![]()
![]()
![]()
A titre purement indicatif, le Groupe a procédé à un calcul de la biomasse totale disponible en Mauritanie et du niveau actuel d'exploitation en utilisant les données de biomasse obtenues par chalutage (CNROP, 1985 et Domain, 1985).
| Biomasse minimale | |
| Plateau 0–200 m (toutes espèces) | 57 500 t(1) |
| Banc d'Arguin et Baie du Lévrier (toutes espèces) | 28 000 t(2) |
| 85 500 t |
(1) Source Domain (Annexe M) données N'DIAGO
On doit noter que les évaluations ont été obtenues avec des bateaux de recherche différents.
Si on considère un coefficient d'efficacité du chalut de 0,5 la biomasse réelle serait le double de la valeur calculée soit enviorn 171 000 t. On admet généralement la formule C = F.B où C représente la capture annuelle, F la mortalité par pêche et B la biomasse annuelle moyenne.
La capture débarquée de toutes les espèces démersales en Mauritanie pour 1983 est de 51 700 t environ. Une part importante de la capture est cependant rejetée par les bateaux, surtout par la pêche des céphalopodes. Ces rejets attenidraient 30 pour cent en moyenne (selon Gréboval, 1982). La capture réelle serait donc de l'ordre de 67 000 t.
On obtiendrait dont une grossière estimation de F par F = C/B, F = 67 000 / 171 000 = 0,39
Si on compare cette valeur de F avec la valeur moyenne de M (la mortalité nautelle) calculés par Mennes (1984) pour les sparidés (M = 0,48) et si l'on admet cette valeur pour le stock démersal du plateau de Mauritanie on obtient alors la mortalité total Z = M + F soit Z = O, 48 + 0,39 = 0,87 et le taux d'exploitation E = F/Z = 0,448 ce qui indiquerait que le stock de poissons démersaux de Mauritanie est globaement proche de la pleine exploitation (où F = M et E = 0,50).
Il faut cependant souligner des nombreuses limitations à ce type de calcul pourtant souvent utilisé sans précaution:
le coefficient d'efficacité du chalut n'est pas connu et le choix de 0,5 est assez arbitraire; le coefficient peut varier de 0,25 à 0,75.
les estimations de biomasses obtenues en utilisant le N/O ALMORAVIDE sur la baie du Lévrier et le banc d'Arguin et le N/O N'DIAGO sur le plateau ne sont pas forcément comparables dans la mesure où l'éfficacité des deux bateaux comme engis de prélèvement peut être différente;
le taux d'exploitation varie géographiquement et il apparait clairement dans les résultats obtenus par d'autres méthodes (Annexe M) que le nord de la mauritanie (cap Blanc) est beaucoup plus exploité que le sud. La valeur moyenne de E = 0,448 ache donc, au delà des limites de validité qui lui sont propres, le fait que l'état des stocks démersaux du plateau dans le nord est plus préioccupant.
Un diagramme de production par recrue (Figure VII.1) a été en utilisant les paramètres de croissance de Domanievski (1985) et la valeur de la mortalité naturelle (M = 0,24) estimée à partir de l'équation de Pauly (1980).
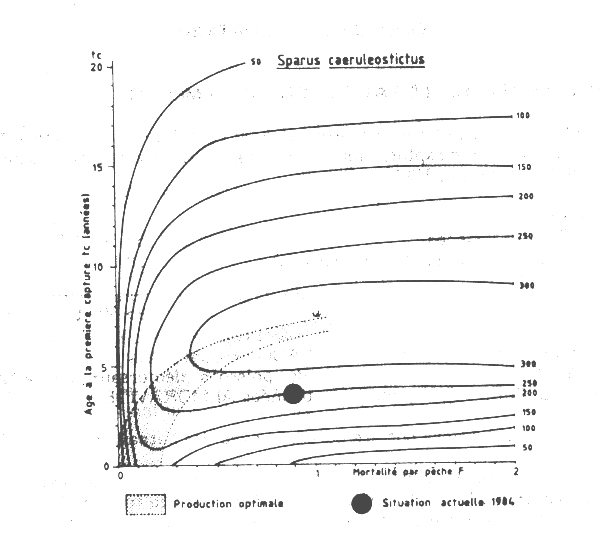
Figure VII.1 Courbes d'isoproduction par recrue de Sparus caeruleostictus de Mauritanie
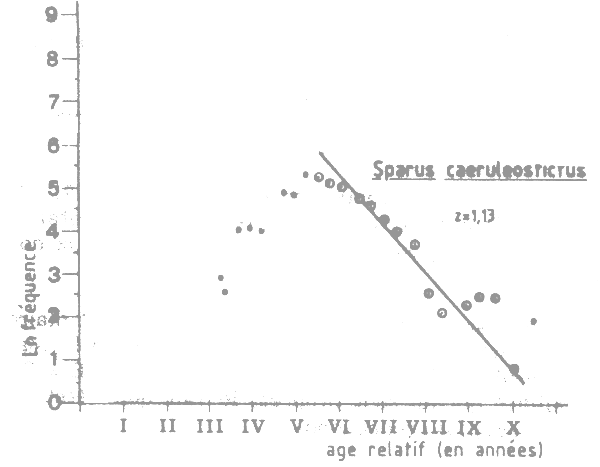
Figure VII.2 Courbe de capture pour Sparus caeruleostictus (1984) en Mauritaine (Banc d'Arguin et Banc du Lévrier)
Le niveau actuel d'exploitation de Sparus caeruleostictus est estimé en utilisant les données de fréquence des tailles (9 échantillons représentant un total de 487 individus) collectées par le N/O ALMORAVIDE du CNROP entre le 10 avril et le 28 juillet 1984 (Tableau VII.1). La longueur fourche estimée à la première capture est lc = 21 cm correspondant à un âge tc de 3,5 ans. L'analyse da la courbe de capture établie à partir des longueurs (Pauly, 1984) (Figure VII.2) donne une mortalité totale estimé Z = 1,13. La mortalité naturelle étant de 0,24 la mortalité par pêche est donc F = 0,89 et le taux d'exploitation E = F/Z = 0,78. Des valeurs similaires de Z sont obtenues en utilisant la relation entre Z et la longueur moyenne dans l'équation de Beverton et Holt, 1956 (in Pauly 1984, p.70).
La situation actuelle (tc = 3,5 ans et F = 0,89) est représentée sur le diagramme de production. Ce point se trouve en dessous et à droite de la ligne de production optimale pour ce stock, ce qui laisse supposer que ce stock est fortement surexploité avec un âge à la première capture trop faible et une mortalité par pêche F plus de deux fois supérieure à celle nécessaire pour obtenir une production par recrue optimale. On estime que la production par recrue de Sparus caeruleostictus pourrait être améliorée par une réduction d'au moins 30 pour cent de la mortalité par pêche et/ou par une augmentation de l'âge à la première capture. Cependant, la totalité des effets de telles mesures doit être discutée dans le contexte de la pêcherie des céphalopodes où la plus grande partie des Sparus est capturée comme prise accessoire.
Le diagramme de production par recrue (figure VII.3) a été établi en utilisant les paramètres de croissance établis par Mennes (1984) et la valeur de la mortalité naturelle M = 0,3 (Caverivière et al., 1985).
Pour estimer l'état atuel d'exploitation de Dentex canariensis nous avons utilisé les données de fréquence des tailles (8 échantillons, 485 individus) collectées par le CNROP à bord du N/O ALMORAVIDE entre le 10 avril et le 28 juillet 1984 (Tableau VII.2). La taille à la première capture a été estimée à lc = 19 cm correspondant à un âge tc = 1,8 ans. L'analyse de la courbe de capture établie à partir des longueurs, ainsi que l'analyse des données de longueur moyenne, conduisent toutes deux à des estimations très élevées de la mortalité totale (respectivement Z = 3,1 et 2,9) ce qui montre que ce stock est trop fortement surexploité par la pêcherie actuelle de céphalopodes. La mortalité par pêche sur ce stock de Dentex est trop forte, et les poissons capturés trop petits.
La production par recrue de Dentex pourrait probablement être augmentée de plus de 300 pour cent par une réduction de la mortalité par pêche, et un accroissement de l'âge (ou de la taille) à la première capture. Cependant, comme pour les autres Sparidae capturés en Mauritanie, tout changement dans le modèle d'exploitation est à considérer en relation avec la pêcherie de céphalopodes.
Le diagramme de production (Figure VII.4) a été établi en utilisant les paramètres de croissance adoptés par le dernier Groupe de travail COPACE sur les chinchards et les maqueraux (FAO, 1983). La mortalité naturelle (M = 0,5) est égale à la moyenne des deux estimations présentées à ce même groupe.
Les fréquences des tailles collectées par le CNROP sur les navires de la flottille pélagique soviétique (34 403 poissons mesurés entre novembre 1982 et avril 1985) ont été utilisées pour estimer la taille et l'âge à la première capture ainsi que la mortalité totale Z. Les échantillons n'ont pas été pondérés par les captures, les données n'étant pas disponibles. Une simple somme de tous les échantillons (Tableau VII.3) a été effectuée pour l'analyse de la courbe de capture établie a partir des longueurs (Figure VII.5). La valeur estimée de la mortalité totale est Z = 0,8 ce qui correspond à un F = 0,3 et à un taux d'exploitation E = 0,375 (si M = 0,5). L'âge à la première capture tc est d'environ 0,6 an, ce qui correspond à une longueur lc = 18,5 cm.
Les valeurs de tc et de F décrivant la situation actuelle sont représentées sur le diagramme de la Figure VII.4. Ces valeurs indiquent que le stock de Trachurus trecae est modérément à pleinement exploité avec une production par recrue très proche de la valeur optimale pour le maillage actuellement utilisé. La production par recrue pourrait apparemment être augmentée, par un accroissement simultané de l'effort de pêche (donc de F) de la taille de première capture (et donc du maillage) 1. Cependant toute augmentation de F serait suivie par une diminution de l'abondance totale et des rendements. Par exemple pour obtenir une augmentation de 50 pour cent de la production par recrue (de 40 à 60 g) il serait nécessaire de doubler l'âge à la première capture (par un accroissement du maillage d'environ 30 pour cent) et pratiquement tripler l'effort de pêche (ce qui aurait pour conséquences une augmentation des coûts et une réduction de 40 pour cent de l'abondance totale et des rendements).
