![]()
![]()
![]()
Une masse importante de données sur la distribution, la biologie de la sardine, les stocks, les paramétres démographiques et sur la disponibilité de la ressource en fonction des facteurs de l'environnement a été acquise depuis le précédent Groupe de travail et permis d'améliorer la qualité des évaluations.
Aucun progrés n'a été constaté dans ce domaine depuis la tenue du Groupe de travail sur la standardisation des techniques de lecture d'âge sur la sardine (Santa Cruz de Tenerife, 15–20 Octobre 1979).
Le Groupe, aprés avoir constaté l'hétérogénéité des paramétres de croissance obtenus sur la sardine durant la période 1978–1984, insiste sur la necessité de mettre en place un programme effectif d'échanges d'otolithes et d'écailles suivi d'une réunion afin d'évaluer les résultats de ce programme.
En ce qui concerne l'échantillonnage, le Groupe a rappelé l'importance de la réalisation d'une évaluation des méthodes d'échantillonnages biologiques actuellement utilisées afin d'établir un programme de collecte de données applicables à chaque pays impliqué dans la pêcherie sardiniére.
La composition par classe de taille de captures, dans la zone A, des trois ports de Safi, Essaouira et Agadir depuis 1973 est présentée dans le Tableau 18 et la Figure 4.
La Figure 5 montre que la distribution annuelle de fréquences de tailles des sardines capturées, exprimées en pourcentage pour l'ensemble des trois ports, est unimodale pour les années impaires (1979, 1981, 1983) et bimodale pour les années paires (1980, 1982). Le mode dominant pendant une année correspond au creux entre les modes de la distribution de l'année précédente.
| Cm | 1973 | 1974 | 1975 | 1976 | 1977 | 1978 | 1979 | 1980 | 1981 | 1982 | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 7 | 14 | 16 | ||||||||||||||||||
| 7,5 | 625 | 2 | 0 | |||||||||||||||||
| 8 | 552 | 0 | - | 16 | 30 | 101 | ||||||||||||||
| 8.5 | 1 440 | -1 | 77 | 75 | 109 | 147 | ||||||||||||||
| 9 | 10 | 2 21 | 0.1 | 1 | 0 | 315 | 246 | 426 | ||||||||||||
| 9.5 | 139 | 5373 | 0.1 | 30 | 111 | 319 | 131 | 203 | 746 | 1 446 | 48 | |||||||||
| 10 | 109 | 6 203 | 0.1 | 4 105 | 0.1 | 169 | 500 | 327 | 611 | 916 | 2 162 | 72 | ||||||||
| 10.5 | 41 | 19 437 | 0.4 | 9 499 | 0.3 | 5 217 | 0.2 | 3 020 | 0.2 | 3 439 | 0.2 | 1 664 | 0.2 | 1 800 | 3 567 | 0.1 | 400 | |||
| 11 | 207 | 17 873 | 0.4 | 7 553 | 0.2 | 7 498 | 0.2 | 5 807 | 0.4 | 6 270 | 0.4 | 2 763 | 0.1 | 5 586 | 0.1 | 744 | ||||
| 11.5 | 64 | 29 212 | 0.6 | 9 690 | 0.3 | 18 659 | 0.6 | 9 133 | 0.6 | 11 542 | 0.7 | 5 129 | 0.1 | 3 335 | 0.1 | 10 308 | 0.2 | 1 400 | 0.1 | |
| 12 | 245 | 40 747 | 0.9 | 14 505 | 0.5 | 26 696 | 0,8 | 14 019 | 0,9 | 30 110 | 1.9 | 8 172 | 0.2 | 23 464 | 0.5 | 14 948 | 0.3 | 3 090 | 0.1 | |
| 12.5 | 479 | 80 515 | 1.7 | 24 567 | 0.8 | 39 556 | 1.2 | 21 498 | 1.3 | 54 764 | 3.5 | 13 779 | 0.4 | 81 566 | 1.8 | 31 722 | 0.7 | 19 677 | 0.9 | |
| 13 | 1 418 | 132 812 | 2.9 | 42 724 | 1.4 | 54 069 | 1.6 | 37 842 | 2.4 | 74 453 | 4.7 | 29 929 | 0.8 | 190 272 | 4.3 | 66 309 | 1.5 | 71 974 | 3.2 | |
| 13.5 | 4 560 | 0.1 | 204 992 | 4.5 | 101 271 | 3.3 | 64 134 | 1.9 | 53 877 | 3.4 | 99 214 | 6.3 | 52 355 | 1.4 | 304 602 | 6.9 | 119 720 | 2.8 | 147 500 | 6.5 |
| 14 | 33 517 | 0.6 | 270 772 | 5.9 | 166 704 | 5.4 | 62 946 | 1.9 | 73 436 | 4.6 | 137 766 | 8.8 | 94 881 | 2.6 | 495 500 | 11.2 | 165 967 | 3.0 | 215 144 | 9.5 |
| 14.5 | 113 922 | 109 | 301 103 | 7.8 | 187 880 | 6.0 | 71 033 | 2.1 | 8 910 | 4.9 | 132 575 | 5.9 | 199 559 | 5.5 | 423 232 | 9.5 | 207 578 | 4.6 | 269 576 | 11,9 |
