since the concept of seed orchards was first recognized as an integral part of tree breeding activities. In fact, there are several investigations from the Nordic countries that indicate that the functioning of a seed orchard as a genetic unit can be studied quantitatively (Sarvas 1970, Eriksson et al. 1973, Jonsson et al. 1976, Koski 1975, Bhumibhamon 1978, Chung 1981).
Legislation requiring official registration of seed orchards and other sources of forest reproductive materials is already in force in many countries at a national scale, and the number of such countries is likely to increase in the future. If the concept of genetically improved seed is to be respected, some guarantees for the composition and properties of the seedlots provided in accordance with such legislation must be achieved. In most cases, certification by means of actual evaluation of sufficiently old progeny tests would mean a delay of anything between one to several decades. To avoid such delays, scientists working on forest tree species are often compelled to rely more heavily on genetic theory than their colleagues in the fields of agriculture and horticulture.
Several factors influence the genetic composition of the seed collected in a seed orchard. Many of these can be quantified, such as the number of genotypes included in the orchard, intensity of selection, probability of inbreeding, and amount of outside pollen contamination. Based on earlier studies, Sarvas 1970 and Bhumibhamon 1978 have suggested the use of an index system for the registration of seed orchards. Each factor contributing to the index would be evaluated separately according to a point scale; the total number of points would determine the genetic value of the seed collected in the seed orchard, enabling a consistent grading of different orchards.
Even though an index system is more objective than an intuitive judgement, it has its limitations in practical application: a high score in one factor might compensate for an unacceptably low value in another. It is therefore questionable whether the different grades arrived at by using such an index would reflect the genetic value of the reproductive material and its monetary worth, even in relative terms. It is considered more reliable and practical by the author of this paper to: (i) classify the seed orchards (e.g. clonal orchards of Pinus sylvestris; seedling seed orchards of Pinus sylvestris; clonal orchards of Picea abies, etc.); and (ii) based on this classification, define minimum requirements for certain measurable factors which influence the genetic quality of the seed.
Among the different types of seed orchards identified by the OECD Scheme (see e.g. Barner 1976), first-generation clonal seed orchards, which would fall within the OECD category “Region Orchards” 1, are presently among the most common ones at a global scale. This is the case also in Finland, which officially joined the OECD Scheme in 1980 (Hagman 1982).
Minimum requirements for clonal seed orchards of Pinus sylvestris in Finland have recently been drawn up and published (Koski 1980). These guidelines are presently operational. Most likely, similar technical guidelines for quality control of seed orchards exist also elsewhere, although published information is not known to the author. The purpose of this paper is to make the Finnish minimum requirements more widely known, and to seek comments and suggestions based on experiences in other countries and other species. Although it is unlikely that the present guidelines will find a direct, wider application, they may aid other countries through suggesting a methodology for determining quantitatively specified standards to be followed in seed orchard establishment and classification.
1 Organization for Economic Cooperation and Development.
FINNISH MINIMUM REQUIREMENTS FOR CLONAL PINE SEED ORCHARDS
General considerations
The main objective of the established requirements is to ensure that the expected genetic gain is realized when using seed collected in registered seed orchards. There are three main considerations: (1) the identification of the ramets in the orchard; (2) the clonal composition of the orchard; and (3) the pollen parents of the seed. Although important, formal features such as equal number of ramets per clone and a specific planting design, are not considered critical in the present context.
(1) Strict adherence to the rule that every single graft in a seed orchard should be identifiable, has sometimes been called pedantic and considered of doubtful value in a scheme aimed at producing commercial quantities of seed. However, it must be realized that selective thinnings, progeny tests and calculations of crossing probabilities will be nonsense if the labelling and documentation are unreliable. It is therefore essential that thorough surveys are carried out in seed orchards to reveal possible errors and alterations in the original layout, and to ensure that root stocks have not taken over and suppressed the grafted scions.
(2) The clonal composition refers to (i) the number of clones; (ii) the number of ramets per clone; and (iii) the origin of the clones. These factors influence the genetic diversity of the seed; the crossing pattern, through the synchronization of flowering; and the spatial distribution of the grafts. They also determine the area in which seed from the orchard can be utilized.