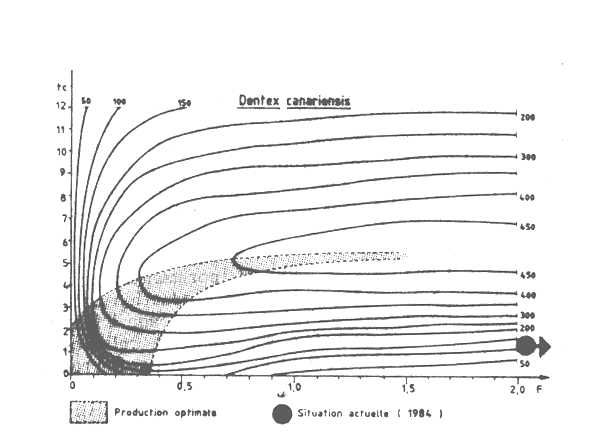
Figure VII.3 Courbes d'soproduction par recrue de Dentex canariensis de Mauritanie (1984) M = 0.3 ; K = 0.15 ; W oo = 13,7 kg; to = 0 ; et t = 20 ans Le point correspondant à la situation actuelle est hors du diagramme (tc = 1,8 ans et F = 3/an)
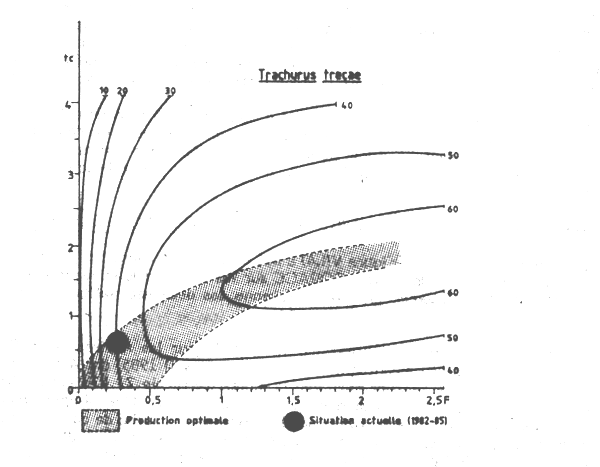
Figure VII.4 Courbes d'isoprosuction de Trachurus trecae de Mauritanie M = 0,5 ; K = o.345 ; W oo = 615,3 gr; t = -1,1 ; t = 9 ans
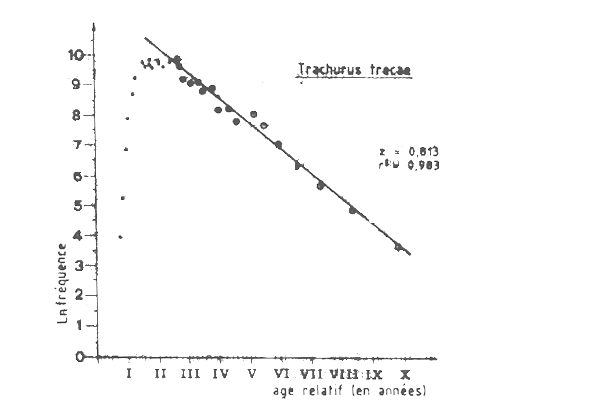
Figure VII.5 Courbe de capture pour Trachurus trecae capture en Mauritanie pr L'URSS entre 1982 et 1985
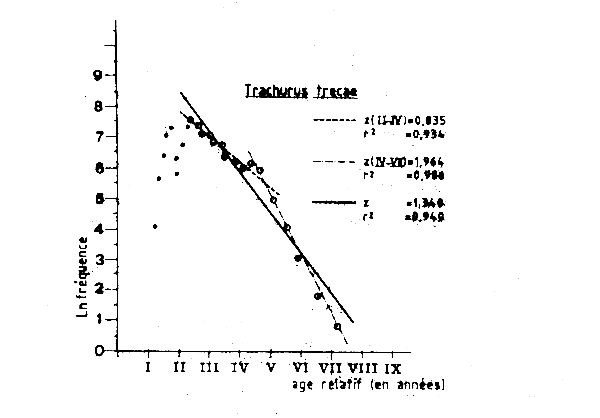
Figure VII.6 Courbe de capture de Trachurus trecae capture en Mauritanie par la flotte roumaine en 1982
Les données de fréquence des tailles pour la flottille roumaine en 1982 étaient aussi disponibles au groupe de travail, et l'analyse de la courbe de capture établie à partir des longueurs (Figure VII.6) donne une estimation plus forte de la mortalité totale. On obtient ainsi Z =1,3 ce qui correspond à une mortalité par pêche F de 0,8 et à un taux d'exploitation E = 0,62 ce qui indiquerait qu'en 1982 le stock était déjá pleinement exploité, pour la valeur correspondant de l'âge á la premiére caputre (tc= 0,9 an ). Un autre fait intéressant que montre l'analyse de la courbe de capture établie á partir des longueurs utilisant les données de la flottille roumaine (Figure VII.6), c'est qu'il est possible d'estimer deux valeurs différentes de Z. Jusq'à âge de quatre ans (âge relatif V sur ce graphique) Z = 0,835 et après cet âge Z = 1,964. Ce changement de Z peut avoir pusiurs causes:
une augmentation brutale de la mortalité naturelle à partir de quatre ans (hypothèse peu probable);
une augmentation importante de la capturabilité (et donc de F) à partir de 4 ans (hypothèse également peu probable);
une brusque diminution de la mortalité par pêche partir de 1978 (quand les poissons de quatre ans en 1982 sont entrés dans la pêcherie) et au moins jusq'en 1982. Cette dernière hypothèse semble être confirmée par les information disponibles sur l'extension des zones de pêche et sur l'effort de pêche de ces flottilles ces années là;
une diminution importante du recrutement. La dispartion récente des concentrations côtières et l'effondrement des biomasses de chinchard tendentà confirmer fortement cette dernière hypothèse.
Un diagramme de production pour Scomber japenicus a étéétabli (Figure VII. 7), en utilisant les paramètres de croissance adopté par le Groupe de travail COPACE sur les chinchards et les maqueraux (FAO, 1983). La mortalité naturelle a été à M = 0, 45, cette valeur étant la moyenne des deux estimation présentées au groupe de travail préédemment mentionné.
Les données de fréquence des tailles, relatives à la pêcherie industrielle soviétique, et collectées par le CNROP entre 1982 et 1985 (8 874 mensuration en 12 mois entre novembre 1982 et avril 1985) ont été utilisées pour estimer la mortalité totale Z ainsi que la longuer et l'âge à la première capture. Ces échantillons n'ont pas été pondérés par la capture totale (les données nécessaires pour le faire n'étant pas disponibles) et une simple somme de tous les échantillons (Tableau VII.4) a été effectuée pour l'analyse de la courbe de capture établie à partir des longueurs (Figure VII.8).
L'estimation de la mortalité totale serait Z = 0, 62, ce qui nous donnerait F = 0, 17 et E = 0,27. L'âge à la première capture a été estimée à tc = 1, 6 ans correspondant à une taille lc de 24, 4 cm. La représentation de ces valeurs sur le diagramme de production montre que ce stock serait faiblement exploité et que la production totale pourrait être augmentée par un accroissement de l'effort de pêche et que conservant la taille et l'âge actuels de première capture. Cependant, tout accroissement de l'effort de pêche (pour accro&lcirc;tre la production totale) conduirait à réduire les rendements et à réduire l'abondance totale. Une augmentation de F de 0, 17à 0, 45 (pour avoir F =M) par exemple pourrait en théorie conduire à une augmentation de 40 pour cent du rendement par recrue (Y/N)et peut-être de la production totale, si le recrutement n'est pas affecté, mais aménerait également à une diminution de 30 pour cent environ du niveau actuel de l'abondance. II faut cependant insister sur le fait que le stock de maquereaux est un stock don't le potentiel varie avec les conditions climatiques. II est vraisemblable qu'en cas d'arrivée de bons recrutements dans les populations l'effort de recherche (et de pêche) orienté sur cette espèce augmenterait immédiatement par transfert à partir des autres pêcherie pélagiques et la taille de prenmiére capture diminueratit par concentration de l'effort sur les classes jeunes et abondantes.
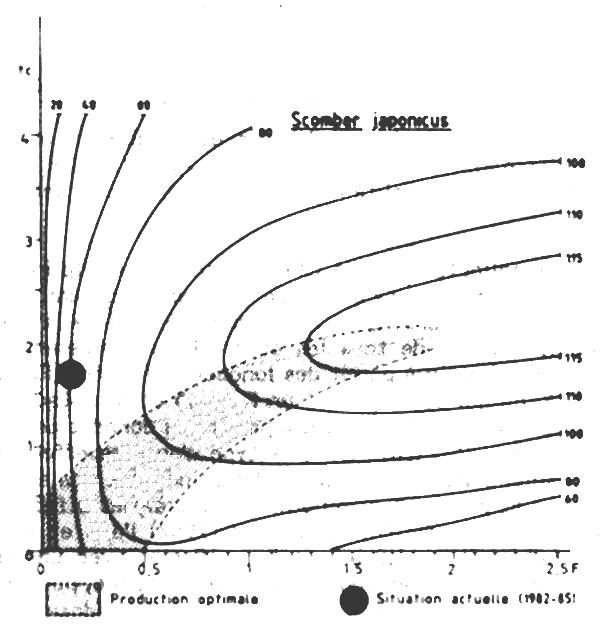
Figure VII.7 Courbes d'isoproduction de Scomber japonicus M = 0,45 ; K = 0,309 ; W oo = 1,27 kg; to = -1,01 ; t = 10 ans (= 3/K)
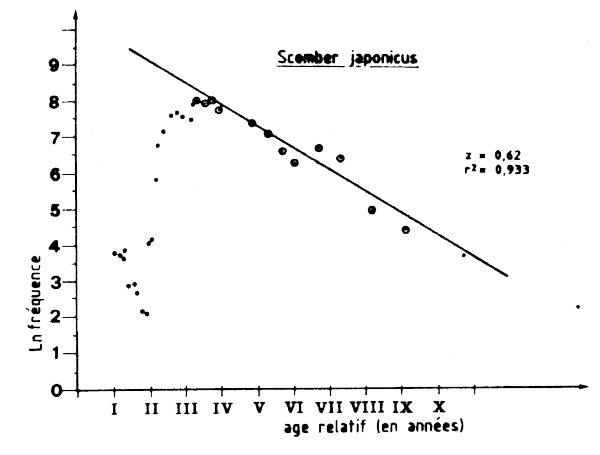
Figure VII.8 Courbe de capture pour Scomber japonicus capture en Mauritanie par L'URSS entre 1982 et 1985
Le diagrmme de production de la Figure VII.9 a été étabil en utilisant les paramètres de croissance présentés par la Roumanie lors du Groupe de travail COPACE déjà précédemment (FAO,1983). Un coefficient de mortalité naturelle M = 0,3 a été à partir de l'équation de Pauly (1980).
Les fréquences de tailles collectées par le CNROP sur la flottille industirielle soviétique entre 1982 et 1985 (10 655 poissons échantillonés en douze mois entre déceembre 1982 et avril 1985) ont été utilisées pour obtenir des estimations de la longueur et de l'âge à la premiére capture et de la mortalité totale Z. Une simple somme de tous les échantillons (Tableau VII>5) a é5é utilisée pour l,analyse de la courbe de capture établie à partir des longueurs (Figure VII.10). La valeur estimée de la mortalité totale est Z =1,8 ce qui correspond à un F= 1,5 et à un taux d'exploitation trés élevé E =0,8 . L'âge et la longueur à la premiére capture, à partir des donnés de la flottille socviétique sont respectivement tc = 3,3 ans et lc= 25,7 cm. Le même type d,analyse effectuésur ,les captures de la flottille roumaine en 1982 (Tableau VII.6) donne une estimation de Z égale à 1,84 (FigureVII.11) pour les groupes de poissons de 5 ans et plus tout conmme dans les analyses effectuées à la premiére soviériques. Dans les données de la flottille roumaine (1982) l'âge et la l;ongueur à la prmiére captre semblent être polus faible s(tc =1.8 ans et lc =19 cm) avec une mortalité par pêche (F) calculée par l'analyse des cohortes de jones comprise entre 0,3 et 0,6 pour des poissons dont la taille varie entre 19 et 29 cm.
Il est bon de nter la différence entre les structures de tailles des captures roumaines et soviétiques les roumains ayant apparemment tendance à exploiter de plus jeunes individus.