| 15 | 269 632 | 4.6 | 336 790 | 7.3 | 146 995 | 4.7 | 58 130 | 1.7 | 89 475 | 5.6 | 92 175 | 5.9 | 199 559 | 5.5 | 423 232 | 9.5 | 207 678 | 4.8 | 206 808 | 9.1 |
| 15.5 | 362 901 | 6.2 | 279 639 | 6.1 | 166 014 | 5.3 | 84 205 | 2.5 | 91 428 | 5.7 | 47 146 | 3.0 | 262 769 | 7.2 | 288 928 | 6.5 | 291 073 | 140 452 | 6.2 | |
| 16 | 419 610 | 7.2 | 198 463 | 4.3 | 188 995 | 6.1 | 111 215 | 3.3 | 102 402 | 6.4 | 22 147 | 1.4 | 347 368 | 9.6 | 212 583 | 5.2 | 384 103 | 8.2 | 100 359 | 4.4 |
| 16.5 | 433 513 | 7.4 | 183 814 | 4.0 | 247 445 | 7.9 | 160 327 | 4.7 | 97 204 | 6.0 | 17 843 | 1.1 | 458 745 | 12.2 | 180 689 | 4.1 | 479 099 | 11.0 | 87 880 | 3.9 |
| 17 | 576 121 | 9.8 | 208 007 | 4.5 | 236 637 | 7.6 | 237 779 | 7.0 | 109 981 | 6.8 | 20 983 | 1.3 | 593 144 | 16.4 | 193 028 | 4.3 | 531 112 | 12.2 | 100 046 | 4.4 |
| 17.5 | 695 486 | 11.9 | 257 761 | 5.6 | 251 525 | 8.1 | 329 162 | 9.7 | 125 195 | 7.8 | 32 540 | 2.1 | 519 171 | 14.3 | 248 503 | 5.6 | 543 752 | 12.5 | 114 204 | 5.0 |
| 18 | 882 526 | 15.0 | 302 490 | 6.6 | 263 439 | 8.5 | 415 601 | 12.3 | 167 868 | 10.4 | 54 443 | 3.5 | 396 555 | 10.9 | 347 583 | 7.8 | 472 367 | 10.0 | 161 869 | 7.1 |
| 18.5 | 740 193 | 12.6 | 394 373 | 8.6 | 334 983 | 10.8 | 466 005 | 13.8 | 171 461 | 10.7 | 71 143 | 4.5 | 208 362 | 5.7 | 303 174 | 8.2 | 371 953 | 8.6 | 182 840 | 8.1 |
| 19 | 599 019 | 10.2 | 387 449 | 8.4 | 281 824 | 9.0 | 407 712 | 12.0 | 141 308 | 8.8 | 82 229 | 5.2 | 87 773 | 2.4 | 257 586 | 5.8 | 220 233 | 5.1 | 168 356 | 7.4 |
| 19.5 | 378 243 | 6.4 | 384 914 | 8.4 | 210 048 | 6.7 | 332 677 | 9.8 | 95 118 | 5.9 | 88 883 | 5.7 | 48 273 | 1.3 | 136 780 | 3.1 | 125 658 | 2.9 | 125 378 | 5.5 |
| 20 | 224 143 | 3.8 | 261 248 | 5.6 | 123 562 | 4,0 | 218 725 | 6,5 | 64 811 | 4,0 | 97 361 | 6,2 | 17 736 | 1,0 | 65 528 | 1,5 | 40 642 | 1,1 | 82 189 | 3,6 |
| 20.5 | 88 283 | 1,5 | 151 441 | 3,3 | 63 786 | 2,0 | 118 239 | 3,5 | 31 762 | 2,0 | 119 824 | 7,6 | 38 410 | 1,1 | 36 923 | 0,8 | 23 783 | 0,5 | 41 777 | 1,8 |
| 21 | 31 949 | 0,5 | 55 170 | 1,2 | 22 911 | 0,7 | 59 528 | 1,8 | 12 409 | 0,8 | 121 468 | 7,7 | 31 261 | 0,9 | 21 301 | 0,5 | 7 984 | 0,4 | 19 989 | 0,9 |
| 21.5 | 6 889 | 0.1 | 18 895 | 0,4 | 5 683 | 0,2 | 23 254 | 0,7 | 4 194 | 0,3 | 84 758 | 5,4 | 24 849 | 0,7 | 12 045 | 0,3 | 5 531 | 0,1 | 6 324 | 0.3 |
| 22 | 2 045 | 4 861 | 0,1 | 932 | 12 010 | 0,4 | 1 630 | 0,1 | 43 968 | 2,8 | 14 982 | 0,4 | 5 963 | 0,1 | 1 556 | 1 309 | 0,1 | |||
| 22.5 | 688 | 1 476 | 117 | 3 382 | 0,1 | 596 | 16 317 | 1,0 | 8 417 | 0,2 | 1 002 | 1 648 | 568 | |||||||
| 23 | 227 | 89 | 44 | 445 | 347 | 4 808 | 0,3 | 1 982 | 0,1 | 256 | 126 | 47 | ||||||||
| 23.5 | 109 | 74 | 434 | 1 241 | 0,1 | 614 | 39 | 59 | ||||||||||||
| 24 | 78 | 343 | 690 | 6 | ||||||||||||||||
| 24.5 | 62 | 298 | 91 | 68 | ||||||||||||||||
| 5 866 368 | 4 601 176 | 3 115 504 | 3 389 106 | 1 607 047 | 1 570 558 | 3 625 384 | 4 440 286 | 4 346 438 | 2 270 021 | |||||||||||
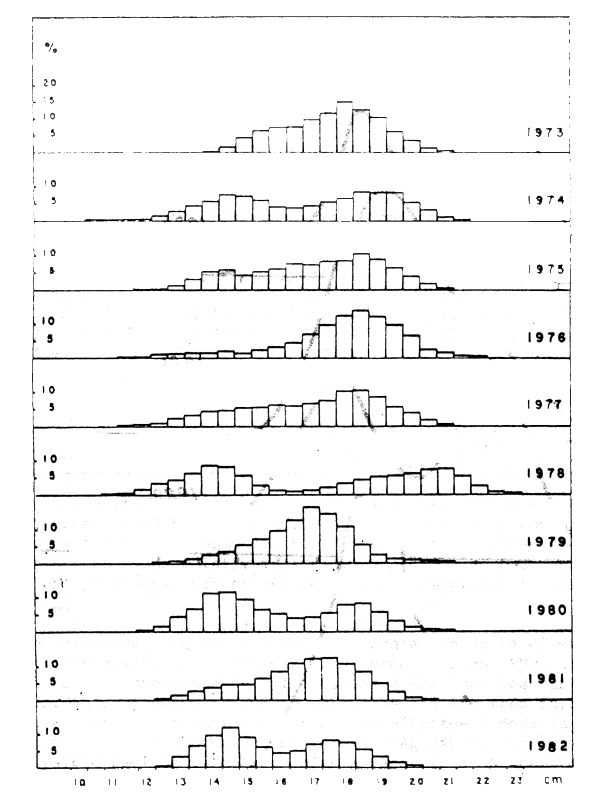
Figure 4 : Distribution relative (en pourcentages) des captures de sardines par classes de taille de 1/2 centimètres dans la zone A (Safi, Essaouira et Agadir) de 1973 à 1982
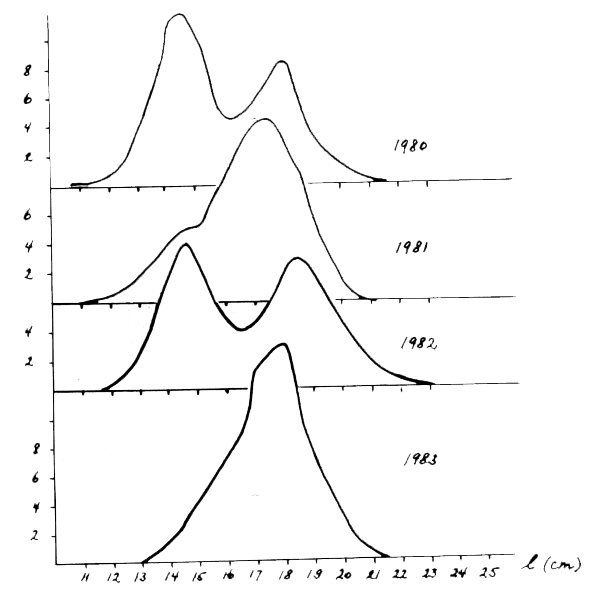
Figure 5 : Variation de la distribution des fréquences des tailles de la capture totale de la sardine (Safi, Essauira et Agadir) entre 1980 et 1983
La présence de sardine de petite taille, dans les captures de la la zone A, se manifeste en moyenne une année sur deux et particuliérement dans les années impaires.