(3) The most complex requirement is the one referring to the pollen parent of the seed. All definitions for seed orchards agree that both self-pollination and outside pollen contamination should be minimized. Implicitly, this means that crossing between the select geno-types included in the orchard is promoted. Thus, all clones should ideally contribute equal proportions of female and male gametes. In practice, self-pollination and pollen contamination from outside sources can never be completely eliminated, and due to individual differences in the abundance and time of flowering, the contributions of different clones are far from equal. Only when the phenological characteristics of each of the clones included in the orchard have been determined, can the crossing pattern within the orchard be accurately evaluated; studies to clarify this aspect should therefore be started as early as possible. Actual measurements of degree of self-pollination and subsequent inbreeding should also be carried out on a single-clone basis, and their harmful effects should be quantified through progeny trials.
The simplest way of reducing inbreeding is to ensure an adequate distance between ramets of the same clone. The magnitude of outside pollen contamination can be relatively accurately measured in the field, but its importance in actual pollination is difficult to determine. In general, it is difficult to reduce the amount of background pollination 1 in already existing seed orchards. In the case of abundant background pollination, the only measure which can be taken to reduce its harmful effects, is to improve the efficiency of the crossing within the seed orchard through promoting abundant flowering and through increasing the size of the orchard.
Minimum requirements
The registration of untested clonal seed orchards in Finland is based on seven main considerations. The numeric values given have been calculated for Pinus sylvestris. These requirements and the numeric values should be viewed as a whole: all the requirements must be simultaneously fulfilled. The numeric values have been estimated allowing for the possibility that one or several of the other factors may be close to the minimum acceptable level.
1. The seed orchard must be in good physical condition and all the documents must be accurate and up to date.
A detailed map which gives the clone number of each ramet as well as spacing, is a prerequisite for any genetic investigations. In addition to the actual position map, there should also be a list that gives the clonal number and position (= row/column) of each graft and the distances between rows and columns. In Finland the layout, listing and calculation of inbreeding, etc., are done by the help of a computer programme developed for this purpose. Before any evaluation or calculations are carried out, the National Authority 1 has to verify the accuracy of the position map by visiting randomly distributed “sample plots” throughout the orchard, for a graft to graft check.
2. The geographic range of the ortets (= original plus trees) must be limited so as to facilitate synchronization of flowering and determination of the area of utilization for the seed 2
When seed orchards were first established in Finland, diverging opinions were expressed about the size of an acceptable range. A desire to increase genetic diversity and to achieve heterosis effects spoke for the inclusion of trees from a very large area; a hypothesis of the existence of highly differentiated, small, local races supported the idea of a narrow range.
Through studies carried out on the annual, physiological growth rhythm of Pinus sylvestris in Finland it has been shown that in this and other commercially important species, there is large, within-stand variation in this trait. If the variation in the annual rhythm is kept to the same magnitude in a seed orchard as it is in any one continuous natural stand, the flowering times of all clones will, however, overlap and consequently match fairly well. The assumption for this is that the phenotypic selection carried out, favouring trees of superior growth and quality has not changed the distribution patterns of the annual rhythm.
For quantifying the limits of a recommended range, Sarvas (1967, 1969, 1972) concluded that in the cool, temperate region, the duration and the accumulated heat sum of the growing season are of decisive importance for the on-set of specific physiological processes such as flowering. Based on means and standard deviations of the sum 3 calculated for synchronized natural populations, and on the coefficient of variation in such populations, it is presently specified in Finland that the ortets included in a seed orchard must originate in areas which do not differ for more than ± 130 d.d. From experience to date, it is likely that these limits will be narrowed down further in the next generation of seed orchards.
