![]()
![]()
![]()
Les résultats des campagnes de prospection acoustiques effectuées entre 1979 et 1984 ont été présentées (Lamboeuf et al., 1984).
Les évaluations fondées sur les données de l'échointégration ont conduit à des chiffres largement sous estimés en raison des réactions d'évitement des sardines au passage du bateau, (Tableau 25, Figure 13). Pour pallier cet inconvénient une méthode fondée sur le comptage des bancs détectés au sonar dirigé latéralement par rapport au bateau a permis d'estimer le nombre total de bancs de sardines. Le poids moyen des bancs nécessaire à la transformation de ce nombre en abondance, a été déterminé à la transformation de ce nombre en abondance, a fournies par l'échointégrateur et le sonar, étalonnées à l'aide du poids des captures par calées dans la région d'Agadir. Les résultats obtenus sont présentés dans le Tableau 26 et la Figure 14.
On remarque la4 similitude de l'évolution des estimations pour les trois zones A, B et C au moyen de deux systèmes indépendants : l'échointégrateur d'une part et le sonar d'autre part; les estimations au sonar sont beaucope plus élevées. La tendance générale de l'abondance est stationnaire dans la zone A (moyenne sur 10 campagnes : 80.000 tonnes) et représente le bilan résiduel entre la quantité qui a migré en provenance de B à la saison de pêche et les captures.
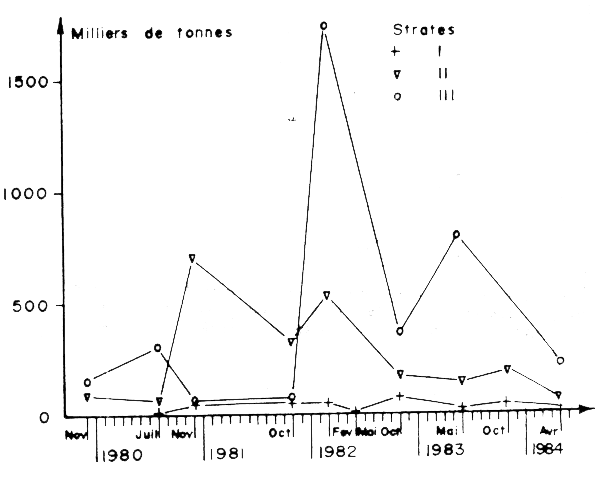
Figure 13 : Biomasse de sardines en limmiers de tonnes, estimée au sondeur couplé à un échointégrateur (données de jour) (Lamboeuf et al., 1984)
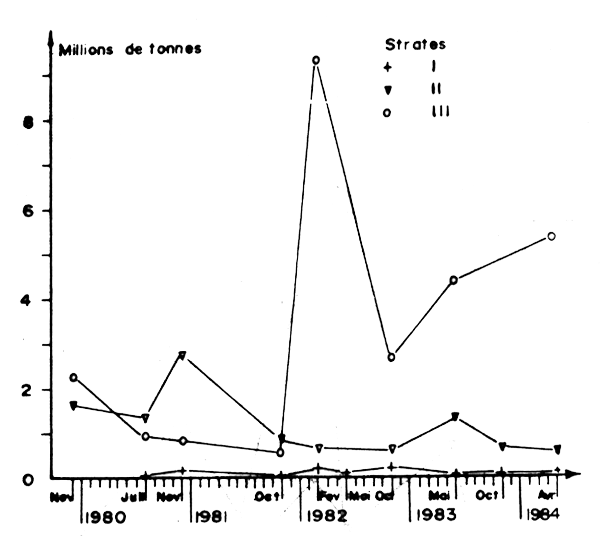
Figure 14 : Biomasse de sardines en millions de tonnes, estimé au sonar (d'après Lamboeuf et al., 1984)
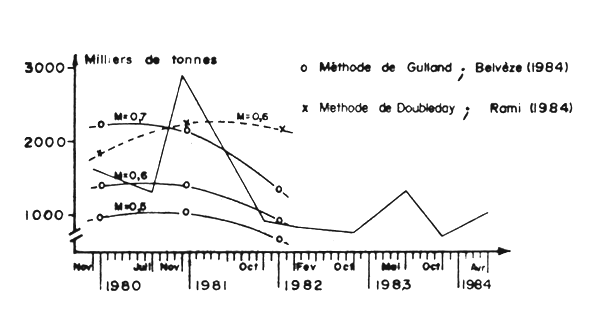
Figure 15 : Comparaison de la biomasse de sardine des strates I et II obtenues par analyse de cohorted et prospection sonar, (Lamboeuf et al., 1984)
| STRATES | NOV | JUIL | NOV | OCT | FEV | MAI | OCT | MAI | OCT | AVR | MOYENNE |
|---|---|---|---|---|---|---|---|---|---|---|---|
| 79 | 80 | 80 | 81 | 82 | 82 | 82 | 83 | 83 | 84 | ||