| Anné | Maroc (zone A, A et en 1983) | Espagne(zone B) | Roumanie (zone C) | URSS (zoneC) | Pologne (zone ) | RDA (zone C) |
|---|---|---|---|---|---|---|
| 1972 | II III | II III | I | |||
| 1973 | I II III | III IV | II III | II III | II | |
| 1974 | I II III | III IV | II III | II III | III | |
| 1975 | II III | III IV | III IV | III IV | III | |
| 1976 | I II | II III IV | III IV V | III IV | III IV | |
| 1977 | 0 I II | II III | III IV | III IV | III IV | |
| 1978 | 0 II III | I II III | III IV V | |||
| 1979 | I II | I II III | - | III IV | ||
| 1980 | 0 I II | I II | - | III IV | ||
| 1981 | 0 I II | I III | - | III IV | ||
| 1982 | 0 I II | I II IV | - | III IV | ||
| 1983 | I III | IV V | III IV |
La composition en âge des captures est nettement différente dans les trois zones de pêche A, B et C. Dans la zone C les groupes d'âge III et IV prédominent dans les captures. Par contre dans les zones A et B oú sont exploités des poissons relativement plus jeunes, la composition par âge varie selon les années. Les prises de la zone A montrent les plus jeunes poissons (groupe 0).
Par ailleurs, une trés faible représentation de la classe d'âge zéro a été enregistrée dans les captures marocaines en 1983. Comme on ne connait pas l'importance de cette classe d'âge dans les captures espagnoles, on peut supposer que le recrutement a été trés faible cette année-lá. En effet, les captures marocaines effectuées dans la zone B et destinées á alimenter les usines de farine d'Agadir comportent également une trés faible part de sardine de classe zéro. Il semblerait donc qu'il ne s'agisse pas d'un phénoméne de disponibilité mais plutôt d'un manque de recrutement. Si on se référe aux années antérieures, on constate depuis 1977 une alternance d'années de fort et de faible recrutement dans le stock (voir paragraphe 4.10.3).
Le modéle de von Bertalanffy a été utilisé pour décrire la croissance de la sardine des zones de pêche A, B et C. Dans l'ensemble (Tableau 20), les résultats obtenus sont hétérogènes et traduisent la diversité des données de base et des méthodes de calcul qui ont servi à étudier la croissance. En effet, le modéle a été ajusté soit à des cohortes individuelles et moyennes (Belvéze, 1984), soit à des longueurs moyennes par âge observés dans les captures (Belvèze, 1984/zone A) ou dans les échantillons (zone B: Zouiri, sous presse; Delgado et al., 1980; zone C: Barkova et al., Annexe 3). On peut remarquer que la croissance dans la zone C est la plus forte (22,21 t<L∞ <25,6).
| Zone A 0,441<K<0,828 | 20,02<L∞ cm<21,82 | -1,33<t0 année<-0,38 |
| (1973-1982) | ||
| (Belvèze, 1984) | ||
| (Belvèze et Rami, 1978) | ||
| Zone B 0,370<K<0,93 | 19,44<L∞ <22,65 | -1,68<t0 année<-0,35 |
| (1979-1983) | ||
| (Zouiri) | ||
| (Delgado et al., 1980) | ||
| Zone C0,274<K<1,080 | 22,16<L∞ <25,6 | -3,03<t0 année<-0,205 |
| (1979-1983) | ||
| (Zouiri) | ||
| (Barkova, et al., Annexe 3) | ||
| (Staicu et Maxim, Annexe 4) |
Le Groupe a surtout retenu que la comparaison des paramètres de von Bertalanffy n'avait pas une grande signification biologique : toutefois les données issues du modèle c'est à dire les longueurs moyennes estimées par âge devraient être comparées entre elles car à des valeurs de L∞ , K et t0 différentes, peuvent correspondre des tailles moyennes par âge semblables.
La mortalité totale a été estimée (Z = 0,80) pour le stock central (zone A + B) à l'aide de modéle de Chapman et Robson (1960) appliqué aux captures dez zones A + B, ventilées par âge (Rami, 1984). Par analyse de la courbe de capture, Belvèze (1984) a obtenu Z = 0,85.