3. The number of clones must be at least 30.
This requirement is based on the following facts: (i) in P. sylvestris, clones do not flower prolifically each year and, as profuse flowering has not been used as a selection criterion in Finland, some clones consistently flower more than others (see e.g. Erickson et al. 1973; Jonsson et al. 1976; Bhumibhamon 1978; Koski and Tallqvist 1978). If the total number of clones is small, the probability will increase that the year-to-year contribution of certain clones in pollen and seed production will be improportionately large. Although no theoretical model for the optimum number of clones has been presented, several authors recommend a minimum number of clones ranging from 20–30; (ii) a small number of clones increases the probability of inbreeding, because the distances between the ramets of any one clone become small in any planting design. This will be aggravated if there is a large variation in the number of ramets per clone; a large number of ramets in some clones combined with a low number of clones in another one, inevitably causes clusters of some clones and, consequently, an increased risk of inbreeding. In Finland, the accepted variation in number of ramets is ± 30 per cent of the average number in the orchard.
4. The ramets of each clone must be spatially distributed so that the probability of self-pollination is always below 20 per cent.
Due to the presence in P. sylvestris of embryonic lethals, the proportion of living embryos originating from self-pollination is much lower than the proportion of actual self-pollination. The efficiency of such biological screening of inbreeding varies between species and individuals (see e.g. Koski 1973). Lindgren (1975) and the present author (Koski 1982) have calculated the probabilities of inbred embryos after different proportions of self-pollination. In a mode genotype 1 of Pinus sylvestris or Picea abies, the above mentioned 20 per cent self-pollination would yield 3 to 4 per cent selfed individuals in the offspring. There is no absolute criterium to set the limit at 20 per cent. However, a more strict lower limit would be difficult to apply in practice.
Estimation of the proportion of self-pollination can be made based on (i) the average values of natural self-pollination and (ii) pollen dispersal. Without doubt the actual values in some cases deviate considerably from the expectations. A more sophisticated scheme would require detailed information of the abundance of male flowering, the number of embryonic lethals in each clone, the wind conditions, etc. At the present stage of knowledge it is assumed that the proportion of self-pollination within the crown of each tree is 10 per cent in a normal stand. This assumption is connected with the abundance of male flowering (point 5) and the minimum area of the orchard (point 6). Adjacent ramets of the same clone naturally increase the probability of self-pollination.
In the computer programme presently in use in Finland, the computer checks in turn the neighbourhood of every graft within 30 meters circumference 2. If it detects a tree with the same clonal number, it calculates the exact distance between the two ramets. The program contains a function for pollen density relative to distance: P = 10 e - d/8,5, where P is the probability of self-pollination (%); e is the base of natural logarithm; and d is the distance (meters). The coefficient 8,5 has been modified from earlier empirical values (Koski 1970, Müller 1977, Shen et al. 1981). If the cumulative probability of self-pollination exceeds 20%, corresponding action must be taken in the field to lower this estimate, by removing one or several ramets. The limit of 20% is fairly liberal; in fact, it is about twice as high as the estimated, natural self-pollination in P. sylvestris, as evidenced by recent biochemical studies (see e.g. Müller 1976; Müller-Stark 1979; Shen et al. 1981; Rudin and Ekberg 1982).
5. The pollen produced by the orchard per year and hectare must be at least 20 kg.
Wind pollination functions properly only when the amount of pollen is large. Generally, insufficient pollination results in abscission of conelets and abortion of ovules, in other words, in a reduced seed crop (Sarvas 1962). In a clonal seed orchard, a small pollen crop increases the probability of self-pollination in relatively abundantly flowering clones and, above all, it increases the probability of outside pollen contamination.
As a rule, grafts of Pinus sylvestris do not start producing male flowers until they have reached a certain size. Once the male flowering has started, its quantity is for several years proportional to the age and size of the grafts. It can be roughly predicted that when the grafts have reached a height of 6 – 8 meters, or a breast height diameter of 14 – 16 cm, the pollen production will be satisfactory, provided that the number of trees per hectare is at least 250. On the other hand, too close spacing in the orchard causes early death of the lower branches, where the majority of the male strobili are produced.