| IV | - | - | - | - | - | 281 | - | - | 121 | - | 201 |
| I | 0 | 25 | 154 | 18 | 199 | 76 | 176 | 29 | 56 | 34 | 77 |
| II | 1633 | 1302 | 2748 | 818 | 652 | - | 600 | 1296 | 668 | 573 | 1143 |
| III | 2318 | 950 | 863 | 564 | 9307 | - | 2665 | 4375 | - | 5185 | 3278 |
| I+II | 1633 | 1327 | 2902 | 836 | 851 | - | 776 | 1325 | 724 | 607 | 1220 |
| I+II+III | 3951 | 2277 | 3765 | 1400 | 10158 | - | 3441 | 5700 | - | 5792 | 4498 |
(Lamboeuf et al., 1984)
| STRATES | NOV | JUIL | NOV | OCT | FEV | MAI | OCT | MAI | OCT | AVR | MOYENNE |
|---|---|---|---|---|---|---|---|---|---|---|---|
| 79 | 80 | 80 | 81 | 82 | 82 | 82 | 83 | 83 | 84 | ||
| IV | - | - | - | - | - | 46 | - | - | 8 | - | 27 |
| I | 0 | 9 | 24 | 22 | 25 | 6 | 74 | 16 | 24 | 5 | 23 |
| II | 88 | 61 | 705 | 319 | 535 | - | 171 | 140 | 189 | 25 | 248 |
| III | 162 | 303 | 64 | 67 | 1741 | - | 255 | 796 | - | 215 | 464 |
| I+III | 88 | 70 | 729 | 341 | 560 | - | 245 | 156 | 213 | 30 | 271 |
| I+II+III | 250 | 373 | 793 | 408 | 2301 | - | 610 | 952 | - | 245 | 735 |
| géographiqueéde bancsémoyenneé | 79 – 84 | 79 – 84 | Moyennes 3 – 83 (1) | |||
|---|---|---|---|---|---|---|
| IV | Cap Spartel El Jadida | 45.000 | 200.000 | Augmenttation | Marco | 11.000 |
| I | El Jadida Cap Aglou | 19.000 | 80.000 | Stble | Maroc | 158.000 |
| II | Cap Aglou Cap Bojador | 122.000 | 100.000 | Diminutation | Maroc Espagne | 123.000 |
| III | Cap Bioador Cap Blanc | 246.000 | 3.200.000 | Augmentation | Pays de l' est | 158.000 |
| I+II | El Jadida | 387.000 | 4.380.000 | - | - | 450.000 |
| +III | Cap Blanc |
Dans la zone B, on remarque une tendance à la diminution de l'abondance (moyenne 1.200.000 t) et à l'augmentation dans la zone C (moyenne 3.000.000 t). Le Tableau 27 résume ces résultats.
Pour mémoire, l'abondance du stock Nord (Tanger, Casablanca), a été évalué à 200.000 tonnes au cours de deux campagnes; ce chiffre semble élevé au regard des captures, il est possible qu'il s'agisse d'une biomasse plurispécifique, mais le peu de données dont nous disposons ne permet pas de préciser davantage.