Deux estimations indépendantes de la mortalité naturelle M ont été données par Belvèze (1984). La première, M = 0,65 a été obtenue à l'aide de la relation qui lie la mortalité naturelle à l'âge maximum (Beverton et Holt, 1959). La seconde, M = 0,56 a été obtenue selon une équation développée par Pauly (1980). Rami (1984) a estimé M = 0,60 à partir des résultats des campagnes acoustiques de l'ISPM et des taux de mortalité totale Z présentés ci-dessus. Cet auteur a également obtenu M = 0,63 en appliquant la technique de Paloheimo (1961, 1980) à la classe d'âge 1976, la seule qui ait permis de suivre une cohorte compléte dans les captures durant la période d'étude. Il est à noter que cette estimation basée principalement sur les données de CPUE en nombre par âge est d'une part comparable à toutes les valeurs présentées ci-dessus et d'autre part proche de M = 0,59 obtenu à l' aide d'expériences de marquage réalisées sur la sardine du Sud-Ouest africain (Newman, In Crawford, 1979).
En conclusion, on adoptera M = 0,6 d'autant plus que cette valeur a étéutilisée dans les analyses de cohortes effectuées par les chercheurs de la RDA, de l'URSS et d'Espagne.
Une compilation des données nouvelles (Staicu et Maxim, Annexe 4, Belvèze 1984, Delgado et Fernandez 1983, Zouiri, sous presse) est présentée au Tableau 21. Les exposants de la taille dans la relation sont régulièrement supérieurs à 3 (compris entre 3,2 et 3,4) sauf en ce qui concerne les sardines échantillonnées au cours des campagnes de prospection acoustique et pour lesquelles l'exposant est inférieur ou égal à 3. Ces derniers échantillons ont été congelés à bord ; la taille et le poids des individus ont été mesurés au laboratoire après décongélation; il est probable que cette différence avec les échantillons mesurés à l' état frais soit liés à une perte de poids à la décongélation. Le Groupe de travail a convenu de ne prendre comme relation taille-poids que celles issues des mesures sur le poisson frais. Le fait que l'exposant de la longueur soit supérieur à 3 indique une allométrie de croissance. L'emploi du modéle analytique de Beverton et Holt qui présuppose une croissance isométrique (exposant = 3) n'est donc pas souhaitable pour la sardine marocaine.
| Zone A | ||
| Belvèze, 84 | ||
| Résultats globaux 76-82 | 0,003066 | 3,353 |
| (ler tr. | 0,00295 | 3,19 |
| (2ème tr. | 0,00226 | 3,25 |
| Rami, 1980 | ||
| (3ème tr. | 0,000898 | 3,24 |
| données 78 | ||
| (4 ème tr. | 0,00103 | 3,40 |
| Rami, 84, | ||
| 1976–82 | 0,003477 | 3,33 |
| Zone B | ||
| Zouiri, sous presse : | ||
| Acoustique, Mai 83 | 0,0097 | 2,9205 |
| Acoustique, Octobre 83 | 0,0076 | 3,0158 |
| Delgado-Fernandez | ||
| 1976–1982 | 0,00357 | 3,20506 |
| Zone C | ||
| Acoustique, Mai 83 | 0,0132 | 2,8492 |
| Staicu et Maxim 1973 | 0,055 | 2,536 |
| (Annexe 4) 1974 | 0,018 | 2,824 |
| 1975 | 0,009 | 3,098 |
| 1976 | 0,021 | 2,827 |
| 1977 | 0,009 | 3,106 |
| 1978 | 0,009 | 3,114 |
| 1983 | 0,016 | 2,910 |
La sardine a développé une stratégie de reproduction assurant les meilleurs chances de survie des larves pendant leurs premières semaines d'existence. Des études comme celles de Lasker (1978), Parrish et al., (1981) ont analysé les conditions d'environnement assurant la survie ou la perte des larves des petits pélagiques dans les zones d'upwelling côtier.
La localisation géographique et les époques où se produisent la majeure partie de la ponte assurent un minimum de perte des larves par dérive vers le large et par famine lorsque le brassage d'eau disperse les micro-strates de particules alimentaires.
Dans les zones A et B, la période de reproduction est essentiellement l'hiver (Belvèze 1984 ; Delgado et Fernandez 1983) c'est à dire l'époque où les alizé s sont les plus faibles et l'upwelling est minimum. Dans la zone C, la ponte de la sardine s'effectue à deux périodes : principalement Octobre-Décembre et secondairement Avril-Mai, coindidant avec deux périodes d'affaiblissement de l'upwelling (Barkova et Domanevsky, Annexe 3).
Les zones de reproduction (Figure 1) sont localisées aux endroits où la côte prend une orientation vers l'Ouest et où le plateau continental est large.
Les larves de sardine ont le temps d'effectuer leur développement et leur métamorphose avant d'être rejetées hors du plateau continental par la dérive des eaux superficielles qui est orientée approximativement à 45° par rapport à la côte dans ces zones-là.
On peut trouver des oeufs et des larves de sardine toute l'année et tout au long de la côte entre 35° et 20°N mais les concentrations maximales se trouvent en hiver dans trois zones définies : entre Casablanca et Rabat, entre Agadir et Cap Juby et entre le Cap Bojador et le Cap Barbas.
Quatre principales aires de distribution de la sardine ont été identifiées et localisées lors de douze campagnes de prospection acoustique effectuées dans la période 1979–1984. Les limites de séparation de ces aires de distribution coincident avec les zones où le plateau continental est le plus étroit.
Trois zones de concentration sont permanentes : la zone Nord entre Casablanca et Larache, la zone B entre 26° et 30°N et la zone C entre 23° et 25°N. La quatrième zone (A), située entre 30° et 32°N n'est pas observée en hiver et correspond à une migration vers le Nord d'une partie de la B au printemps (Figures 1 et 2).
Des échanges entre le stock C (maximum d'abondance en hiver et au printemps) et le stock B (maximum d'abondance en été et en automne) ont été également décrits (Ikzarn, 1984).
En résumé, les aires de distribution s'étendent vers le Nord en été/automne et vers le Sud en hiver et au printemps. Ces mouvements plus ou moins saisonniers sont en accordance avec le déplacement des flottilles de pêche polonaises et soviétiques qui suivent les concentrations de poisson dans la zone C (Barkova et al., 1981 ; Krzeptowski 1978).
On peut dire que 80% des concentrations se situent entre les isobathes 20 et 50 m en toutes saisons. En automne, on observe un large déplacement vers la côte et vers le large en été.
Le niveau moyen d'immersion de la sardine en bancs le jour est situé à une profondeur plus grande que celui des poissons dispersés de nuit. L'amplitude verticale des concentrations des poissons est plus faible le jour que la nuit (Ikzarn, 1984).