Dans les deux cas (Figure VII.9) le stock semble être surexploité avec une mortalité par pêche deux à trois fois supérieure à celle requise pour obtenir une production par recrue optimale.
A partir des mensurations effectuées sur le plateau contiental entre le cap Blanc et le cap Timiris et de 20 à 600m de profondeur lors de la campagne expérimentale du BAKHTCHISARAY, il a été possible d'estimer la mortalité totale Z de Merluccius sene galensis. Les données brutes ont d'abord été pondérees par la capture moyenne par heure de chalutage (Tableay VII.7) puis par la surface relative des différentes strates prospectées (Tableau VII.8). Ce calcul permis d,obtebnir une unique distribution de fréquence refrésentative de la sturcture démographique de l'espéce entre 20 et 600 m au moment de la campagne de chalutage.
La mortalité totale Z a ensuite été calculée par la méthode des courbes de capture en longueur décrite par Pauly (1984). Les paramétres de croissance utilisés sont: l =90cm , K=0,14 et to = 0,9. La valeur ainsi obtenue de Z est Z = 1,03. Une estimation de l'état d,exploitation de pêcherie peut être obtenue en comparant cette valeur à la mortalité par pêche F obtenue par différence avec la valeur de la mortalité naturelle M donnée par le rapport de Groupe de travail sur les merlus (CECAF/REVII/84/3). On a ainsi:
F =Z -M = 1,03-0,50 =0,53
et le niveau d'exploitation:
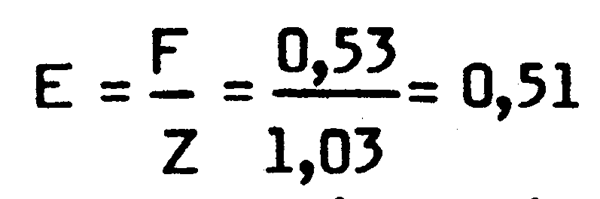
qui indique que le stock de merlu serait pleinement exploité.
(Les résultats sont présentés sur la Figure VII>12).
1 Par p. Boukatine, CNROP,B.P.22, Nouadhibou,Mauritanie
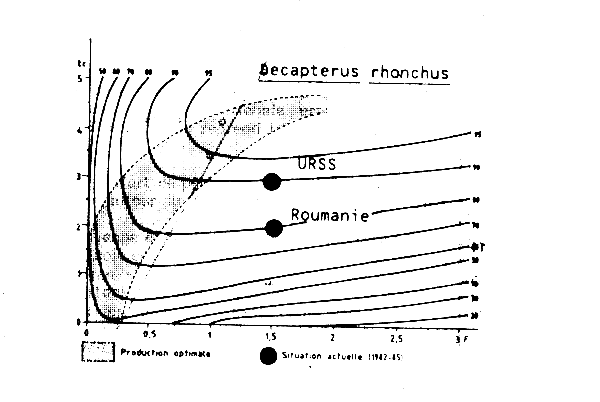
Figure VII.9 Courbee d'isoproduction par recrue de Decapterus rhonchus en Mauritanie M = 0,30 ; K = 0,136 ; W oo = 2,39 kg ; to = -1,2 ; t = 10 ans
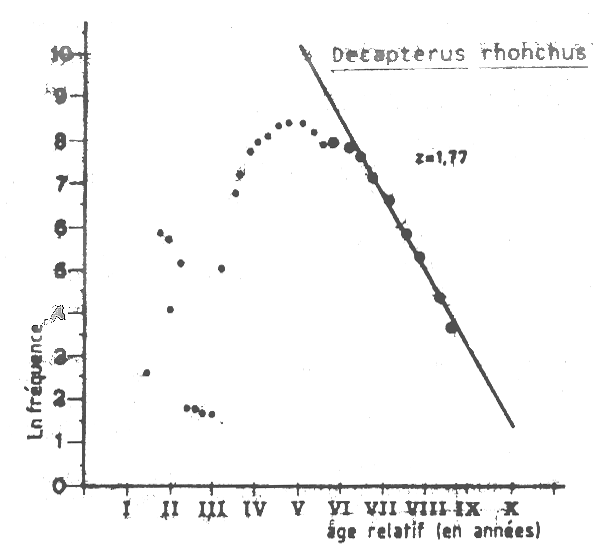
Figure VII.10 Courbe de capture pour Decapterus rhonchus de Mauritanie dans les captures de la flotte de L'URSS (1982–1985
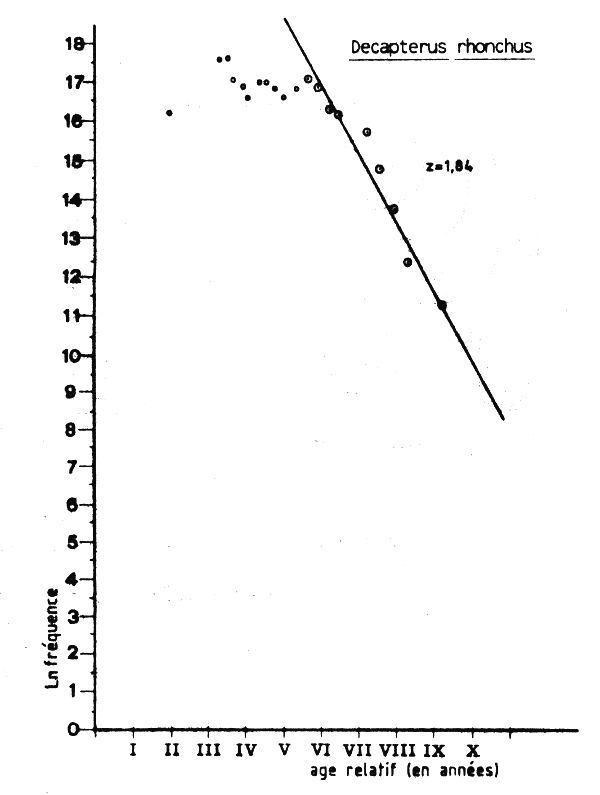
Figure VII.11 Courbe de capture de Decapterus rhonchus capture en Mauritanie par la flottille roumaine
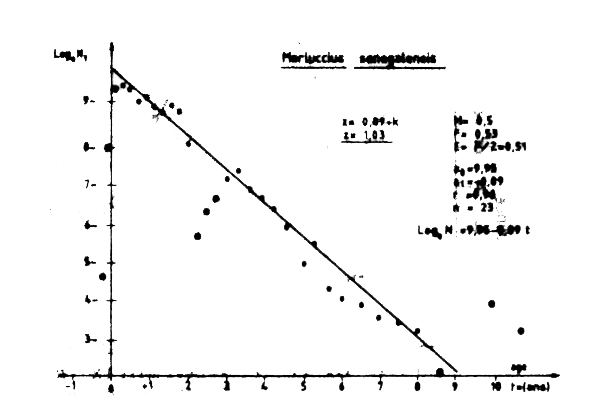
Figure VII.12 Merluccius senegalensis capture pendant la campange experimentale de N/O BAKHTICHSARAI (Maillage 10mm) (zone Nord: c. Timiris - c. Blanc) Donnée Boukatine
Cette langouste côtiére est exploitée par la pêche artisanale avec deux stocks : un stock nord et un stock sud.
Le stock nord s'étend du cap Blanc aux côtes du Sahara occidental (Villa Cisneros). II était exploitéavant 1975 par une flottille artisanale espagnole et quelques langoustiers francomaurtaniens. Aprés 1976, l'exploitation reprend lentement à partir de la Guerra par une flottille artisanale mauritanienne.
Suite à réduction de l'effort de pêche en 1976-1977, la langouste verte réapparait à partir de 1981-82 sur les fonds de pêche autour du cap Blanc et dans la baie du Lévrier.
La capture annuelle moyenne en période de pleine exploitation (1968-1975) est de 232 t pour une longueur de 333 km de côte, soit environ 700 kg/km. Environ 1/4 de cette côte peut être exploitée par la Mauritanie soit environ 60 t pour la Mauritanie. Ce chiffre est trés sous-estimé car la ,oyenne est calculée sur un stock trés exploité ce qui n'est plus le cas en 1985. La pêcheartisanale a produit 230 t en 1984 dans des conditions d'exploitation mal connues. Les données de captures ne sont pas suffisamment précises pour appliquer un modé;e de production. L'estimation réalisée en 1978 sur les données 1955-1968 nous avait donné une Prise Maximale Equilibrée (PME) de 298 t.
Un échantillonnage des tailles effectué en 1980 montrait un mode à 20/23 cm donc encore trés faible, semblable à ceux rencontrés par Maigret (1978) et Maigret (1980) ett Maigret (1980). Cependant les individus de taille supérieure à 24 cm étaient relativement plus nombreux (14,4 pour cent de captures contre 5,5 pour cent en 1973), ce qui laisse supposer une laisse supposer une reconstitution du stock.
Nous ne disposons pas de fréquence de tailles postérieures à 1980 pour cette pêcheire.
Le stock sud occupe les côtes sud mauritaniennes de Nouakchott à St. Louis. II est exploité uniqument par la pêche arisanale basée à Nouakchott et St. Louis au Sénégal, où est débarqué une partie des captures. Les chalutiers glaciers à la recherche des céphalopodes en capturent occasionnellement.
Si l'on estime que la densité de ce stock est identique à celle de la zone nord, les captures potentielles peuvent être estimées à 175 t/an.
Les tailles observées notamment dans les apports des chalutiers, sont beaucoup plus élevées que dans le stock nord (Mode 31-35 cm) et confirment le faible niveau d'exploitation du stock.
Le stock de langouste rose a été exploité à partir de 1956 sur une aire de répartition qui correspondait à un stock consiéré comme voerge ety réparti sur l'ensembale du plateau contiental entre 40 et 250 m de profondeur dans une région située entre le cap Barbas et le sud du cap Timiris.
Dés 1963, le stock surexploité par la pêche langoustiére (63 bateaux pour plus de 12 000 jours de pêche) aidée par la pêche chalutiére qui se développait sur le plateau contiental a été progressivement éliminé de son aire de répartition traditionnelle pour être cantonné dans les fosses qui entaillent le plateau.
A partir de 1970/71 le stock trés réduit ne se rencontre plus que dans des fosses profondes où il est difficile de travailler. Corrélativement, la flottille est réduite à 10 bateaux qui travaillent en permanence depuis, développant un effort relativement stable.
Afin de présenter l'évolution de la pêcherie, nous avons employé le modèle de Schaeffer en tenant compte de l'aire de répartition de la langouste et représentant les captures en kg/km2 en fonction de l'effort (J de P 1 000 /km)(Figure VII.13)
Ce modèle global peut être décomposé en fonction de trois phases d'exploitation corespondant à des aires de répartition de plus en plus réduites (Figure VII.14).
La premiére qui peut être considérée comme le stock vierge
PME = 3 138 T
fPME =7 946 Jours de pêche.
La deuxième, en période d'effondrement du stock (1963 à1970); l'estimation de la PME etde fPMEne représente pas l'évolution réelle car il n,est pas possible de se rendre compte de la diminution de l'aire de répartion avec exactitude.
La troisième , de 1977 â 1984
PME = 663 t
fPME = 1 681 j de P.