Although a rough prediction of pollen production can be made based on size of the grafts, a more accurate assessment must also be made once the grafts have actually started flowering. A direct counting of all the male strobili would be too laborious to be applied in practice. In P. sylvestris, the male inflorescences can be counted even two or three years after the flowering, by counting the “scars”, or needleless gaps, on the twigs, whirl by whirl. Generally, it is enough to count the number of scars in one branch per whirl and multiply by the number of branches in the whorl. The accuracy of this method must, however, first be ascertained by comparing the number of scars in different branches.
As each centimeter of scar will correspond to a specific amount (= weight) of pollen, which can be experimentally determined, the average pollen crop per graft can be easily calculated. In Finland, the estimation of the pollen crop of a clone is based on counts of 5 ramets per clone. The total of all clones equals the pollen production of the whole orchard. In Southern Finland, the maximum pollen crops recorded in seed orchards of Pinus sylvestris have been of the order of 90 kg/ha/year 1.
6. The area of the seed orchard must be at least 5 hectares and the shortest diameter at least 150 meters.
A relatively large area of grafts is needed to generate a satisfactorily dense pollen cloud. In a small “island” of trees, the pollen is immediately transportated out of the stand and pollen density will remain low. In such circumstances, the proportion of self-pollination and possible background pollination will be high even if male flowering in the orchard is abundant. The applied, numeric values were obtained from a regression of the efficiency of pollination on the diameter of the stand (Koski 1975).
7. The proportion of outside pollen contamination (background pollination) must be below 20 per cent.
The existence of considerable pollen migration is often not adequately taken into account. Isolation belts of 100 – 200 meters are frequently recommended as a complete guard against outside pollen contamination. The production of germinable seed soon after the establishment of a seed orchard, when the male flowering has not yet commenced or is scarce, is convincing evidence of inadequate isolation. The amounts of foreign pollen and the peak periods for contamination can be easily measured during the juvenile stage of an orchard.
In Finland, where the established minimum distance to hybridizing trees is 100 meters, it has been found that the quantity of outside pollen can be directly related to the abundance of the tree species in the region, rather than in the immediate neighbourhood. If the background pollination is plentiful, the only effective measures to reduce it are to (i) enhance the within-seed orchard pollination or (ii) create phenological barriers between the seed orchard and the surrounding forests. The guidelines given in points 2, 5 and 6 above have been made considering also the problem of background pollination.
Phenological barriers can sometimes be created by physically transferring a seed orchard to a region where flowering is out-of-phase with local populations. However, the effect of a transfer is not always linearly correlated with the length of the transfer (Chung 1981): this means that the probability of undesirable crossings cannot be accurately predicted, introducing an element of uncertainty into such transfers.
The suggested minimum requirement of 20 per cent contamination or less, may seem overly liberal to many tree breeders. In many regions, more strict limits can certainly be applied. In Finland it would, however, be self- delusion to set the limit lower than this, considering the existing, vast areas of natural stands of P. sylvestris. On the other hand, the decrease in genetic gain due to a background pollination of 20 per cent is, when calculated theoretically, only one tenth as compared to a situation of complete isolation. If the expected gain were, say, 15 per cent, a background pollination of 20 per cent would thus decrease it to 13,5 per cent. Considering the fact that we are talking about first-generation orchards, in which the gene frequencies have not yet been greatly changed compared to the original populations, a decrease of this magnitude has been considered acceptable. However, when advanced-generation seed orchards are planned, a more efficient isolation must be assured.
2 Planting distance is generally 5 m × 5 m in seed orchards in Finland.
REFERENCES
Barner, H. 1976 Certification of Forest Reproductive Material. In Report on the FAO/DANIDA Training Course on Forest Seed Collection and Handling. Vol. II, pp. 65–78.
Bhumibhamon, S. 1978 Studies on Scots Pine Seed Orchards in Finland with Special Emphasis on the Genetic Composition of the Seed. Commun. Inst. For. Fenn. 94.4: 1–118.
Chung, M.S. 1981 Flowering Characteristics of Pinus sylvestris L. with Special Emphasis on the Reproductive Adaptation to Local Temperature Factor. Acta For. Fenn. 169: 1–69.