Le Groupe a rappelé que les chercheurs soviétiques ont commencé à étudier systématiquement les ressources de la région Nord du COPACE dès 1957 et que les travaux menés sur une base annuelle à partir de 1960 entre Dakar et Tan-Tan ont pu montrer que les concentrations de sardine étaient absentes au Sud de la région Cap Juby-Cap Bojador. Ce n'est que vers 1967 qu'apparurent les premiers indices de migration de la sardine vers le Sud en liaison avec l'apparition de conditions hydroclimatiques et trophiques favorables au large des côtes du Sahara.
Trois zones de reproduction sont définies entre Gibraltar et le Cap Blanc (Figure 1). Les zones de concentration et de pêche sont au nombre de quatre : zone Nord (entre Casablanca et Larache), Zone A, Zne B et zone C. La zone A est en grande partie désertée par les sardines en hiver. L'importance des captures qui sont réalisées en été laisse penser que les sardines proviennent de la zone B où l'essentiel de la biomasse se concentre en hiver.
Un certain nombre de critères ont été présentés par Belvèze (1984) concenrnant l'identification des populations. Si aucun ne peut séparement apporter de preuve définitive, l'ensemble permet néanmoins de tirer des conclusions acceptables.
Les observations, basées sur la biométrie ou sur des données biologiques peuvent se ré sumer dans un tableau :
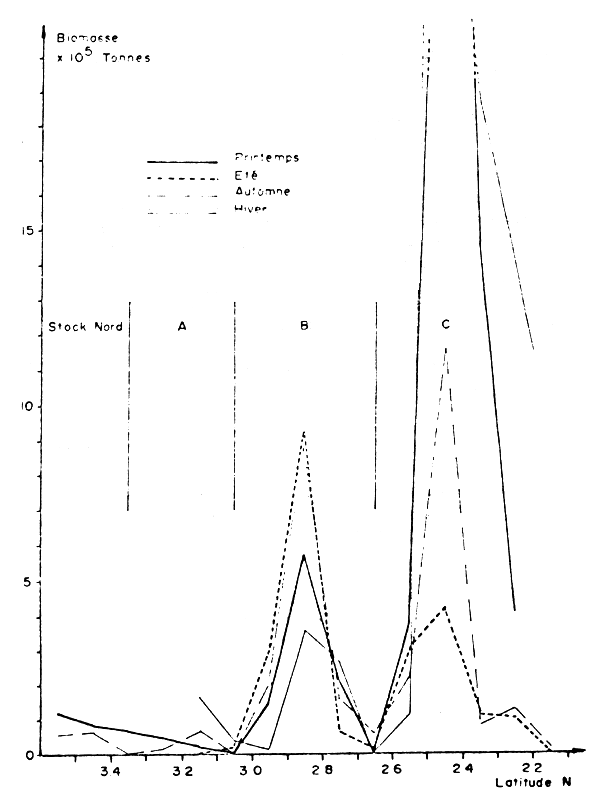
Figure 6 : Répartition de la biomase de sardine en fonction de la latitude
| Critères | ZONES | |||
|---|---|---|---|---|
| Nord | A | B | C | |
| ReproduPêche | X | X | X | X |
| Moyennes vertébrales | ≠ | = | = | ≠ |
| Indice céphalique | ≠ | = | = | = |
| Longueur prédorsale | ≠ | = | = | ≠ |
| Longueur préanale | = | = | = | ≠ |
| Parasitisme | X | 0 | 0 | 0 |
| (Peroderma cylindrium) | ||||
| Croisance | = | = | ≠ | |
L'examen de ce tableau montre qu'il n'y a pas de différence ( = ) pour chaque paramètre observé entre les sardines capturées dans les zones A et B.
Par contre, des différences significatives (≠) existent avec les sardines capturées dans la zone Nord et dans la zone C.
En conclusion, trois unités de stock, correspondent aux trois aires de reproduction : une petite population au Nord de Casablanca, une population qui se reproduit dans la zone B et qui alimente en été la pêcherie A, et une population dans la zone C, appelée population “saharienne” par Barkova et Domanevsky. D'après une communication de M. Koukhorenkho au Groupe de travail, cette dernière population n'existait pas en 1960. La colonisation de la zone C par la sardine aurait débuté en 1967, et se serait fortement développé au cours des années 1968–1972.
Les CPUE tendent à diminuer dans la zone A et à augmenter dans la zone B. Le manque de standardisation entre les puissances de pêche des bateaux constituant les différentes flottes ne permet pas l'expression d'un effort de pêche global pour l'ensemble des pêcheries.
Deux analyses de cohorte ont été présentées basées sur les captures par classes d'âge marocaines et espagnoles additionnées et extrapolées aux captures totales pour la période 1976–1982 (Belvèze, 1984 ; Rami, 1984).
La première utilise la méthode de Gulland (1965) avec M = 0,6 et Ft = 0,3 pour les cohortes entièrement pêchées. La biomasse moyenne au cours de l'année est calculée en multipliant les abondances de chaque classe d'âge par les poids moyens obtenus dans l'échantillonnage des captures à Agadir.
La seconde utilise la méthode de Doubleday (1976) basée sur une régression non linéaire des prises par âge. La biomasse au début de l'année est calculée en multipliant les abondances par un poids moyen théorique pour chaque classe d'âge issues de la courbe de croissance moyenne et de la relation taille-poids moyenne.
Les résultats (Tableau 23) montrent une variation identique des biomasses annuelles sauf pour les deux dernières années.
La biomasse est en diminution à partir de 1976. Elle commence à réaugmenter dès 1979, mais si cette augmentation se poursuit jusqu'en 1982 avec la méthode de calcul de Doubleday, par contre la méthode de Gulland met en évidence un tassement de la ressource dès 1980.
| Année | Méthode de Gulland Biomasse moyenne au cours de l'année | Méthode de Doubleday Biomasse exploitable au début de l'année |
|---|---|---|
| 1976 | 979 | 1450 |
| 1977 | 705 | 1131 |
| 1978 | 788 | 948 |
| 1979 | 1282 | 1159 |
| 1980 | 1159 | 1379 |
| 1981 | 1158 | 1602 |
| 1982 | 951 | 1750 |
Les différences observées dans l'évaluation de la biomasse par ces deux méthodes semblent principalement provenir de l'estimation des poids moyens attribués aux sardines d'âge O recrutées à la pêcherie. La biomasse totale au début de l'année calculée par la méthode de Doubleday montre cependant une tendance à la diminution en 1982 ce qui semble, avec un certain décalage en accord avec la méthode de Gulland.