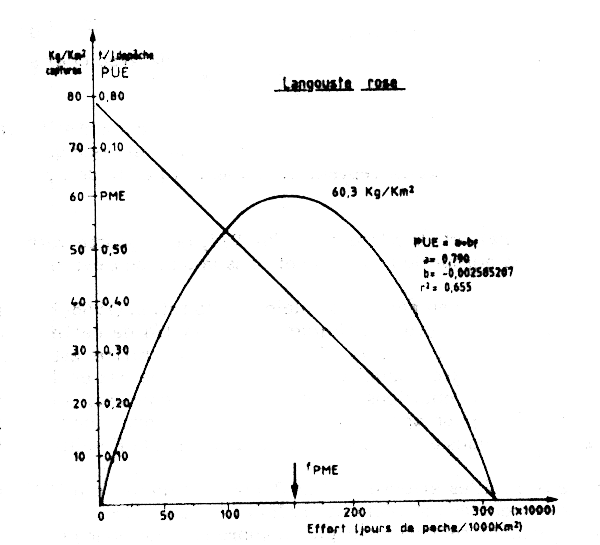
Figure VII.13 Modéle global pour la pêcherie de langouste rose standardisé pour les changements de superficie occupée par le stock
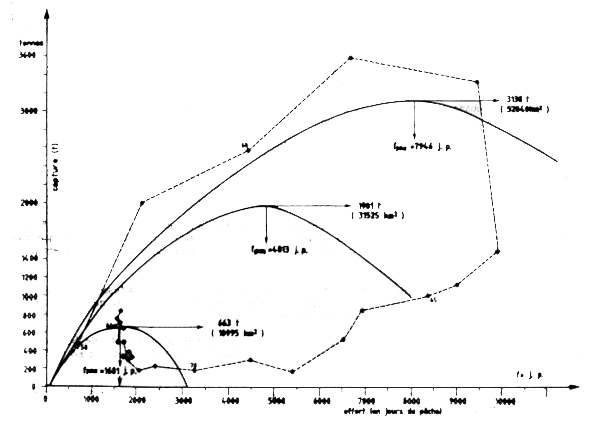
Figure VII.14 Modéle de production en fonction de l'aire de répartition de la langouste rose
En fait ce dernier mofè;r rdy probablement encore provisoire car l'ensemble des points 71-84 est group'e et la capture en hausse permanete ne dépend pas de l'effort développé. Cette hausse se poursuit actuellement et le modèle indique un potentiel en accroisment constant:
données 1969-1975, PME = 390 t (Maigret,1978)
données 1969-1679, PME = 597 t (Maigret,1980)
données 1958-1984, PME = 663 t
Deux hypothèes existent pour expliquer le fairt que les captures varient à la hausse de facon indépendante de l'effort:
l'effort serait mal évalé; il est en effet diffcile dans l'ètat actuel de préciser cet effort car les bateaux pêchent en flottille à la fois au chalut et aux casiers. Pourtant depusis une dizaine d'années les engins de pêche m'ont pas varriè, la flottille est resstée stable avec 10 bateaux toujours identiques. Il ne semble pas possible dans l'état actuel d'affiner les définitions de l'effort de pêche ;
le stock se reconstituersit et aprés avoir atteint sa densité optimale dans l'aire de répartition réduite les fosses profondes, il commence à s'étendre sur le plateau continental favoriséd par la diminution de l'effort des flottilles travaillant dans cette région à la recherche du merlyu, des crevettes profondes et de Sparidae.
Il est indiaspensable de poursuivre l'observation de cette pêcherie adfin de voir son évolution. Si l'exploitation continu avev ;e ,êche on peut penser que les captures continuieront à crotre pour se stabiliser sur la premièr courbe autour de 1 200 t/an, soit une P.U.E. autout de 710/720 kg/J de mer. Dans ces conditions, il serait probablement possible d'augmenter légèrement l'effort de pêche en autorisamt 1 ou 2 bateaux supplémentaires. A cette occasion il serati intéressant de réaliser un modèle économiques de cette pêcherie afin d'éaluer sa rentabilité actuele et envisager l'avenir de la flottille.
Une petite quantité de languoustes roses est pêche [ar ;a flottille de chalutiers espangnols exploitant le merlu noir sur le talus. Les captures n'en sont pas connues; il est indispensable de dissposer de ces quantitiés dans les années à années à venir afin d'intégreer ces captures à celles de la pêche langousitère. D'autre part l'augmentation des captures et de la P.U.E . de langouste dans cettle pêcherie serait une confirmation di faot qie ;a ;amgpiste recolonise en [artoe son ancienne aire de répartition sur ;e plateau.
Les plus récentes évaluations sont celles du Groupe de tracvail aad hoc du COPACE (FAO,1979; 1982 ) et celles contenues dans la compilation de Bravo de Laguna (1983)
Pendant la premirère session du Groupe de travail (FAO.1979), trois stocks de pouples ffurent identifiés, au large de Dakhla , du cap Blanc et de Noukchott respectivementy. IIs furent analyusés séparment à l'adie ede modèles de [production . Plus tartd, et au cours de la deuxième session du dGroupe de travail (FAO,1982), les évaluations firenmt mises à jour avec des données jusqi'à 1980 et des méthodese aussi bien globales qi'analytiques.
L'information disponible sur l;es captures féalisées en Maurityanie permet seulement d'utiliser les modèles de production de schaefer, de Fox ,ou généralisés, Dans cd dernier cas on a considéré emble touttes les données mauritaniennes que l'on considère comme provenant du stock du cap Blanc. Les données de base utilisées furent celles communiquées par Hatanaka (1979). La conclusion du Groupe de travail fut que, à ce moment là, le sstock était sous -exploité.
Tout de suite, après cette réunion se tint la 6ème sessopm du Groupe de travail deCOPACE pour l'évalutation des ressources au cours de laquelle furent présentées des données des données novelles coréemmes qui permirent la mise à jour des évaluations du Groupe de travail (FAO,1982). Les résultats furent légérement différents sur le plan quantitatif mais pas sur le plabn qualitatif.
PME =42 600 t, f1980f0,1 =0,33 stock très sous-exploité
Bravo de Laguna /(1983) révisa ces évaluation appliquant le modèle de Fox au stock de Dakhla et du capp Blanc. Dans le premier cas il utilise les données 1967-82; dans le second les données 1969-80 (Tableau VII.11 Figure VII.15). La prise maxmale équilibrée (PME) obtebnue fut de 42 600 t et le rapppor f1980f0,1 = 0,33. Le stock apparauissait sous-exploité.
L'historique des évaluations des stocks de deeiches a été similaire à celui des stocks de poulpes. On n'a cependant pas pu identfier de stocks séparés. En oute les statistiques disponibles étaient regroupées par Division statisique du COPACE sans plus de précision. Pour cela le Groupe put seulement évaluer séparément les stocks des Dicisions34.1.3 et 34.3.1. Bravo de Laguna (1983) révisa les évaluations jusquà 1982 pour la Division 34.1.3. Cette révision concerne les captures réalisées dans les eaux mauritaniennes.
Les données de capture furent obtenues auprès de la FAO (1983) pour 1969-80, et de Shimura (1983) aussi que de la flottille espagnole pour 1981-82. Les CPUE utilisées sont les CPUE espangnoles. Le modèle de Fox élaboréindique que la PME est de 32200 t et que le rapport f1982/f0,1 = 1,82. LE stock apparit donce comme très fortement surexploité en 1982 et une réducion de l'effort de 46,4 pour cent serait nécessarie pour obtenir la capture otpinale correspondant à f0,1 (Tableau VII.12 et Figure VII.16).
Les données disponibles sur le calmar ont imposé de traiter globaalement le stock de la Division 34.1.3. Les données utilisés par Bravo de Lagjna (1983) furent la captrure totale et la CPUE espagnole pour 1969–80, fournies lors de la 7éeme session du Groupe de travail sepécial du COPACE (FAO,1982) ainsi que les données de Shimura (1983) complétées par les dobnnées par les données espagnoles pour 1981-82 (Tableau VII.13). Le modèle de Fox indique une forte surpêche (FigureVII.17). La PME est de 20 300 t. Le rapport f 1982/f0,1est de 2,93 indiquant, pour l'ensesmble de la régionune trés grave surpêche. Une réduction globale de l'effort de 66 pour cent serait nécessaire pour atteindre le niveau f0,1.
Sy Moussa et Tchernichkov (1985) ont analysé les variations de la C[UE de céphalopodes au cap Blanc en liaison avec les anomalies de tempéreature pour la période 1966-82. Les anomalies de] température furent utilisées par ces auturs comme des indices d'upwelling et ils déclaraent avoir décelé une relation nette entre l'intensité de l'upwelling et les CPUE de poulpes.
Pour vérifier ces résultats le Groue a procédé à une analyse similaire disposant pour cela des données définitives et correctes de prise et d'effort pour 1981–82 (dans le document cité ci-dessus, les chiffres provisoires utilisés étaient eronnés). Les indices d'upwelling ont été détermines graphiquement à partir de la figure 2 de Sy et Tchernichkov (1985). Les anomalies de CPUE ont été calculées comme la différence entre les valeurs observées et prédites par les modèles exponentiels et linéaires de production (Tableau VII.14, Figure VII.18). Ces anomalies sont portées sur les Figures VII.19 et VII.20. Elles représentent la variabilité de la valeur de la CPUE qui ne peut être expliquée par des changements de l'effort de pêche.
Les résultats du Groupe ne montrent aucune relation entre anomalies thermiques et anomalies de CPUE et diffèrent donc sensiblement de ceux présentés par 5y et Tchernichkov (1985). La différence tient essentiellement au mode de calcul des anomalies de CPUE par ces deux auteurs qui n'est pas entièrement correcte dans la mesure où l'influence de l'effort d pêche (une variable dominante) n'a pas été d'abord extraite. Quoi qu'il en soit l'etude mérite d'étre poursuivie.