Eriksson, G., Jonsson, A. & Lindgren, D. 1973 Flowering in a Clone Trial of Picea abies Karst. Stud. For. Suec. 110: 1–45.
Hagman, M. 1982 Die neue Finnische Gesetzgebung über den Handel von forstlichem Verjüngungs-material und die dazu gehörige Regelung der Versorgung von Waldbaumsamen. Allgem. Forstz. (Jahrgang 1982, in press).
Jonsson, A. Ekberg, I. & Eriksson, G. 1976 Flowering in a Seed Orchard of Pinus sylvestris L. Stud. For. Suec. 135: 1–38.
Koski, V. 1970 A Study on Pollen Dispersal as a Mechanism of Gene Flow in Conifers. Commun. Inst. For. Fenn. 70.4: 1–78.
Koski, V. 1973 On Self-Pollination, Genetic Load and Subsequent Inbreeding in Some Conifers. Ibid. 78.10: 1–42.
Koski, V. 1975 Natural Pollination in Seed Orchards with Special Reference to Pines. Forestry Commission, Bull. 54: 83–91.
Koski, V. 1980 Minimivaatimukset männyn siemenviljelyksille Suomessa. Summary: Minimum Requirements for Seed Orchards of Scots Pine in Finland. Silva Fenn. 14: 136–149.
Koski, V. 1982 How to Study the Rate of Inbreeding in Populations of Pinus sylvestris and Picea abies. Ibid. 16 (in press).
Koski, V. and Tallqvist, R. 1978 Tuloksia monivuotisista kukinnan ja siemensadon määrän mittauksista metsäpuilla. Summary: Results of Long-Time Measurements of the Quantity of the Flowering and Seed Crop of Forest Trees. Folia For. 364: 1–60.
Lindgren, D. 1975 The Relationship between Self-Fertilization, Empty Seeds and Seeds Originating from Selfing as a Consequence of Polyembryony. Stud. For. Suec. 126: 1–24.
Müller, G. 1976 Untersuchungen über die natürliche Selbst-befruchtung in Bestanden der Fichte (Picea abies (L.) Karst.) und Kiefer (Pinus sylvestris L.) Silvae Gen. 26: 207–217.
Müller-Stark, G. 1979 Estimates of Self- and Cross-Fertilization in a Scots Pine Seed Orchard. Proc. Confer. Bioch. Gen. For. Trees. Umea. Sweden 1978: 170–179.
OECD 1974 OECD Scheme for the Control of Forest Reproductive Materials Moving in International Trade. Organization for Economic Cooperation and Development, 2 rue André Pascal, 75016 Paris, France.
Rudin, D. and Ekberg, I. 1982 Genetic Structure of Open Pollinated Progenies from a Seed Orchard of Pinus sylvestris. Silva Fenn. 16. (In press).
Sarvas, R. 1962 Investigations on the Flowering and Seed Crop of Pinus sylvestris. Commun. Inst. For. Fenn. 53.4: 1–198.
Sarvas, R. 1967 The Annual Period of Development of Forest Trees. Proc. Finn. Acad. Sci. Lett. 1965: 211–231.
Sarvas, R. 1969 Genetical Adaptation of Forest Trees to the Heat Factor of the Climate. Papers of the 2nd World Congress on Forest Tree Breeding, Washington, FAO/IUFRO, FO, TTB 69/2: 1–11.
Sarvas, R. 1970 Establishment and Registration of Seed Orchards. Folia For. 89: 1–24.
Sarvas, R. 1972 Investigations on the Annual Cycle of Development of Forest Trees, Active Period. Commun. Inst. For. Fenn. 76.3: 1–110.
Shen, H.H., Rudin, D. and Lindgren, D. 1981 Study of the Pollination Pattern in a Scots Pine Seed Orchard by Means of Isozyme Analysis. Silva Genetica 30:7–15.
Manuscript received in February 1982.
![]()
![]()
![]()
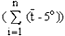 . It thus reflects the
joint effects of temperature and time.
. It thus reflects the
joint effects of temperature and time.