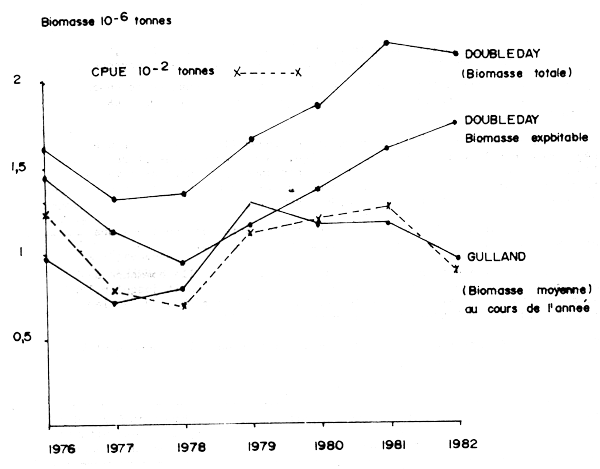
La comparaison de ces biomasses avec les CPUE enregistrées dans la zone A (Figure 7) met en évidence le parallèlisme des évolutions. Néanmoins les CPUE dans la zone A diminuent en 1982 comme la biomasse estimée par la méthode de Gulland.
Une relation étroite a étémise en évidence par Belvèze (1984) entre le coefficient de condition moyen annuel des sardines capturées dans la zone A et la biomasse moyenne annuelle de l'ensemble de la population. Une relation identique avait été mise en évidence par Mac Grégor (1959) sur la sardine de Californie. Cette relation :
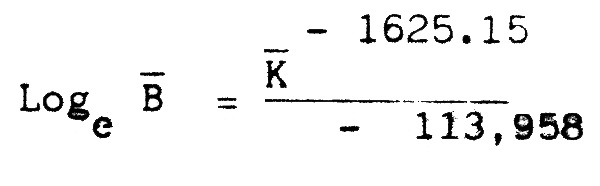
permet de calculer une estimation de biomasse pour les années antérieures à l'analyse de cohorte. Les données sont disponibles de 1971 à 1982, excepté pour l'année 1975 (Figure 8a).
L'évaluation de biomasse par la méthode de Gulland suit très étroitement les coefficients de condition moyens observés en 1976 à 1982 (Figure 8a).
La biomasse du stock central (A + B) a décliné fortement de 1971 à 1977, s'est accrue rapidement de 1977 à 1979 et redécline à nouveau jusqu'en 1982.
Les variations du coefficient de capturabilité q de la relation F = qf ont été établies sur la base des taux de mortalité par pêche dues à la seule flotte marocaine et l'effort effectif dans la zone A (Belvèze, 1984).
| Année | Effort effectif“f” | Mortalité par pêche “F” | 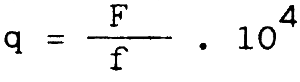 |
|---|---|---|---|
| 1976 | 1429.8 | 0,159 | 1,112 |
| 1977 | 1306.0 | 0,075 | 0,574 |
| 1978 | 1324.8 | 0,107 | 0,808 |
| 1979 | 1456.7 | 0,118 | 0,810 |
| 1980 | 1515.8 | 0,120 | 0,792 |
| 1981 | 1576.6 | 0,104 | 0,660 |
Le coefficient de capturabilité q est loin d'être constant. Le diagramme d'exploitation de cette pêcherie n'a pas évolué au cours de cette période, par contre la tendance qui se dégage est une diminution de q de 1976 à 1981. Cet affaiblissement du coefficient de capturabilité peut être la conséquence d'une diminution de la disponibilité en sardine dans la zone A.
L'étude des variations mensuelles du carré de la vitesse moyenne des vents alizés qui engendrent l'upwelling au centre de la zone A met en évidence la relation étroite entre le phénomène et la disponibilité locale en sardine (Figure 8b).
Figure 8a: Biomasse moyenne estimée par l'analyse de cohorte pour M = 0,6 et Ft = 0,3 en milliers de tonnes (trait tireté) et biomasse “prédite” par la relation entre le coefficient de condition moyen et la biomasse moyenne au cours de l'année (trait plein) (Belvèze, 1984)
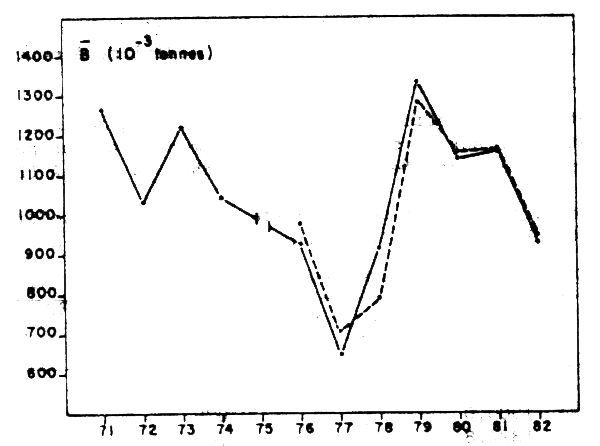
Figure 8b: Carré des vitesses moyennes mensuelles des vente de Nord et de Nord-Est à Essaouira et CPUE locales moyennes dans la zone Safi-Essaouira de 1968 à 1982 (Belvèze, 1984)
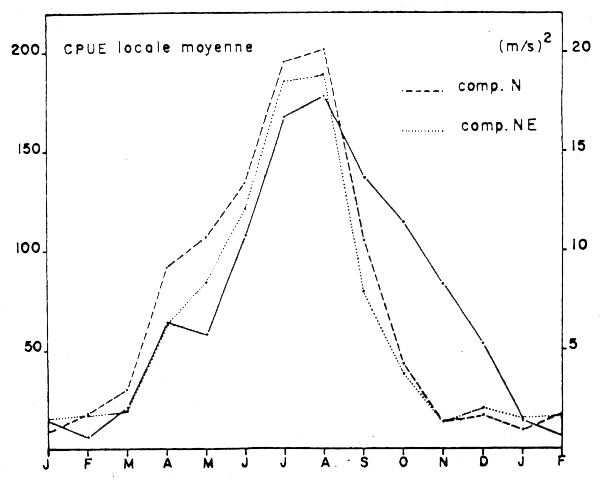
Les études réalisées au cours du programme CINECA (Grall et al., 1982) montrent que l'énergie transmise à l'océan et provoquant la remontée des eaux profondes provient essentiellement des vents du Nord-Nord/Est (alizés). Les enregistrements anémométriques effectués à Essaouira, c'est à dire au cours de l'upwelling de la zone A, de 1945 à 1980 montrent un affaiblissement progressif de la vitesse moyenne des vents alizés depuis 1959 (Figure 9), ainsi qu'une tendance à une orientation vers l'Est de la direction moyenne d'où soufflent ces vents. L'upwelling résultant tend donc à diminuer d'intensité depuis cette date et plus particulièrement depuis 1975.