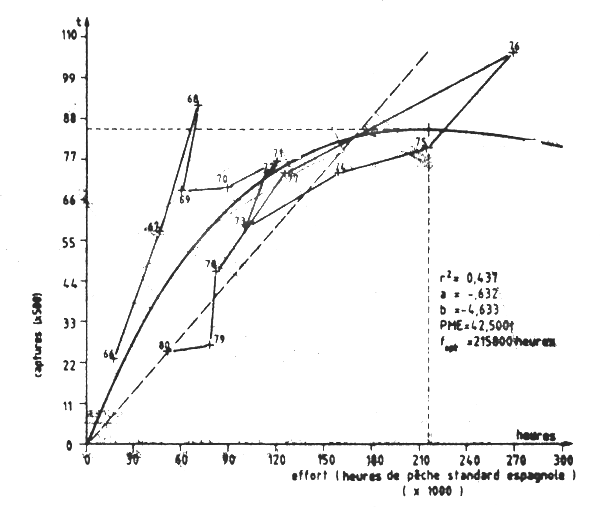
Figure VII.15 Modéle de production de Fox du stock de poulpes de cap Blanc (Division 34.1.3) Données 1966–1980
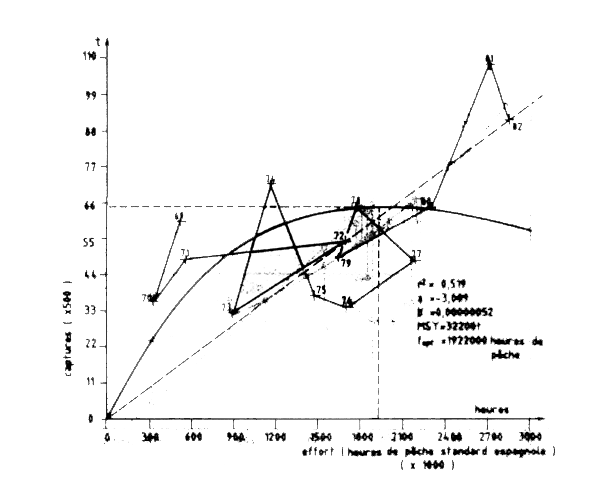
Figure VII.16 Modéle de production de Fox de la seiche Division statistique 34.1.3
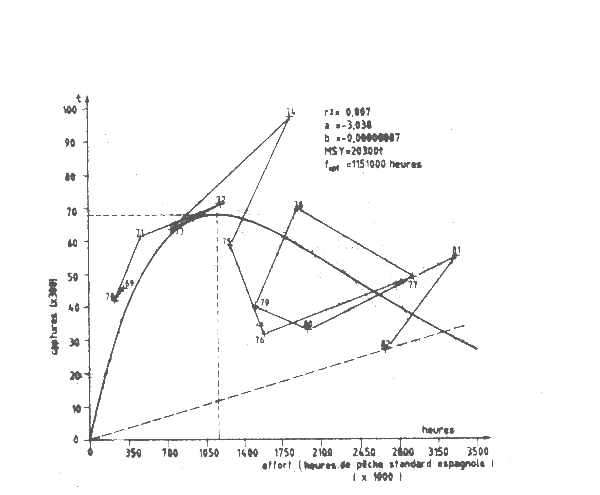
Figure VII.17 Modéle de production de Fox du calmar Division statistique 34.1 du COPACE
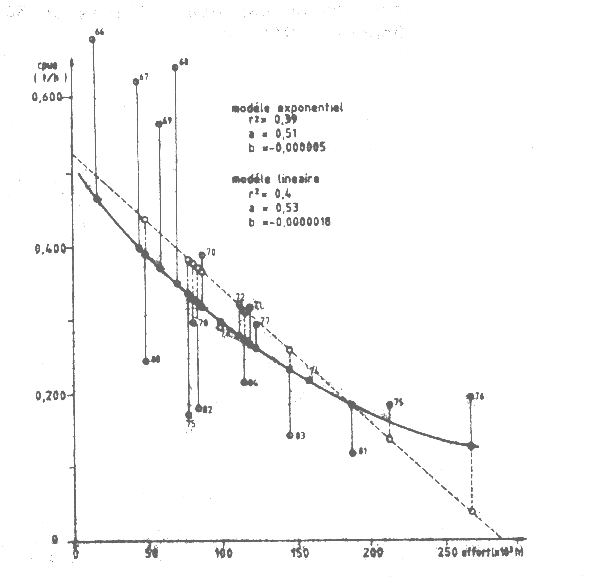
Figure VII.18 Modéles linéaire et exponentiel de la relation entre espèce et effort (octopus/cap Blac) et anomalies de cpue = différences entre valeurs calculées théoriques et valeurs observées
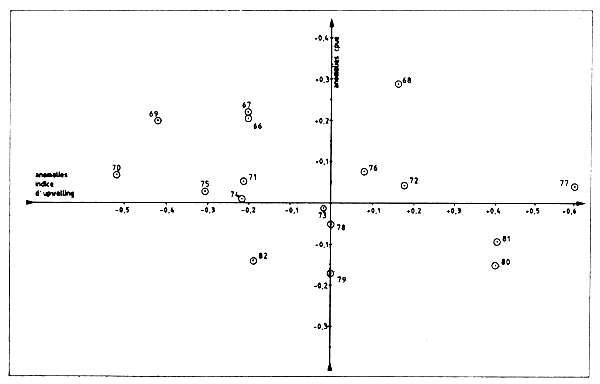
Figure VII.19 Relation entre les anomalies de l'indice d'upwelling et les anomalies de cpue (tirées de la figure VII.17) Poulpe/c. Blanc - Modéle linéaire
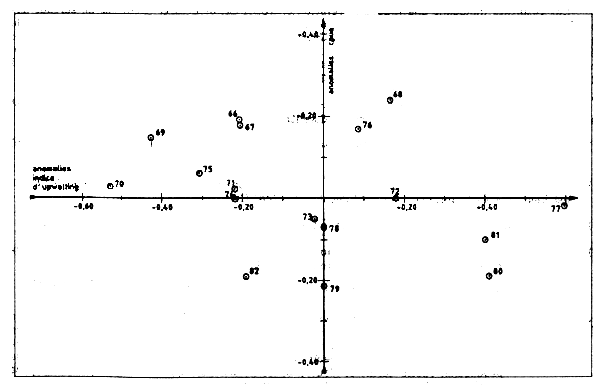
Figure VII.20 Relation entre les anomalies de l'indice d'upwelling et les anomalies de cpue Poulpe/c. Blanc - Modéle exponentiel
- Poulpes
Les données disponibles à la réunion étaient les suivantes:
modéle de production pour les octopus du cap Blanc et données japonaises de capture et d'effort de 1966 à 1980 (FAO, 1982);
données mauritaniennes de capture d'octopus de 1981 à 1984. On suppose qu'il s'agit lè des seules captures réalisées au cap Blanc pendant ces années;
données mauritaniennes de CPUE en tonnes/jour de pêche de chalutiers de type 1 (250 TJB) de 1974 à 1979 et type 2 (250 TJB) de 1981 à 1984.
Pour mettre à jour les évaluations effectuées par les Groupes de travil du COPACE il fallait exprimer l'effort mauritanien en équivalent effort japonais (pour pouvoir constituer une seule série homogène de données comparables). Pour cela on a calculé le rapport entre la CPUE moyenne 1974-79 de bateaux japonais (données Hatanaka, 1979) et des bateaux mauritaniens1 de la MAFCO (données CNROP). Ce rapport était égal à 2,11. On a donc multiplé par 2,11 la PUE mauritanienne de 1981 à 1984 pour la rendre équivalente à celle que les bateaux japonais auraient obtenu sur le même stock pendant la même période.
En divisant la capture totale en Mauritanie de 1981 à 1984 (telle que rapportée dans les statistiques officielles du CNROP) par la PUE ainsi corrigée, on obtient un effort total pour la période 1981–84 exprimé en heures de pêche d'un chalutier de type japonais.
Grâace à cet étalonnage de l'effort mauritanien en unités japonaises, la série historique s'étend alors de 1966 à 1984 (Tableau VII.11). Les données ont été utilisées dans un modèle de Schaefer (Figure VII.2l) car l'ajustement était meilleur que pour le modèle de Fox.
Les résultats indiquent une mauvaise relation entre CPUE et effort. Ceci est dû surtout aux données des quatre dernières années et plus particulièrement 1981–83 et 84. La prise maximale équilibrée (PME) est de 35 600 t et l'effort exercé en 1984 est pratiquement égal a fPME, l'effort nécessaire à l'extraction de cette de cette PME. Ceci signific que le stock serait exploité au maximum, avec un effort correspondant à 232 000 h de chalutage d'un chalutier type mauritanien actuel, soit à 11 600 jours de pêche à raison de 20 h de chalutage par jour.
La mauvaise qualité de l'ajustement des points correspondant aux quatre dernières années n'est pas facile à expliquer et plusieurs hypothèses existent:
les déclarations de capture mauritaniennes seraient incorrectes et sous-estimées d'environ 50 pour cent (!);
la conversion de la CPUE mauritanienne en CPUE japonaise sur la base des données MAFCO, est inappropriée à cause de la qualité des données MAFCO;
l'effort nominal mauritanien 1981-84 est mal estimé;
le recrutement d'octopus est en régression pour des raisons inconnues et indépendantes de la pêche et par conséquent le potentiel du stock a baissé.
Ces hypothèses sont examinées dans l'ordre ci-après (cf. Figure VII.22).
Hypothèse 1 Si la capture totale est sous-estimée et l'effort mauritanien observé est correct, la CPUE mauritanienne serait sous-estimée (parce que calculée par le rapport entre prise et effort)(les données réelles se trouvent en Al (Figure VII.22) et le stock serait bien exploité au niveau du maximum de production en 1984; mais sa production réelle serait le double de la production apparente. Le problème posé serait celui de la sous-déclaration des captures.
1 Rendements exprimés en kg/heure de pêche, en supposant 20 h de pêche par jour
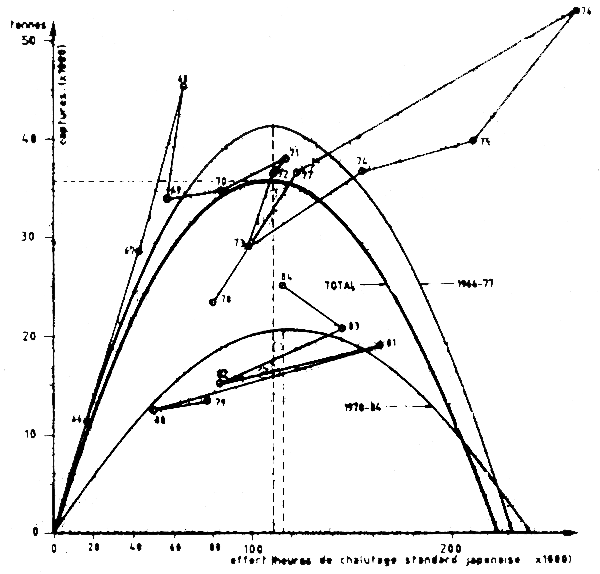
Figure VII.21 Modéles de production de Schaeffer - Octopus/c. Blanc
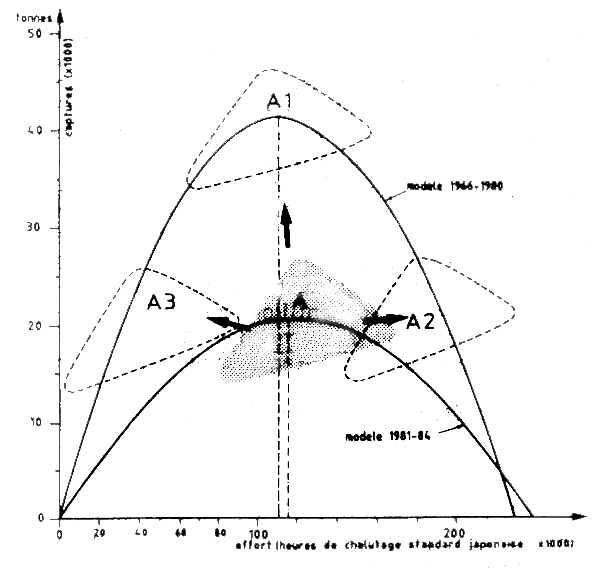
Figure VII.22 Illustration géographique des diverses hypothèses concernant l'état des stocks de poulpes du cap Blanc A = position actuelle des données capture-effort pour la période 1981–84 (données mauritaniennes) A1 A3 = positions possibles en fonction des diverses hypothèses (voir texte)
Hypothèse 2 Si, au contraire, la capture totale déclarée de 1981 à 1984 est correcte mais que le rapport PUE japonaise/PUE MAFCO n'a pas de sens (par exemple si la qualité des anciennes données MAFCO est douteuse) la correction par un facteur 2,11 serait injustifiée. Dans ce cas une autre possibilité serait que les bateaux mauritaniens (qui sont en réalité à l'origine des bateaux japonais et bateaux mauritaniens l'efficacité qu'ils avaient dans le passé. Dans ce cas la CPUE observée ne doit pas être corrigée car elles est directement comparable avec celle donnée par Hatanaka et l'effort réellement exercé de 1982 à 1984 est le double de celui indiqué sur la Figure VII.21. Les données 1981–84 se trouveraient en A2 (Figure VII.22) et le stock, dont le potentiel serait toujours de l'ordre de 40 000 t, serait très fortement surexploité ce qui expliquerait la faiblesse des captures et des rendements.