La variabilité des estimations est le résultat de variations naturelles de l'abondance et de l'accessibilité de la ressource aux instruments de détection.
La comparaison des estimations acoustiqued effectuées au sonar, aver les estimations d'autres navires montre qu'elles ont le même ordre de grandeur.
La comparaison avec les estimations obtenues jusqu'en 1982 par analyse de cohortes selon les méthodes de Gulland et de Doubleday, pour les zones A et B, montrent que la diminution de l'abondance dans cette région est confirmé par la méthode de Gulland, alors que celle de Doubleday ne présente qu'un inflechissement de la tendance (Figure 15).
Pour la zone C, les estimations au sonar sont très supérieures à celles obtenues par analyse de cohortes (Barkova et Domanevsky, Annexe 3). Il est possible que cette différence soit le résultat d'une surestimation du poids moyen des bancs dans cette région. A partir de 1983 une tendance à l'augmentation de la biomasse est decelée par les deux méthodes. Cette tendance semble se confirmer en 1984 par les évaluations acoustiques.
Les estimations obtenues au moyen du sonar sont dépendantes du choix de la valeur du poids moyen des bancs, et il convient donc de déterminer ce paramètre avec une bonne précision. Il faut donc considérer les valeurs absolues de ces estimations avec précaution. Elles permettent cependant de mettre en évidence des tendances dans un délai plus court que les autres méthodes.
Trois évaluations des mortalités par pêche et des abondances, effectuées par analyse de cohorte avec M = 0.6 ont été présentées au Groupe de travail (cf. §4.10).
Les deux premières, concernant l'unité de stock centrale (zone A + B), couvrent la période 1976–1982 (Belvèze 1984 ; Rami 1984). L'actualisation de ces évaluations n'a pas été possible pour le Groupe de travail, les captures par classe d'âge effectuées par les flottes espagnole et marocaine opérant dans la zone B n'étant pas disponibles pour 1983.
La troisième est présentée par Barkova et Domanevsky (voir Annexe 3) concernant l'unité de stock Sud (zone C), et couvre la période 1972–1983.
a) Dans le stock central (A + B)
Le taux instantané de mortalité par pêche a été évalué pour chaque année par deux méthodes différentes : celle de Gulland et celle de Doubleday, sur les âges pleinement recrutés, c'est à dire supérieurs à 1 an. Les deux méthodes mettent en évidence la très faible mortalité par pêche qui s'applique aux classes d'âge O chaque année. La classe d'âge I subit également une mortalité par pêche faible, en moyenne la moitié de celle qui s'applique aux classes d'âge pleinement recrutées (de deux à huit ans).
Les taux de mortalité annuels sur les sardines pleinement recrutées (2 ans et plus), calculés par les deux méthodes sont présentées au Tableau 28 suivant :
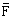 (1) (1) | F (2) | |
|---|---|---|
| 1976 | 0,33 | 0,22 |
| 1977 | 0,25 | 0,16 |
| 1978 | 0,37 | 0,20 |
| 1979 | 0,31 | 0,18 |
| 1980 | 0,17 | 0,20 |
| 1981 | 0,21 | 0,16 |
| 1982 | - | 0,28 |
Les taux de mortalité calculés par la première méthode sont en moyenne supérieurs à ceux calculés par la seconde, ce qui peut provenir du choix du F terminal dans la méthode de Gulland. Par contre si ce choix empêche d'avoir une donnée pour 1982, la méthode de Doubleday s'affranchit de cette contrainte et fournit pour cette dernière année une valeur plus élevée (0,28) du taux de mortalité par pêche. Le changement du diagramme d'exploitation de la pêcherie en 1982 où les bateaux marocains ont commencé à exploiter la zone B plus dense en sardine, et où une société mixte a également entrepris la pêche est la cause de cette élévation du taux de mortalité par pêche. Cet aspect devra être pris en compte dans les mesures d'aménagement à proposer pour la pêcherie.