La diminution de la disponibilité en sardine dans la zone A marquée par la baisse du coefficient de capturabilité est liée en grande partie à cet affaiblissement de l'intensité de l'upwelling estival, dans cette zone.
Inversement dans la zone B, les CPUE espagnoles augmentent depuis 1970. Les enregistrements des vitesses des vents ne sont pas disponibles et il a été nécessaire de calculer un indice d'upwelling mensuel basé sur une estimation des vents géostrophiques engendrés par les gradients de pression athmosphérique, selon la méthode mise au point par Bakun (1973). Les résultats exprimés en tonnes déplacées par seconde et par 100 mètres de littoral mettent en évidence une augmentation de cet indice d'upwelling de 1950 à 1980 et plus particulièrement depuis 1970, (Figure 10).
Il apparait donc que les variations observées de la disponibilité en sardine dans les zones A et B soient liées à des évolutions différentes de l'intensité de l'upwelling dans ces deux zones : affaiblissement au Nord et renforcement au Sud.
Dans la zone C, les fluctuations d'abondance de la population de sardine sont d'une très grande amplitude. Les données disponibles sont:
les captures de 1969 à 1983 ;
une évaluation de la biomasse par analyse de cohorte présentée au Groupe de travail par Barkova et Domanevsky de 1972 à 1983 ;
un indice annuel d'upwelling en tonnes/seconde pour 100 mètres de côte publié par Sedykh et al., (1979);
les anomalies de température de surface calculées par Belvèze (1984).
La preprésentation graphique de l'évolution de ces différents paramètres (Figure 11) montre un accroissement important de l'upwelling de 1969 à 1973.
Figure 9 : Vitesse et directions moyennes annuelles du vent à Essaouira de 1945 à 1980 (Belvèze, 1984
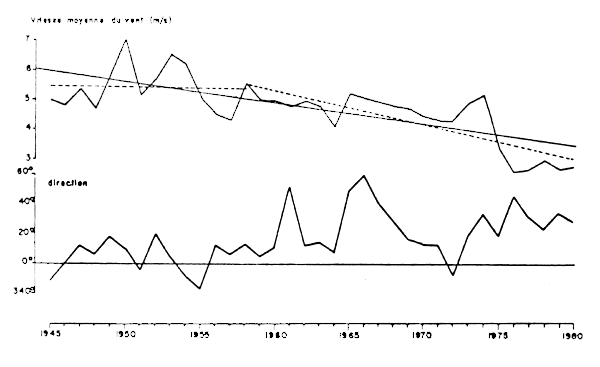
Figure 10 : Moyennes annuelles de l;indice d'upwelling de 1946 à 1981, par 28°N et 13°W et tonne/seconde pour 100 mètres de cote calculées d'après les données mensuelles de Bakun (Belvèze, 1984)
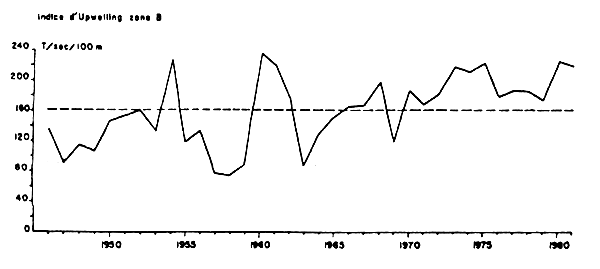
Figure 11: Anomalies des temperaturos de surface
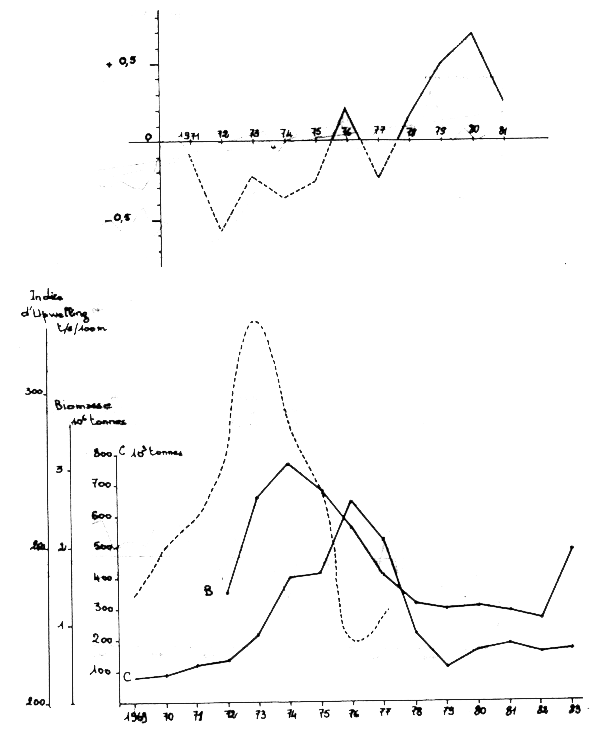
Il y a une bonne concordance entre les indices d'upwelling et les anomalies de température de surface pendant la période de recouvrement de ces deux séries d'observations (1971–1977).
La biomasse du stock évolue parallèlement à l'intensité de l'upwelling mais avec un décalage d'un an ce qui est normal si on fait l'hypothèse que l'upwelling a une action sur le recrutement. Les captures s'accroissent également mais avec un décalage de deux ans par rapport à la biomasse, les classes d'âge principalement exploitées étant celles de III et IV ans.