Hypothèse 3 L'effort mauritanien est surestimé. Les données originales sont exprimées en tonnes/jour d'un chalutier de type 2. Ces données peuvent elles-mêmes être biaisées si l'effort est estimé à partir des jours d'absence au port. De plus pour convertir les jours de pêche mauritaniens en heures de pêche, pour des raisons de compatibilité avec les données historiques, on a considéré 20 h de pêche/jour. Ce chiffre pourrait être trop élevé car le nombre d'heures de pêche/jour semble varier beaucoup d'un type de bateau à l'autre. Dans ce cas si la prise totale est correcte, la réduction d'un heures de pêche calculées conduirait à un accroissement de la CPUE estimée, à un niveau plus comparable à la série des CPUE de bateaux de type japonais. Les données se situeraient en A3 et le stock serait sous-exploité (Figure VII.22).
Il est donc fortement recommandé que les données de CPUE 1981 à 1984 soient révisées et que tout soit mis en oeuvre pour assurer la qualité de ces données. La révision de la série 1981-84 devra être accomplie pour présentation au Groupe de travail ad hoc sur les céphalopodes en décembre 1985 à Ténérife.
Hypothèse 4 Baisse du recrutement et du potentiel du stock. Finalement il est aussi possible que le recrutement des octopus ait diminué fortement dans la région du cap Blanc. Selon les informations disponibles il est difficile de l'affirmer mais il faut rappeler qu'une telle baisse du recrutement fut observée de 1967 à 1975 (Figure VII.23). Ceci n'a pas pu être contrôlé pour la période récente faute de disposer des données de capture detaillées par catégories de taille mais l'hypothèse doit être considérée. Dans ce cas, le modèle historique (1966–80, Figure VII.22) n'est plus valable et la Mauritanie disposerait d'un stock de poulpes réduit, suivant le modèle 1982–84 (Figure VII.22) avec un potentiel de 20 000 t environ pleinement exploité. Le Groupe de travail ad hoc des céphalopodes a suivi les changements du recrutement du stock de Dakhla et il est souhaitable de le faire également au cap Blanc. Ceci pourrait être fait si on met en place un programme d'échantillonnage biologique des captures à Nouadhibou. Cette mise en place est donc recommandée. Un premier pas serait de collecter les statistiques de débarquement par catégories de tailles commerciales.
Toutes ces hyptohèses doivent être testées avec de meilleures données dans un futur aussi proche que possible.
- Seiches et calmars
Les informations disponibles n'ont pas permi de mettre à jour les évaluations déjá effectuées par Bravo de Laguna (1983) et données sur les Tableau VII.12 et VII.13 et les figures VII.16 et VII.17 du paragraphe 2.9.1.
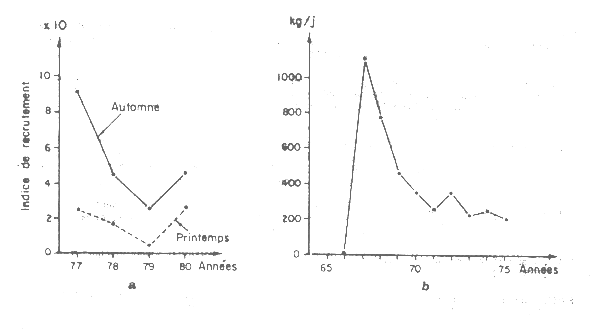
Figure VII.23 Changements de l'indice de recrutement du stock de Dakhla (a) et du cap Blanc (b) d'après FAO (1982, Fig 16 et 17)
Bas, C. et A. Guerra,1976 Catch and effort statistics and length frequency distribution regarding the Spanish cephalopod fishery in the CECAF region. FAO Fish.Rep., (183):111–3
Boukatine, P., 1985 Les ressources biologiques du talus continental mauritanien. In Rapport du Groupe de travail CNROP/FAO/ORSTOM sur les ressources halieutiques de la ZEE mauritanienne, Nouadhibou, 16–27 septembre 1985 (Annexe E)
Bravo de Laguna, J., Los recursos pesqueros del area de afloramiento del NW africano. Paper 1983 presented too the International Symposium on the most important upwelling areas of West Africa (Cape Blanc and Beguela, Barcelona, 21–25 November 1983: 61 p. Also Inv.Pesq. (in press))
Caveriviere, A.,1985 F. Domain et D. Thiam, Quelques estimations de mortalités pour les principales espèces de poissons démersaux du plateau continental mauritanien. In Rapport du Group de travail CNROP/FAO/ORSTOM sur les ressources halieutiques de la ZEE mauritanienne, Nouadhibou, 16–27 septembre 1985 (Annexe K)
Domain, F., Estimation par chalutage des ressources démersales du plateau continental mauritanien. 1985 In Rapport du Groupe de Travail CNROP/FAO/ORSTOM sur les ressources halieutiques de la ZEE mauritanienne, Nouadhibou, 16–27 septembre 1985
FAO, Rapport du Groupe de travail ad hoc sur l'évaluation des stocks de céphalopodes. Rome, FAO, 1979 135 p., COPACE/PACE Séries/78/11 (Fr)
FAO, Rapport du Groupe de travail spécial sur les stocks de céphalopodes de la région nord du 1982 COPACE. Rome, FAO, 180p., COPACE/PACE Séries/82/84(Fr)
FAO, 1983 Comité des pêches pour l'Atlantique centre-est, Rapport du Groupe de travail sur l'évaluation chinchards et les maqueraux de la zone nord du COPACE. Nouadhibou, Mauritanine, 30 janier–4 février 1983. COPACE/PACE Séries 83/27
FAO,1984 Comité des pêches pour l'Atlantique centre-est, Rapport du Groupe de travail ad hoc sur l'évaluation des ressources, Septiéme session, Santa Cruz de Ténérife, Espagne, 10–15 mai 1984. Premier groupe de travail spécial sur les pêcheries de merlus et de crevettes profondes dans la zone nord du COPACE (CECAF/RE/VII/84/3).
Fox, W.W.,1970 An exponential surplus-yield model for optimizing exploited fish populations. Trans.Am. Fish.Soc., 99(1):80–8
Gregoval, D., Analyse bioéconomique des principales pêcheries démersales dans la région nord 1982 COPACE. Dakar, projet COPACE, 118p., COPACE/TECH/82/85
Hatanaka, H., Studies on the fisheries biology on common octopus off the northwest coast of Africa. 1979 Bull.Far Seas Fish.Res.Lab., 17:13–124
Hennes, F., A multispecies assessment of the demersal fish stocks off southern Morocco with 1984 emphasis on the sparid resources. In Assessments of the stocks of Sparidae and Trachurus trachurus off Morocco. CECAF/TECH/84/62:1–146
Ikeda, I.,1976 Japanese research report. FAO Fish.Rep., (183):57–71
Lim Joo Yeul,1982 Review on cephalopod catches by Korean trawlers in the northern zone of CECAF, 1977–80. Annexe 15 au Rapport de la sixième session du Groupe de travail du COPACE sur l'évaluation des ressources, Dakar, Sénégal, février 1982
Maigret, J.,1976 Contribution à l'étude des langoustes de la côte occidentale d'Afrique (Crustace, Décapodes, Palinuridae). 2. Données historiques sur l'effort et la prise par unité d'effort dans la pêcherie langoustière sur les côtes du Sahara. Bull.IFAN, 38, sér.A (3):643–59
Maigret, J.,1978 Contribution à l'étude des langoustes de la côte occidentale d'Afrique (Crustace, Décapodes, Palinuridae). 6. Etude de la pêcherie langoustiére sur les côtes du Sahara. Bull.IF AN, 40, sér.A (3):553–86
Maigret, J.,1979 Etat du stock de langoustes roses (Palinurus mauritanicus, Gruvel, 1911) au large des côtes de Mauritanie. Invest.Pesq. , 23(1):83–94
Maigret, J., 1980Contribution à l'étude des langoustes de la côteoccidentale d'Afrique. Etat des stocks sur les côtes du Sahara en 1979. Bull.Centr.Nat.Rech.Océanogr.et Pêches, Nouadhibou, 9(1):84–101
Maigret, J.,1985Les stocks de Palinuridae de la côte nord ouest africane. Invest.Pesq., (sous presse)
Pauly, D.,1980 On the interrelationships between natural mortality, growth parameters and mean environmental temperature in 175 fish stocks. J.Cons.CIEM, 39(3):195–212
Pauly, D.,1982 Une sélection de méthodes simples pour l'estimation des stocks de poissons tropicaux. FAO Fish.Circ., 729
Pauly, D., 1984 Fish population dynamics in tropical waters; a manual for use with programmable calculators. ICLARM studies and reviews, 8, 325 p. International Center for Living Aquatic Resources Management, Manila, Philippines
Pella, J.J.1969 et P.K. Tomlinson, A generalized stock production model. Bull.IATTC, 13(3):421–96
Schaefer, M.B.,1954 Some aspects of the dynamics of populations important to the management of the commercial marine fisheries. Inter.Am.Trop.Tuna Comm., Bull.1(2):27–56
Shimura, T., Statistical survey of Las Palmas based international trawlers for 1982. CECAF/TECH/ 1983 83/49:46 p.