Le taux d'exploitation E = F/Z varie d'une année à l'autre et d'une estimation à l'autre mais reste compris entre 0,21 et 0,38.
Les valeurs comprises dans cette fourchette sont inférieures aux taux d'exploitation atteints dans les autres pêcheries de petits pélagiques juste avant leur effondrement total.
b) Dans le stock Sud (zone C)
Dans cette pêcherie, le plein recrutement ne semble atteint qu'à partir de l'âge III et les taux de mortalité par pêche restent très faibles sur les classes plus jeunes. Une valeur très élevée du taux de mortalité par pêche terminal Ft = 0,8 a été adoptée pour le calcul, mais il n'y a
d'estimation d'un taux moyen de mortalité par pêche annuel ni du taux d'exploitation de la ressource. Cependant, ce taux peut se déduire du rapport entre la capture pondérale et la biomasse. Sur la base des données présentées des captures dans la zone C et des biomasses estimées par analyse de cohorte (Tableau 29) on peut constater que le prélèvement s'est accru fortement de 1972 à 1977, passant de 5% à 35% de la biomasse. A partir de 1978, la limitation d'accès à la resource à environ 30% de l'aire de distribution du stock et la diversification des captures vers les autres espèces pélagiques (chinchard et maquereau) font chuter le taux du prélèvement qui oscille jusqu'en 1983 entre 17% et 9% de la biomasse. On peut en déduire que depuis cette période le taux d'exploitation de la sardine dans la zone C est réduit, vraisemblablement la moitié de ce qu'il est dans la zone A où un tiers de la biomasse disponsible est capturée.
| Captures | Siomasse | Capture Biomasse | |
|---|---|---|---|
| 1972 | 64 | 1428 | 5% |
| 1973 | 244 | 2641 | 9% |
| 1974 | 436 | 3086 | 14% |
| 1975 | 429 | 2732 | 15% |
| 1976 | 647 | 2238 | 28% |
| 1977 | 570 | 1626 | 35% |
| 1978 | 222 | 1273 | 17% |
| 1979 | 114 | 1200 | 9% |
| 1980 | 207 | 1222 | 17% |
| 1981 | 196 | 1187 | 17% |
| 1982 | 164 | 1071 | 15% |
| 1983 | 174 | 1962 | 9% |
Les analyses de cohorte fournissent une évaluation du recrutement annuel dans les deux unités de stock.
a) Dans la population centrale (zone A + B)
Les recrutements obtenus par l'analyse de cohorte de 1976 à 1982 fluctuent de façon considérable d'une annéa à l'autre. Larégression non linéaire (méthode de Doubleday) fournit une évaluation plus rrécise que la méthode de Gulland en ce qui concerne les années les plus récentes.
Après deux faibles recrutements en 1976 et 1977 succède une alternance de forts et faibles recrutements jusqu'en 1982 (Figure 12).
Une comparaison entre ces valeurs et les captures par unité d'effort des classes d'âge ) et I additionnées sur une même cohorte (en millions de sardines) montrent une concordance étroite entre les deux séries de données de 1976 à 1982. Les CPUE des classes O + I peuvent donc servir d'indice de recrutement pour les années antérieurs à 1976.
Aux recrutements moyens de 1968 à 1969 succède un très fort recrutement en 1970 qui dominera la pêcherie jusqu'en 1974 et procurera les très fortes captures de 1973. A partir de 1971 les recrutements sont en déclin progressif jusqu'à la nouvelle augmentation de 1978–1982.
b) Dans l'unité de stock Sud (zone C)
Les variations de l'intensité de l'upwelling dans cette zone ont entrainé une augmentation rapide de la biomasse à partir de 1971 (cf. sec 4.10) suivie d'une chute non moins rapide. L'évolution du recrutement est à la base de ces larges variations d'abondance.