L'affaiblissement de l'upwelling s'amorce en 1974 et se prolonge jusqu'en 1980 d'après les anomalies de températures de surface. La biomasse chute dès 1975, et les captures qui atteignent leur maximum en 1976 chutent à leur tour pour se stabiliser en dessous de 200 000 tonnes de 1979 à 1983.
En 1981, un refroidissement se manifeste dans les températures de surface, signe d'une reprise de l'upwelling et en 1983, la biomasse qui s'était stabilisée aux alentours de 1,2 millions de tonnes depuis 1978, augmente brusquement à nouveau à la suite d'un fort recrutement. Les évaluations acoustiques réalisées dans cette zone enregistrent cet accroissement qui semble se poursuivre en 1984.
Une analyse de l'évolution de l'upwelling côtier semaine par semaine au moyen de thermographie satellitaires (Meteosat II et NOAA-7) dans la zone atlantique marocaine, pendant la période 28 Juin-8 Août 1984 a été effectuée (Refk, sous presse).
Les résultats de cette analyse correspondent aux diverses descriptions du phénomène, trouvé dans la littérature concernant la région.
Simultanément aux observations par télédétection les débarquements hebdomadaires de sardine et de maquereau ont été suivis au cours de la période d'étude pour la zone de pêche traditionnelle. Dans la zone d'Agadir l'évolution de l'upwelling, du rendement, pour la sardine et pour le maquereau vont en parallèle mais, avec respectivement une semaine de décalage. Dans la zone Safi-Essaouira où l'upwelling et le dévelopement de la production primaire sont considérablement influencés par les vents locaux, on a constaté que le rendement pour la sardine diminue pendant que l'upwelling s'intensifie, progresse et regresse; les forts vents ont bien provoqué des remontées d'eaux froides mais ont empêché en même temps le développement de la production primaire en créant un vigoureux brassage.
Les thermographies satellitaires ne sont pas les seuls produits pouvant déterminer l'existence de zone potentiellement favorables à la pêche. La recherche des zones productives à l'aide de mesures par télédétection de la teneur de la mer en chlorophyle devait donner des résultats aussi satisfaisants et complèterait les renseignements des thermographies.
Les recrutements obtenus par l'analyse de cohorte de 1976 à 1982 fluctuent de façon considérable d'une année à l'autre. La régression non linéaire (méthode de Doubleday) fournit une évaluation plus précise que la méthode de Gulland en ce qui concerne les années les plus récentes.
Après deux faibles recrutements en 1976 et 197è succède une alternance de forts et faibles recrutements jusqu'en 1982 (Figure 12).
Une comparaison entre ces valeurs et les captures par unité d'effort des classes d'âge O et I additionnées sur une même cohorte (en millions de sardines) montre une concordance étroite entre les deux séries de données de 1976 à 1982. Les CPUE des classes O + I peuvent donc servir d'indice de recrutement pour les années antérieures à 1976.
Aux recrutements moyens de 1968 à 1969 succède un très fort recrutement en 1970 qui dominera la pêcherie jusqu'en 1974 et procurera les très fortes captures de 1973. A partir de 1971 les recrutements sont en déclin progressif jusqu'à la nouvelle augmentation de 1978–1982.
Des variations d'une telle amplitude ne peuvent être mises en relation avec les biomasses du stock reproducteur. La relation stockrecrue, en ce qui concerne les petits pélagiques, apparait donc comme complètement masquée par l'action de l'environnement qui agissent sur les conditions de survie des larves, assure le succès ou l'échec du recrutement.
Une estimation des coefficients de sélectivité vis à vis de l'âge des captures a été réalisée par Rami (1984) notamment pour les classes 0 et I qui sont incomplètement recrutées. L'âge de sélection traduit dans ce cas le recrutement partiel des classes d'âge dans la pêcherie et la sélectivité de l'engin de pêche. Les coefficients So = 0,14 et S1 = 0,55 donnent les proportions respectives des classes O et I pleinement vulnérables aux engins de pêche. Le modèle de Doubleday (1976) a été appliqué à cet effet aux données de captures par âge en nombre des pêcheries A et B combinées.
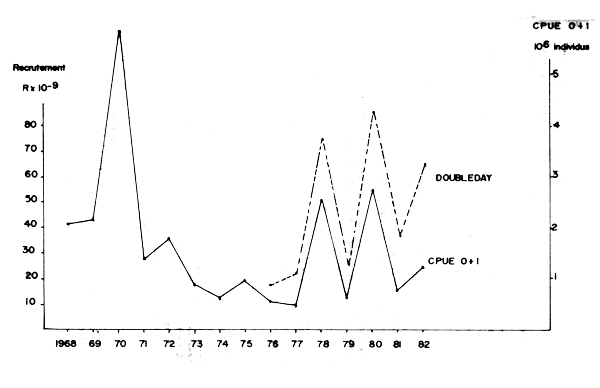
Figure 12 : Comparaison du recrutement à la CPUE des classes 0 + 1 appartenant à une même cohorte
Rami a également calculé ces coefficients respectivement pour la flottille marocaine en zone A et la flottille espagnole en zone B et constaté que les valeurs obtenues dans le premier cas sont beaucoup plus fortes que dans le second. Ceci est également montré par la composition en âge des captures marocaines où l'importance des classes O et I est plus forte que dans les captures espagnoles. La faiblesse des groupes O et I dans les captures espagnoles est due à des impératifs commerciaux et non à des différences dans la représentation des classes d'âge dans la zone B (Belvèze, 1984). En effet, les échantillonnages biologiques lors des campagnes acoustiques de l'ISPM montrent bien l'importance des classes O et I dans la zone B.
Par ailleurs, le Groupe a souligné que compte tenu du taux élevé de mortalité naturelle des petits pélagiques, toute tentative d'augmentation productivité n'est à attendre d'une capture de sardines plus âgées. Les simulations effectuées par Belvèze (1984) sur les effets attendus des changements de la taille à la première capture appuient ce point de vue.
Ceci n'exclut pas la réalisation d'expériences de sélectivité sur les engins utilisés et notamment sur les chaluts pélagiques.
![]()
![]()
![]()