Sy Moussa and P.P. Tchernichkov,1985 Influences des conditions thermiques sur le rendement de la pêche des poulpes (Octopus vulgaris) dans la région du cap Blanc
Anonyme, Etat des stocks de poissons démersaux dans la zone économique mauritanienne. Bull. 1985 Centr.Nat.Rech.Océanogr.et Pêches, Nouadhibou, 13(1):138–68
| Longueur cm | 10.04.84 | 16.04.84 | 17.04.84 | 20.05.84 | 31.05.84 | 17.07.84 | 19.07.84 | 22.07.84 | 28.07.84 | |
|---|---|---|---|---|---|---|---|---|---|---|
| 16 | 4 | 4 | ||||||||
| 17 | 3 | 3 | ||||||||
| 18 | 2 | 9 | 2 | 13 | ||||||
| 19 | 1 | 10 | 2 | 1 | 14 | |||||
| 20 | 1 | 1 | 9 | 2 | 1 | 14 | ||||
| 21 | 3 | 1 | 8 | 3 | 1 | 16 | ||||
| 22 | 3 | 6 | 12 | 7 | 1 | 4 | 38 | |||
| 23 | 3 | 2 | 1 | 2 | 15 | 1 | 9 | 33 | ||
| 24 | 2 | 2 | 7 | 4 | 21 | 7 | 1 | 12 | 56 | |
| 25 | 4 | 1 | 8 | 2 | 8 | 13 | 6 | 1 | 14 | 57 |
| 26 | 6 | 2 | 1 | 7 | 18 | 2 | 2 | 9 | 47 | |
| 27 | 4 | 2 | 4 | 4 | 14 | 8 | 9 | 45 | ||
| 28 | 3 | 1 | 4 | 3 | 9 | 7 | 10 | 37 | ||
| 29 | 5 | 1 | 4 | 2 | 7 | 3 | 1 | 9 | 32 | |
| 30 | 2 | 3 | 4 | 2 | 4 | 1 | 8 | 24 | ||
| 31 | 1 | 5 | 2 | 2 | 2 | 7 | 19 | |||
| 32 | 6 | 6 | 1 | 1 | 14 | |||||
| 33 | 1 | 5 | 6 | |||||||
| 34 | 2 | 1 | 3 | |||||||
| 35 | 3 | 1 | 1 | 4 | ||||||
| 36 | 1 | 3 | 1 | 5 | ||||||
| 37 | 1 | 4 | 5 | |||||||
| 38 | 1 | 1 | ||||||||
| 39 | 3 | 3 | ||||||||
| Nombre d'individus | 48 | 40 | 15 | 26 | 44 | 154 | 53 | 6 | 98 | 487 |
| Longueur moyenne (cm) | 26,75 | 33,00 | 24,93 | 26,91 | 25,45 | 23,79 | 23,45 | 25,33 | 26,23 |
| Longueur cm | 10.04.84 | 17.04.84 | 19.05.84 | 20.05.84 | 31.05.84 | 17.05.84 | 22.07.84 | 28.07.84 | |
|---|---|---|---|---|---|---|---|---|---|
| 10 | 1 | 1 | |||||||
| 11 | 1 | 1 | |||||||
| 12 | 4 | 4 | |||||||
| 13 | 0 | ||||||||
| 14 | 1 | 0 | |||||||
| 15 | 1 | 1 | |||||||
| 16 | 11 | 11 | |||||||
| 17 | 2 | 3 | 1 | 6 | |||||
| 18 | 2 | 1 | 9 | 5 | 17 | ||||
| 19 | 4 | 3 | 8 | 6 | 7 | 3 | 31 | ||
| 20 | 5 | 19 | 7 | 6 | 6 | 40 | |||
| 21 | 3 | 3 | 24 | 7 | 26 | 2 | 65 | ||
| 22 | 9 | 2 | 11 | 11 | 20 | 6 | 59 | ||
| 23 | 3 | 3 | 1 | 7 | 4 | 19 | 5 | 42 | |
| 24 | 5 | 12 | 6 | 3 | 4 | 4 | 9 | 2 | 45 |
| 25 | 3 | 19 | 7 | 10 | 1 | 7 | 6 | 53 | |
| 26 | 15 | 2 | 3 | 4 | 1 | 7 | 3 | 35 | |
| 27 | 1 | 8 | 4 | 1 | 7 | 2 | 1 | 24 | |
| 28 | 1 | 2 | 1 | 8 | 1 | 3 | 16 | ||
| 29 | 1 | 1 | 2 | 2 | 1 | 7 | |||
| 30 | 1 | 2 | 2 | 3 | 8 | ||||
| 31 | 2 | 2 | 1 | 5 | |||||
| 32 | 2 | 2 | 4 | ||||||
| 33 | 1 | 1 | |||||||
| 34 | 0 | ||||||||
| 35 | 1 | 1 | 2 | ||||||
| 36 | 0 | ||||||||
| 37 | 1 | 3 | 4 | ||||||
| 38 | 0 | ||||||||
| 39 | 0 | ||||||||
| 40 | 0 | ||||||||
| 41 | 1 | 1 | |||||||
| 42 | 1 | 1 | |||||||
| Nombre d'individus | 37 | 60 | 54 | 20 | 117 | 53 | 112 | 32 | 485 |
| Longueur moyenne (cm) | 23,11 | 25,17 | 21,78 | 26,05 | 23,95 | 21,09 | 22,39 | 23,25 |
| Longueur (cm) | 1982 | 1983 | 1984 | 1985 | Total |
|---|---|---|---|---|---|
| 10 | 2 | 1 | 1 | 4 | |
| 1 | 0 | 8 | 9 | 17 | |
| 2 | 12 | 15 | 54 | 81 | |
| 3 | 58 | 29 | 14 | 4 | 235 |
| 4 | 219 | 170 | 124 | 12 | 525 |
| 5 | 369 | 366 | 157 | 39 | 931 |
| 6 | 480 | 633 | 240 | 106 | 1 459 |
| 7 | 289 | 689 | 300 | 196 | 1 474 |
| 8 | 135 | 640 | 303 | 457 | 1 535 |
| 9 | 32 | 740 | 321 | 411 | 1 504 |
| 20 | 23 | 1 000 | 564 | 314 | 1 901 |
| 1 | 50 | 1 019 | 531 | 331 | 1 931 |
| 2 | 53 | 763 | 761 | 270 | 1 847 |
| 3 | 41 | 612 | 1 197 | 353 | 2 203 |
| 4 | 53 | 730 | 1 504 | 413 | 2 700 |
| 5 | 58 | 596 | 1 160 | 405 | 2 219 |
| 6 | 41 | 535 | 754 | 247 | 1 577 |
| 7 | 48 | 510 | 661 | 132 | 1 351 |
| 8 | 98 | 683 | 672 | 131 | 1 584 |
| 9 | 198 | 761 | 242 | 120 | 1 321 |
| 30 | 270 | 927 | 151 | 164 | 1 512 |
| 1 | 380 | 719 | 82 | 90 | 1 271 |
| 2 | 416 | 620 | 91 | 59 | 1 186 |
| 3 | 170 | 473 | 89 | 47 | 779 |
| 4 | 576 | 328 | 88 | 31 | 1 023 |
| 5 | 504 | 149 | 69 | 19 | 741 |
| 6 | 462 | 47 | 32 | 14 | 555 |
| 7 | 282 | 26 | 11 | 11 | 330 |
| 8 | 196 | 6 | 5 | 9 | 216 |
| 9 | 138 | 3 | 1 | 7 | 149 |
| 40 | 74 | 0 | 0 | 5 | 79 |
| 1 | 30 | 1 | 2 | 33 | |
| 2 | 27 | 1 | 1 | 29 | |
| 3 | 0 | 0 | |||
| 4 | 10 | 2 | 12 | ||
| 5 | 3 | 3 | |||
| 6 | 2 | 2 | |||
| 7 | 0 | 0 | |||
| 8 | 1 | 1 | |||
| 9 | 3 | 3 | |||
| 50 | |||||
| 34 23 |
| Longueur (cm) | N |
|---|---|
| 12 | 4 |
| 3 | 4 |
| 4 | 5 |
| 5 | 2 |
| 6 | 2 |
| 7 | 1 |
| 8 | 1 |
| 9 | 1 |
| 20 | 9 |
| 1 | 49 |
| 2 | 129 |
| 3 | 211 |
| 4 | 334 |
| 5 | 348 |
| 6 | 347 |
| 7 | 536 |
| 8 | 669 |
| 9 | 619 |
| 30 | 751 |
| 1 | 556 |
| 2 | 610 |
| 2 | 623 |
| 3 | 583 |
| 4 | 466 |
| 5 | 315 |
| 7 | 282 |
| 8 | 470 |
| 9 | 448 |
| 40 | 146 |
| 1 | 112 |
| 2 | 80 |
| 3 | 69 |
| 4 | 46 |
| 5 | 27 |
| 6 | 10 |
| 7 | 3 |
| 8 | 4 |
| 9 | 1 |
| 50 | 1 |
| Total | 8 874 |
| Longueur (cm) | N |
|---|---|
| 10 | 2 |
| 1 | 58 |
| 2 | 48 |
| 3 | 10 |
| 4 | 3 |
| 5 | 1 |
| 6 | 1 |
| 7 | 1 |
| 8 | 0 |
| 9 | 29 |
| 20 | 186 |
| 1 | 197 |
| 2 | 486 |
| 3 | 596 |
| 4 | 751 |
| 5 | 1 043 |
| 6 | 1 128 |
| 7 | 1 154 |
| 8 | 1 061 |
| 9 | 762 |
| 30 | 786 |
| 1 | 726 |
| 2 | 629 |
| 3 | 404 |
| 4 | 243 |
| 5 | 118 |
| 6 | 74 |
| 7 | 30 |
| 8 | 17 |
| 9 | 10 |
| 40 | 0 |
| 1 | 0 |
| 2 | 0 |
| 3 | 1 |
| 4 | |
| Total | 10 655 |
| Longueur (cm) | N×10-3 |
|---|---|
| 18 | 879 |
| 19 | 3 739 |
| 20 | 3 556 |
| 21 | 2 562 |
| 22 | 2 150 |
| 23 | 1 702 |
| 24 | 2 511 |
| 25 | 2 579 |
| 26 | 2 086 |
| 27 | 1 877 |
| 28 | 2 620 |
| 29 | 2 952 |
| 30 | 2 889 |
| 31 | 1 681 |
| 32 | 1 573 |
| 33 | 1 202 |
| 34 | 976 |
| 35 | 316 |
| 36 | 143 |
| 37 | 37 |
| 38 | 54 |
| 39 | 12 |
| Total | 38 096 |
| Espéce | Merluccius senegalensis | |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Prof.m | 20 | 50 | 75 | 100 | 200 | 400 | 600 | |||||||
| L.cm | N | % | N | % | N | % | N | % | N | % | N | % | N | % |
| 8 | 5 | 0.2 | ||||||||||||
| 10 | 136 | 6.0 | ||||||||||||
| 12 | 3 | 0.9 | 621 | 27.4 | 6 | 0.2 | ||||||||
| 14 | 11 | 3.3 | 612 | 27.0 | 59 | 2.3 | ||||||||
| 16 | 80 | 24.3 | 344 | 15.2 | 270 | 10.7 | ||||||||
| 18 | 97 | 29.5 | 177 | 7.8 | 253 | 10.0 | ||||||||
| 20 | 100 | 30.4 | 127 | 5.6 | 476 | 18.9 | 3 | 0.4 | ||||||
| 22 | 26 | 7.9 | 95 | 4.2 | 587 | 23.2 | 11 | 1.4 | 6 | 0.4 | ||||
| 24 | 9 | 2.7 | 86 | 3.8 | 440 | 17.4 | 95 | 11.9 | 28 | 1.8 | ||||
| 26 | - | - | 32 | 1.4 | 305 | 12.1 | 327 | 41.0 | 230 | 14.6 | ||||
| 28 | 3 | 0.9 | 23 | 1.0 | 94 | 3.7 | 300 | 37.6 | 372 | 23.6 | ||||
| 30 | 9 | 0.4 | 29 | 1.1 | 58 | 7.3 | 241 | 15.3 | 1 | 0.8 | ||||
| 32 | 6 | 0.2 | 3 | 0.4 | 28 | 1.8 | - | - | ||||||
| 34 | 68 | 4.3 | 1 | 0.8 | ||||||||||
| 36 | 91 | 5.8 | 4 | 3.3 | ||||||||||
| 38 | 139 | 8.8 | 13 | 10.7 | ||||||||||
| 40 | 162 | 10.3 | 21 | 17.4 | 2 | 1.7 | ||||||||
| 42 | 91 | 5.8 | 24 | 19.8 | 1 | 0.9 | ||||||||
| 44 | 57 | 3.6 | 32 | 26.4 | 6 | 5.2 | ||||||||