Les estimations d'abondance de la classe d'âge I obtenues par analyse de cohorte reflètent l'intensité du recrutement (classes d'âge O) l'année précédente. Pour fixer dans le temps l'apparition de ces recrutements il a donc été attribué l'année antérieure aux évaluations d'abondance de la classe I (Figure 16).
Après un pic de recrutement en 1972, un déclin rapide se manifeste jusqu'en 1976, suivi d'une stabilisation à un faible niveau de 1977 à 1981. En 1982 un nouveau très fort recrutement se manifeste, provoquant une augmentation de la biomasse évaluée par analyse de cohorte en 1983 ou par campagne de prospection acoustique en 1984.
a) Modèles globaux
Ces types de modèles sont basés sur les hypothèses restrictives suivantes :
le recrutement est considéré comme une fonction de la biomasse de géniteurs;
les fluctuations d'abondance sont dûes à la pêche et le stock tend constamment à revenir à un niveau d'équilibre théorique.
Ces hypothèses ne sont absolument pas vérifiées en ce qui concerne les stocks de sardine : recrutement variable, dépendant de l'hydroclimat et fluctuations d'abondance en grande partie indépendantes de la pêche. Le Groupe de travail a jugé qu'en raison de ces limitations d'une part et de la qualité des données d'effort et de que d'autre part il n'était pas possible d'utiliser ce type de modélisation. Cependant, avec une amélioration de la collecte des données concernant les efforts (en termes biologiques et économiques) et les relations entre abondance et hydroclimat, il serait possible d'obtenir non plus un modèle déterministe mais une famille de courbes correspondant à des conditions ambiantes et de capacité biologique du milleu différentes, ou un modèle multivariable (Garcia, 1984).
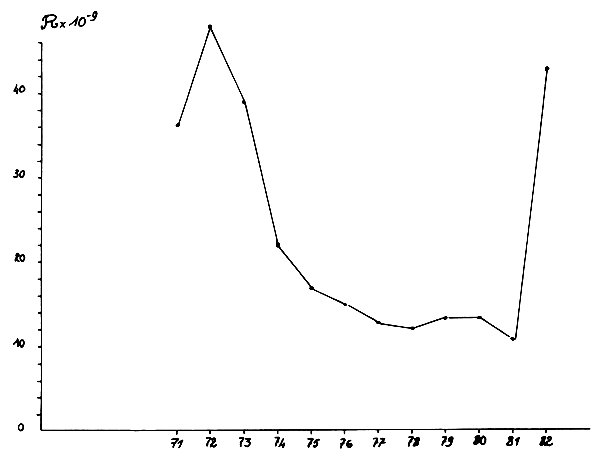
Fig. 16 : Evolution du recrutement de 1971 à 1982 dans la zone C
b) Modèles analytiques
Ces modèles permettent d'analyser les effets d'une modification du diagramme d'exploitation sur la production.
Les hypothèses de base du modèle de Beverton et Holt (1957) (F constant pour toutes les années, croissance isométrique des poissons) et l'inadaptation du modèle à l'analyse de deux pêcheries différentes exploitant le même stock ne permettent pas son utilisation pour la pêcherie de sardine dans les zones A et B.
Des simulations sont par contre possibles avec le modèle de Ricker plus souple d'emploi. Trois types de simulation ont été présentés par Belvèze au Groupe de travail.
La première concernant les effets d'une modification de l'âge de première capture sur la production pondérale totale et par bateau (Figure 17). Cette simulation montre qu'il n'y a aucun bénéfice à attendre d'un recul de l'âge de première capture quel que soit le niveau d'effort de pêche utilisé.
Cette conclusion est liée au taux élevé de mortalité naturelle qui fait que la biomasse d'une cohorte commence à décliner après l'âge I. Une augmentation de l'effort se traduit, pour un recrutement donné par une augmentation de la production totale et par diminution de la prise par bateau.
La seconde simulation concerne les interactions entre la pêcherie de type marocain et la pêcherie de type espagnol dans l'exploitation de la ressource (A + B) en termes de production. La Figure 18 montre les variations de production (en milliers de tonnes) à attendre de différents niveaux d'effort marocain en fonction de différents niveaux d'effort espagnol.
Si l'on veut maintenir le taux d'exploitation actuel qui ne doit pas augmenter dans cette zone, on peut voir sur cette figure qu'une augmentation de 2/3 de l'effort marocain nécessite une diminution de 90% de l'effort espagnol. Un doublement de l'effort de pêche type marocain nécessite, pour ne pas augmenter le taux d'exploitation de la ressource, la suppression totale de l'effort type espagnol. Tout plan d'aménagement de la ressource dans le contexte d'une extension de l'effort marocain dans la zone B devra prendre ces résultats en considération.
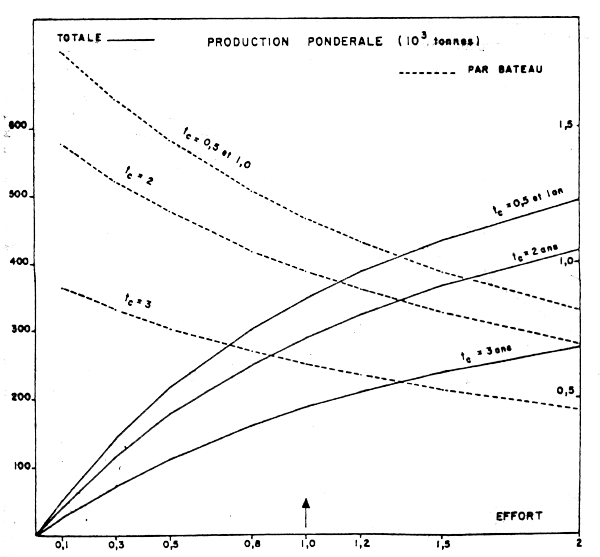
Figure 17 : Productions pondérales de la pêcherie (trait plein) et rendements annuels moyens pat bateau (trait tireté) en milliers de tonnes, en fonction de l'effort et pour différents âges à la première capture (In Belvèze, 1984)
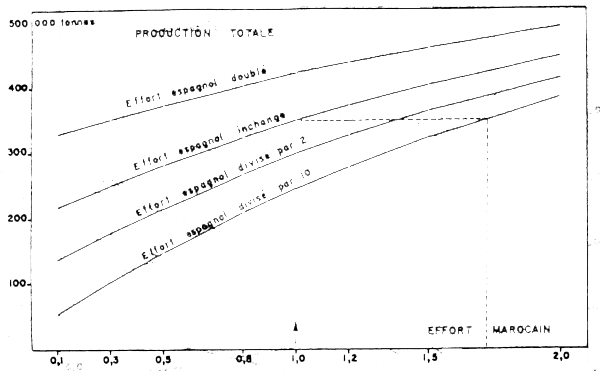
Figure 18 : Productions pondérale totale de la pêcherie en fonction de l'effort marocain pour divers niveaux d'effort espagnol (In Belvèze, 1984)
La troisième simulation concerne les évaluations de la biomasse féconde résiduelle (pourcentage de sardines femelles adultes survivantes par rapport à un stock non exploité) en fonction de l'effort de pêche. En conservant l'âge de la première capture actuel et le niveau d'effort actuel, la biomasse féconde résiduelle est de 58%. Si l'effort est double, cette biomasse féconde chutera à 39,5%. Il semble donc d'après ces résultats que la fécondité du stock ne soit pas le facteur limitant de l'exploitation. Il est admis qu'une biomasse féconde supérieure à 30% de la biomasse féconde du stock vierge n'entraine pas pour la plupart des stocks de problème de recrutement. En ce qui concerne les petits pélagiques les conditions naturelles de survie des oeufs et des larves influencent beaucoup plus le recrutement que le nombre d'oeufs pondus, et des explosions démographiques peuvent être observées pour des valeurs très basses de la biomasse féconde (Kawasaki, 1983; Tanaka, 1983).
![]()
![]()
![]()