| 46 | 26 | 1.0 | 15 | 12.4 | 21 | 18.3 | ||||||||
| 48 | 8 | 0.5 | 4 | 3.3 | 22 | 19.1 | ||||||||
| 50 | 3 | 0.2 | 2 | 1.6 | 32 | 27.8 | ||||||||
| 52 | 3 | 0.2 | 3 | 2.5 | 15 | 13.0 | ||||||||
| 54 | - | - | 6 | 5.2 | ||||||||||
| 56 | - | - | 5 | 4.3 | ||||||||||
| 58 | - | - | 4 | 3.5 | ||||||||||
| 60 | 3 | 0.2 | 1 | 0.9 | ||||||||||
| 62 | 6 | 0.4 | ||||||||||||
| 64 | 3 | 0.2 | ||||||||||||
| 66 | - | - | 1 | 0.8 | ||||||||||
| 68 | - | - | ||||||||||||
| 70 | 6 | 0.4 | ||||||||||||
| 72 | 3 | 0.2 | ||||||||||||
| Total | 329 | 2267 | 2525 | 797 | 1574 | 121 | 115 | |||||||
| 99.9 | 100.0 | 99.8 | 100.0 | 100.0 | 99.8 | 99.9 | ||||||||
| capture par heure | |||||||
|---|---|---|---|---|---|---|---|
| (kg) | 17.4 | 76.6 | 225.5 | 137.8 | 452.7 | 79.8 | 104.6 |
| Profond.m | 20–35 | 35–62.5 | 62.5–87.5 | 87.5–150 | 150–300 | 300–500 | 500–700 | 20–700 | |
|---|---|---|---|---|---|---|---|---|---|
| Surface/ % m2 | 835.03/36.20 | 471.59/20.45 | 178.31/7.73 | 201.69/8.74 | 180.26/7.82 | 176.36/7.65 | 263.08/11.41 | 2306.32/100.00 | |
| L. cm | N | N | N | N | N | N | N | N | % |
| 8 | 102 | 102 | 0.1 | ||||||
| 10 | 2781 | 2781 | 2.8 | ||||||
| 12 | 109 | 12699 | 46 | 12854 | 12.9 | ||||
| 14 | 398 | 12515 | 456 | 13369 | 13.5 | ||||
| 16 | 2896 | 7035 | 2087 | 12018 | 12.1 | ||||
| 18 | 3511 | 3620 | 1956 | 9087 | 9.2 | ||||
| 20 | 3620 | 2597 | 3679 | 26 | 9922 | 10.0 | |||
| 22 | 941 | 1943 | 4538 | 96 | 47 | 7565 | 7.6 | ||
| 24 | 326 | 1759 | 3401 | 830 | 219 | 6535 | 6.6 | ||
| 26 | - | 654 | 2358 | 2858 | 1799 | 7669 | 7.7 | ||
| 28 | 109 | 470 | 727 | 2622 | 2909 | 6837 | 6.9 | ||
| 30 | 184 | 224 | 507 | 1885 | 8 | 2808 | 2.8 | ||
| 32 | 46 | 26 | 219 | - | 291 | - | 0.3 | ||
| 34 | 532 | 8 | 540 | 0.5 | |||||
| 36 | 712 | 31 | 743 | 0.7 | |||||
| 38 | 1087 | 99 | 1186 | 1.2 | |||||
| 40 | 1267 | 161 | 23 | 1451 | 1.5 | ||||
| 42 | 712 | 184 | 11 | 907 | 0.9 | ||||
| 44 | 446 | 245 | 68 | 759 | 0.7 | ||||
| 46 | 203 | 115 | 240 | 558 | 0.5 | ||||
| 48 | 63 | 31 | 251 | 345 | 0.3 | ||||
| 50 | 23 | 15 | 365 | 403 | 0.4 | ||||
| 52 | 23 | 23 | 171 | 217 | 0.2 | ||||
| 54 | - | - | 68 | 68 | 0.1 | ||||
| 56 | - | - | 57 | 57 | 0.1 | ||||
| 58 | - | - | 46 | 46 | 0.1 | ||||
| 60 | 23 | - | 11 | 34 | 0.03 | ||||
| 62 | 47 | - | 47 | 0.1 | |||||
| 64 | 23 | - | - | 23 | 0.02 | ||||
| 66 | - | 8 | 8 | 0.01 | |||||
| 68 | - | - | - | ||||||
| 70 | 47 | 47 | 0.1 | ||||||
| 72 | 23 | 23 | 0.02 | ||||||
| Total | 99300 | 99.98 | |||||||
| Captures totales Nouadhibou + Sahara Occ. | Exportation Nouadhibou | Pêche arti- Nouadhibou | Pêche Lang Nouakchott | Pêche arti- Nouakchott | Chalutiers Nouakchott | |
|---|---|---|---|---|---|---|
| 1968 | 186 | |||||
| 1969 | 301 | |||||
| 1970 | 289 | |||||
| 1971 | 289 | |||||
| 1972 | 249 | 22 | ||||
| 1973 | 210 | (16) | ||||
| 1974 | 180 | (6) | ||||
| 1975 | 150 | |||||
| 1976 | 13,4 | |||||
| 1977 | 12,6 | |||||
| 1978 | 16 | |||||
| 1979 | 34 | |||||
| 1980 | 51 | |||||
| 1981 | 129 | |||||
| 1982 | 197 | 11,2 | 4,2 | |||
| 1983 | 109 | 20,3 | 4,6 | |||
| 1984 | 242,6 | 35,9 | 29.8 | |||
| Stock nord | Stock sud | |||||
| Anneé | Captures (1) | Nombre de bateaux | Nombre de marées | Rendement par marée | PUE | Effort total | Effort corrigé |
|---|---|---|---|---|---|---|---|
| 1968 | 272 | 14 | 19 | 14,31 | 92 | 2 953 | 5 465 |
| 1969 | 273 | 14 | 21 | 13,00 | 82 | 3 338 | 4 448 |
| 1970 | 200 | 14 | 16 | 12,50 | 114 | 1 748 | 3 254 |
| 1971 | 247 | 10 | 16 | 14,96 | 150 | 1 641 | 2 420 |
| 1972 | 215 | 9 | 18 | 11,94 | 147 | 1 457 | 2 046 |
| 1973 | 324 | 14 | 27 | 14,24 | 149 | 2 177 | 1 756 |
| 1974 | 391 | 10 | 24 | 16,14 | 175 | 2 231 | 1 876 |
| 1975 | 336 | 10 | 20 | 16,80 | 181 | 1 850 | 1 929 |
| 1976 | 348 | 10 | 17 | 20,47 | 259 | 1 343 | 1 900 |
| 1977 | 479 | 9 | 21 | 22,80 | 297 | 1 609 | 1 758 |
| 1978 | 482 | 10 | 22 | 21,90 | 313 | 1 541 | 1 585 |
| 1979 | 683 | 10 | 26 | 26,26 | 375 | 1 821 | 1 578 |
| 1980 | 691 | 10 | 25 | 28,00 | 419 | 1 648 | 1 654 |
| 1981 | 692 | 10 | 25 | 28,00 | 389 | 1 778 | 1 697 |
| 1982 | 556 | 8 | 16 | 34,72 | 505 | 1 100 | 1 507 |
| 1982 | 840 | 10 | 24 | 35,01 | 530 | 1 585 | 1 610 |
| Année | Capture ×103 | Effort ×103(h. pêche) | c.p.u.e (tonnes/heure) |
|---|---|---|---|
| 1966 | 11.6 | 17 | 0.682 |
| 1967 | 28.7 | 46 | 0.624 |
| 1968 | 45.7 | 71 | 0.644 |
| 1969 | 34.2 | 60 | 0.570 |
| 1970 | 34.6 | 89 | 0.389 |
| 1971 | 38.2 | 120 | 0.318 |
| 1972 | 36.6 | 114 | 0.321 |
| 1973 | 29.2 | 100 | 0.292 |
| 1974 | 36.6 | 159 | 0.230 |
| 1975 | 39.9 | 214 | 0.186 |
| 1976 | 52.9 | 268 | 0.197 |
| 1977 | 36.6 | 125 | 0.293 |
| 1978 | 23.3 | 82 | 0.284 |
| 1979 | 13.4 | 78 | 0.172 |
| 1980 | 12.5 | 51 | 0.245 |
| 1981 | 19.2 | 41.3 | 0.46 |
| 1982 | 15.1 | 21.3 | 0.71 |
| 1983 | 20.7 | 36.8 | 0.56 |
| 1984 | 25.5 | 29.0 | 0.88 |
Source: 1966–1980: FAO 1982
1981–1984: CNROP
| Année | Capture (x103 t) | Effort (x103 h) | CPUE (t/h) |
|---|---|---|---|
| 1969 | 30.0 | 526 | 0.057 |
| 1970 | 17.9 | 331 | 0.054 |
| 1971 | 24.3 | 565 | 0.043 |
| 1972 | 27.0 | 1688 | 0.015 |
| 1973 | 16.1 | 894 | 0.018 |
| 1974 | 35.8 | 1155 | 0.030 |
| 1975 | 18.8 | 1466 | 0.013 |
| 1976 | 17.0 | 1700 | 0.010 |
| 1977 | 24.0 | 2182 | 0.011 |
| 1978 | 31.8 | 1767 | 0.018 |
| 1979 | 24.6 | 1640 | 0.015 |
| 1980 | 32.1 | 2293 | 0.013 |
| 1981 | 53.9 | 2695 | 0.020 |
| 1982 | 45.4 | 2838 | 0.016 |
Source: Bravo de Laguna (1983)
| Année | Capture (× 103 t) | Effort (× 103 h) | CPUE (t/h) |
|---|---|---|---|
| 1969 | 13.7 | 311.4 | 0.044 |
| 1970 | 12.6 | 233.3 | 0.054 |
| 1971 | 18.4 | 460.0 | 0.040 |
| 1972 | 21.2 | 1177.8 | 0.018 |
| 1973 | 19.0 | 730.8 | 0.026 |
| 1974 | 29.1 | 1818.8 | 0.016 |
| 1975 | 17.6 | 1257.1 | 0.014 |
| 1976 | 9.5 | 1583.3 | 0.006 |
| 1977 | 14.6 | 2920.0 | 0.005 |
| 1978 | 20.8 | 1890.9 | 0.011 |
| 1979 | 12.0 | 1500.0 | 0.008 |
| 1980 | 9.9 | 1980.0 | 0.005 |
| 1981 | 16.5 | 3300.0 | 0.005 |
| 1982 | 8.0 | 2666.7 | 0.003 |
Source: Bravo de Laguna (1983)
| Année | Indice1 d'upwelling | Δ cpue exponentiel | Δcpue linéaire |
|---|---|---|---|
| 1966 | - 0.21 | + 0.21 | + 0.19 |
| 1967 | - 0.21 | + 0.22 | + 0.18 |
| 1968 | + 0.16 | + 0.29 | + 0.24 |
| 1969 | - 0.43 | + 0.2 | + 0.15 |
| 1970 | - 0.53 | + 0.07 | + 0.03 |
| 1971 | - 0.22 | + 0.05 | + 0.02 |
| 1972 | + 0.18 | + 0.04 | + 0.00 |
| 1973 | - 0.02 | - 0.01 | - 0.05 |
| 1974 | - 0.22 | + 0.01 | 0 |
| 1975 | - 0.31 | + 0.03 | + 0.06 |
| 1976 | + 0.08 | + 0.08 | + 0.17 |
| 1977 | + 0.6 | + 0.03 | - 0.01 |
| 1978 | 0.0 | - 0.05 | - 0.07 |
| 1979 | 0.0 | - 0.17 | - 0.21 |
| 1980 | + 0.41 | - 0.15 | - 0.19 |
| 1981 | + 0.40 | - 0.09 | - 0.10 |
| 1982 | - 0.19 | - 0.14 | - 0.19 |
1 Obtenu graphiquement de sy Moussa st Tchernikov (1985), Bull NROP 1 (13)
![]()
![]()
![]()